Application of cell-free DNA sequencing in characterization of bloodborne microbes and the study of microbe-disease interactions - PeerJ
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Application of cell-free DNA sequencing in characterization of
bloodborne microbes and the study of microbe-disease
interactions
Kuo-Ping Chiu Corresp., 1, 2 , Alice L. Yu 3, 4
1
Genomics Research Center, Academia Sinica, Taipei, Taiwan
2
Departent of Life Sciences, National Taiwan University, Taipei, Taiwan
3
Department of Pediatrics, University of California, San Diego, San Diego, United States
4
Institute of Stem Cell and Translational Cancer Research, Chang Gung University, Taipei, Taiwan
Corresponding Author: Kuo-Ping Chiu
Email address: chiukp@gate.sinica.edu.tw
Background. It is an important issue whether and how microorganisms can live harmoniously
withnormal cells in the circulatory system. Answers to these issues will have enormous impact on
medical microbiology. To address these issues, it is essential to identify and characterize the blood-borne
microbes in an efficient and comprehensive manner.
Methodology. Traditional approaches using PCR or microarray are not suitable for the purpose due to
the complexity and composition of large amount of unknown microbial species in the circulatory system.
Recent reports indicated that cell-free DNA (cfDNA) sequencing using advanced sequencing technologies,
including next-generation sequencing (NGS) and single-molecule sequencing (SMS) together with
associated bioinformatics approaches, possess a strong potential enabling us to address these issues at
the molecular level.
Results. Multiple studies using microbial cfDNA sequencing to identify microbes for septic patients have
shown strong agreement with cell culture. Similar approaches have also been applied to reveal
previously unidentified microorganisms or to demonstrate the feasibility of comprehensive assessment of
bloodborne microorganisms for healthy and/or diseased individuals. Single-molecule sequencing (SMS)
using either SMRT (single-molecule real-time) sequencing or Nanopore sequencing are providing new
momentum to reinforce this line of investigations.
Conclusions. Microbial cfDNA sequencing provides a novel opportunity allowing us to further
understand the involvement of blood-borne microbes in development of diseases. Similar approaches
should also be applicable to the study of metagenomics for sufficient and comprehensive analysis of
microbial species isolated from various environments. This article reviews this line of research and
discuss the methodological approaches that have been developed, or are likely to be developed in the
future, which may have strong potential to facilitate cfDNA- and cfRNA-based studies of cancer and
chronic diseases, in the hope that a better understanding of the hidden microbes in the circulatory
system would improve the accuracy of diagnosis, prevention, and treatment of problematic diseases.
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 20191 Application of cell-free DNA sequencing in
2 characterization of bloodborne microbes and the
3 study of microbe-disease interactions
4
5 Kuo-Ping Chiu1, 2, Alice L. Yu3, 4
6
7
8
9 1Genomics Research Center, Academia Sinica, Taipei, Taiwan
10 2Department of Life Sciences, College of Life Sciences, National Taiwan University, Taipei,
11 Taiwan
12 3Institute of Stem Cell and Translational Cancer Research, Chang Gung Memorial Hospital at
13 Linkou and Chang Gung University, Taoyuan, Taiwan
14 4Department of Pediatrics, University of California in San Diego, San Diego, California, USA
15
16
17
18
19
20 Corresponding Author:
21 Kuo-Ping Chiu
22 128 Academia Road, Sec. 2, Nankang District, Taipei, 115, Taiwan
23 Email address: chiukp@gate.sinica.edu.tw
24
25
26
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 201927 Abstract
28 Background. It is an important issue whether and how microorganisms can live harmoniously
29 with normal cells in the circulatory system. Answers to these issues will have enormous impact
30 on medical microbiology. To address these issues, it is essential to identify and characterize the
31 blood-borne microbes in an efficient and comprehensive manner.
32 Methodology. Traditional approaches using PCR or microarray are not suitable for the purpose
33 due to the complexity and composition of large amount of unknown microbial species in the
34 circulatory system. Recent reports indicated that cell-free DNA (cfDNA) sequencing using
35 advanced sequencing technologies, including next-generation sequencing (NGS) and single-
36 molecule sequencing (SMS) together with associated bioinformatics approaches, possess a
37 strong potential enabling us to address these issues at the molecular level.
38 Results. Multiple studies using microbial cfDNA sequencing to identify microbes for septic
39 patients have shown strong agreement with cell culture. Similar approaches have also been
40 applied to reveal previously unidentified microorganisms or to demonstrate the feasibility of
41 comprehensive assessment of bloodborne microorganisms for healthy and/or diseased
42 individuals. Single-molecule sequencing (SMS) using either SMRT (single-molecule real-time)
43 sequencing or Nanopore sequencing are providing new momentum to reinforce this line of
44 investigations.
45 Conclusions. Microbial cfDNA sequencing provides a novel opportunity allowing us to further
46 understand the involvement of blood-borne microbes in development of diseases. Similar
47 approaches should also be applicable to the study of metagenomics for sufficient and
48 comprehensive analysis of microbial species isolated from various environments. This article
49 reviews this line of research and discuss the methodological approaches that have been
50 developed, or are likely to be developed in the future, which may have strong potential to
51 facilitate cfDNA- and cfRNA-based studies of cancer and chronic diseases, in the hope that a
52 better understanding of the hidden microbes in the circulatory system would improve the
53 accuracy of diagnosis, prevention, and treatment of problematic diseases.
