Habitat preferences of the Eurasian Griffon Vulture (Gyps fulvus) in Bulgaria to support species management
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ornis Fennica 98: 116–127. 2021
Habitat preferences of the Eurasian Griffon Vulture
(Gyps fulvus) in Bulgaria to support species management
Dobromir D. Dobrev* & Georgi S. Popgeorgiev
D.D. Dobrev, Bulgarian Society for the Protection of Birds, 5 Leonardo da Vinci Str.,
PO box 562, 4000 Plovdiv, Bulgaria & Plovdiv University, Faculty of Biology,
24 Tzar Assen Str., Plovdiv 4000, Bulgaria * Corresponding author’s e-mail: dobromir.
dobrev1@gmail.com
G.S. Popgeorgiev, Bulgarian Society for the Protection of Birds, 5 Leonardo da Vin-
ci Str., PO box 562, 4000 Plovdiv, Bulgaria & National Museum of Natural History,
Bulgarian Academy of Sciences, 1 Tsar Osvoboditel Blvd., 1111 Sofia, Bulgaria
Received 16 December 2020, accepted 31 July 2021
The Eurasian Griffon Vulture (Gyps fulvus) is a large scavenger with a population
ranging between Portugal and India. The species is an obligate scavenger with a narrow
ecological niche and is therefore particularly dependent on, and limited by habitat
availability. The study aimed, for the first time in Eastern Europe, to identify the habitat
preferences of the Griffon Vulture at landscape and cliff scales. We used long-term
monitoring data between 1987–2018 to analyze habitat preferences of the natal Griffon
Vulture population in the Eastern Rhodope Mountains, Bulgaria. We employed single
explanatory variable tests to reveal the species habitat preferences at two spatial scales.
The results revealed Griffon Vultures’ high preferences towards rocky habitats at the
landscape level. At a cliff scale, the height and length of the cliff, the distance to the
nearest conspecific colony and the distance to the nearest feeding site were the best
predictors for the species habitat preferences. We stress the importance of these findings
considering the current status of the species within the region. Our results are useful to
support the future conservation of the Griffon Vulture population in Bulgaria and provide
a starting point for future research in the Balkans and Eastern Europe.
necessary because resources may vary (Martínez
1. Introduction et al. 2003). Therefore, a multi-scale approach
has been proposed and used in avian research to
Determining resources available to animals is identify different factors outlining the relationship
essential to preserve species and is mandatory to between the environment and the spatial distribu-
understand their requirements (Manly et al. 2002). tion of raptors and scavengers (García-Ripollés et
Resources are not uniform and hence the usage of al. 2005, López-López et al. 2006, 2007, Margalida
environmental features by animals through the et al. 2008, Mateo-Tomás & Olea 2010). These
process of habitat selection and/or preferences is relationships have been broadly examined in
important to study (Johnson 1980). Understanding the last decades (Guisan & Zimmermann 2000,
species distribution at different spatial scales is Guisan & Thuiller 2005, Olea & Mateo-Tomás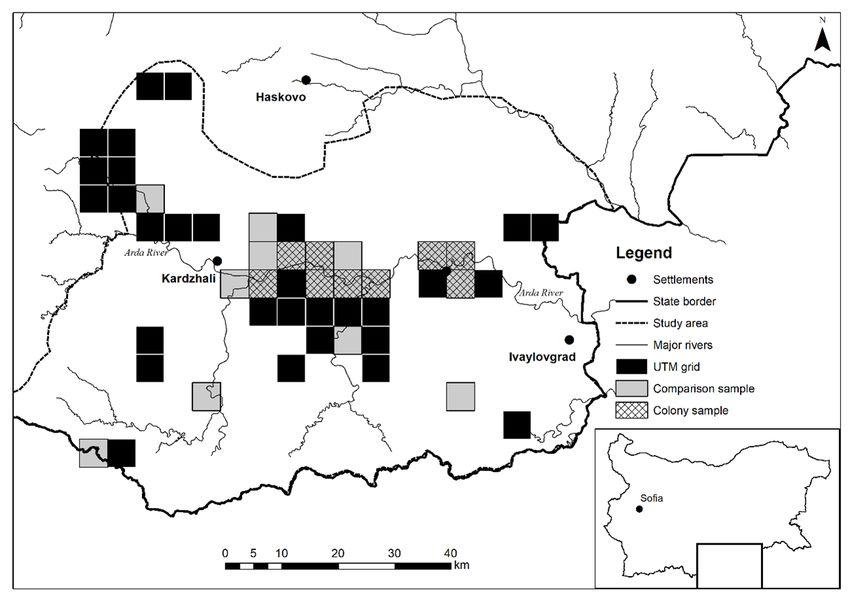
Dobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 117
2013). The main objectives of most of these species is also breeding in the Balkan Mountain
studies was to identify the habitat preferences range and Pirin Mountains as a result of a re-
and subsequently to address conservation actions introduction program in Bulgaria, and the national
(Martínez et al. 2003, Muñoz et al. 2005, Mateo- population now numbers 130 pairs distributed in
Tomás & Olea 2009, Olea & Mateo-Tomás 2011, five breeding nuclei (Demerdzhiev et al. 2014a,
Di Vittorio et al. 2012, Donázar et al. 2016). Dobrev et al. 2021b).
The Eurasian Griffon Vulture (Gyps fulvus) The relationship between the Griffon Vulture
(hereafter Griffon Vulture) is a large Old-World and its habitat requirements at different spatial
vulture with a distribution spanning from the scales in different areas of the Iberian Peninsula
Iberian Peninsula to the Himalayan region has been well-studied (Parra & Telleria 2004,
(Cramp & Simmons 1980). The species plays a García-Ripollés et al. 2005, Mateo-Tomás & Olea
key ecological role to recycle animal carcasses, 2010, Morales-Reyes et al. 2017). An extensive
limit the spread of diseases, and maintain the study conducted in Spain revealed the importance
nutrient return in natural ecosystems (DeVault of environmental factors for their distribution at
et al. 2016, Sebastián-González et al. 2020). two spatial scales (García-Ripollés et al. 2005).
Griffon Vulture is one of the most prominent At the colony level, the distance to the nearest
providers of multiple natural non-material cottage, the distance to the nearest colony, and
contributions to people thus also plays a major role the average altitude above sea level were the
in social-ecological systems (Aguilera-Alcalá et best predictors of Griffon Vulture preferences
al. 2020). The most abundant species population for a breeding site. They also found that climatic,
at a global scale is found in Spain (Botha et al. geomorphologic, disturbance-related, trophic
2017). The species is classified globally as ‘Least and vegetation variables were involved in the
Concern’ with an increasing population and range habitat preferences at the landscape level (García-
(Birdlife International 2017). Griffon Vulture Ripollés et al. 2005). The presence of vultures was
was widely distributed across the Balkans in the related to traditional livestock practices based on
past with many colonies known from all over the the movement of livestock between summer and
region (Andevski 2013). However, because of the winter pastures, e.g. transhumance; vultures relied
use of poison baits to extirpate large terrestrial on transhumant animals for up to four months per
predators, the species was brought to the brink of year in the Cantabrian Mountains (Olea & Mateo-
extinction from almost all of the area after World Tomás 2009). Rocky habitats and livestock
War II (Demerdzhiev et al. 2014a). Nowadays the abundance were the most important variables
species survives in scattered fragments across five affecting vulture distribution (Mateo-Tomás &
Balkan countries with a population estimate of Olea 2010).
