Influence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot Disease in Mungbean - AWS
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

RESEARCH ARTICLE Legume Research- An International Journal, Volume 45 Issue 1: 104-109 (January 2022)
Influence of Temperature on the Pathogenesis and Gene
Expression of Rhizoctonia solani Causing Web Blight/Wet Root
Rot Disease in Mungbean
Bishnu Maya Bashyal1, Bhupendra Singh Kharayat1, Pooja Parmar1,
Ashish Kumar Gupta3, S.C. Dubey2, Rashmi Aggarwal1 10.18805/LR-4342
ABSTRACT
Background: Mungbean [Vigna radiata (L.) W ilzeck] is one of the most important pulse crops and grown in almost all parts of the
India. Web blight/wet root rot disease of mungbean is caused by Rhizoctonia solani Kühn. Crop environmental factors plays a vital
role in the development of web blight disease caused by R. solani. An understanding of the role of environmental factors on the
infection and survival of the pathogen is necessary to develop disease management practices.
Methods: The effect of different temperatures (4C, 20C, 25C, 30C and 35 C) on mycelial growth of seven different R. solani
isolates belonging to different anastomosis group were evaluated under in vitro conditions. Effect of different temperatures on the
development of root rot/web blight disease of mungbean was also evaluated under phytotron conditions at various temperatures with
constant relative humidity (85%) and illumination (alternate dark and light period of 12 h). Effect of temperatures on the expression of
selected pathogenicity related genes was evaluated through real time PCR.
Result: Maximum radial growth in R. solani isolates was observed at 25 and 30C after 48 hrs of incubation. Maximum disease
incidence was observed with R. solani isolate RUPU-18 (73.11%) followed by R-17 (68.75%), RDLM-1 (63.45%) at 25C on mungbean
genotype Pusa Vishal. Expression of genes like ABC transporter was observed only at 35 C, while other genes like 1, 3 glucan
hydrolase expressed maximum at 25C after 24, 48 and 72 hrs post inoculation. Present study suggested that the expression of
pathogenicity related genes in mungbean-R. solani system is dependent on the temperature and time interval post pathogen inoculation.
Key words: Mungbean, Pathogenesis-associated genes, Real time-PCR, Rhizoctonia solani.
INTRODUCTION 1
Division of Plant Pathology, ICAR-Indian Agricultural Research
Mungbean [Vigna radiata (L.) W ilzeck] is one of the most Institute, New Delhi-110 012, India.
important pulse crop and grown in almost all parts of the 2
Division of Plant Quarantine, ICAR-National Bureau of Plant
India. With increased irrigation facilities, remunerative prices Genetic Resources, New Delhi-110 012, India.
and availability of short duration cultivars, this crop now 3
National Institute for Plant Biotechnology, New Delhi-110 012, India.
occupies considerable area during summer season also in
Corresponding Author: Bishnu Maya Bashyal, Division of Plant
several parts of India. Mungbean is inflicted by a number of
Pathology, ICAR-Indian Agricultural Research Institute, New Delhi-
diseases caused by fungi, bacteria and viruses.
110 012, India. Email: bishnumayabashyal@gmail.com
Wet root-rot and web/aerial bight caused by Rhizoctonia
solani Kühn (Thanatephorus cucumeris Donk) is a How to cite this article: Bashyal, B.M., Kharayat, B.S., Parmar,
destructive disease of mungbean and it is considered one P., Gupta, A.K., Dubey, S.C. and Aggarwal, R. (2022). Influence of
of the important causes for stagnated productivity of the crop Temperature on the Pathogenesis and Gene Expression of
in India. The disease was observed to reduce 33 to 40 per Rhizoctonia solani Causing Web Blight/Wet Root Rot Disease in
cent grain yield and 28.6 per cent in test weight at different Mungbean. Legume Research. 45(1): 104-109. DOI: 10.18805/LR-4342.