54
55
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 201956 Introduction
57 It is important to understand whether microorganisms can live harmoniously with normal cells in
58 blood circulation, which is equipped with immune surveillance evolved to combat
59 microorganisms. The answer will not only have significant impact on medicine, but also pave the
60 way towards further studies to address key questions, such as, how hidden microbes can be
61 tolerated by, or escape from, the immune surveillance? What are the roles played by the hidden
62 microbes in disease development? How microbiota interact with the immune system along the
63 development of a disease? To stratify the interactions between bloodborne microbes and
64 diseases, identification and characterization of the hidden microbes is essential.
65
66 Conventional procedure of cell culture followed by laboratory characterization with PCR or
67 microarray has been the gold standard for microbial species identification and characterization.
68 However, it has severe limitations not only in sensitivity and specificity for detecting known
69 species but also being incapable of detecting unknown species. Many microbes in clinical
70 samples are fastidious and either cannot be culturable with conventional approaches or cannot be
71 precisely assigned to specific species. These limitations result in significant false negative in
72 detection and ambiguity in species annotation (Hall and Lyman, 2006; Lamy et al., 2016; Pham
73 and Kim, 2012).
74
75 With the advances in sequencing technology, including next-generation sequencing (NGS) and
76 single molecule sequencing (SMS), and coupled bioinformatics approaches, the above-
77 mentioned issues can now be more effectively addressed at the molecular level using cfDNA
78 sequencing. Recent reports have demonstrated the feasibility of using NGS- and SMS-coupled
79 cell-free DNA sequencing for comprehensive microbiota profiling in bloodstream (De Vlaminck
80 et al., 2015; Grumaz et al., 2016; Hong et al., 2018; Huang et al., 2018; Kowarsky et al., 2017).
81 The same approach is expected to unravel the potential correlation of diseases (e.g., chronic
82 diseases and cancer) and microbes in blood circulation, especially the involvement of microbes
83 in disease onset and progression. Additionally, similar approaches should be able to enhance the
84 analyses of metagenomics, gastrointestinal microbiota, as well as microbial species in various
85 environments.
86
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 201987 There are reviews on applications of cfDNA sequencing in noninvasive prenatal testing (NIPT)
88 and oncology (Bianchi and Chiu, 2018; Finotti et al., 2018; Oellerich et al., 2017), except for
89 blood-borne microbial species identification and characterization. Here, we review the associated
90 technologies, experimental designs and potential applications, in hope to facilitate the progress of
91 scientific research in this direction.
92
93 Survey methodology
94 We surveyed literatures in PubMed and Google Scholar websites using the keywords cell-free
95 DNA, cfDNA and "cell-free DNA" AND "microbial" and included the articles related to the
96 focus of current review. We comprehensively and unbiasedly cover recent articles published in
97 Feb. 2019 and beyond. This review is aimed to provide a up-to-date research trend pertaining to
98 the identification of bloodborne microbial species associated with cancer using cell-free DNA.
99
100 Microbes and human diseases
101 Microbes, including bacteria, fungi and viruses, together with phages that infect prokaryotic
102 cells, form the largest population of biological entities on Earth. Due to constant and intimate
103 contact with humans, their role for the onset and development of diseases, including cancer, have
104 long been investigated by academic laboratories, clinics and hospitals worldwide.
105
106 Studies have associated numerous types of cancer with viral infections. As absolute pathogens,
107 viruses rely on the biological functions of the host cells for replication. This is achieved by
108 internalization of viral genetic material into the host cell followed by manipulation of host’s
109 biological machineries to favor the synthesis of viral nucleic acids and proteins for the assembly
110 of new viral particles. During the process, the host genome may be altered to induce
111 tumorigenesis, such as that shown in Kaposi’s sarcoma induced by HIV (human
112 immunodeficiency virus) or KSHV (Kaposi’s sarcoma herpesvirus) (Mehta et al., 2011; Pyakurel
113 et al., 2007), cervical cancer induced by HPV (human papillomavirus) (Burd, 2003),
114 hepatocarcinoma induced by HBV (hepatitis B virus) (Levrero and Zucman-Rossi, 2016), etc.
115
116 On the other hand, bacteria and fungi are free-living microorganisms. Little is known about their
117 roles in cancer or other diseases. However, some unexpected observations were reported. For
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019118 instance, low levels of pleomorphic bacteria were found in blood circulation of healthy
119 individuals (McLaughlin et al., 2002; Potgieter et al., 2015; Tedeschi et al., 1976). Bacteria were
120 also detected in tumor tissues (Cummins and Tangney, 2013). Since bacteria and fungi,
121 especially the former, may involve in a broad spectrum of host physiology, including
122 metabolism, inflammation, immunity and hematopoiesis, they may play a role in tumorigenesis
123 or cancer development (Roy and Trinchieri, 2017). However, this line of evidence has been
124 elusive and discrete. To stratify the association of microbial cells with diseases, more data are
125 required and the identification and characterization of bloodborne microbes is essential.