445–565 pairs (Dobrev et al. 2021b). In contrast, in the Balkans, the distribution of
In Bulgaria, the Griffon Vulture population the Griffon Vulture in respect to habitat prefer-
showed a steep decline and by the beginning of the ences and selection is largely unexplored because
1970s was considered extinct from the country. such ecological surveys are limited in the region
At the end of that decade, a breeding colony was (Xirouchakis & Mylonas 2004, 2005). As excep-
discovered in the Eastern Rhodope Mountains tions, McIntyre (2010) modelled Griffon Vulture
(Iankov 2007, Demerdzhiev et al. 2014a). Since distribution relative to food availability in Crete,
the 1980s, this population was subject to regular and Polce (2004) and Dobrev & Popgeorgiev
and intensive monitoring of its colony occupancy, (2021) assessed the species habitat suitability
number of pairs and breeding rates. As a result in Romania and Bulgaria, respectively. Only
of numerous conservation activities, such as species distribution and population size have
supplementary feeding, anti-poisoning actions been relatively well documented. However, those
and active work to engage local communities in parameters are insufficient to ensure the conser-
the protection of the species, the Griffon Vulture vation of Griffon Vulture breeding and foraging
population slowly recovered in the last 30 years habitat, without being supplemented with studies
and reached over 100 pairs recently (Dobrev revealing the species ecological requirements.
& Stoychev 2013, Dobrev et al. 2019). The Therefore, habitat preference studies are urgently118 ORNIS FENNICA Vol.98, 2021
needed to explain species distribution, to evaluate 2. Materials and methods
Griffon Vultures habitat carrying capacity and
therefore to provide important information for the 2.1. Study area and data collection
management and conservation of the species in
the light of the current habitat changes and threats The study encompassed the territory of the
(Newton 1979, Dobrev & Stoychev 2013). This Eastern Rhodope Mountains, Bulgaria, part of
knowledge is essential for the future conservation the Rilo-Rhodopean massif (Fig. 1). This low
of the species in Bulgaria, but also in the Balkans mountainous area (100 to 1463 m a.s.l.) comprises
and Еastern Europe. diverse heterogeneous habitats and low-impact
The current study aims to identify Griffon human activities, mainly extensive livestock
Vulture habitat preferences and the specific envi- breeding. Over 150,000 cattle, sheep and goats are
ronmental variables determining its distribution raised in the territory (BFSA 2018) with the most
at two spatial scales: landscape and cliff, in the free grazing for much of the year. The climate is
Eastern Rhodopes. We analysed species prefer- continental-Mediterranean characterized with
ences and discuss our findings broadly to ensure average January temperatures of 1–2°C and hot
a better understanding of the Griffon Vulture and dry summers, with average July temperatures
resource selection and to support the conservation of 24°C. The annual rainfall is 600–800 mm
management of the species in Bulgaria and other (Kopralev et al. 2002). The area hosts the highest
areas of its range. diversity of breeding raptors in Bulgaria (23
species) and 171 breeding bird species overall
(Stoychev et al. 2004).
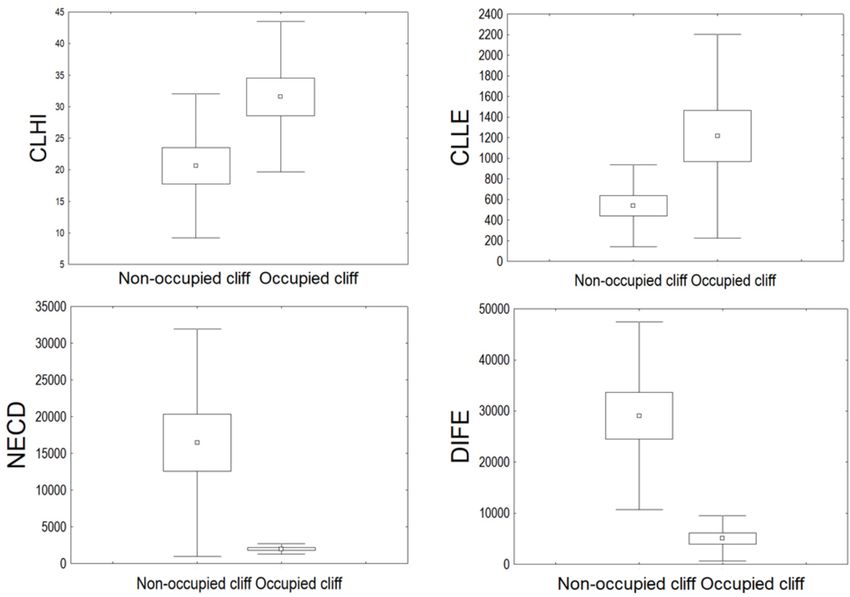
Fig. 1. Map showing the study area and 5 × 5 km cells on the UTM grid used for comparisons between occupied and
non-occupied cliffs analysed at two spatial scales.Dobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 119
The data set we used for both cliff and The same number of non-occupied grid cells
landscape preferences of the species was based on (n = 9, comparison sample) was selected at
long-term breeding distribution surveys conducted random from the same grid and compared with
during 1987–2018 (Demerdzhiev et al. 2014a, each other (analysed sample = 18 cells in total,
Dobrev et al. 2019). The survey was carried out see Fig.1). Also, colony, climate, disturbance and
mainly along the Arda River and its tributaries trophic parameters of the known breeding cliffs
to locate all occupied breeding cliffs by Griffon (n = 16) were compared at this spatial scale with
Vultures. However, more distant cliffs that offered the same number of randomly selected non-occu-
suitable nesting sites were also annually checked pied cliffs within the analyzed grid cells (García-
within the study area to locate other breeding Ripollés et al. 2005). Non-occupied cliffs were
locations (Demerdzhiev et al. 2014a). We selected randomly from a pool of cliffs (n = 26)
monitored the area yearly in January/August with encompassed within the analysed sample. Thus,
at least four visits to each potential breeding cliff occupied and non-occupied cells and cliffs were
between 1987 and 2018. We used the method for independently sampled to collect information
observation from an elevated viewpoint (Bibby et on the colony, climate, disturbance, topography,
al. 1999) and recorded the status of the different trophic and habitat variables. The spatial calcula-
Griffon Vulture colonies, following López-López tions and measurements of these variables for each
et al. (2004). Thereby, species distribution in the of the studied cliffs and cells were processed in
study area was determined to adequately qualify QGIS (QGIS Development Team 2016). Colony
presences and absences. and disturbance related variables were measured
based on the ortho-photography layer of Bulgaria
(scale 1:2000) (http://gis.mrrb.government.bg/)
2.2 Cliff (colony) and landscape (home range) and obtained from field surveys and NSI (2012),
preferences respectively. Climate-related variables were
obtained from free internet climate databases
All identified Griffon Vulture breeding localities at (rp5.ru, weatheronline.co.uk, stringmeteo.com).
