Association between oncogenic human papillomavirus type 16 and Killian polyp
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3
https://doi.org/10.1186/s13027-020-00342-3
RESEARCH ARTICLE Open Access
Association between oncogenic human
papillomavirus type 16 and Killian polyp
Lucia Oton-Gonzalez1†, John Charles Rotondo1†, Luca Cerritelli2, Nicola Malagutti2, Carmen Lanzillotti1,
Ilaria Bononi1, Andrea Ciorba2, Chiara Bianchini2, Chiara Mazziotta1, Monica De Mattei1, Stefano Pelucchi2,
Mauro Tognon1* and Fernanda Martini1*
Abstract
Background: Killian polyp (KP) is a benign lesion that arises from the maxillary sinus. The etiology of KP is
unknown. The aim of this study was to investigate the potential involvement of human papilloma- (HPV) and
polyoma-viruses (HPyV) infections in the onset of KP.
Methods: DNA from antral (n = 14) and nasal (n = 14) KP fractions were analyzed for HPV and HPyV sequences,
genotypes, viral DNA load and physical status along with expression of viral proteins and p16 cellular protein.
Results: The oncogenic HPV16 was detected in 3/14 (21.4%) antral KPs, whilst nasal KPs tested HPV-negative (0/14).
The mean HPV16 DNA load was 4.65 ± 2.64 copy/104 cell. The whole HPV16 episomal genome was detected in
one KP sample, whereas HPV16 DNA integration in two KPs. P16 mRNA level was lower in the KP sample carrying
HPV16 episome than in KPs carrying integrated HPV16 and HPV- negative KPs (p< 0.001). None of the antral and
nasal KP samples tested positive for HPyV DNA (0/28).
Conclusions: A fraction of KP tested positive for the oncogenic HPV16. HPV16 detection in the KP antral portion
may be consistent with HPV16 infection derived from the maxillary sinus. HPV16 DNA integration represents a novel
finding. Altogether, these data improve our knowledge on the association between KP and HPV infection, whereas
it indicates that the KP onset is heterogeneous.
Keywords: Killian polyp, Human papillomavirus, Polyomavirus, Infection, Nasal polyps
Introduction and a nasal/choanal fraction, emerging through an en-
Killian polyp (KP), or antrochoanal polyp, is a benign le- larged maxillary accessory ostium [3].
sion of the upper respiratory tract arising from the max- The etiopathogenesis of KP is not known. Several
illary anthrum, which may extend through the nasal studies have suggested that autoreactivity, allergies and/
cavity to the choana. KP represents about 5 and 33% of or chronic inflammation could be risk factors for KP on-
nasal polyps in adults and children, respectively [1, 2]. set [4–6]. KPs are indeed inflammatory polyps [7].
KP usually presents as unilateral pedunculated mass Schryver et al. questioned if autoreactivity contributed to
composed by an antral portion, which is usually cystic, the KP onset or it resulted from a chronic inflammation,
and proposed to investigate other inflammation causes,
* Correspondence: tgm@unife.it; mrf@unife.it such as viral infections [8]. In fact, KP recurrence after
†
Lucia Oton-Gonzalez and John Charles Rotondo contributed equally to this its incomplete surgical removal suggests that viral infec-
work.
1 tions may play a role [3, 9, 10].
Department of Medical Sciences, Laboratories of Cell Biology and Molecular
Genetics, School of Medicine, University of Ferrara, 64/B, Fossato di Mortara Different viruses are able to infect the oropharyngeal
Street, 44121 Ferrara, Italy region, and play a role in various head and neck diseases
Full list of author information is available at the end of the article
© The Author(s). 2021 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License,
which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if
changes were made. The images or other third party material in this article are included in the article's Creative Commons
licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the
data made available in this article, unless otherwise stated in a credit line to the data.Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 2 of 9
[11, 12]. Specifically, human papillomaviruses (HPV) and underwent surgical removal at the Ear, Nose and Throat
polyomaviruses (HPyV) such as BKPyV, JCPyV and Mer- Unit, University Hospital of Ferrara (Italy). Inclusion cri-
kel cell polyomavirus (MCPyV), are DNA viruses infect- teria were unilateral polyp with histopathological diagno-
ing the tonsillar tissues [13–16], and have been sis of KP and age between 18 and 80 yrs. Exclusion
associated to the development of respiratory diseases as criteria were bilateral polyps not coincident with KP.
well as to head and neck cancer [17–20]. HPV and Written informed consent was obtained from all pa-
HPyV display similar biological behavior in infected tar- tients. The study was conducted in accordance with the
get tissues. After infection of epithelial cells, HPV and Declaration of Helsinki. The protocol was approved by
HPyV may multiply and spread in different anatomical the County Ethical Committee (ID:160986).
sites, or enter lifelong latent phase, whereby viral DNA
is maintained at low copy number [21, 22]. In some in- Nucleic acids extraction
stance, long term latency of the oncogenic HPV and KP tissue samples (n=14) were divided into two por-
HPyV types may result in viral DNA integration into the tions: the antral (n=14) and the nasal portion (n=14).
host cell genome, leading to cell transformation upon Samples (n=28) were incubated overnight with protein-
viral oncoprotein overexpression [21–25]. ase K at 56 °C to allow tissue digestion. Then, nucleic
The association between HPV infection and KP has acids were simultaneously extracted from samples using
been poorly investigated, whereas studies on HPyV in KP the All Prep DNA/RNA extraction kit (Qiagen, Milan,
are missing. HPV sequences have been found at different Italy). DNA from KPs was isolated/purified together with
prevalence, ranging 0–54% [26–28]. Moreover, oncogenic a salmon sperm DNA (ssDNA) sample and a mock sam-
HPV genotypes such as HPV16, have been found to be ple lacking DNA [30]. After purification, DNAs/RNAs
prevalent in KPs, raising the question if HPV may play were quantified spectrophotometrically (NanoDrop
role in cell transformation. One recent study focusing on 2000, Thermo Scientific) [31]. DNA amplification suit-
tumor marker expression, such as p16 and viral oncopro- ability was evaluated by β-globin gene PCR [32]. DNA/
teins, did not find any correlation between HPV DNA RNAs were stored at − 80 °C until time of analysis.