126
127 Limitations of cell culture on characterization of bloodborne pathogens
128 Cell culture using either liquid media or agar plates is a gold standard as well as the mainstream
129 clinical tool for the identification and characterization of infectious agents. Blood agar plates are
130 commonly used in the lab to isolate fastidious bacteria such as Streptococcus, Enterococcus and
131 Aerococcus. Bacterial colonies producing hemolysins capable of lysing red blood cell or
132 hemoglobin are distinguishable based on the color and the degree of transparency in the
133 background (Lorian, 1971).
134
135 However, besides the requirement of a long period of waiting (about a week or so) to obtain
136 results and tedious laboratory works, blood culture is hindered by limitation in sensitivity. A
137 literature survey reported that among patients with bloodstream infections, only 5 to 13% of
138 blood cultures turn out to be positive (Lamy et al., 2016). Furthermore, 0.6-6% of positive results
139 were in fact caused by contamination by etiologically irrelevant microbes which are frequently
140 introduced by indwelling vascular access devices (Hall and Lyman, 2006). A much higher ratio
141 of false positive was even estimated independently by another group (Lamy et al., 2016). Even
142 for microbial inhabitants in oral cavity, which acts as a buffer zone between the circulatory
143 system and outside environment, only about 50% of oral microorganisms are culturable (Pham
144 and Kim, 2012; Wade, 2002).
145
146 The above-mentioned problems are more or less correlated with the difficulty in culturing
147 microbes growing in natural habitats. It was shown that less than 1% of soil bacteria were
148 culturable in the lab (Pham and Kim, 2012). Since many environmental bacteria may enter
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019149 human body through wounds or via respiratory or digestive tracks, numerous environmental
150 microbes have the potential to become opportunistic pathogens in hospital (Buonaurio et al.,
151 2002; Huang et al., 2018).
152
153 Cell-free DNA and its potential in diagnostics
154 Liquid biopsies represent one of the most valuable and readily accessible material for diagnosis
155 and prognosis. For example, plasma cell-free DNA (cfDNA) samples are much more readily
156 available and accessible compared to diseased tissues (e.g., tumor or diseased tissues). A cfDNA
157 sample acts like a genetic reservoir which carries genetic information from all cells within the
158 body, as DNA fragments are generated by apoptosis or necrosis of virtually all cells within the
159 body, including healthy and diseased cells and microbes.
160
161 Although cfDNA samples can be analyzed with PCR or microarray using specific primers or
162 probes to target 16S rRNA or particular genetic sequences in microorganisms, these approaches
163 are not suitable for detecting novel microorganisms. Moreover, primer binding sites or probe-
164 recognition sites may be degraded or fragmented in cfDNA samples.
165
166 NGS-based cfDNA analysis as a robust molecular diagnostic tool
167 Compared to Sanger Sequencing, NGS conveys a number of advantages including higher speed,
168 yield and completeness, but with lower cost, while Sanger Sequencing retains highest accuracy.
169 These advantages have revolutionized biological investigations, making a number of previously
170 untouchable research territories workable (Levy and Myers, 2016), including accelerated human
171 genome sequencing, antibody-coding gene variable region sequencing (Boyd and Crowe, 2016),
172 single-cell sequencing (Chiang et al., 2018; Gawad et al., 2016), and recently developed
173 microbial species identification using cfDNA (see below).
174
175 By combining the prestigious features of cfDNA with the technological strength of massively
176 parallel sequencing, NGS-based cfDNA sequencing is gaining tremendous momentum in
177 diagnostics. Cell-free DNA sequencing is becoming a robust approach not only for the analysis
178 of disease-associated genetic mutations in cancer and NIPT (noninvasive prenatal testing)
179 (Butler et al., 2015; Mehrotra et al., 2018), but also for the identification of fastidious microbes
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019180 in the circulatory system (Blauwkamp et al., 2019; De Vlaminck et al., 2015; Grumaz et al.,
181 2016; Huang et al., 2018).
182
183 Previously cfDNA sequencing focused mainly on the study of human subjects. During past few
184 years, the potential of NGS-based cfDNA sequencing in analysis of cancer susceptibility genes
185 have also been well evaluated (Fernandez-Cuesta et al., 2016; Volik et al., 2016). Relatively less
186 attention was paid to microbial sequences in cfDNA sample. For microbial DNA analysis, it is
187 practical to just focus on specific sequence markers such as 16S rDNA/rRNA previously or PCR
188 amplicons (Wade, 2002). Current cfDNA sequencing mainly relies on NGS, while SMS, which
189 is a robust tool for long-read sequencing and which does not necessarily rely on molecular
190 cloning or PCR, is severely hampered by high error rates and random deletions and insertions
191 (Weirather et al., 2017).
192
193 Applications of cell-free DNA in Microbiology
194 We now return to the focus on the application of cfDNA sequencing in microbial species
195 characterization. Here, we will survey some reports published during the past few years which
196 we think are important and relevant to our subject. Although there are many more related reports
197 which are more or less related to our subject, our review is constrained and serious readers are
198 recommended to refer to the literature online.
199
200 Application in monitoring infection and rejection in lung transplantation patients
201 It is difficult to use traditional methods to distinguish infection and rejection resulted from organ
202 transplantation, because both have similar clinical symptoms. In their attempt to apply cfDNA
203 sequencing in monitoring infection and rejection of lung transplantation, Vlaminck and
204 colleagues developed a method using single nucleotide polymorphisms (SNPs) to distinguish
205 donor and recipient cfDNA fragments (De Vlaminck et al., 2015) for the study of transplant
206 rejection and infection. Levels of donor-originated cfDNA were found directly correlated with
207 data from invasive test of rejection. This noninvasive approach avoids invasive tissue biopsies
208 commonly used for detecting rejection. On the other hand, levels of cytomegalovirus-derived
209 cfDNA were found correlated with clinical results of infection.