the cliff and landscape-scale were geo-referenced Topography features were obtained from a Digital
using GPS and an ortho-photography layer of Elevation Model (DEM) of Bulgaria with a 40 m
Bulgaria. Those data were processed in Quantum pixel size. Trophic features were obtained from
GIS (QGIS) to match the study design. Thus, the the country’s official database, obtained by BFSA
study area was divided into a grid comprising 48 (2018), and habitat variables were based on 2012
cells of the UTM 5 × 5 km grid using the MGRS Corine Land Cover layer classification (but see
naming of cells (UTM zone 35N, datum WGS Table 1) (Bossard et al. 2000). These variables
1984), containing rocky landscapes and cliffs were selected because they are indirectly related
(Fig. 1), following a stratified random design to known breeding habitat preferences of the
(Sutherland 2000, Xirouchakis & Mylonas 2004, Griffon Vulture and were expected to affect the
Di Vittorio et al. 2015). The Griffon Vulture has a realized ecological niche of the species (Guisan
large home range (Monsarrat et al. 2013, Genero & Zimmermann 2000, López-López et al. 2007).
et al. 2020, Martin-Díaz et al. 2020). However, the Thus, our results would be comparable to similar
Griffon Vulture breeding distribution in our area studies in Spain (García-Ripollés et al. 2005) and
is confined along the Arda river valley between would stem from a standard procedure, useful for
Kardzhali and Madzharovo (Demerdzhiev et al. future assessment in Eastern Europe.
2014а). Therefore, in our study, we used a 5 × 5
km grid cell to measure species preferences at
landscape scale according to the concise distribu- 2.3 Statistical procedures
tion of the Griffon Vulture, studied by GPS/GSM
transmitters in the Eastern Rhodopes (Arkumarev Before starting the statistical analysis, we processed
et al. 2021, BSPB/Birdlife Bulgaria unpubl.). the data for multi-collinearity. We used the
Breeding colonies were identified in nine of Spearman correlation coefficient (r) to explore the
the 48 UTM 5 × 5 km grid cells (colony sample). inter-correlations among the continuous variables120 ORNIS FENNICA Vol.98, 2021
Table 1. Complete list of environmental variables considered in the Griffon Vulture habitat preferences comparison.
Variables included in the cliffs analysis are marked with (1) and those included in the landscape analysis with (2).
Dropped
because of
Abbreviation Variable description (measurement unit)
correlation
(r > 0.7)
Colony
CLLE1 Linear length of the cliff (m) No
CLHI1 Height of the cliff, measured at the highest point (m). No
The linear distance between the base and the top of the cliff.
CLHIma1 Highest (summit) level of the breeding cliff (m) Yes
NECD 1 Linear distance to the nearest neighbouring colony (m) No
CAMA 1 Aspect of the breeding cliff (degrees). The variable was categorized No
into 16 categories (per every 22.5°).
Climate
APJA1,2 Average rainfall during January (mm) Yes
Disturbance
SEDI1 Linear distance to the nearest human settlement (m) No
RODI 1 Linear distance to the nearest paved road (m) No
INDE2 Inhabitant density in the surveyed grid cell (people/km2) No
POLE2 Length of the low and medium voltage grid system ≤ 110 kV (m) Yes
Topography
ASAV2 Average aspect of the cell (degrees), created by a triangular irregular No
network (TIN) method based on vector data of contour lines with 40
m accuracy as the maximum rate of change in exposure across each
in the TIN. For the aim of the analysis, the variable was categorized
into 16 categories (per every 22.5°).
Trophic
DELI2 Density of the livestock in livestock units (LU): (1) 1 cow = 5 LU; (2) 1 No
sheep/goat = 1 LU; (3) pig = 2 LU. A method to quantify the biomass
of the available livestock in the study area, based on Mateo-Tomás &
Olea 2010.
DIFE1 Linear distance to the nearest feeding station (m) No
Habitat
PEOH2 Area of the open habitats in the cell in hectares (Categories from No
the Corine Land Cover describing the open habitat types: code
321 – Natural grasslands; 324 – Transitional woodland-shrub; 231
– Pastures; 243 – Land principally occupied by agriculture, with
significant areas of natural vegetation; 112 – Discontinuous urban
fabric; 333 – Sparsely vegetated areas; 131 – Mineral extraction
sites; 142 – Open sports and leisure areas).
PERO2 Area of rock habitats in the cell in hectares NoDobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 121
(Table 1). From pairs of variables that showed length (CLLE), cliff height (CLHI), nearest neigh-
high correlation (r > 0.7), we used only one to be bouring Griffon Vulture colony distance (NECD)
analyzed further on. The decision on which variable and the distance to the nearest feeding station
to retain was based on the results of a Principal (DIFE). Cliffs occupied by Griffon Vultures were
Component Analysis (PCA), selecting the one longer (U = 2.13, p = 0.03) and higher (U = 1.73,
with a higher factor score. As a result, we removed p = 0.05) in comparison to non-occupied ones.
three variables (CLHIma – highest (summit) level The simple Mantel test showed that occupied
of the breeding cliff; APJA – Average rainfall and non-occupied cliffs have no similarities in
during January; and POLE – Length of the low their spatial pattern of distribution (normalized Z
and medium voltage grid system ≤ 110 kV) and = 29.27, p = 0.85), neither there is a correlation
retained 6 variables for the cliff analysis (CLLE – between cliffs distance matrices (r = 0.35, p =
cliff length; CLHI – cliff height; NECD – linear 0.12). Nevertheless, the NECD of occupied cliffs
distance to the nearest neighbouring colony; SEDI is significantly lesser in comparison to that of
– linear distance to the nearest human settlement; non-occupied cliffs (U = –3.31, p = 0.00) (Fig.
RODI – linear distance to the nearest paved road; 2). The distance to the nearest feeding station was
and DIFE – linear distance to the nearest feeding significantly lower in occupied than in non-
station) and 4 variables for the cell analysis occupied cliffs (U = –4.52, p < 0.001). Cliffs
(INDE – inhabitants density in the surveyed grid occupied by the Griffon Vulture had predomi-
cell; PEOH – an area of the open habitats in the nantly southern exposure (CAMA) in contrast to
cell; PERO – an area of rock habitats in the cell; randomly selected cliffs (χ2 = 12.4, p = 0.05).
and DELI – density of the livestock in livestock
units) from a set of a priori selection (Table 1).