level of disease severity and in different varieties of Submitted: 03-02-2020 Accepted: 05-06-2020 Online: 09-11-2020
mungbean (Singh, 2006; Gupta et al. 2010). Web blight
symptoms on mungbean appears on roots, stems, petioles, of the lower leaf from where it extends upwards. The infected
as well as on pods, but the disease is devastating on foliage branches shrivel at the point of infection, leaves on it loose
(Bash yal et al. 2018). Symptoms initiate with the their normal green colour and wither. Leaves and branches
development of small water soaked lesion which enlarged fall down leaving behind the main stem bare. Affected plant
giving blighted appearance with web like mycelium of the parts rot rapidly in warm and wet weather.
pathogen on upper surface (Singh, 2006). The seedling is Rhizoctonia solani, teleomorph Thanatephorus
killed when the lesions on hypocotyle; girdles the stem after cucumeris, is a polyphagous necrotrophic plant pathogen
they coalesce or enlarge in size. During the period of high belongs to order the Basidiomycete (Rioux et al. 2011). To
humidity, the disease spreads very fast. In the beginning, date, R. solani has been splited into 14 anastmosis groups
the infection occurs only on the basal parts including petiole (AGs) designated as AG 1 through 13 and bridging isolate
104 Legume Research- An International Journal
Influence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot...
(BI) group (Carling et al. 2002; Rioux et al. 2011). Several Source of R. solani isolates
AGs are further subdivided into intraspecific groups (ISGs) The culture of R. solani belonging to different anastomosis
based on morphology of cultural growth, nutritional
groups (AGs) were obtained from Pulse Pathology Lab,
requirements, temperature effect on growth, host specificity,
Division of Plant Pathology, ICAR-IARI, New Delhi, which
frequency of hyphal anastomosis and pathogenicity (Sneh
were originally isolated from diseased host plant (Table 1)
et al. 1991). Recent protein and DNA-based studies support
with typical symptoms. The anastomosis grouping and
the separation of R. solani into genetically distinct groupings,
pathogenicity test using Koch’s postulates of these cultures
but has also revealed considerable genetic diversity within
one anastomosis group. Rhizoctonia solani casual agent of were also determined earlier (Dubey et al. 2014).
web blight of mungbean belongs to anastomosis group 1, Effect of temperature on mycelial growth of R. solani
intraspecific group IB (Singh, 2006). Aerial blight pathogen
The influence of different temperature (4C, 20C, 25C, 30C
(R. solani) of soybean belongs to AG-1IA group (Yang and
and 35C) on mycelial growth of seven different R. solani
Chen, 1989) while both IA and IB intraspecific groups are
isolates belonging to different anastomosis group were
responsible for causing foliar blight of legume crops especially
investigated on Potato Dextrose Agar (PDA) medium (200 g
in soybean (Harvillae et al. 1996). Dubey et al (2014) identified
potatoes; 20 g agar; 20 g dextrose in 1 L of water). Each
the seven anastomosis groups (AGs) of R. solani associated
Petri plate was centrally inoculated with 5 mm mycelial disc
with pulse crops. Ganeshamoorthi and Dubey (2015)
suggested AGs did not show host specificity in legumes. from the 48 hours old culture of test fungus. Five replications
However, effect of these AGs on the pathogenicity of were maintained for each treatment and colony diameter
mungbean needs to be studied. was measured after 48 hrs of inoculation.