positivity and KP development, concluding that HPV la-
tently infects KP [27]. However, HPV DNA load and phys- Detection of HPV and HPyV DNAs
ical status, which are two main hallmarks of latent or KP tissue samples were tested for HPV and BKPyV,
active infection, have not been assessed yet in KP [29]. JCPyV and MCPyV DNA sequences, by quantitative
Even though maxillary sinus viral infections are con- PCR (qPCR). Fifty ng of human genomic DNA were
sidered risk factors for KP, there is no evidence proving used in 10 μl qPCR reactions. For HPV DNA detection
the KP etiopathogenesis from this infection. So far, stud- the universal primers GP5+/GP6+ (Table 1) were used,
ies focusing on the identification of viral infections have as previously reported [33, 34]. These primers allow sim-
analyzed bulk KP tissues without diversifying between ultaneous amplification of several HPV types [35, 36],
the antral and nasal components. This distinction would including those frequently detected in KP, such as
be particularly important to understand if viral infections HPV6/11/16/18 [27, 28]. QPCR reactions included 2x of
may play a role in the KP onset. In fact, any viral se- the SsoAdvanced Universal SYBR Green Supermix, Bio-
quences detected in the antral region might account for Rad (Hercules, CA, USA) and a final concentration of
maxillary sinus infections, and therefore potentially in- 0.5 μM for each GP5+/GP6+ primer. For HPyV DNA
volved in the onset of KP, while those in the nasal region detection, specific primers for BKPyV, JCPyV and
might be due to nasopharyngeal infections after the KP MCPyV were employed [25, 38, 39]. QPCR reactions in-
formation, thus not relevant for KP onset. cluded 2x of the TaqMan Universal Master Mix II, no
The aim of this study was to investigate the potential UNG, Thermo Fisher Scientific (Waltham, MA, USA),
involvement of HPV and HPyV infections in the onset and 1X of primers and probe assays (Table 1). Recom-
of KP. To this purpose, tissue samples from KP were di- binant plasmids containing HPV16 genome and BKPyV,
vided into antral and nasal parts, and analyzed separately JCPyV, and MCPyV genomes were used as positive con-
for HPVs and HPyVs sequences, genotypes, DNA load trols [25, 38, 39], whereas ssDNA and mock samples
and physical status (episomal vs integrated), and expres- lacking of DNA, as negative controls of DNA extraction
sion, along with expression levels of p16, which is a cell and PCR amplification. Each assay was run in triplicate.
protein strictly associated to active HPV infection.
HPV DNA load, genotype and physical status analyses
Materials and methods HPV DNA load was quantified by qPCR assay using the
Samples GP5+/GP6+ primers and a 10-fold dilutions standard
Killian polyp (KP) tissue specimens were collected from curve, from 108 to 102 copies, of recombinant plasmids.
14 patients (Mean age ± SD; 44 ± 18 years) who HPV DNA load values were reported as viral copies perOton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 3 of 9
Table 1 Primers used in qPCR to detect and quantify HPV, PyV DNA, viral, cellular genes
Target Primers Primers sequence (5′→ 3′) Amplicon Annealing References
names size (bp) temp. (°C)
DNA
Viral HPV L1 GP5+ TTTGTTACTGTGGTAGATACTAC 139–145 48 Malagutti et al. 2020 [33]; Tognon et al. 2020
[34]; Rotondo et al. 2020a [35]
GP6+ GAAAAATAAACTGTAAATCATATTC
HPV16 E2 E- HPV16 E2 AACGAAGTATCCTCTCCTGAAATTAT 82 60 Peitsaro, Johansson, e Syrjänen 2002 [37]
F TAG
E- HPV16 E2 CCAAGGCGACGGCTTTG
R
E Probe [ROX] CACCCCGCCGCGACCCATA
16E2PRO [BHQ2]
HPV16 E6 I+E- HPV16 GAGAACTGCAATGTTTCAGGACC 81 60
E6 F
I+E- HPV16 TGTATAGTTGTTTGCAGCTCTGTGC
E6 R
I+E Probe [6FAM] CAGGAGCGACCCAGAAAG
16E6PRO TTACCACAGTT [BHQ1]
MCPyV RQ MCPyV_ CCACAGCCAGAGCTCTTCCT Tagliapietra et al. 2020 [38]
LT.1F
RQ MCPyV_ TGGTGGTCTCCTCTCTGCTACTG
LT.1R
RQ MCPyV_ [6FAM] TCCTTCTCAGCGTCCCAG
LT Probe GCTTCA [MGB]
JCPyV Assay_JCyV AI1RWNE Tagliapietra et al. 2019 [39]
BKPyV Assay_BKyV AI20UTM
β-Globin β-Globin F TGGGTTTCTGATAGGCACTGACT 152 56 Contini et al. 2018 [32]
Host
β-Globin R AACAGCATCAGGAGTGGACAGAT
RNA
Viral HPV16 E2 HPV16 E2 F AACGAAGTATCCTCTCCTGAAATTAT 82 60 Peitsaro, Johansson, e Syrjänen 2002 [37]
TAG
HPV16 E2 R CCAAGGCGACGGCTTTG
HPV16 E6 HPV16 E6 F GAGAACTGCAATGTTTCAGGACC 81 60
HPV16 E6 R TGTATAGTTGTTTGCAGCTCTGTGC
HPV16 E5 16-E5 FWD CGTCCGCTGCTTTTGTCTGTGTCTAC 89 60 Weyn et al. 2011 [40]
ATAC
16-E5 REV CACCTAAACGCAGAGGCTGCTGTT
ATCCAC
HPV16 E7 E7 FWD AGGAGGATGAAATAGATGGTCCAG 112 60 Pett et al. 2006 [41]
E7 REV CTTTGTACGCACAACCGAAGC
P16INK4A p16 ink4a CCAACGCACCGAATAGTTACG 58 60 Marcoux et al. 2013 [42]
Host FWD
p16 ink4a GCGCTGCCCATCATCATG
REV
GAPDH GAPDH F GAAGGTGAAGGTCGGAGTC 226 60 Xiao et al. 2011 [43]
GAPDH R GAAGATGGTGATGGGATTTC
human cell equivalents (viral copy/cell). Samples were differential melting temperature (Tm), adding a high
normalized vs. HPV16-positive SiHa cell line, which resolution melting (HRM) step, from 65 °C to 95 °C
contains one HPV16 copy/cell. Human β-globin gene (ramping 0.1 °C every 0.03 s), to the qPCR analysis, as
was used to determine the human cell equivalents of done before for detection of the HPV16 and HPV18 ge-
each sample [32]. HPV genotype was determined by notypes [44]. HPV6/11/16/18 plasmids were used asOton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 4 of 9
positive controls. HPV DNA physical status was investi- Results
gated using the E2/E6 ratio by qPCR, as previously de- Prevalence of HPV and HPyV sequences
scribed (Table 1) [37]. Briefly, 50 ng of template DNA DNAs isolated from KP tissue samples (n=28) repre-
were analyzed in 10 μl multiplex PCR reactions, 2x Taq- sented by antral (n=14) and nasal (n=14) portions were
Man Universal Master Mix II, no UNG, Thermo Fisher tested for viral DNA sequences of HPV and HPyVs. The
Scientific (Waltham, MA, USA); 0.3 μM of each HPV16 qPCR analyses showed that 3/14 (21.4%) of the antral
E2 primer; 0.5 μM of each HPV16 E6 primer; and KP tissues were positive for HPV DNA (Table 2). None
0.1 μM of each E2 and E6 probe. E2/E6 ratio equal to 1 of the nasal KP samples (n=14) tested positive for HPV
indicated episomal form, less or more than 1 mixed DNA (0/14; 0%) (Table 2). KP tissue samples analyzed
forms, i.e. episomal and integrated, whereas no E2 DNA for BKPyV, JCPyV and MCPyV DNA sequences gave
detection indicated full integration. Each assay was run negative results in both antral (n=14) and nasal (n=14)
in triplicate. portions (0/14; 0%) (Table 2).