Non-parametric tests were applied because data 3.2. Habitat preferences at landscape scale
did not approach the normal distribution (Fowler
& Cohen 1995). We compared occupied and Cells occupied by Griffon Vultures colonies had a
non-occupied cliffs and grid cells through Mann- significantly more area of rocky habitats (PERO)
Whitney U-test to search for differences in the than non-occupied ones (U = 2.78, p = 0.003). We
medians of the explored variables separately. We did not find significant differences between the
further compared the categorical variables (ASAV aspect (ASAV) of the occupied and non-occupied
– aspect of the cell at home range scale and CAMA cells (χ2 = 27.46, p = 0.3). Density of human
– aspect of the breeding cliff at colony scale) using population (INDE) (U = 1.19, p = 0.25), the area
the Chi-Square test (χ2). We separately computed of open habitats (PEOH) (U = 1.02, p = 0.34) and
a simple Mantel test (Z) with 30 000 permutations livestock densities (DELI) (U = 0.39, p = 0.73) did
to assess the similarity and correlation between not differ significantly between the occupied and
the nearest neighbouring conspecifics distances of non-occupied cells.
the occupied and non-occupied cliffs and species
presence/absence dissimilarity matrices (Pierre-
Rossi 1996, Schick et al. 2004). Results with 4. Discussion
p < 0.05 were considered significant. Statistica
for Windows, Release 7 (StatSoft Inc. 2004) and This is the first study aimed at providing prime
R v.3.6.1 (R Development Core Team 2012) were knowledge for the most important environmental
used for the statistical analysis of the data variables affecting the Griffon Vulture’s prefer-
ences at two spatial scales in Bulgaria and, to a
broader extent, in Eastern Europe. The availability
3. Results of suitable breeding sites is a substantial driver of
site occupancy (Newton 1979, Mateo-Tomás &
3.1. Habitat preferences at cliff scale Olea 2011, Di Vittorio & López-López 2014). We
demonstrated that variables related to the breeding
Statistically significant differences (p < 0.05) cliffs have high significance for the habitat prefer-
were found between the following variables: cliff ences of the Griffon Vulture. Cliff height, length122 ORNIS FENNICA Vol.98, 2021 Fig. 2. Box-Whiskers plots presenting the comparison of the variables showing statistically significant differences between occupied and non-occupied cliffs: CLLE – cliff length (m); CLHI – cliff height (m); NECD – Linear distance to the nearest neighbouring colony (m); and DIFE – Linear distance to the nearest feeding station (m) (± standard error presented with , standard deviation presented with and mean, presented with ). and distance to the nearest conspecific colony were Nesting substrate (niche or ledge) may affect highlighted as important factors for occupancy by species breeding (López-López et al. 2004) and other studies as well (Cramp & Simmons 1980, occupancy (Iezekiel et al. 2004). A previous study Xirouchakis & Mylonas 2005). Similarly, the in our study area did not find such a relationship best predictors affecting the distribution of large (Demerdzhiev et al. 2014a). However, the nesting birds of prey such as the Golden Eagle (Aquila substrate may positively affect Griffon Vulture chrysaetos) and the Bearded Vulture (Gypaetus breeding success and ensure protection from barbatus) were related to topographic features adverse weather conditions (Ceballos & Donázar (López-López et al. 2007, Margalida et al. 2008, 1988, Iezekiel et al. 2004, Freund et al. 2017). Di Vittorio & López-López 2014). The Griffon Our results also underline the significance Vulture, being a large cliff-nesting raptor is highly of the nearest colony distance as a major factor selective to topographic features too, as highlight- affecting species preferences and confirm ed by our and others’ results (Parra & Telleria findings from previous studies (Xirouchakis & 2004, Xirouchakis & Mylonas 2005). Larger cliffs Mylonas 2004, Margalida et al. 2007, Skartsi present more suitable sites for breeding (Mateo- et al. 2009). The positive trend of any given Tomás & Olea 2011). Hence, the Griffon Vultures’ population is dependent on the surplus of birds cliffs in the study area usually present considera- immigrating from other populations (Demerdzhiev ble space, height, and length situated along rivers, et al. 2014b). The occupancy of new territories steep slopes, and gorges where birds can easily is highly dependent and related to neighbouring take off and land (Xirouchakis & Mylonas 2005). conspecifics, occupying the high-quality habitats
Dobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 123
(Carrete et al. 2007). This has been observed in whole range of the species (McIntyre 2010).
our study area where the newly emerging colonies This highlights the strong connection between
were always close to the core ones (Demerdzhiev Griffon Vulture and livestock and supports our
et al. 2014a). Thus, occupancy of new territories statement that food availability and abundance
and the expansion of local range by the Griffon dependent from the extensive livestock husbandry
Vulture is strongly dependent on the safe perma- are of key importance for the species presence and
nence and breeding of neighbour populations. distribution (Xirouchakis & Mylonas 2004, Olea
We revealed that Griffon Vulture has higher & Mateo-Tomás 2009, Mateo-Tomás & Olea
preferences towards cliffs with southern exposure. 2015, Martin- Díaz et al. 2020).
This may be related to climatic factors in our study We found a significant positive relationship for
area, which we accounted for in our analysis but Griffon Vultures breeding colonies to be situated
did not show enough statistical power to be studied closer to feeding stations. This finding supports
in detail. However, Griffon Vultures breeding research in the same study area which reported
season starts with the egg-laying in January that Griffon Vultures have high seasonal prefer-
(Ferguson-Lees et al. 2001) and thus vultures are ences towards usage of the feeding sites without
particularly dependent on the climatic conditions being entirely reliant on them (Arkumarev et al.