Environmental factors play a vital role in the development Effect of temperature on web blight disease development
of web blight disease caused by R. solani. An understanding of mungbean under phytotron conditions
of the role of environmental factors on the infection and
Four sets of pot experiments were conducted under
survival of any root pathogen is necessary to develop cultural
controlled environment (plant growth chamber) in phytotron
disease management practices as well as assays that
evaluate host resistance or efficacy of seed treatments. at ICAR-IARI, New Delhi to determine the disease
Documenting the independent and interactive effects of soil development on susceptible variety of mungbean Pusa
moisture and temperature on plant disease is essential for Vishal. The condition for these sets were temperature of
developing effective management strategies or screening 20 C, 25 C, 30 C and 35 C, relative humidity (85%) and
for host resistance. Temperature is relatively easy to control illumination (alternate dark and light period) of 12 hrs. To
experimentally, especially in climate-controlled chambers. study the virulence of different isolates on susceptible
There are very limited information available about the genotype Pusa Vishal, isolates were multiplied on sorghum
influenc e of enviro nm en tal fac to rs fo r exam ple grains and 15 days old inoculums was mixed on soil at the
temperature and relative humidity on the development of rate of 10 g/kg of soil (Bashyal et al. 2018). Disease
root-rot/web blight/areal blight disease of mungbean and incidence was observed at the interval of 15 days and final
th e expressio n o f genes which having vital role in data was calculated as mean value of five observations.
pathogenesis process of R. sola ni. Therefore, the Experiment was conducted on randomized block design
objectives of this study were to investigate the effect of (RBD) with five replications per treatment and analysis of
temperature on web blight disease development and to variance was calculated using two factor (isolates and
elucidate the effect of temperature on the expression of temperature) analysis.
selected pathogenicity associated genes of R. solani
during the pathogenesis process in mungbean. Effect of temperature on pathogenicity related gene
expression
MATERIALS AND METHODS To study the effect of temperature on pathogenicity related
Experiments were conducted in the year 2016-17 at Division gene expression most virulent isolate RUPU-18 was
of Plant Pathology and National Phytotron Facility of ICAR- selected. The above mentioned temperatures (20C, 25C,
Indian Agricultural Research Institute, New Delhi. 30 C and 35C) and humidity conditions were selected for
Table 1: Details of the R. solani isolates taken for the study.
Isolate code Anastomosis group (AG) Geographic locations Host origin
RDLM-1 AG3 Delhi Mungbean
RUPU-58 AG2-2 Uttar Pradesh Urdbean
RUPU-18 AG2-3 Uttar Pradesh Urdbean
RASC-27 AG2-3 Assam Chickpea
RJKM-5 Not determined Jammu and Kashmir Mungbean
RPBC-1 AG3 Punjab Cowpea
Rice-17 AG1-1A Uttar Pradesh Rice
Volume 45 Issue 1 (January 2022) 105
Influence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot...
the experiments under phytotron conditions. For each Rad Labs, Inc). The final volume of 20 µl PCR mixture
temperature twenty seeds of susceptible variety of prepared with 2X dynamo colour flash SYBR green mix dye
mungbean Pusa Vishal were sown in 4 inch pots filled with (Thermo Scientific, USA): 10 µl; forward and reverse
sterilized soil. After 15 days of sowing of mungbean, in all primers: 100 nM and cDNA template: 100 ng, no template
sets of experiment, the soil in pots was inoculated with the controls (NTC) and a reference gene (Actin) was used for
mycelial suspension (10 g in 50 ml water) of 15 days old each treatment. Thermal cycling conditions were as
sorghum grain multiplied inoculums of R. solani isolate follows: 95C for 10 mins followed by 39 cycles of 95C
RUPU-18. After 24 hrs, 48 hrs, 72 hrs and 96 hrs of for 15 secs and 53C for 30 secs and a melt curve from
inoculations, plants were uprooted carefully, preserved in 65C to 95C. Expression of target gene was normalized
liquid nitrogen and stored at -80 C. The experiment was by reference gene and fold expression of target gene was
conducted in completely randomized design with 5 calculated using 2 CT method of Livak and Schmittgen
replications per treatments. (2001). Three biological replicates with 3 technical
Primer designing for pathogenicity associated genes of replicates for each biological replicate was used for the
R. solani entire experiment.
Putative pathogenesis related genes as described by Rioux
RESULTS AND DISCUSSION
et al. (2011) were selected for the gene expression studies.