210
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019211 Application in identification of microbes in sepsis
212 In an attempt to fight septicemia, Grumaz and colleagues reported a cfDNA sequencing approach
213 to detect the predominant microbial species in bloodstream (Grumaz et al., 2016). Empirically,
214 cfDNA samples were isolated at different time points from three groups including septic patients,
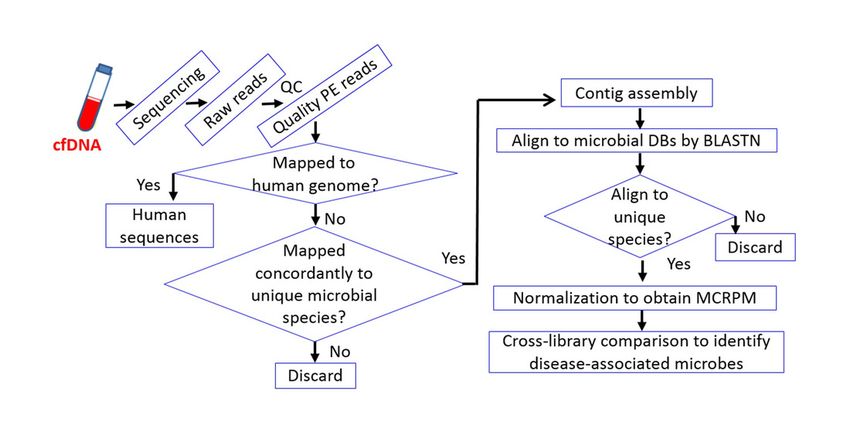
215 surgical patients and normal volunteer controls. Sequencing libraries were constructed and
216 sequenced with HiSeq 2500 to generate 25-30 million 100 bp single-end reads. They first
217 mapped quality reads to human genome assembly hg19. Unmappable reads were then mapped to
218 RefSeq database comprising bacterial, viral and selected fungal genomes. Libraries, including
219 those from septic patients, normal volunteer controls and surgical patients, were normalized and
220 compared.
221
222 They were able to obtain results in within a period of time shorter than traditional cell culture,
223 which normally requires about 5 days. The identified microbial species were strongly correlated
224 with that obtained by blood cultures. In general, septic patients have highest cfDNA
225 concentrations than surgical patients and volunteer controls. Both Gram-negative and Gram-
226 negative bacteria well matched that identified by corresponding blood cultures. Quantification of
227 microbial species is essential for both cross-species and cross-sample comparisons. In their
228 report, the authors employed SIQ values, calculated by complicate formulas, to estimate dynamic
229 microbial cfDNA levels. Some false negatives by cell culture were detected and rescued by
230 cfDNA sequencing.
231
232 This work represents a great achievement for sepsis. However, although applicable for septic
233 patients and potentially for other bacteremic cases, mapping with single-end reads of 50 - 100 bp
234 in length in conjunction with a minimum of 80% identity between read and reference genome
235 may not be stringent enough for the identification of low abundance microorganisms.
236 Additionally, calculation of SIQ seems to be complicate and not very user friendly.
237
238 Application in discovery of novel bacteria and viruses in bloodstream
239 By a deep sequencing of 1,351 cfDNA samples isolated from 188 patients, Kowarsky and
240 colleagues reported that among a total of 37 billion cfDNA fragments sequenced, 0.45% of reads
241 were unable to align to reference human genome assembly; and among those, only 1% aligned to
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019242 reference microbiome database composed of about 8,000 species of known bacteria, viruses,
243 fungi and pathogens (Kowarsky et al., 2017). By de novo assembly of the non-human sequences,
244 they generated 7,190 contigs. With a series of filtering, testing and merging, they identified
245 3,761 novel contigs, which were validated not to be artifacts or contaminants. Further analysis
246 indicated that these novel contigs were derived from a vast number of uncharacterized microbes,
247 suggesting that unexpected diversity of novel phages and viruses is present in human body. Thus,
248 microbiome in human body was substantially undersampled previously.
249
250 Application in identification of bloodborne microbes in healthy females and early-onset breast
251 cancer patients
252 Our recent work further demonstrated the advantages of cfDNA sequencing in microbial species
253 identification for both healthy individuals and early-onset breast cancer (EOBC) patients (Huang
254 et al., 2018). The work relies on a stringent pipeline for mapping and alignment of NGS paired
255 end (PE) reads and contigs, respectively, to identify microbial inhabitants in healthy individuals
256 and early-onset breast cancer (EOBC) patients. The relative levels of microbe-associated cfDNA
257 were compared under MCRPM (microbial cfDNA reads per million) basis after normalization
258 (Fig. 1). Healthy individuals were found to have similar bacterial profiles. Patient with a profile
259 similar to that of healthy individuals indicated a fair recovery, while patients with advanced
260 EOBC were found to inhabit with opportunistic pathogens. Microbial profiles can thus be
261 associated with health conditions. The approach paves the way toward a better understanding of
262 the role played by microbes in the development and prognosis of cancer and chronic diseases.