at that time (Newton 1979, Grande et al. 2009). 2021) in contrast to other areas of the species
Sudden temperature changes and extreme precip- range (Harel et al. 2017). Indeed, the settlement
itation can negatively affect incubation and lead of supplementary feeding stations has become
to breeding failure (Donázar 1993, Elliott et al. a major conservation and management tool for
2005). In the Eastern Rhodopes, regardless of the vulture species in Europe (Cortes-Avizanda &
low altitudes, such climatic extremes are not a rare Pereira 2016, Cortes-Avizanda et al. 2016) and
event at the beginning of the breeding season and elsewhere (Brink et al. 2020). In Bulgaria, this
can ultimately alter reproduction causing failures management tool has been used exclusively to
and/or clutch replacement. Our findings support direct the Griffon Vulture population conservation
similar studies, where the Griffon Vulture avoids after its re-discovery in Bulgaria at the end of the
northern exposure to counteract the fluctuations 1970s (Dobrev& Stoychev 2013). The network
in the temperatures, precipitation (Demerdzhiev of feeding sites in Bulgaria is an important
et al. 2014a) and wind (Xirouchakis & Mylonas conservation tool for the Griffon Vulture popula-
2005). tion in the Balkans (Stoynov et al. 2017). Besides,
We did not find a difference between occupied our study confirms previous findings where the
and non-occupied cells about livestock density, proximity to the feeding site was determined as
very likely because the density of livestock in our one of the best predictors for Griffon Vultures
area is high in most of the territory and thus such roosting cliff selection in the pre-breeding season
differences would be difficult to establish on this (Dobrev et al. 2021a). However, the spatial and
scale. Nevertheless, livestock abundance shapes temporal predictability of the feeding sites can
the foraging behaviour of Griffon Vulture in our rapidly change vultures’ foraging strategies and
study area (Arkumarev et al. 2021). The species behavioural responses (Deygout et al. 2009,
is an obligate scavenger, searching food at great Moreno-Opo et al. 2010, 2015, Margalida et al.
distances and is highly dependent on the availabil- 2014). Consequently, the management of the
ity of dead animals and adapted to use carrion as a feeding sites must be species and/or area-specific
food source (DeVault et al. 2016, Martin-Díaz et al. and considered gently with respect to species
2020). Therefore, vultures are reliant on livestock conservation (Margalida et al. 2014).
abundance and are highly related to agro-pastoral Our study in the Eastern Rhodopes comple-
ecosystems (Xirouchakis & Mylonas 2004, Olea ments other studies from Europe for the factors
& Mateo-Tomás 2009, Donázar et al. 2009), affecting the Griffon Vulture habitat preferences
including in our area (Hristov 1997, Angelov et at two spatial scales – landscape and cliff. We
al. 2006, Arkumarev et al. 2021). For example, confirmed also for an eastern European population
the Griffon Vulture on Crete is probably one of that a combination of larger cliffs, the presence of
the most pastoralism-bonded populations in the conspecifics and suitable breeding and foraging124 ORNIS FENNICA Vol.98, 2021
habitats explain Griffon Vultures preferences. maisematasolla Itä-Euroopassa. Käytimme
Territory quality is an important measure of pitkäaikaisaineistoa (1987–2018) Bulgariasta
occupancy and fitness (Fahrig 2001, Sergio & Rodopivuorilta. Havaitsimme, että maisema-
Newton 2003). Hence, we recommend conserva- tasolla hanhikorppikotkat suosivat kivikkoisia
tion of the species’ prime breeding habitats, but habitaatteja. Kalliojyrkänteiden korkeus ja pituus,
also territories part of the historical breeding range etäisyys lähimpään hanhikorppikotkakoloniaan
in Bulgaria (Olea & Mateo-Tomás 2009, Dobrev sekä etäisyys lähimpään ruokintapaikkaan selit-
& Popgeorgiev 2021) that can be used either for tivät parhaiten lajin esiintymistä. Tulokset ovat
roosting (Dobrev et al. 2021a), but also re-occu- tärkeitä lajin nykyisen statuksen ymmärtämisen
pied in future colonisation (Dobrev et al. 2021b). kannalta, auttavat suojelutoimia suunniteltaessa,
Areas of extensive livestock breeding, like the ja toimivat lähtökohtana tuleville tutkimuksille
Eastern Rhodopes, must be sustained viable to Itä-Euroopassa.
contribute to vultures’ persistence (Mateo-Tomás
& Olea 2011, Martin- Díaz et al. 2020). The animal Acknowledgements. The current research was financed by
husbandry there might be favoured by application National program “Young scientists and Postdoctoral can-
of relevant EU agri-environmental measures to didates” funded by the Bulgarian Ministry of Education
support livestock breeders and search regional and Science. We express our gratitude to Hristo Hristov,
measures to buffer human-wildlife conflict and Dimitar Demerdzhiev, Marin Kurtev, Vladimir Dobrev,
the use of poisons. Thеreby, vultures will benefit Volen Arkumarev, Ivaylo Angelov, Nedko Nedyalkov,
from the availability of safe food and will be Vladimir Trifonov, Stoycho Stoychev, Anton Stamenov,
possibly less affected and/or exposed to severe Atanas Delchev, Vera Dyulgerska and Nikolay Terziev
threats like poisoning. Furthermore, the protection for their contribution in collecting monitoring data and
of the currently occupied breeding cliffs and support. We are grateful to Yurii Korniliev for providing
foraging habitats will ensure population growth valuable comments on the edited draft of the manuscript.
in future and its connection to neighbouring
breeding population nuclei of the species (Dobrev
& Popgeorgiev 2021, Dobrev et al. 2021b). We References
suggest similar studies to be conducted elsewhere
in the Balkans and Eastern Europe to bring insight Aguilera-Alcalá, N., Morales-Reyes, Z., Martín-López, B.,
into the species habitat selection process and Moleón, M. & Sánchez-Zapata, J. 2020: Role of scaven-
gers in providing non-material contributions to people.
preferences there. Application of cognate conser-
— Ecological Indicators 117: 106643.
vation approaches in neighbouring countries may Andevski, J. (ed) 2013: Vulture Conservation in the Balkan
bring homogeneity and consistency at a regional Peninsula and Adjacent Regions. 10 Years of Vulture
level and possibly ease the Griffon Vulture popu- Research and Conservation. Action Plan for the Recov-
lation recovery. The usage of GPS telemetry must ery and Conservation of Vultures on the Balkan Penin-
be considered to assist this process and reveal how sula and Adjacent Regions. — Vulture Conservation
this reverberates the survival and fitness of the Foundation & Frankfurt Zoological Society, Skopje.
Angelov, I., Demerdzhiev, D. & Stoychev, S. 2006: Use of
Griffon Vulture.
carcasses from wolf kills by Griffon Vultures (Gyps ful-
vus) in Eastern Rhodopes, Bulgaria. – In Proceedings of
the International Conference on Conservation and Man-
Hanhikorppikotkan elinympäristövaatimukset agement of Vulture Populations (eds. Houston, D.C. &
Bulgariassa – tukena suojelutoimille S.E. Piper). — Natural History Museum of Crete &
WWF Greece, Thessaloniki.