Effect of temperatures on growth of R. solani isolates
Sequences of these genes were retrieved from National
Center for Biotechnology Information (NCBI). Primers Mycelial growth of seven R. solani isolates was investigated
sequences were designed by the IDT oligo analyser (https:/ under in vitro conditions at different temperatures. Maximum
/www.idtdna.com/Primerquest/Home/Index) at default radial growth in R. solani isolates was observed at 30 C
settings. Primer specificity test was conducted through in followed by 25 C and 20 C after 48 hrs of incubation
silico analysis using Basic Local Alignment Search Tool (Fig 1). However, growth of R. solani at 25C was also at par
(BLAST) of NCBI non redundant data with the identification with 30 C. Significant difference in growth among seven
of short and nearly exact match’s parameters. isolates was observed at 20C. R. solani isolates could not
grow at 4C. Radial growth of one isolate RDLM-1 was found
Real time PCR based expression analysis for the
statistically at par at three temperatures 20C, 25C and 30C.
selected genes
Kumar et al. (2014), in similar studies, have also recorded
Seedlings were kept in pre-chilled mortar and homogenized that the optimum hyphal growth of Rhizoctonia occurred at
in liquid nitrogen with the help of pestle. Total RNA was temperature range of 20 to 30C with an optimum at 25C.
isolated from the powdered samples (100 mg each) using Development of root rot/web blight disease
Tri-reagent (Invitrogen) following the manufacturer protocol.
Further, 2 mg of RNA was treated with RNase- free DNase Development of root rot/ web blight disease in mungbean
(Genetix) and kept at 37 C for 30 minutes. For the was evaluated under phytotron conditions as described
inactivation of DNase, DNase stop solution was given for above (Fig 2). Effect of isolates and temperatures was
10 minutes at 65C. Oligo (dT) 12-18 primers were added significant for the disease development (Table 2). Maximum
to the treated RNA (2 μg) and the sample (20 μl) was briefly disease incidence (73.11%) was produced by R. solani
denatured at 65C for 10 min and chilled on ice for 2 min. isolate (RUPU-18) followed by R-17 (68.75%), RDLM-1
Protocol of MuMLv (Fermentas) was followed for the reverse (63.45%) at 25C (Fig 3). Only RASC-27 isolate produced
transcription. Gene expression was quantified through real disease (36.75%) on mungbean at 20C. When plants were
time PCR detection system (MJ miniopticon-48 wells; Bio- grown at 30C, maximum disease incidence (35.95%) was
10
Diameter of radial mycelial growth (cm)
9
8
7
4 C
6
20C
5
25C
4
30C
3
35C
2
1
0
RPBC-1 RDLM-1 RUPU-58 RUPU-18 RASC-27 RJKM-5 R-17
Different R. solani isolates
Fig 1: Radial growth of different isolates of Rhizoctonia solani at different temperatures.
106 Legume Research- An International Journal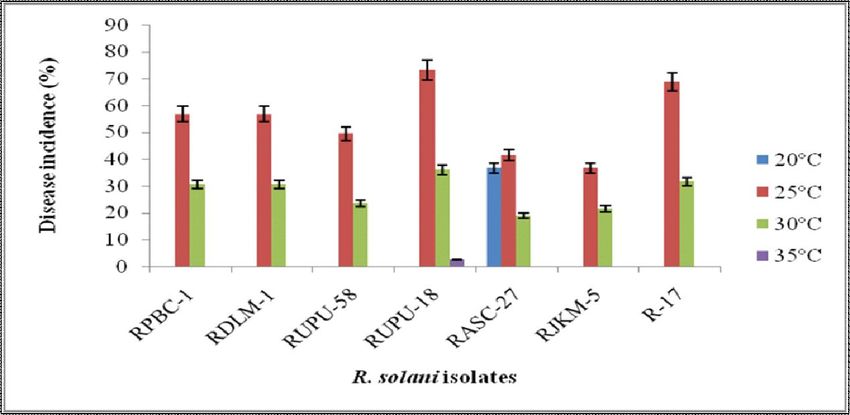
Influence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot...