263
264 For microbial species identification, long read alignment is more accurate and thus a better
265 choice than short reads, because large number of microbial genomes have high degree of
266 sequence similarity. Microbial genomes are much smaller (in the range of a few Mb) than the
267 human genome (about 3 Gb for 1N cells). In order to obtain carbon and energy sources, certain
268 genomic regains are able to alter quickly through mutations or horizontal gene transfer, while
269 other regions remain largely unchanged, making species identification by alignment a great
270 challenge, especially when using cfDNA-derived paired-end reads. Thus, contig-based alignment
271 is supposed to be more accurate and reliable than single-end, or even paired-end reads. The work
272 suggested that cfDNA-derived microbial profile can function as a prognostic indicator or
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019273 reference. Since spreading of microbes is strongly influenced by environment, it is not possible
274 to directly correlate a particular disease, or its clinical status, with a particular microbial profile.
275
276 Validation of microbial cfDNA sequencing test showed a strong agreement with blood culture
277 Very recently, an analytical study was conducted by a group at Stanford to validate a microbial
278 cfDNA sequencing test (Blauwkamp et al., 2019). In the study, factors that may affect accuracy,
279 precision or that may introduce bias were examined, and results validated with in silico
280 simulation, to evaluate the reliability and robustness of microbial cfDNA sequencing for
281 assessing clinically relevant microbes or eukaryotic parasites (1,250 predefined). Results were
282 then compared to cell culture.
283
284 To ensure the reliability of the validation, the authors used 13 microorganisms as reference.
285 Various cfDNA concentrations were prepared and calculated, in microbe-specific cfDNA per
286 microliter of plasma (MMP), so that factors that may affect reliability (e.g., limit of detection,
287 limit of quantitation, accuracy, precision and linearity, etc.) can be validated with different levels
288 of concentrations (i.e., low, medium and high). The authors also employed fragmented DNA
289 samples from two pairs of closely related bacteria (i.e., E. coli/S flexneri and S. aureus/S.
290 epidermidis) to ensure similar microbial species can be properly discriminated.
291
292 Results showed 93.7% agreement with blood culture for a cohort of 350 patients with sepsis alert
293 and 62 (37.3%) out of 166 samples with no sepsis aetiology were found to contain microbial
294 cfDNA. Genetic factors such as GC content, genome size, and sequence similarity (when
295 difference > 3%) seemed to have no significant effect on the results.
296
297 Thus, the strong positive agreement for patients with sepsis alert indicated that microbial cfDNA
298 sequencing is a reliable approach for the assessment of microbial species in blood circulation.
299
300 Applications in prognosis and the study of chronic diseases
301 Certain chronic diseases may result in substantial alteration in physiology, which may be
302 reflected by microbial profile and detected by cfDNA sequencing. Diabetes and Alzheimer’s
303 disease are good examples.
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019304
305 In theory, elevated glucose level in diabetic patients may selectively provoke an outgrowth of
306 certain types of microbes yet to be identified. These microbes may themselves participate in the
307 progression of diabetes and/or patients’ health conditions. Probing the microbes in diabetic
308 patients may provide important information for treatment.
309
310 Alzheimer’s disease represents a good example for combinatorial analysis of a cfDNA sample.
311 Progressive neuronal cell death along the progression of the disease would release increasing
312 amount of related cfDNA into the bloodstream or other body fluids. Cell-free DNA sequencing
313 should be able to provide real time information to expedite the treatment of the disease.
314 Moreover, a recent study has associated herpesviruses HHV-6A and HHV-7 with the
315 development of Alzheimer’s disease (Readhead et al., 2018). Cell-free DNA sequencing may
316 allow us to correlate herpesviral cfDNA titer with the progression of the clinical symptom.
317 Furthermore, since Alzheimer’s disease is characterized as a disease with accumulation of beta-
318 amyloid peptide, it would be attempting to correlate herpesviral profile with beta-amyloid
319 profile.
320
321 Other diseases, such as Parkinson’s disease and gout, may also be good putative candidates to be
322 studied by cfDNA sequencing.
323
324 Analysis of cfDNA in non-blood body fluids
325 Besides blood plasma, there are a number of liquid biopsies and all of these specimen deserve a
326 careful scrutiny. The most accessible and feasible human liquid biopsies include saliva, urine,
327 tears, lymph, vaginal discharge, semen, cerebrospinal fluid, etc. These specimens represent
328 useful alternatives being able to provide meaningful information for the detection of a target
329 disease, such as cancer, in close proximity. Reports have indicated the strong potential of non-
330 blood cfDNA in diagnostics of diseases, especially for local neoplasms (Peng et al., 2017) and
331 infections (Burnham et al., 2018).
