Recent insights on indirect mechanisms in developmental toxicity of nanomaterials - DORA 4RI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Dugershaw et al. Particle and Fibre Toxicology (2020) 17:31
https://doi.org/10.1186/s12989-020-00359-x
REVIEW Open Access
Recent insights on indirect mechanisms in
developmental toxicity of nanomaterials
Battuja Batbajar Dugershaw1, Leonie Aengenheister1, Signe Schmidt Kjølner Hansen2,3,
Karin Sørig Hougaard2,4 and Tina Buerki-Thurnherr1*
Abstract
Background: Epidemiological and animal studies provide compelling indications that environmental and
engineered nanomaterials (NMs) pose a risk for pregnancy, fetal development and offspring health later in life.
Understanding the origin and mechanisms underlying NM-induced developmental toxicity will be a cornerstone in
the protection of sensitive populations and the design of safe and sustainable nanotechnology applications.
Main body: Direct toxicity originating from NMs crossing the placental barrier is frequently assumed to be the key
pathway in developmental toxicity. However, placental transfer of particles is often highly limited, and evidence is
growing that NMs can also indirectly interfere with fetal development. Here, we outline current knowledge on
potential indirect mechanisms in developmental toxicity of NMs.
Short conclusion: Until now, research on developmental toxicity has mainly focused on the biodistribution and
placental translocation of NMs to the fetus to delineate underlying processes. Systematic research addressing NM
impact on maternal and placental tissues as potential contributors to mechanistic pathways in developmental
toxicity is only slowly gathering momentum. So far, maternal and placental oxidative stress and inflammation,
activation of placental toll-like receptors (TLRs), impairment of placental growth and secretion of placental
hormones, and vascular factors have been suggested to mediate indirect developmental toxicity of NMs. Therefore,
NM effects on maternal and placental tissue function ought to be comprehensively evaluated in addition to
placental transfer in the design of future studies of developmental toxicity and risk assessment of NM exposure
during pregnancy.
Keywords: Nanomaterials, Developmental toxicity, Indirect toxicity pathways, Placental barrier, Pregnancy
Background toxicity of nanosized particles came from epidemio-
Since the thalidomide scandal in the early 1960s, it has logical studies, showing association of particulate matter
become evident that the placenta does not provide a (PM) exposure with adverse pregnancy outcomes such
tight barrier, and that fetuses are exceptionally suscep- as low birth weight, preterm birth and preeclampsia [2–4].
tible to potentially toxic substances compared to adults, Recently, it has been confirmed that environmental black
due to the phases of rapid growth, range of developmen- carbon reaches the fetal side of the placenta in exposed
tal events and often irreversible nature of the induced pregnant women [5]. With the advent of nanotechnology,
changes [1]. The first indications of developmental novel NMs with unique properties can be industrially pro-
duced at large scales for application in food (reviewed in
* Correspondence: tina.buerki@empa.ch [6, 7]), cosmetics (reviewed in [7, 8]), medicine (reviewed
1
Laboratory for Particles-Biology Interactions, Empa, Swiss Federal in [9, 10]) and high-technology products (reviewed in [10,
Laboratories for Materials Science and Technology, Empa, Lerchenfeldstrasse
5, 9014 St.Gallen, Switzerland
11]). These engineered NMs further contribute to human
Full list of author information is available at the end of the article exposure to nanosized particles, and due to their high
© The Author(s). 2020 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License,
which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if
changes were made. The images or other third party material in this article are included in the article's Creative Commons
licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the
data made available in this article, unless otherwise stated in a credit line to the data.
Dugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 2 of 22 reactivity, pose additional health risks. However, investiga- limited [30, 31]. Direct effects on embryonic and fetal tions of the toxicological effects of engineered NMs, espe- tissues have been described for a variety of NMs in sev- cially in vulnerable populations such as pregnant women eral in vitro studies as well as across species, including and their unborn children, have lagged behind the deve- fish, chicken, and in vitro human stem cell (SC) models lopment of new applications. Importantly, to support safe- (reviewed in [32]). However, findings from organisms by-design and sustainable use of NMs, it is imperative to that lack a placenta or have a distinctly different placen- gain knowledge on the potential developmental toxicity of tal structure might not directly correlate to the human NMs and to understand the mechanisms underlying such condition. toxicity. The potential for NMs to affect fetal development by In principle, NMs can affect fetal development indirect pathways has been only marginally investigated through two fundamentally different pathways: a direct and