HPV DNA load, genotyping, and physical status analyses
Rolling circle amplification (RCA) assay
HPV DNA load was determined by comparison to the
The episomal viral DNAs were detected by rolling circle
HPV plasmid standard curve in qPCR assay. The mean
amplification (RCA) assay using the TempliPhi™ 100
viral DNA load in HPV-positive antral KPs (n=3) was
Amplification Kit (GE Healthcare, Chicago, USA) [45],
4.65±2.64 copy/104 cell. In detail, in the three HPV-
and in accordance with manufacturer’s instructions.
positive antral KP samples, the viral DNA load was 8.32
Briefly, reactions were prepared with 25 ng of genomic
copy/104 cell, 3.43 copy/104 cell, and 2.21 copy/104 cell.
DNA and 175 μM of dNTP mix (Thermo Scientific,
HPV genotype analyses were carried out by HRM qPCR
Massachusetts, USA). The specificity of the RCA prod-
assay. Firstly, the optimal Tm range for discriminating
ucts was assessed by DNA restriction enzyme digestion
HPV6/11/16/18 types from GP5+/GP6+ amplicons was
in a final volume of 10 μL (Thermo Scientific, Massachu-
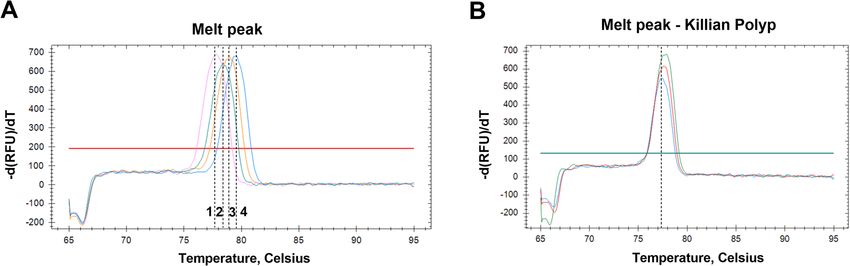
identified, which was between 75.4–79.5±0.2 °C (Fig. 1a).
setts, USA). RCA and digested RCA products were visu-
HPV genotype analyses were carried out by comparing
alized onto a 0.8% agarose gel. Positive and negative
qPCR Tm with the positive controls. Results indicated
controls were used in the RCA assay.
that the three HPV-positive antral KP samples carried
the HPV type 16 (3/3; 100%) (Fig. 1b).
Gene expression analysis HPV16 DNA physical status was assessed by E2/E6 ra-
Total RNA was retrotranscribed using the Improm II tio in the three HPV16-positive antral KP samples. The
(Promega, Wisconsin, USA) reverse transcription sys- E2/E6 ratio was 1.01 in one sample (1/3; 33.3%) indicat-
tem [46]. cDNAs were analyzed for the expression of ing presence of HPV16 in the episomal form. In the two
HPV16 E2, E6, E7 and E5 genes and p16 cellular other samples only the E6 sequence was found (2/3;
gene (Table 1) [37, 40–42]. Briefly, 50 ng of cDNA 66.6%), indicating that HPV16 was integrated into the
were used in 10 μl reaction, 2x of the SsoAdvanced host cell genome.
Universal SYBR Green Supermix, Bio-Rad (Hercules,
CA, USA) and a final concentration of 0.5 μM for HPV physical status validation by RCA
each primer [47]. GAPDH gene was employed as con- Antral KP DNAs (n=14) were investigated by RCA for
trol for the gene expression analysis [43]. SiHa cell validating the HPV DNA episomal physical status. Suc-
line was used as positive control for HPV gene ex- cessfully amplification was obtained only in the KP sam-
pression and mock sample as negative control. Each ple detected with E2/E6 ratio of 1.01, that was
assay was run in triplicate. predictable of the episomal form. The molecular weight
for the positive band corresponding to approximately
8000 bp was consistent of the HPV genome (Fig. 2, lane
Statistical analyses S2). Digestion with Bam HI enzyme, which cuts once
Statistical analyses were performed using the GraphPad
Prism for Windows (version 6.0, GraphPad, California,
Table 2 Prevalence of HPV and HPyV in antral and nasal KP
USA) [48, 49]. For mRNA, fold change was calculated by tissues
the 2-ΔΔCt method and represented in Log2 scale, using
Tissue sample Number of positive samples/samples analyzed (%)
HPV-negative samples as controls [31]. One-way ana-
HPV MCPyV JCPyV BKPyV
lysis of variance was used to compare fold-change
Antral KP 3/14 (21.4) 0/14 (0) 0/14 (0) 0/14 (0)
among samples [50]. P values less than 0.05 were consid-
ered statistically significant (p< 0.05) [51]. Nasal KP 0/14 (0) 0/14 (0) 0/14 (0) 0/14 (0)Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 5 of 9
Fig. 1 HPV differential melt peaks. a Melting temperature (Tm) for; 1) pUC19_HPV16; 2) pUC19_HPV11; 3) pUC19_HPV6 and 4) pUC19_HPV18. b
Tm for KP samples, corresponding to that of pUC19_HPV16
into HPV16 genome, further confirmed the positivity for fold lower in HPV16-episome KP sample than in HPV-
HPV16. negative samples (p< 0.0001, Fig. 3). Although not statis-
tically significant, p16 expression was slightly higher in
Gene expression analysis HPV16-integrated KP samples compared to HPV-
Viral gene expression was studied for the HPV-positive negative samples (p> 0.05, Fig. 3).