332 Saliva consists of a large spectrum of informative materials, including cfDNA and exosome
333 (Hyun et al., 2018), mRNA, and proteins of immunological, toxicological, hormonal and
334 therapeutic values. Applications of salivary fluid cover not only the genomics (including cancer
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019335 genomics), transcriptomic and proteomics of the individual, but also the microbiota in the oral
336 cavity. Reports have demonstrated the feasibility of using saliva in oral cancer diagnosis, and the
337 study of EGFR mutation for lung cancer patients. Lymphatic fluid has close contact with neurons
338 and thus may be used for the detection of neuronal pathogens. Vaginal discharge and semen
339 samples can be useful specimen for testing pathogens associated with sexually transmitted
340 diseases (e.g., gonorrhea and syphilis) or cancer (e.g., cervical cancer) such as Neisseria
341 gonorrhoeae that causes gonorrhea, spirochete Treponema pallidum that cause syphilis and HPV
342 that causes cervical cancer. These body fluids have close contact with certain types of tissues
343 mediated with particular pathogenic infections and thus may have collected desired genetic
344 material or microbial cells for clinical application.
345
346 Urine has also been reported to be useful for the study of cancer, such as colorectal and prostate
347 cancers, and the BRAF V600e mutations in Erdheim-Chester syndrome. The cell-free DNA
348 fragments in urine are known to be between 150–250 bp in size. However, high false negative
349 rate was reported due to limited volume or lack of advanced technology.
350
351 Urinary cfDNA was reported to be a versatile specimen for monitoring urinary tract infection
352 (UTI) (Burnham et al., 2018). UTI is a common problem for kidney transplant recipients (Abbott
353 et al., 2004), ~20% of the recipients are affected by UTI during the first year after transplantation
354 (Ariza-Heredia et al., 2014), and over 50% are affected within the first three years (Chuang et al.,
355 2005). The authors analyzed 141 urinary cfDNA samples of a cohort composed of 82 kidney
356 transplant recipients. Results indicated that a single urinary cfDNA sample is informative enough
357 to provide multiple pieces of information including microbial composition and dynamics, kidney
358 allograft injury, host response to UTI, antimicrobial susceptibility and bacterial growth
359 dynamics.
360
361 Since cfDNA fragments are predominantly from normal cells, finding those released from
362 diseased cells is like finding a needle in a haystack. To get around the problem, cfDNA analysis
363 normally requires advanced methodologies such as PCR and NGS. Moreover, selection of body
364 fluid in close proximity to diseased tissue for cfDNA analysis is also one way to improve the
365 efficacy (Burnham et al., 2018; Lin et al., 2017; Markopoulos et al., 2010; Salvi et al., 2015).
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019366
367
368 Conclusions
369 After microbiome profiling, it would be useful to track down the source tissues for microbes of
370 interest. Vlaminck et al. have demonstrated the feasibility of using organ-specific SNPs as
371 markers to distinguish between donor and recipient cfDNA sequences in lung transplantation (De
372 Vlaminck et al., 2015). To track down the source tissues for microbes, additional independent
373 information is required, but yet to be developed.
374
375 Cell-free DNA-mediated microbial species relies on NGS or SMS for detection followed by
376 alignment to microbial databases, the former determines the sensitivity, while the latter
377 determines specificity. In terms of microbial databases to be used, one has to carefully choose
378 reliable microbial genomic sequences for building reference microbial databases. Although
379 microbial genome assemblies in the public domain have been constantly updated and newly
380 discovered microbial species are constantly integrated to facilitate future studies, undoubtedly
381 significant number of microbial genomes are not yet identified and thus not included in public
382 databases. The volume is likely to grow fast in the near future.
383
384
385 Acknowledgements
386 Here, we would like to acknowledge all people who donated their samples for the study and our
387 team members for their hard work on cell-free DNA sequencing and analysis.
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019388 References
389 Abbott, K.C., Swanson, S.J., Richter, E.R., Bohen, E.M., Agodoa, L.Y., Peters, T.G., Barbour,
390 G., Lipnick, R., and Cruess, D.F. (2004). Late urinary tract infection after renal transplantation in
391 the United States. Am J Kidney Dis 44, 353-362.
392 Ariza-Heredia, E.J., Beam, E.N., Lesnick, T.G., Cosio, F.G., Kremers, W.K., and Razonable,
393 R.R. (2014). Impact of urinary tract infection on allograft function after kidney transplantation.
394 Clin Transplant 28, 683-690.
395 Bianchi, D.W., and Chiu, R.W.K. (2018). Sequencing of Circulating Cell-free DNA during
396 Pregnancy. N Engl J Med 379, 464-473.
397 Blauwkamp, T.A., Thair, S., Rosen, M.J., Blair, L., Lindner, M.S., Vilfan, I.D., Kawli, T.,
398 Christians, F.C., Venkatasubrahmanyam, S., Wall, G.D., et al. (2019). Analytical and clinical
399 validation of a microbial cell-free DNA sequencing test for infectious disease. Nat Microbiol.
400 Boyd, S.D., and Crowe, J.E., Jr. (2016). Deep sequencing and human antibody repertoire
401 analysis. Curr Opin Immunol 40, 103-109.
402 Buonaurio, R., Stravato, V.M., Kosako, Y., Fujiwara, N., Naka, T., Kobayashi, K., Cappelli, C.,
403 and Yabuuchi, E. (2002). Sphingomonas melonis sp. nov., a novel pathogen that causes brown
404 spots on yellow Spanish melon fruits. Int J Syst Evol Microbiol 52, 2081-2087.
405 Burd, E.M. (2003). Human papillomavirus and cervical cancer. Clin Microbiol Rev 16, 1-17.
406 Burnham, P., Dadhania, D., Heyang, M., Chen, F., Westblade, L.F., Suthanthiran, M., Lee, J.R.,
407 and De Vlaminck, I. (2018). Urinary cell-free DNA is a versatile analyte for monitoring infections
408 of the urinary tract. Nat Commun 9, 2412.