recorded with RUPU-18 isolate. At temperature, 35C, only disease development on groundnut. The pathogenic ability
two isolates (RDLM-1 and RUPU-18) were able to produce of R. solani at different temperatures enhances the fitness in
disease on Pusa Vishal. Sharma and Tripathi (2001) have nature and might be responsible for making it most successful
noted that the higher aerial temperature (26 to 32C), relative pathogen with wide range under diverse climatic conditions.
humidity near 100% and soil temperature 30-33C favors the
Effect of temperature on expression of pathogenesis
development of high disease severity of web blight of urdbean
related genes
caused by Rhizoctonia solani Kuhn. They also noted that the
rainfall (91-97 mm) has a significant role in severe development List of primers designed for the real time PCR based studies
of web blight during early stage of crop in urdbean. Dubey were presented in Table 3. Primers were screened for the
(1997) have shown that the high temperature (26-28C) and constitutive expression and those present constitutively were
high relative humidity (90-100%) favor severe web blight further selected for the study. Real Time-PCR analysis of
Table 2: Effect of Rhizoctonia solani isolates and temperature on web blight disease development in mungbean.
Source of variation DF Sum of squares Mean squares F-Calculated
Isolates 2 70.77
Temperature 6 28,414.14 4,735.69 632.13
Interaction (Isolates Temperature) 3 548.13 182.71 24.39
Error 18 6,623.29 367.96 49.12
Total 54 404.55 7.49
Table 3: List of genes selected and their primers.
Primers pairs Product Tm
Genes Gene annotations Putative roles in
pathogenesis size (bp) (C)
G1 Glutathione-s-transferase Inactivation and degradation TCCTTATTTCCTCGGTGGTG 169 52
kappa 1 of toxic compounds GGCCACTCCCATTGTATTTG
G2 RAB GTPase Vesicular trafficking CGTCTACGATGTCACGGATG 225 53
and protein secretion GCATCTTCGACACCCTCATT
G3 Pyruvate carboylase Metabolism and CACATCAAGCGAGTTGTGGT 202 52
gluconeogenesis CGAAACCAGACTCTTCGTCA
G4 β-1,3-glucan-hydrolase Cell wall degradation GCAAATATGCGGCATTCTCT 248 50
and host infiltration AAACTATGATCGCGCTGCTT
G5 F-box containing protein/ Translocation of presecretary TTGGACAAAGGGTCATTTCC 165 50
translocation protein Sec72 proteins into the ER GGTGGTTTTGTCGCTTCAAT
G6 AAA perxisomal Appresorium formation TAATCGCGGACTCACTGTTG 190 52
biogenesis protein and toxin production CCATATCGCACTTCCTGGAT
G7 Polysaccharide synthase/gyco- Appresorium formation ACCCGTTGGCAAAAATATGA 196 51
syltransferase family 2 protein CCTCTCGCGATCTCTTGAAC
G8 Potassium: hydrogen antiporter Regulation of pH and ATCGTGGTTGCTTTTCTTGG 223 51
stress response AAGGCGATGTGATGAAGGTC
G9 Potassium: hydrogen antiporter Regulation of pH and ATCGTGGTTGCTTTTCTTGG 223 51
stress response AAGGCGATGTGATGAAGGTC
G10 Phosphatidyl transferease Signaling TTAGATGCCGTTGATGGACA 214 52
ACCTGCCACAAGAGAGCCTA
G11 Nuclear pore protein (Nic 96) Essential for cell death; TGCGTTCTAACAGAGGCGTA 211 52
regulation of gene expression ATTCGTGGACTCAACCAAGC
G12 Nuclear pore protein (Nic 96) Essential for cell death; TGCGTTCTAACAGAGGCGTA 211 52
regulation of gene expression ATTCGTGGACTCAACCAAGC
G13 ABC transporter Efflux and protection GGTTAACGAAGAACCCAGCA 191 53
against plant defense CTGGATGGTCGGTCTTTCAC
G14 Major facilitator superfamily Toxin secretion; efflux; protection AGGTCGTGCATACTCCATCC 169 52
multigrug-DHA1 sub-family protein against plant defense CGCTGTGAAACATGGAAGAA
G15 O-glycosyl hydrolase Cellp wall degradation; CCATCTTGATTTGCGTTGAA 170 50
host infiltration; hydrolysis AGCCCAACATCTTGTGAACC
Volume 45 Issue 1 (January 2022) 107Influence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot...