antral KP samples (n=3). No expression for HPV16 E2,
E5, E6, E7 genes was detectable in any of the samples Discussion
analyzed, indicating either that HPV16 is not transcrip- The etiopathogenesis of KP is still not completely under-
tionally active in KP or that viral mRNA levels were too stood. Viral infections have been suggested to be in-
low to be detected under qPCR conditions. To gain volved in KP onset [52, 53]. Herein, with the aim to
insight into this topic, p16, which is considered a surro- verify the putative role of the viral infections, HPV and
gate marker of active HPV infection, was analyzed for HPyV were investigated in KP samples. Independent
mRNA expression in the three antral HPV-positive KP analyses of the antral and nasal region were useful in un-
samples, containing the episomal HPV16 (n=1) and the derstanding whether the KP infections depended on the
integrated HPV16 (n=2). Results indicated that p16 maxillary sinus or the nasal cavity.
mRNA level was 8.01-fold lower in HPV16-episome KP HPV sequences were detected in 21% of the antral KP
sample than in HPV16-integrated KP samples and 7.05- samples, while none of the nasal samples tested positive
Fig. 2 Rolling circle amplification assay performed on antral KP DNAs. MW: Molecular Weight. Negative controls: H2O, Salmon Sperm DNA (SS),
Neg digestion (H2O). KP DNAs 1–14Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 6 of 9
reported in HPV-positive normal cervical samples [70].
Regard KP, evidences proving its neoplastic transform-
ation do not exist, although some cases mimicking ma-
lignant transformation have been reported [71].
Nevertheless, HPV carcinogenesis in KP, if any, could be
difficult to be assessed, since KPs are removed early after
presentation, whereas HPV transformation process oc-
curs in long lasting time, needing many years to be de-
tected. Altogether, our data indicate that HPV16 is
present at low DNA load in both episomal and inte-
grated form, consistent with latent/persistent infection
in the antral KP. Nevertheless, the detection of the onco-
genic HPV16 combined with its DNA integration in the
KP is intriguing. Further studies are needed to assess the
HPV DNA integration in KP over the time.
HPV mRNA expressions occur during active viral
infection. Accordingly, in this study, viral expression
of E2, E5, E6 and E7 sequences was not detected in
Fig. 3 p16 mRNA expression. KPs (n=14) were stratified according to
HPV positivity/negativity. HPV-positive were further divided in the HPV-positive KP samples. Although E6/E7 ex-
episomal (n=1) and integrated (n=2). ****p< 0.0001 pression in HPV-positive KP samples carrying viral
DNA integration would be expected, HPV latency in
normal and pathological tissues presenting viral DNA
for HPV. This result indicates that the KP antral region integration is also common [72]. Some other explana-
is target of HPV infection and suggests a possible link tions may account for lack of viral expression. For in-
between maxillary sinus infections and KP development. stance, KPs are covered by ciliated cylindrical
In term of prevalence, our data are in agreement with epithelium, which may be not permissive for HPV
previous studies reporting HPV rates ranging from 0 to E6/E7 gene expression [7, 67]. Also, it is possible that
54% in KP samples, although the new methodological viral mRNA levels were too low to be detected under
approach used herein does not allow our and previous our qPCR conditions. Further studies with more sen-
data to be compared adequately [26, 27, 54–60]. sitive assays may clarify this matter [46].
HPV genotypes have been investigated in two previous Since no HPV transcriptional activity was found, the
studies reporting HPV16 to be frequently detected at higher surrogate marker of active HPV infection, the p16, was
rate, 61.9 and 85.72%, respectively, than HPV11, 14.3 and studied in correlation to infection. During HPV infection
14.28%, respectively [27, 28]. In this study HPV16 was the the viral E7 protein inactivates pRb tumor suppressor pro-
only viral genotype detected in KP. These results are of tein leading to p16 overexpression [73]. In this study, no
interest as HPV16 is the high risk oncogenic type involved difference between HPV16-positive KP samples carrying
in development of different tumors [24, 61–63], including integrated viral DNA and HPV-negative KPs was observed
head and neck cancer [64, 65]. (p> 0.05), although a slightly higher p16 mRNA level was
HPV viral load and physical status are indicative of ac- found in HPV16-positive KPs. Likely, the small samples
tive or latent infection in the infected tissues [29, 66]. size used in the study did not allow statistical significance
For the first time, DNA load and physical status was in- to be reached. In contrast, the KP sample carrying epi-
vestigated in HPV-positive KP samples. The viral DNA somal HPV DNA showed stronger p16 down-expression
load was lower than 1 copy/cell, which is consistent with compared to HPV-positive and HPV-negative KP samples
latent or persistent infection occurring in normal tissues (p< 0.001). Mutations at the p16 coding gene may explain
[67, 68]. When HPV physical status was analyzed a het- its down-expression [74, 75]. Alternatively, methylation at
erogeneous trend was found among the HPV-positive p16 promoter may silence the gene leading to decrease ex-
KP samples. One sample carried HPV16 in episomal pression, as previously shown in HPV-positive samples
form, which was confirmed amplifying the whole HPV carrying HPV in episomal form [76].