Hanhikorppikotka on kookas haaskalintu, Arkumarev, V., Dobrev, D., Stamenov, A., Terziev, N.,
Delchev, A. & Stoychev, S. 2021: Seasonal dynamics in
jonka levinneisyys ulottuu Portugalista Intiaan. the exploitation of natural carcasses and supplementary
Lajilla on kapea ekolokero, sillä se käyttää ravin- feeding stations by a top avian scavenger. — Journal of
nokseen vain haaskoja, ja täten sen esiintyminen Ornithology. DOI 10.1007/s10336-021-01865-1.
on hyvin rajattu tiettyihin elinympäristöihin. BFSA 2018: Reference at Art.37m, par.3 of the Property and
Tässä tutkimuksessa selvitettiin ensimmäistä ker- agricultural lands use Act. Downloaded from http://www.
taa hanhikorppikotkan elinympäristövaatimuksia babh.government.bg/en/ on 24/1/2018. (In Bulgarian)Dobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 125
Bibby, C., Jones, M. & Marsden, S. 1999: Expedition Field ture scavenging service efficiency. — Ecological Mod-
Techniques. – Bird Surveys-Royal Geographical Socie- elling 220: 1826–1835.
ty, London. Di Vittorio, M. & López-López, P. 2014: Spatial distribution
BirdLife International 2017: Gyps fulvus (Griffon Vulture). and breeding performance of Golden Eagles (Aquila
– IUCN Red List for birds. Downloaded from http:// chrysaetos) in Sicily: implications for conservation. —
www.birdlife.org/ on 01/01/2020. Acta Ornithologica 49: 33–45.
Bossard, M., Feranec, J. & Otahel, J. 2000: CORINE Land Di Vittorio, M., Ciaccio, A., Grenci, S. & Luiselli, L. 2015:
Cover Technical Guide-Addendum 2000. Technical re- Ecological modelling of the distribution of the Lanner
port No 40. Copenhagen (EEA). Downloaded from falcon Falco biarmicus feldeggii in Sicily at two spatial
http://terrestrial.eionet.eu.int. scales. — Ardeola 62: 81–94.
Botha, A.J., Andevski, J., Bowden, C.G.R., Gudka, M., Saf- Di Vittorio, M., López-López, P. & Sarà, M. 2012: Habitat
ford, R. J., Tavares, J. & Williams, N. P. 2017: CMS preference of Bonelli’s eagle (Aquila fasciata) in Sicily.
Multi – species Action Plan to conserve African – Eura- — Bird Study 59: 207–217.
sian Vultures. Abu Dhabi, United Arab Emirates: Coor- Dobrev, A., Arkumarev, V., Dobrev, V., Stamenov, A. & De-
dinating Unit of the CMS Raptors MOU. (CMS Raptors merdzhiev, D. 2021a: Use and selection of roost sites by
MOU Technical Publication No. 5. CMS Technical Se- Eurasian Griffon Vultures Gyps fulvus in Bulgaria. —
ries No. 35). Bird Study. DOI: https://doi.org/10.1080/00063657.202
Brink, C., Santangeli, A., Amar, A., Wolter, K., Tate, G., 1.1950122
Krueger, S., Tucker, A. S. & Thomson, R. L. 2020: Dobrev, D., Arkumarev, V., Skartsi, T., Stamenov, A. & Kret,
Quantifying the spatial distribution and trends of sup- E. 2019: Griffon Vulture population trend and size in the
plementary feeding sites in South Africa and their po- Eastern Rhodopes at the crossroads to Asia. — Poster,
tential contribution to vulture energetic requirements. European vulture conference, Algarve, Portugal.
— Animal Conservation 23: 491–501. Dobrev, D. & Popgeorgiev, G. 2021b: Modeling breeding
Carrete, M., Grande, J., Tella, J., Sanchez-Zapata, Donázar habitat suitability of Griffon Vultures (Gyps fulvus) in
J., Dıaz-Delgado, R. & Romo, A. 2007: Habitat, human Bulgaria and conservation planning. — North-Western
pressure, and social behavior: Partialling out factors af- Journal of Zoology 17(2).
fecting large-scale territory extinction in an endangered Dobrev, D. & Stoychev, S. 2013: Vulture conservation in
vulture. — Biological conservation 136: 143 – 154. Bulgaria. — Proceedings of the Griffon Vulture Confer-
Ceballos, O. & Donázar, J. 1988: Egyptian vulture (Neo- ence, 6-8 March 2013, Limassol.
phron percnopterus) nest-site in Navarra. — Munibe Dobrev, D., Tsiakiris, R., Skartsi, T., Dobrev, V., Arkumarev,
40: 3–8. V., Stara, K., Stamenov, A., Probonas, N., Kominos, T.,
Cortes-Avizanda, A. & Miguel Pereira, H. 2016: Rewilding: Galanaki, A., Kret, E., Hallmann, B., Grubač, B., Sušić,
Vulture restaurants cheat ecosystems. — Nature 540: G., Marinković, S., Hribšek, I., Skorić, S., Jerrentrup,
525–525. H., Lucić, V., Kapelj, S., Stoyanov, G., Zakkak, S., Hris-
Cortes-Avizanda, A., Blanco, G., DeVault, T., Markandya, tov, H., Stoychev, S., Sidiropoulos, L., Bino, T. & De-
A., Virani, M., Brandt, J. & Donázar, J. 2016: Supple- merdzhiev, D. 2021: Long-term size and range changes
mentary feeding and endangered avian scavengers: of the Griffon Vulture Gyps fulvus population in the
Benefits, caveats, and controversies. — Frontiers in Balkans: a review. — Bird Conservation International.
Ecology and the Environment 14: 191–199. DOI: https://doi.org/10.1017/S0959270921000198
Cramp, S. & Simmons, K. 1980: The Birds of the Western Donázar, J.A. 1993: Los buitres ibéricos. Biología y con-
Palearctic. — Oxford Univ. Press, Oxford. servación. — Reyero J. M., Madrid.
Demerdzhiev, D., Hristov, H., Dobrev, D., Angelov, I. & Donázar, J.A., Avizanda, A., Fargallo, J., Margalida, A.,
Kurtev, M. 2014а: Long-term population status, breed- Moleon, M., Morales-Reyes, Z., Moreno-Opo, R.,
ing parameters and limiting factors of the Griffon Vul- Perez-Garzia, J., Sanchez-Zapata, J., Zuberogoitia, I. &
ture (Gyps fulvus) population in Eastern Rhodopes, Bul- Serrano, D. 2016: Roles of raptors in a changing world:
garia. — Acta Zoologica Bulgarica 66: 373–384. from flagships to providers of key ecosystem services.
Demerdzhiev, D., Stoychev, S., Dobrev, D., Spasov S. & — Ardeola 63: 181–234.
Oppel, S. 2014b: Studying the demographic drivers of Donázar, J., Margalida, A. & Campión, D. 2009: Vultures,
an increasing Imperial Eagle population to inform con- Feeding Stations and Sanitary Legislation: A conflict
servation management. — Biodiversity and Conserva- and its consequences from the perspective of conserva-
tion 24: 627–639. tion biology. — Sociedad de Ciencias Aranzadi San
DeVault, T., Beasley, J., Olson, Z., Moleón, M., Carrete, M., Sebastián, Spain.