409 Butler, T.M., Johnson-Camacho, K., Peto, M., Wang, N.J., Macey, T.A., Korkola, J.E., Koppie,
410 T.M., Corless, C.L., Gray, J.W., and Spellman, P.T. (2015). Exome Sequencing of Cell-Free
411 DNA from Metastatic Cancer Patients Identifies Clinically Actionable Mutations Distinct from
412 Primary Disease. PLoS One 10, e0136407.
413 Chiang, Y.S., Huang, Y.F., Midha, M.K., Chen, T.H., Shiau, H.C., and Chiu, K.P. (2018). Single
414 cell transcriptome analysis of MCF-7 reveals consistently and inconsistently expressed gene
415 groups each associated with distinct cellular localization and functions. PLoS One 13,
416 e0199471.
417 Chuang, P., Parikh, C.R., and Langone, A. (2005). Urinary tract infections after renal
418 transplantation: a retrospective review at two US transplant centers. Clin Transplant 19, 230-
419 235.
420 Cummins, J., and Tangney, M. (2013). Bacteria and tumours: causative agents or opportunistic
421 inhabitants? Infect Agent Cancer 8, 11.
422 De Vlaminck, I., Martin, L., Kertesz, M., Patel, K., Kowarsky, M., Strehl, C., Cohen, G., Luikart,
423 H., Neff, N.F., Okamoto, J., et al. (2015). Noninvasive monitoring of infection and rejection after
424 lung transplantation. Proc Natl Acad Sci U S A 112, 13336-13341.
425 Fernandez-Cuesta, L., Perdomo, S., Avogbe, P.H., Leblay, N., Delhomme, T.M., Gaborieau, V.,
426 Abedi-Ardekani, B., Chanudet, E., Olivier, M., Zaridze, D., et al. (2016). Identification of
427 Circulating Tumor DNA for the Early Detection of Small-cell Lung Cancer. EBioMedicine 10,
428 117-123.
429 Finotti, A., Allegretti, M., Gasparello, J., Giacomini, P., Spandidos, D.A., Spoto, G., and
430 Gambari, R. (2018). Liquid biopsy and PCR-free ultrasensitive detection systems in oncology
431 (Review). Int J Oncol 53, 1395-1434.
432 Gawad, C., Koh, W., and Quake, S.R. (2016). Single-cell genome sequencing: current state of
433 the science. Nat Rev Genet 17, 175-188.
434 Grumaz, S., Stevens, P., Grumaz, C., Decker, S.O., Weigand, M.A., Hofer, S., Brenner, T., von
435 Haeseler, A., and Sohn, K. (2016). Next-generation sequencing diagnostics of bacteremia in
436 septic patients. Genome Med 8, 73.
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019437 Hall, K.K., and Lyman, J.A. (2006). Updated review of blood culture contamination. Clin
438 Microbiol Rev 19, 788-802.
439 Hong, D.K., Blauwkamp, T.A., Kertesz, M., Bercovici, S., Truong, C., and Banaei, N. (2018).
440 Liquid biopsy for infectious diseases: sequencing of cell-free plasma to detect pathogen DNA in
441 patients with invasive fungal disease. Diagn Microbiol Infect Dis 92, 210-213.
442 Huang, Y.F., Chen, Y.J., Fan, T.C., Chang, N.C., Chen, Y.J., Midha, M.K., Chen, T.H., Yang,
443 H.H., Wang, Y.T., Yu, A.L., et al. (2018). Analysis of microbial sequences in plasma cell-free
444 DNA for early-onset breast cancer patients and healthy females. BMC Med Genomics 11, 16.
445 Hyun, K.A., Gwak, H., Lee, J., Kwak, B., and Jung, H.I. (2018). Salivary Exosome and Cell-Free
446 DNA for Cancer Detection. Micromachines (Basel) 9.
447 Kowarsky, M., Camunas-Soler, J., Kertesz, M., De Vlaminck, I., Koh, W., Pan, W., Martin, L.,
448 Neff, N.F., Okamoto, J., Wong, R.J., et al. (2017). Numerous uncharacterized and highly
449 divergent microbes which colonize humans are revealed by circulating cell-free DNA. Proc Natl
450 Acad Sci U S A 114, 9623-9628.
451 Lamy, B., Dargere, S., Arendrup, M.C., Parienti, J.J., and Tattevin, P. (2016). How to Optimize
452 the Use of Blood Cultures for the Diagnosis of Bloodstream Infections? A State-of-the Art. Front
453 Microbiol 7, 697.
454 Levrero, M., and Zucman-Rossi, J. (2016). Mechanisms of HBV-induced hepatocellular
455 carcinoma. J Hepatol 64, S84-S101.
456 Levy, S.E., and Myers, R.M. (2016). Advancements in Next-Generation Sequencing. Annu Rev
457 Genomics Hum Genet 17, 95-115.
458 Lin, S.Y., Linehan, J.A., Wilson, T.G., and Hoon, D.S.B. (2017). Emerging Utility of Urinary Cell-
459 free Nucleic Acid Biomarkers for Prostate, Bladder, and Renal Cancers. Eur Urol Focus 3, 265-
460 272.