genome by RCA assay. Instead, two KP samples showed Finally, HPyV DNA sequences were analyzed in KP.
the HPV16 DNA in integrated form. This is an interest- HPyVs have been found associated to different diseases,
ing finding because high risk HPV integration into the including cancer and polyposis [77]. Specifically, JCPyV
host cell genome is a common event preceding cell has been studied in correlation to colon polyposis [78],
transformation [69]. On the other hand, HPV integration whereas BKPyV has been investigated in the prostate and
occurs up to 42.8% of normal tissues, as previously colon cancer onset [79]. MCPyV is the main cause of theOton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 7 of 9
Merkel cell carcinoma, a rare but very aggressive non- Author details
1
melanoma skin cancer [25]. Moreover, MCPyV is consid- Department of Medical Sciences, Laboratories of Cell Biology and Molecular
Genetics, School of Medicine, University of Ferrara, 64/B, Fossato di Mortara
ered to be a part of the skin microbiota, and viral DNA se- Street, 44121 Ferrara, Italy. 2Department of Biomedical Sciences and
quences have been found in nasal swabs, blood, chorionic Specialistic Surgeries, ENT Section, University of Ferrara and University
villi, eyebrows and adrenal glands [38, 77, 80]. In this Hospital of Ferrara, 8, Aldo Moro Square, 44124 Cona, Italy.
study, HPyV sequences were not found neither in antral Received: 19 November 2020 Accepted: 25 December 2020
nor nasal KPs, thus excluding their role in KP formation.
References
Conclusions 1. Chaiyasate S, Roongrotwattanasiri K, Patumanond J, Fooanant S.
The present study investigated HPV and HPyV as poten- Antrochoanal polyps: how long should follow-up be after surgery? Int J
tial pathogenic risk factors in KP. While no implication Otolaryngol. 2015;2015:297417.
2. Aksakal C. Bilateral antrochoanal polyp in a child. J Craniofac Surg. 2018;
was found for HPyV, a fraction of KPs showed positivity 29(8):2368–9.
for HPV16. New information on HPV DNA load and 3. Frosini P, Picarella G, De Campora E. Antrochoanal polyp: analysis of 200
physical status in KPs were also provided. Specifically, cases. Acta Otorhinolaryngol Ital Organo Uff Della Soc Ital Otorinolaringol E
Chir Cerv-facc. 2009;29(1):21–6.
HPV16-positive KPs presented viral DNA at low load and 4. Piquet JJ, Chevalier D, Leger GP, Rouquette I, Leconte-Houcke M. Endonasal
in episomal or integrated form. The reduced sample size microsurgery of antro-choanal polyps. Acta Otorhinolaryngol Belg. 1992;
employed in this pilot study could be considered a limita- 46(3):267–71.
5. Cook PR, Davis WE, McDonald R, McKinsey JP. Antrochoanal polyposis: a
tion, and further studies in a larger samples size are review of 33 cases. Ear Nose Throat J. 1993;72(6):401–2 404–10.
needed, especially for clarifying the oncogenic HPV16 in- 6. Lee T-J, Huang S-F. Endoscopic sinus surgery for antrochoanal polyps in
tegration into the KPs. Of note, KP samples were divided children. Otolaryngol--Head Neck Surg Off J Am Acad Otolaryngol-Head
Neck Surg. 2006;135(5):688–92.
in antral and nasal portions, whereas HPV sequences were 7. Hirshoren N, Neuman T, Gross M, Eliashar R. Angiogenesis in chronic
found only in the antral region, providing a possible ex- rhinosinusitis with nasal polyps and in antrochoanal polyps. Inflamm Res.
planation for polyp formation from sinus maxillary infec- 2011;60(4):321–7.
8. Schryver ED, Calus L, Bonte H, Natalie DR, Gould H, Donovan E, et al. The
tions. We suggest that a HPV latent infection of the quest for autoreactive antibodies in nasal polyps. J Allergy Clin Immunol.
maxillary sinus might be responsible for its recurrence, 2016;138(3):893–895.e5.
after KP surgical removal, highlighting the importance of 9. Stierna PL. Nasal polyps: relationship to infection and inflammation. Allergy
Asthma Proc. 1996;17(5):251–7.
complete surgical removal of the HPV-positive patho- 10. Yaman H, Yilmaz S, Karali E, Guclu E, Ozturk O. Evaluation and management
logical tissue to prevent further recurrences. of antrochoanal polyps. Clin Exp Otorhinolaryngol. 2010;3(2):110.
11. Kobayashi K, Hisamatsu K, Suzui N, Hara A, Tomita H, Miyazaki T. A review of
HPV-related head and neck cancer. J Clin Med. 2018;7(9).
Abbreviations
12. Poluschkin L, Rautava J, Turunen A, Wang Y, Hedman K, Syrjänen K, et al.
KP: Killian polyp; HPV: Human papillomavirus; HPyV: Polyomaviruses;
Polyomaviruses detectable in head and neck carcinomas. Oncotarget. 2018;
RCA: Rolling circle amplifications; MCPyV: Merkel cell polyomavirus;
9(32):22642–52.
ssDNA: Salmon sperm DNA; qPCR: quantitative PCR
13. Goudsmit J, Wertheim-van Dillen P, van Strien A, van der Noordaa J. The
role of BK virus in acute respiratory tract disease and the presence of BKV
Authors’ contributions DNA in tonsils. J Med Virol. 1982;10(2):91–9.
For research articles with several authors, a short paragraph specifying their 14. Monaco MC, Jensen PN, Hou J, Durham LC, Major EO. Detection of JC virus
individual contributions must be provided. The following statements should DNA in human tonsil tissue: evidence for site of initial viral infection. J Virol.
be used “Conceptualization, F.M. and S.P.; methodology, L.O.G. and J.C.R..; 1998;72(12):9918–23.
software, M.D.M.; validation, M.T., S.P., F.M.; formal analysis, L.O.G., J.C.R.; 15. Kantola K, Sadeghi M, Lahtinen A, Koskenvuo M, Aaltonen L-M, Möttönen
investigation, L.O.G., J.C.R., C.L., C.M., I.B. M.D.M.,; resources, L.C., N.M., A.C., C.B., M, et al. Merkel cell polyomavirus DNA in tumor-free tonsillar tissues and
S.P.; data curation L.O.G., J.C.R., C.L., C.M., I.B..; writing-original draft prepar- upper respiratory tract samples: implications for respiratory transmission and
ation, L.O.G., J.C.R..; writing, review and editing, C.L., M.T., S.P., F.M..; latency. J Clin Virol Off Publ Pan Am Soc Clin Virol. 2009;45(4):292–5.
visualization, L.O.G..; supervision, M.T., S.P., F.M..; project administration, M.T., 16. Rieth KKS, Gill SR, Lott-Limbach AA, Merkley MA, Botero N, Allen PD, et al.