Margalida, A. & Sanchez-Zapata, J.A. 2016: Ecosystem Elliott, K., Gill, C. & Elliott, J. 2005: The influence of tide
Services Provided by Avian Scavengers. — USDA Na- and weather on provisioning rates of chick-rearing Bald
tional Wildlife Research Center-Staff Publications. Eagles in Vancouver island, British Columbia. — Jour-
Deygout, C., Gault, A., Sarrazin, F. & Bessa-Gomes, C. nal of Raptor Research 39: 1–10.
2009: Modeling the impact of feeding stations on vul- Fahrig, L. 2001: How much habitat is enough? — Biological126 ORNIS FENNICA Vol.98, 2021
Conservation 100: 65–74. use at different spatial scales. — Journal of Ornithology
Ferguson-Lees, J., Christie, D.A., Franklin, K., Mead, D. & 147: 97–106.
Burton, P. 2001: Raptors of the World. — Houghton López-López, P., Ripollez, C.G., Soutullo, A., Cadahia, L. &
Mifflin Company, New York. Urios, V. 2007: Identifying potentially suitable nesting
Fowler, J. & Cohen, L. 1995: Statistics for Ornithologists: habitat for golden eagles applied to ‘important bird are-
2nd ed. — BTO, London. as’ design. — Animal Conservation 10: 208–218.
Freund, M., Bahat, O. & Motro, U. 2017: Breeding success López-López, P., García-Ripollés, C. & Verdejo, J. 2004:
and its correlation with nestsite characteristics: a study Population status and reproductive performance of Eur-
of a Griffon Vulture colony in Gamla, Israel. — Journal asian Griffons (Gyps fulvus) in eastern Spain. — Journal
of Raptor Research 51:136–144. of Raptor Research 38: 350–356.
García-Ripollés, C., López-López, P., García-López, F., Manly, B., McDonald, L., Thomas, D.,McDonald, T., & Er-
Aguilar, J.M. & Verdejo, J. 2005: Modeling nesting hab- ickson, W. 2002: Resource selection by animals. Statis-
itat preferences of Eurasian Griffon Vulture Gyps fulvus tical design and analysis for field studies, 2nd edition.
in eastern Iberian Peninsula. — Ardeola 52: 287–304. — Dordrecht, The Netherlands:Kluwer Academic Pub-
Genero, F., Franchini, M., Fanin, Y., Filacorda, S. 2020: Spa- lisher.
tial ecology of non-breeding Eurasian Griffon Vultures Margalida, A., Campion, D. & Donázar, J. 2014: Vultures vs
(Gyps fulvus) in relation to natural and artificial food livestock: Conservation relationships in an emerging
availability. — Bird Study 67: 53–70. conflict between humans and wildlife. — Oryx 48:
Grande, J.M., Serrano, D., Tavecchia, G., Carrete, M., Ce- 172–176.
ballos, O., Diaz-Delgado, R., Tella, J. & Donázar, J. Margalida, A., Donázar, J., Bustamante, J., Hernández, F. &
2009: Survival in a long-lived territorial migrant: Ef- Romero-Pujante, M. 2008: Application of a predictive
fects of life-history traits and ecological conditions in model to detect long-term changes in nest-site selection
wintering and breeding areas. — Oikos 118: 580–590. in the Bearded Vulture (Gypaetus barbatus): conserva-
Guisan, A. & Thuiller, W. 2005: Predicting species distribu- tion in relation to territory shrinkage. — Ibis 150: 242–
tion: offering more than simple habitat models. — Ecol- 249.
ogy Letters 8: 993–1009. Margalida, A., García, D. & Cortés-Avizanda, A. 2007: Fac-
Guisan, A. & Zimmermann, N.E. 2000: Predictive habitat tors influencing breeding density of Bearded Vultures,
distribution models in ecology. — Ecological Model- Egyptian Vultures and Eurasian Griffon Vultures in Cat-
ling 135: 147–186. alonia (NE Spain): management implications. — Ani-
Harel, R., Spiegel, O., Getz, W.M. & Nathan, R. 2017: So- mal Biodiversity and Conservation 30: 189–200.
cial foraging and individual consistency in following Martin-Díaz, P., Cortés-Avizanda, A.,Serrano, D., Arrondo,
behaviour: testing the information Centre hypothesis in E., Sánchez-Zapata, J. & Donázar, J. 2020: Rewilding
free-ranging vultures. — Proceedings of the Royal So- processes shape the use of Mediterranean landscapes by
ciety B: Biological Sciences 284: 10.1098/ an avian top scavenger. — Scientific Reports 10
rspb.2016.2654. (1):2853.
Hristov, H. 1997: Status, monitoring and supplementation of Martínez, J.A., Serrano, D. & Zuberogoitia, I. 2003: Predic-
the vultures in the Eastern Rhodopes. — Eastern Rho- tive models of habitat preferences for the Eurasian Ea-
dopes reports, BSBDP-BSPB, Sofa. (In Bulgarian) gle Owl Bubo bubo: a multiscale approach. — Ecogra-
Iankov, P. (ed.) 2007: Atlas of Breeding Birds in Bulgaria. phy 26: 21–28.
— Bulgarian Society for the Protection of Birds, Con- Mateo-Tomás, P. & Olea, P. P. 2009: Combining scales in
servation series, Book 10, Sofia. habitat models to improve conservation planning in an
Iezekiel, S., Bakaloudis, D.E. & Vlachos, C.G. 2004: The endangered vulture. — Acta Oecologica 35: 489–498.
status and conservation of Griffon Vulture Gyps fulvus Mateo-Tomás, P. & Olea, P.P. 2010: Anticipating Knowl-
in Cyprus. — In Raptors worldwide: proceedings of the edge to Inform Species Management: Predicting Spa-
6th World Conference on Birds of Prey and Owls (eds. tially Explicit Habitat Suitability of a Colonial Vulture
Chancellor, R.D. & B.-U. Meyburg). – WWGBP/ Spreading Its Range. — PLoS ONE 5: e12374.
MME, Budapest, Hungary. Mateo-Tomás, P. & Olea, P.P. 2011: The importance of so-
Johnson, D. 1980: The comparison of usage and availability cial information in breeding site selection increases with
measurements for evaluating resource preference. — population size in the Eurasian Griffon Vulture (Gyps
Ecology 61: 65–71. fulvus). — Ibis 153: 832–845.
Kopralev, I. 2002: Geography of Bulgaria. Physical and so- Mateo-Tomás, P. & Olea, P.P. 2015: Livestock-driven land
cio-economic geography. — BAS, ForKom, Sofia. (in use change to model species distributions: Egyptian vul-
Bulgarian) ture as a case study. — Ecological Indicators 57: 331–
López-López, P., García-Ripolеs, C., Aguilar, J. M., García- 342.