Fig 2: Mungbean genotype Pusa Vishal grown under phytotron conditions A: Experimental set up at 25oC; B: wet root rot development.
90
80
70
Disease incidence (%)
60
50 20C
40 25C
30 30C
20 35C
10
0
-1 -1 -5 8 U-
18 - 27 -5 17
BC LM PU P SC KM R-
RP R D RU RU RA RJ
R. solani isolates
Fig 3: Root rot/web blight disease development at 20, 25, 30 and 35C by different R. solani isolates.
Fig 4: qPCR based analysis of Rhizoctonia solani pathogenicity related genes in mungbean genotype Pusa Vishal at different
temperature and time intervals. Mean values ± standard deviation of triplicate data from three biological replicates are shown.
108 Legume Research- An International JournalInfluence of Temperature on the Pathogenesis and Gene Expression of Rhizoctonia solani Causing Web Blight/Wet Root Rot...
expression of pathogenesis associated genes showed that ACKNOWLEDGEMENT
the expression of these genes is dependent on the Authors are thankful to the ICAR-NASF (project code BSP-
temperature and time interval post pathogen inoculation (ppi) 4011/2013-14) for financial support and Head, Division of
of isolate RUPU-18 (Fig 4). For gene β-1, 3 glucan hydrolase Plant Pathology, for providing facilities to carry out the
significant expression was observed after 24, 48 and 72 hrs research work.
post pathogen inoculation (ppi) at 25 C. At 92 hrs post
inoculation maximum expression of gene was observed at REFERENCES
30 C. Similarly, gene 7 i.e. Polysaccharide synthtase Bashyal, B.M., Kharayat, B.S., Kumar, J. Dubey, S.C. and Aggarwal,
expression was maximum at 25C after 24, 48 and 72hrs R. (2018). Histopathological Studies of Rhizoctonia solani
post inoculation. At 96 hrs significant expression of the gene infection process in different cultivars of mungbean [Vigna
was observed at 20 and 25 C. Gene 8 i.e. Potassium radiata (L.) W ilczek]. National Academy Science Letter.
hydrogen antiporter was expressed maximum at 20C after 41: 269-273. https://doi.org/10.1007/s40009-018-0669-3.
24 and 48 hrs of inoculation. It was maximum at 25C at 72 Carling, D.E., Baird, R.E., Gitaitis, R.D., Brainard, K.A. and Kuninaga,
hrs ppi and expression was maximum at 35C after 96 hrs S. (2002). Characterization of AG-13, a newly reported
of inoculation. Gene 13 i.e. ABC transporter was observed anastomosis group of Rhizoctonia solani. Phytopathology.
to be expressed maximum at 30C after 24, 48, 72 and 96 92(8): 893-899.
hrs post inoculation (Fig 3). Dubey, S.C. (1997). Influence of age of plants, temperature and
Cell wall polysaccharides, such as β-1, 3-glucan and humidity on web blight development in groundnut. Indian
Phytopathology. 50: 119-120.
chitin, are recognized by host cells as pathogen-associated
Dubey, S.C., Tripathi, A., Upadhyay, B.K. and Deka, U.K. (2014).