S.P., F.M..; funding acquisition, J.C.R., M.T., F.M.. All authors have read and Prevalence of high-risk human papillomavirus in tonsil tissue in healthy
agreed to the published version of the manuscript. adults and colocalization in biofilm of tonsillar crypts. JAMA Otolaryngol--
Head Neck Surg. 2018;144(3):231–7.
17. Bialasiewicz S, Lambert SB, Whiley DM, Nissen MD, Sloots TP. Merkel cell
Funding polyomavirus DNA in respiratory specimens from children and adults.
This research was funded by the University of Ferrara, FAR grants (2017/2018 Emerg Infect Dis. 2009;15(3):492–4.
to MT and FM) and FIR grants 2016, 2017, 2018 to FM; Associazione Italiana 18. Abedi Kiasari B, Vallely PJ, Klapper PE. Merkel cell polyomavirus DNA in
per la Ricerca sul Cancro (AIRC), Milan, Contract grant number: IG 21617 to immunocompetent and immunocompromised patients with respiratory
M.T. and 21956 to J.C.R.. J.C.R was a post-doctoral fellow of the Fondazione disease. J Med Virol. 2011;83(12):2220–4.
Umberto Veronesi, Milan, Italy (2019–2020). 19. Gillison ML, Alemany L, Snijders PJF, Chaturvedi A, Steinberg BM, Schwartz
S, et al. Human papillomavirus and diseases of the upper airway: head and
Availability of data and materials neck cancer and respiratory papillomatosis. Vaccine. 2012;30(Suppl 5):
Data and material will be available upon request to the corresponding F34–54.
author. 20. Shikova E, Emin D, Alexandrova D, Shindov M, Kumanova А, Lekov A, et al.
Detection of merkel cell polyomavirus in respiratory tract specimens.
Intervirology. 2017;60(1–2):28–32.
Competing interests 21. Rotondo JC, Mazzoni E, Bononi I, Tognon M, Martini F. Association between
The authors declare no conflict of interest. human tumours and simian virus 40. Fontiers Oncol. 2019;9 In press.Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 8 of 9
22. Krump NA, Liu W, You J. Mechanisms of persistence by small DNA tumor 44. de Araujo MR, De Marco L, Santos CF, Rubira-Bullen IRF, Ronco G, Pennini I,
viruses. Curr Opin Virol. 2018;32:71–9. et al. GP5+/6+ SYBR green methodology for simultaneous screening and
23. Egawa N, Egawa K, Griffin H, Doorbar J. Human papillomaviruses; epithelial quantification of human papillomavirus. J Clin Virol Off Publ Pan Am Soc
tropisms, and the development of neoplasia. Viruses. 2015;7(7):3863–90. Clin Virol. 2009;45(2):90–5.
24. Preti M, Rotondo JC, Holzinger D, Micheletti L, Gallio N, McKay-Chopin S, 45. da Silva FRC, Cibulski SP, Daudt C, Weber MN, Guimarães LLB, Streck AF,
et al. Role of human papillomavirus infection in the etiology of vulvar et al. Novel bovine papillomavirus type discovered by rolling-circle
cancer in Italian women. Infect Agent Cancer. 2020;15:20. amplification coupled with next-generation sequencing. Aguayo FR,
25. Rotondo JC, Bononi I, Puozzo A, Govoni M, Foschi V, Lanza G, et al. Merkel curatore. PLoS ONE. 2016;11(9):e0162345.
cell carcinomas arising in autoimmune disease affected patients treated 46. Torreggiani E, Rossini M, Bononi I, Pietrobon S, Mazzoni E, Iaquinta MR, et al.
with biologic drugs, including anti-TNF. Clin Cancer Res. 2017;23(14):3929– Protocol for the long-term culture of human primary keratinocytes from the
34. normal colorectal mucosa. J Cell Physiol. 2019;234(7):9895–905.
26. Pei F, Chen X-P, Zhang Y, Wang Y, Chen Q, Tan X-J, et al. Human 47. Rotondo JC, Borghi A, Selvatici R, Mazzoni E, Bononi I, Corazza M, et al.
papillomavirus infection in nasal polyps in a Chinese population. J Gen Virol. Association of retinoic acid receptor β gene with onset and progression of
2011;92(Pt 8):1795–9. lichen sclerosus–associated vulvar squamous cell carcinoma. JAMA
27. Knör M, Tziridis K, Agaimy A, Zenk J, Wendler O. Human papillomavirus Dermatol. 2018;154(7):819 Available at: http://archderm.jamanetwork.com/
(HPV) prevalence in nasal and antrochoanal polyps and association with article.aspx?doi=10.1001/jamadermatol.2018.1373. [citato 8 gennaio 2019].
clinical data. PLoS One. 2015;10(10):e0141722. 48. Mazzoni E, Martini F, Corallini A, Taronna A, Barbanti-Brodano G, Querzoli P,
28. Yılmaz E, Alatas N, Ucar F, Cora T, Buruk K, Unlu Y. Investigation of human et al. Serologic investigation of undifferentiated nasopharyngeal carcinoma
papillomavirus (HPV) and epstein-barr virus (EBV) in antrochoanal polyps. and simian virus 40 infection. Head Neck. 2016;38(2):232–6.