Lopez, F. & Verdejo, J. 2006: Modelling breeding habi- McIntyre, A.L. 2010: Modelling foraging distribution of
tat preferences of Bonelli’s eagle (Hieraaetus fasciatus) Griffon Vultures (Gyps fulvus) on the island of Crete,
in relation to topography, disturbance, climate and land Greece. — MSc thesis, University of Twente.Dobrev & Popgeorgiev: Eurasian Griffon Vulture habitat preferences in Bulgaria 127
Monsarrat, S., Benhamou, S., Sarrazin, F., Bessa-Gomes, C. geoinformation science and earth observation Enschede,
& Bouten, W. 2013: How Predictability of Feeding The Netherlands.
Patches Affects Home Range and Foraging Habitat Se- QGIS Development Team. 2016: QGIS: Geographic Infor-
lection in Avian Social Scavengers? — PLoS ONE 8: mation System. — Open Source Geospatial Foundation
10.1371. Project. http://qgis.osgeo.org.
Morales-Reyes, Z., Pérez-García, J., Moleón, M., Botella, F., R Development Core Team. 2012: R: A language and envi-
Carrete, M., Donázar, J., Cortés-Avizanda, A., Arrondo, ronment for statistical computing. — R Foundation for
E., Moreno-Opo, R., Jimenez, J., Margalida, A. & Statistical Computing, Vienna, Austria. http://ww-
Sánchez-Zapata, J. 2017: Evaluation of the network of w.R-project.org/
protection areas for the feeding of scavengers in Spain: Sebastián-González, E., Morales-Reyes, Z., Botella, F.,
From biodiversity conservation to greenhouse gas emis- Naves, L., Pérez-García, J., Mateo-Tomás, P., Olea, P.,
sion savings. — Journal of Applied Ecology 54: 1120– Moleón, M., Barbosa, J., Hiraldo, F., Arrondo, E.,
1129. Donázar, J., Cortés-Avizanda, A., Selva, N., Lambertuc-
Moreno-Opo, R., Margalida, A., Arredondo, Á., Guil, F., ci, S., Bhattacharjee, A., Brewer, A., Abernethy, E.,
Martín, M., Higuero, R., Soria, C. & Guzmán, J. 2010: Turner, K. & Sánchez-Zapata, J. 2020: Network struc-
Factors influencing the presence of the cinereous vul- ture of vertebrate scavenger assemblages at the global
ture Aegypius monachus at carcasses: Food preferences scale: drivers and ecosystem functioning implications.
and implications for the management of supplementary — Ecography 43: 1143–1155.
feeding sites. — Wildlife Biology 16: 25–34. Sergio, F. & Newton, I. 2003: Occupancy as a measure of
10.2981/09-037. territory quality. — Journal of Animal Ecology 72: 857–
Moreno-Opo, R., Trujillano, A. & Margalida, A. 2015: Opti- 865.
mization of supplementary feeding programs for Euro- Schick, R., Goldstein, J., & Lutcavage, M. 2004: Bluefin
pean vultures depends on environmental and manage- tuna (Thunnus thynnus) distribution in relation to sea
ment factors. — Ecosphere 6(7): 1–15. 27. 10.1890/ surface temperature fronts in the Gulf of Maine (1994–
ES15-00009.1. 96). — Fisheries Oceanography 13: 225–238.
Muñoz, A.R., Real, R., Barbosa, A.M. & Vargas, M. 2005: Skartsi, T., Vasilakis, D. & Elorriaga, J. 2009: Population
Modelling the distribution of Bonelli’s eagle in Spain: trends and conservation of vultures in the National Park
implications for conservation planning. — Diversity of Dadia-Lefkimi-Soufli forest. – In The Dadia-Lefki-
and Distributions 11: 477–486. mi-Soufli National Park, Greece: Biodiversity, Manage-
Newton, I. 1979: Population Ecology of Raptors. — Buteo ment and Conservation (ed Catsadorakis, G.): 183–193.
Books, Vermillion SD, U.S.A. — WWF-Hellas, Athens.
NSI 2012: Population and Settlements Census in 2011, StatSoft Inc. 2004: STATISTICA (data analysis software
Vol.1: population by regions, municipalities and settle- system), version 7. — StatSoft Inc.,Tulsa, USA: www.
ments, NSI Sofia. (In Bulgarian). statsoft.com.
Olea, P.P. & Mateo-Tomás, P. 2009: The role of traditional Stoychev S., Hristov, H., Iankov, P. & Demerdzhiev, D.
farming practices in ecosystem conservation: The case 2004: Birds in the Bulgarian part of the Eastern Rho-
of transhumance and vultures. — Biological Conserva- dopes. – In Biodiversity of Bulgaria. Biodiversity of
tion 142: 1844–1853. Eastern Rhodopes (Bulgaria and Greece) (eds Beron P.
Olea, P.P. & Mateo-Tomás, P. 2011: Spatially explicit esti- & Popov A.): 881 – 894. — Pensoft & Natural Museum
mation of occupancy, detection probability and survey of Natural History, Sofia.
effort needed to inform conservation planning. — Di- Stoynov, E., Peshev, H., Vangelova, N. & Grozdanov, A.
versity and Distributions 17: 714–724. 2017: Ranging Behaviour of the Griffon Vulture Gyps
Olea, P. & Mateo-Tomás, P. 2013: Assessing Species Habitat fulvus in Kresna Gorge and South-western Bulgaria. —
Using Google Street View: A Case Study of Cliff-Nest- Technical Report on Action A1 of the LIFE11 NAT/
ing Vultures. — PLoS ONE 8: 10.1371. BG/363, Fund for Wild Flora and Fauna, Blagoevgrad.
Parra, J. & Telleria, J.L. 2004: The increase in the Spanish 10.13140/RG.2.2.31588.58242.
population of Griffon Vulture Gyps fulvus during 1989- Sutherland, W. 2000: The Conservation Handbook: re-
1999: effects of food and nest site availability. — Bird search, management and policy. — Blackwell science,
Conservation International 14: 33–41. Oxford.
Pierre-Rossi, J. 1996: Statistical tool for soil biology. XI. Au- Xirouchakis, S. & Mylonas, M. 2004: Griffon Vulture (Gyps
tocorrelogram and Mantel test. — European journal of fulvus) distribution and density in Crete. — Israeli Jour-
soil biology 32(4): 195–203. nal of Zoology 50: 351–354.
Polce, C. 2004: Application of Geographic Information Sys- Xirouchakis, S. & Mylonas, M. 2005: Selection of Breeding
tems to assess environmental suitability for reintroduc- Cliffs by Griffon Vultures Gyps fulvus in Crete (Greece).
tion of Griffon Vulture (Gyps fulvus) in Retezat National — Acta Ornitologica 40: 155–161.
Park, Romania. — Report, International Institute forYou can also read

















