understood. Here, the concept is that NMs can and an indirect pathway [12] (Fig. 1), that, however, are interfere with fetal development in an indirect manner not mutually exclusive. Direct developmental toxicity without being in direct contact with fetal tissue (Fig. 1). may arise from particles in maternal blood that cross the NMs deposited in primary maternal tissue barriers at placental barrier [13–15] and directly damage fetal tis- the point of entry following oral, inhalation, dermal or sues due to their high surface reactivity and propensity intravenous (i.v.) exposure might induce oxidative stress to induce inflammation [16–18], reactive oxygen species and subsequently inflammation, leading to the release of (ROS) [19] and hence oxidative stress reactions [20–22], inflammatory mediators and soluble signaling factors among others. Several FNMs are able to cross primary that can reach the placenta and fetus to induce potential biological tissue barriers (e.g., lung [23, 24] and toxic effects (maternal mediated developmental toxicity). gastrointestinal (GI) tract [24, 25]) as well as the pla- Alternatively, particles reaching the placenta can cause centa [26–29], even if translocation is usually rather similar responses in the placental tissue, compromising Fig. 1 Scheme illustrating direct and indirect pathways of NM-mediated developmental toxicity
Dugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 3 of 22
placental function and inducing the release of placental I.v. injection would make NMs readily systemically
signaling factors, which might impair embryo-fetal de- available. In contrast, only a low fraction of air and food-
velopment (placental mediated developmental toxicity). borne NMs would be expected to reach the systemic cir-
The aim of this review is to (i) collect the current culation and become bioavailable for maternal, placental
knowledge base on the indirect developmental toxicity and fetal tissues. Dermal exposure is expected to con-
of NMs, (ii) compile and describe already known signal- tribute very little to the systemic burden [27, 31]. Once
ing pathways, (iii) propose novel candidate pathways and NMs have reached the systemic circulation, they can
(iv) suggest directions of future research needs. distribute to maternal organs, including the placenta. As
a highly perfused organ, the placenta is extensively ex-
Risks and opportunities of NMs in pregnancy posed to circulating substances. Placental cells have been
For a proper risk assessment of NMs, a central aspect is described to take up nanosized particles from the blood
to understand the exposure of pregnant women to NMs, stream in experimental animals as well as the ex vivo
including all relevant routes of exposure [33]. Due to the human placenta perfusion model (e.g. [48–51]). Studies
use of NMs in many consumer, high-technology and on placental translocation of NMs in rodents, in the hu-
biomedical products, pregnant women could be exposed man ex vivo and in in vitro placenta models have shown
to NMs via inhalation, absorption through damaged that some types of NMs are retained in the maternal cir-
skin, ingestion or injection (Fig. 1) (reviewed in [34, 35]). culation while others can pass the placenta (reviewed in
At production sites with applications of NMs, pregnant [26, 52]). Placental transfer appears to partially correlate
women can be exposed to NMs by inhalation, since the withphysicochemical properties of NMs, in particular
established protective legislation [36] does not come into particle size [26]. However, other factors such as the ges-
action until the employer is made aware of the preg- tational stage or combined physico-chemical properties
nancy, most often not until after the first 4–6 weeks. can also affect placental translocation of NMs, making
Even then, NM exposure might continue, as the regula- this process difficult to predict [53]. As an example, a re-
tion does not specifically regulate NM relative to preg- cent study demonstrated decreased fetal viability and
nancy [37, 38]. Ingestion of NMs used as food additives, growth, when 13 nm zinc oxide (ZnO) NPs were orally
in food packaging material or personal care products, administered (7.2 mg/mouse) during organogenesis (ges-
constitutes another realistic route of exposure during tational day (GD)7–16) in mice. However, when ZnO
pregnancy. For example, the white food colorant E171 NP exposure occurred during the peri-implantation
consists of particulate titanium dioxide (TiO2), with period (GD1-GD10) no fetal toxicity, but a slight change
approximately 17–35% of the particles being within the in placental weight, was observed [54].
nano-range (reviewed in [7, 39, 40]), and is present in For most routes of uptake (inhalation, ingestion and
toothpaste and various food products such as beverages, injection), gestational NM exposure has been associated
soups, cakes or candy in the European Union [41, 42]. In with developmental toxicity for a variety of different
the United States, the dietary intake of TiO2 is estimated NMs (extensively reviewed in [36, 55–58]). However, we
to be 1–2 mg/kg body weight per day for children, and have yet to identify the underlying mechanisms and
0.2–0.7 mg/kg body weight per day for other age groups which particle properties are of particular concern.