Am J Otolaryngol. 2019;40(3):389–92. 49. Rotondo JC, Oton-Gonzalez L, Selvatici R, Rizzo P, Pavasini R, Campo GC,
29. McBride AA, Warburton A. The role of integration in oncogenic progression et al. SERPINA1 gene promoter is differentially methylated in peripheral
of HPV-associated cancers. PLoS Pathog. 2017;13(4):e1006211. blood mononuclear cells of pregnant women. Front Cell Dev Biol. 2020;8
30. Rotondo JC, Candian T, Selvatici R, Mazzoni E, Bonaccorsi G, Greco P, et al. Available at: https://www.frontiersin.org/article/10.3389/fcell.2020.550543/full.
Tracing males from different continents by genotyping JC polyomavirus in [citato 3 settembre 2020].
DNA from semen samples. J Cell Physiol. 2017;232(5):982–5. 50. Mazzoni E, Pietrobon S, Masini I, Rotondo JC, Gentile M, Fainardi E, et al.
31. Rotondo JC, Giari L, Guerranti C, Tognon M, Castaldelli G, Fano EA, et al. Significant low prevalence of antibodies reacting with simian virus 40
Environmental doses of perfluorooctanoic acid change the expression of mimotopes in serum samples from patients affected by inflammatory
genes in target tissues of common carp. Environ Toxicol Chem. 2018;37(3): neurologic diseases, including multiple sclerosis. PLoS One. 2014;9(11):e110923.
942–8. 51. Mazzoni E, Di Stefano M, Fiore JR, Destro F, Manfrini M, Rotondo JC, et al.
32. Contini C, Rotondo JC, Magagnoli F, Maritati M, Seraceni S, Graziano A, et al. Serum IgG antibodies from pregnant women reacting to mimotopes of simian
Investigation on silent bacterial infections in specimens from pregnant virus 40 large T antigen, the viral oncoprotein. Front Immunol. 2017;8:411.
women affected by spontaneous miscarriage. J Cell Physiol. 2018;234(1): 52. Galluzzi F, Pignataro L, Maddalone M, Garavello W. Recurrences of surgery
100–7. for antrochoanal polyps in children: A systematic review. Int J Pediatr
33. Malagutti N, Rotondo JC, Cerritelli L, Melchiorri C, De Mattei M, Selvatici R, Otorhinolaryngol. 2018;106:26–30.
et al. High human papillomavirus DNA loads in inflammatory middle ear 53. Hong SK, Min YG, Kim CN, Byun SW. Endoscopic removal of the antral
diseases. Pathog Basel Switz. 2020;9(3). portion of antrochoanal polyp by powered instrumentation. Laryngoscope.
34. Tognon M, Tagliapietra A, Magagnoli F, Mazziotta C, Oton-Gonzalez L, 2001;111(10):1774–8.
Lanzillotti C, et al. Investigation on spontaneous abortion and human 54. Becker M, Forslund O, Hansson BG, Malm L. Search for the human
papillomavirus infection. Vaccines. 2020;8(3):473. papillomavirus in nasal polyps, using a polymerase chain reaction-method. J
35. Rotondo JC, Oton-Gonzalez L, Mazziotta C, Lanzillotti C, Iaquinta MR, Tognon Otolaryngol. 1994;23(5):344–6.
M, et al. Simultaneous detection and viral DNA load quantification of different 55. Hoffmann M, Kahn T, Goeroegh T, Lohrey C, Gottschlich S, Meyer J, et al.
human papillomavirus types in clinical specimens by the high analytical Tracing human papillomavirus DNA in nasal polyps by polymerase chain
droplet digital PCR method. Front Microbiol. 2020; In press. Available at: reaction. Acta Otolaryngol (Stockh). 2000;120(7):872–5.
https://www.frontiersin.org/articles/10.3389/fmicb.2020.591452/abstract. 56. Hoffmann M, Klose N, Gottschlich S, Görögh T, Fazel A, Lohrey C, et al.
36. Evans MF, Adamson CSC, Simmons-Arnold L, Cooper K. Touchdown General Detection of human papillomavirus DNA in benign and malignant sinonasal
Primer (GP5+/GP6+) PCR and optimized sample DNA concentration support neoplasms. Cancer Lett. 2006;239(1):64–70.
the sensitive detection of human papillomavirus. BMC Clin Pathol. 2005;5:10. 57. Zaravinos A, Bizakis J, Spandidos DA. Prevalence of human papilloma virus
37. Peitsaro P, Johansson B, Syrjanen S. Integrated human papillomavirus type 16 and human herpes virus types 1-7 in human nasal polyposis. J Med Virol.
is frequently found in cervical cancer precursors as demonstrated by a novel 2009;81(9):1613–9.
quantitative real-time PCR technique. J Clin Microbiol. 2002;40(3):886–91. 58. Sham CL, Tol KF, Chan PKS, Lee DLY, Tong MCF, van Hasselt CA. Prevalence
38. Tagliapietra A, Rotondo JC, Bononi I, Mazzoni E, Magagnoli F, Oton of human papillomavirus, Epstein-Barr virus, p21, and p53 expression in
Gonzalez L, et al. Droplet-digital PCR assay to detect Merkel cell sinonasal inverted papilloma, nasal polyp, and hypertrophied turbinate in
polyomavirus sequences in chorionic villi from spontaneous abortion Hong Kong patients. Head Neck. 2012;34(4):520–33.
affected females. J Cell Physiol. 2020. 59. Rizzo R, Malagutti N, Bortolotti D, Gentili V, Rotola A, Fainardi E, et al.
39. Tagliapietra A, Rotondo JC, Bononi I, Mazzoni E, Magagnoli F, Maritati M, Infection and HLA-G molecules in nasal polyposis. J Immunol Res. 2014;
et al. Footprints of BK and JC polyomaviruses in specimens from females 2014:407430.
affected by spontaneous abortion. Hum Reprod Oxf Engl. 2019. 60. Pagella F, Emanuelli E, Pusateri A, Borsetto D, Cazzador D, Marangoni R,
40. Weyn C, Vanderwinden J-M, Rasschaert J, Englert Y, Fontaine V. Regulation et al. Clinical features and management of antrochoanal polyps in children:
of human papillomavirus type 16 early gene expression in trophoblastic cues from a clinical series of 58 patients. Int J Pediatr Otorhinolaryngol.
and cervical cells. Virology. 2011;412(1):146–55. 2018;114:87–91.
41. Pett MR, Herdman MT, Palmer RD, Yeo GSH, Shivji MK, Stanley MA, et al. 61. Burd EM. Human papillomavirus and cervical cancer. Clin Microbiol Rev.