molecular patterns (PAMPs) and the fungal cells are attacked
Diversity of Rhizoctonia solani associated with pulse crops
and removed by host immune systems. Glycoside
in different agro-ecological regions of India. World Journal
hydrolases degrade β-glucans, mainly to regulate the
of Microbiology and Biotechnology. 30: 1699-1715.
plasticity of the cell wall for cell expansion, cell division, cell FAO. (2016). http://www.fao.org/pulses-2016/en.
separation and sporulation. Potassium hydrogen antiporter Ganeshamoorthi, P. and Dubey, S.C. (2015). Morphological and
regulates the pH and stress response. Polysaccharide pathogenic variability of Rhizoctonia solani isolates
synthase is responsible for the appressorium formation associated with wet root rot of chickpea in India. Legume
(Rioux et al. 2011). Three maj or events involved in Research-An International Journal. 38: 389-395.
pathogenesis process of R. solani in mungbean are early Gupta, R.P., Singh, S.K. and Singh, R.V. (2010). Assessment of losses
host contact or attachment and penetration, adjustment to due to web blight and weather effects on disease development
the host environment and proliferation of pathogen through in mungbean. Indian Phytopath. 63(1): 108-109.
necrotic tissues (Bashyal et al. 2018). Higher expression of Harville, B.G., Russin, I.S. and Habetz, R.J. (1996). Rhizoctonia
β-1, 3 glucan hydrolase and Polysaccharide synthase at foliar blight reactions and seed yields in soybean. Crop
Sci. 36(3): 563-566.
early hrs of inoculation i.e. 24 and 48 hrs at 25C suggests
Kumar, S., Kumar, A., Chand, G., Lal, M. and Kumar, R. (2014).
the early colonization and pathogenicity at this temperature.
Dynamics of mycelia growth and sclerotia production of
The ABC transporter family is a group of membrane proteins
Rhizoctonia solani Kuhn (AG1-IB) of urdbean. The Ecoscan.
that use the hydrolysis of ATP to power the translocation of 8(3 and 4): 273-277.
a wide variety of substrates across cellular membranes. The Rioux, R., Manmathan, H., Singh, P., des los Reyes, B., Jia, Y. and
ABC transporter of Pseudomonas fluorescens was optimally Tavantzis, S. (2011). Compararitive analysis of putative
functional at 20 and 25C. Transporter proteins are one of pathogenesis-related gene expression in two Rhizoctonia
an organism’s primary interfaces with the environment. The solani pathosystems. Current Genetics. 57(6): 391-408.
expressed set of transporters mediates cellular metabolic Sharma, J. and Tripathi, H.S. (2001). Host range of Rhizoctonia
capabilities and influences signal transduction pathways and solani Kuhn, the causal agent of web blight of urdbean
regulatory networks. [Vigna mungo (L.) Hepper]. Journal of Mycology and Plant
Pathology. 31: 81-82.
CONCLUSION Singh, J. (2006). Occurrence, variability and management of web
blight pathogen (Rhizoctonia solani Kuhn) of mungbean
Our study showed that there is direct link between the
[Vigna radiata (L.) W ilczek]. Ph.D. Thes is, B.H.U.,
temperature and the induction level of key genes that likely
Varanasi (U.P.).
to affect the efficiency in the disease development in Sneh, B., Burpee, L. and Ogoshi, A. (1991). Identification of
mungbean and fitness of R. solani in nature. Real Time- Rhizoctonia Species. The American Phytopathological
PCR analysis of expression of pathogenesis associated Society Press, St. Paul, Minn, USA, pp 133.
genes showed that the expression of these genes is Yang, L.M. and Chen, H.Y. (1989). A preliminary on the infestation
dependent on the temperature and time interval post of brown plant hopper and the infection of sheath blight
pathogen inoculation. of rice. Insect Knowledge. 26: 312-322.
Volume 45 Issue 1 (January 2022) 109You can also read

















