461 Lorian, V. (1971). Effect of antibiotics on staphylococcal hemolysin production. Appl Microbiol
462 22, 106-109.
463 Markopoulos, A.K., Michailidou, E.Z., and Tzimagiorgis, G. (2010). Salivary markers for oral
464 cancer detection. Open Dent J 4, 172-178.
465 McLaughlin, R.W., Vali, H., Lau, P.C., Palfree, R.G., De Ciccio, A., Sirois, M., Ahmad, D.,
466 Villemur, R., Desrosiers, M., and Chan, E.C. (2002). Are there naturally occurring pleomorphic
467 bacteria in the blood of healthy humans? J Clin Microbiol 40, 4771-4775.
468 Mehrotra, M., Singh, R.R., Loghavi, S., Duose, D.Y., Barkoh, B.A., Behrens, C., Patel, K.P.,
469 Routbort, M.J., Kopetz, S., Broaddus, R.R., et al. (2018). Detection of somatic mutations in cell-
470 free DNA in plasma and correlation with overall survival in patients with solid tumors.
471 Oncotarget 9, 10259-10271.
472 Mehta, S., Garg, A., Gupta, L.K., Mittal, A., Khare, A.K., and Kuldeep, C.M. (2011). Kaposi's
473 sarcoma as a presenting manifestation of HIV. Indian J Sex Transm Dis AIDS 32, 108-110.
474 Oellerich, M., Schutz, E., Beck, J., Kanzow, P., Plowman, P.N., Weiss, G.J., and Walson, P.D.
475 (2017). Using circulating cell-free DNA to monitor personalized cancer therapy. Crit Rev Clin
476 Lab Sci 54, 205-218.
477 Peng, M., Chen, C., Hulbert, A., Brock, M.V., and Yu, F. (2017). Non-blood circulating tumor
478 DNA detection in cancer. Oncotarget 8, 69162-69173.
479 Pham, V.H., and Kim, J. (2012). Cultivation of unculturable soil bacteria. Trends Biotechnol 30,
480 475-484.
481 Potgieter, M., Bester, J., Kell, D.B., and Pretorius, E. (2015). The dormant blood microbiome in
482 chronic, inflammatory diseases. FEMS Microbiol Rev 39, 567-591.
483 Pyakurel, P., Pak, F., Mwakigonja, A.R., Kaaya, E., and Biberfeld, P. (2007). KSHV/HHV-8 and
484 HIV infection in Kaposi's sarcoma development. Infect Agent Cancer 2, 4.
485 Readhead, B., Haure-Mirande, J.V., Funk, C.C., Richards, M.A., Shannon, P., Haroutunian, V.,
486 Sano, M., Liang, W.S., Beckmann, N.D., Price, N.D., et al. (2018). Multiscale Analysis of
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019487 Independent Alzheimer's Cohorts Finds Disruption of Molecular, Genetic, and Clinical Networks
488 by Human Herpesvirus. Neuron 99, 64-82 e67.
489 Roy, S., and Trinchieri, G. (2017). Microbiota: a key orchestrator of cancer therapy. Nat Rev
490 Cancer 17, 271-285.
491 Salvi, S., Gurioli, G., Martignano, F., Foca, F., Gunelli, R., Cicchetti, G., De Giorgi, U., Zoli, W.,
492 Calistri, D., and Casadio, V. (2015). Urine Cell-Free DNA Integrity Analysis for Early Detection
493 of Prostate Cancer Patients. Dis Markers 2015, 574120.
494 Tedeschi, G.G., Amici, D., Sprovieri, G., and Vecchi, A. (1976). Staphylococcus epidermidis in
495 the circulating blood of normal and thrombocytopenic human subjects: immunological data.
496 Experientia 32, 1600-1602.
497 Volik, S., Alcaide, M., Morin, R.D., and Collins, C. (2016). Cell-free DNA (cfDNA): Clinical
498 Significance and Utility in Cancer Shaped By Emerging Technologies. Mol Cancer Res 14, 898-
499 908.
500 Wade, W. (2002). Unculturable bacteria--the uncharacterized organisms that cause oral
501 infections. J R Soc Med 95, 81-83.
502 Weirather, J.L., de Cesare, M., Wang, Y., Piazza, P., Sebastiano, V., Wang, X.J., Buck, D., and
503 Au, K.F. (2017). Comprehensive comparison of Pacific Biosciences and Oxford Nanopore
504 Technologies and their applications to transcriptome analysis. F1000Res 6, 100.
505
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019Figure 1
Fig. 1. Workflow of cfDNA sequencing andcross-microbial species comparison.
The stringent workflow is designed for the identification of rare (as in healthy individuals) and predominant
opportunistic pathogens in diseased patients. The stringency relies on initial mapping with Illumina paired-
end (PE) reads followed by alignment with contigs assembled from qualified PE reads. Cross-species
comparison is based on MCRPM (microbial cfDNA reads per million) values which correlate with the titers of
corresponding cfDNA fragments in plasma.
PeerJ Preprints | https://doi.org/10.7287/peerj.preprints.27588v1 | CC BY 4.0 Open Access | rec: 15 Mar 2019, publ: 15 Mar 2019You can also read

















