Selection of cervical keratinocytes containing integrated HPV16 associates 2003;16(1):1–17.
with episome loss and an endogenous antiviral response. Proc Natl Acad 62. Ramakrishnan S, Partricia S, Mathan G. Overview of high-risk HPV’s 16 and
Sci. 2006;103(10):3822–7. 18 infected cervical cancer: pathogenesis to prevention. Biomed
42. Marcoux S, Le ONL, Langlois-Pelletier C, Laverdière C, Hatami A, Robaey P, Pharmacother Biomedecine Pharmacother. 2015;70:103–10.
et al. Expression of the senescence marker p16INK4a in skin biopsies of 63. Nicolás-Párraga S, Gandini C, Pimenoff VN, Alemany L, de Sanjosé S, Xavier
acute lymphoblastic leukemia survivors: a pilot study. Radiat Oncol Lond Bosch F, et al. HPV16 variants distribution in invasive cancers of the cervix,
Engl. 2013;8:252. vulva, vagina, penis, and anus. Cancer Med. 2016;5(10):2909–19.
43. Xiao Z, Liu Q, Mao F, Wu J, Lei T. TNF-α-induced VEGF and MMP-9 64. Geißler C, Tahtali A, Diensthuber M, Gassner D, Stöver T, Wagenblast J. The
expression promotes hemorrhagic transformation in pituitary adenomas. Int role of p16 expression as a predictive marker in HPV-positive oral SCCHN--a
J Mol Sci. 2011;12(6):4165–79. retrospective single-center study. Anticancer Res. 2013;33(3):913–6.Oton-Gonzalez et al. Infectious Agents and Cancer (2021) 16:3 Page 9 of 9
65. Castellsagué X, Alemany L, Quer M, Halec G, Quirós B, Tous S, et al. HPV
involvement in head and neck cancers: comprehensive assessment of
biomarkers in 3680 patients. J Natl Cancer Inst. 2016;108(6):djv403.
66. Shukla S, Mahata S, Shishodia G, Pande S, Verma G, Hedau S, et al. Physical
state & copy number of high risk human papillomavirus type 16 DNA in
progression of cervical cancer. Indian J Med Res. 2014;139(4):531–43.
67. Abramson AL, Nouri M, Mullooly V, Fisch G, Steinberg BM. Latent human
papillomavirus infection is comparable in the larynx and trachea. J Med
Virol. 2004;72(3):473–7.
68. Kalantari M, Garcia-Carranca A, Morales-Vazquez CD, Zuna R, Montiel DP,
Calleja-Macias IE, et al. Laser capture microdissection of cervical human
papillomavirus infections: Copy number of the virus in cancerous and
normal tissue and heterogeneous DNA methylation. Virology. 2009;390(2):
261–7.
69. Münger K, Baldwin A, Edwards KM, Hayakawa H, Nguyen CL, Owens M,
et al. Mechanisms of human papillomavirus-induced oncogenesis. J Virol.
2004;78(21):11451–60.
70. Huang J, Qian Z, Gong Y, Wang Y, Guan Y, Han Y, et al. Comprehensive
genomic variation profiling of cervical intraepithelial neoplasia and cervical
cancer identifies potential targets for cervical cancer early warning. J Med
Genet. 2019;56(3):186–94.
71. Thakur JS, Chaitanya A, Minhas RS, Azad RK, Sharma DR, Mohindroo NK.
Killian’s polyp mimicking malignant tumor. Ann Maxillofac Surg. 2015;5(2):
281–3.
72. Leonard SM, Pereira M, Roberts S, Cuschieri K, Nuovo G, Athavale R, et al.
Evidence of disrupted high-risk human papillomavirus DNA in
morphologically normal cervices of older women. Sci Rep. 2016;6:20847.
73. Romagosa C, Simonetti S, López-Vicente L, Mazo A, Lleonart ME, Castellvi J,
et al. p16(Ink4a) overexpression in cancer: a tumor suppressor gene
associated with senescence and high-grade tumors. Oncogene. 2011;30(18):
2087–97.
74. Hibi K, Koike M, Nakayama H, Fujitake S, Kasai Y, Ito K, et al. A cancer-prone
case with a background of methylation of p16 tumor suppressor gene. Clin
Cancer Res Off J Am Assoc Cancer Res. 2003;9(3):1053–6.
75. Wong DJ, Paulson TG, Prevo LJ, Galipeau PC, Longton G, Blount PL, et al.
p16(INK4a) lesions are common, early abnormalities that undergo clonal
expansion in Barrett’s metaplastic epithelium. Cancer Res. 2001;61(22):8284–
9.
76. Carestiato FN, Amaro-Filho SM, Moreira MAM, Cavalcanti SMB. Methylation
of p16 ink4a promoter is independent of human papillomavirus DNA
physical state: a comparison between cervical pre-neoplastic and neoplastic
samples. Mem Inst Oswaldo Cruz. 2018;114:e180456.
77. Prado JCM, Monezi TA, Amorim AT, Lino V, Paladino A, Boccardo E. Human
polyomaviruses and cancer: an overview. Clin Sao Paulo Braz. 2018;73(suppl
1):e558s.
78. Coelho TR, Gaspar R, Figueiredo P, Mendonça C, Lazo PA, Almeida L.
Human JC polyomavirus in normal colorectal mucosa, hyperplastic polyps,
sporadic adenomas, and adenocarcinomas in Portugal: JCV presence in
normal or abnormal colorectal mucosa. J Med Virol. 2013;85(12):2119–27.
79. Tognon M, Corallini A, Martini F, Negrini M, Barbanti-Brodano G. Oncogenic
transformation by BK virus and association with human tumors. Oncogene.
2003;22(33):5192–200.
80. Mazzoni E, Rotondo JC, Marracino L, Selvatici R, Bononi I, Torreggiani E,
et al. Detection of merkel cell polyomavirus DNA in serum samples of
healthy blood donors. Front Oncol. 2017;7:294.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in
published maps and institutional affiliations.You can also read

















