[7, 42]. Dermal uptake of NMs present in personal care
products, such as sunscreen, is expected to be minimal Organ systems of relevance for pathways of indirect
since the intact skin forms a tight barrier for NMs developmental toxicity
(reviewed in [43]). Finally, particles may be directly For sure, the placenta should be a key focus in any
injected into the body in case of medical application of mechanistic study on NM-mediated developmental tox-
NMs (reviewed in [9, 44, 45]), but currently, nano- icity due to its position at the interface between mother
medical therapies during pregnancy are still in the and fetus and its numerous essential functions during
investigational stage. For instance, King et al. demon- pregnancy. As a transient organ, the placenta starts
strated the potential of iRGD (9-amino acid cyclic pep- forming after implantation of the conceptus in the uter-
tide: CRGDKGPDC)-decorated liposomes loaded with ine wall. It consists of tissues of maternal (decidua) and
insulin-like growth factor (IGF)-2 for the treatment of fetal origin (amnion, chorion) [59, 60]. Anatomically, the
fetal growth restriction in mice [46]. An oxytocin recep- maternal side of the placenta comprises the multinuclear
tor coated liposomal carrier loaded with the tocolytic syncytiotrophoblast (ST) layer, which is supported by a
drug indomethacin substantially decreased preterm birth basal membrane, underlying cytotrophoblast cells, mes-
rates in mice [47]. Nevertheless, before clinical use in enchymal tissue and the microvascular endothelium of
pregnant women, not only the efficacy of the potential the fetal small blood vessels (Fig. 2). This interface be-
treatment in humans but also the safety of the NMs dur- tween the inner mucous membrane of the uterus (endo-
ing pregnancy needs to be proven. metrium) and the fetus defines the degree to whichDugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 4 of 22
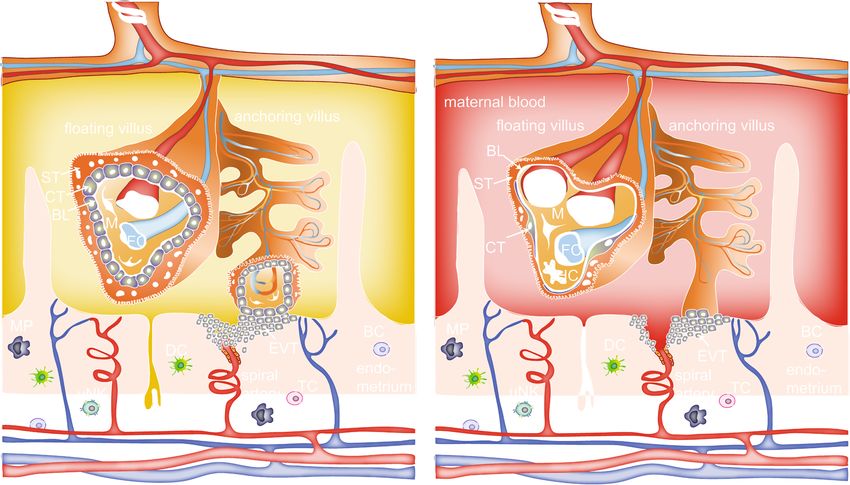
Fig. 2 Scheme of the human placental barrier in early and late pregnancy. In the first trimester, the placental barrier consists of the
syncytiotrophoblast (ST), cytotrophoblasts (CT), basal lamina (BL) and the endothelial cells (E) of the fetal capillaries (FC). Other cell types in the
villous mesoderm include fibroblasts (F) and Hofbauer cells (HC). Various immune cells are also present in the maternal decidual tissue, including
dendritic cells (DC), macrophages (MP), uterine natural killer cells (uNK), T cells (TC) and B cells (BC). Extravillous trophoblasts (EVT) of the
anchoring villi invade the maternal spiral arteries (SA) and form a plug that prevents entry of maternal blood into the intervillous space, and
uterine glands (UG) provide histiotrophic nutrition. After the first trimester, the EVT plug is released and placental villi are now surrounded by
maternal blood. Towards the end of pregnancy, the placental barrier decreases in size by thinning of the ST layer and spreading of the CT layer,
and the FCs move towards the periphery of the floating villi
maternally delivered substances reach the fetal tissue with caution, and the use of physiologically relevant
[61]. During pregnancy, the placenta undergoes dramatic placenta models is encouraged.
structural and functional changes to fulfill the evolving Also, maternal organs could mediate indirect develop-
needs of the developing fetus. During early pregnancy, mental toxicity of NMs. Here, a focus should be on
the placental barrier is relatively thick (20–30 μm) and tissues at the port of entry that are in direct contact with
bilayered [62–64], but thins (2–4 μm) [65], becomes particles such as the lung, the skin or the GI tract upon
predominantly monolayered [62–64], and increases its inhalation, dermal deposition or oral exposure, respect-
surface area tremendously (to approx. 12 m2) towards ively. Uptake and accumulation of NMs in these tissues
the end of pregnancy to allow for efficient exchange of could affect organ functions locally, but effects may
nutrients and gases required to sustain rapid fetal spread to distant sites, including the placenta or the de-
growth. Placental damage, disease or impairment of its veloping fetus, if particles interfere with essential signal-
development or function are responsible for numerous ing pathways. This concept is nicely exemplified in a
pregnancy complications, including preeclampsia [66], recent study in mice, where systemic adverse effects (i.e.
miscarriage [63, 67] and intrauterine growth restriction increased retention of activated leukocytes, secondary
[63, 67], and can likely impact offspring health later in thrombocytosis, and pro-inflammatory responses in
life [68]. It should also be highlighted that the placenta secondary organs) were observed only upon inhalation
is the most species-specific organ among mammals and exposure to carbon NPs, but not after intra-arterial
shows remarkable differences in global structure, tissue injection of an equivalent dose of particles to bypass the
layer organization, trophoblast cell types [69, 70] as well lung [72]. The mechanism(s) underlying the observed
as molecular features [71]. Therefore, translation from indirect systemic toxicity of carbon NPs appeared to
animal studies to the human situation should be done involve inflammatory responses of the lung tissue [72].
In addition to pro-inflammatory actions, NMs may alsoDugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 5 of 22
interfere with essential functions of the lung, skin or GI, cobalt-chrome (CoCr) NPs into pregnant mice on GD 9
such as gas exchange, digestion, nutrient uptake, meta- (0.12 mg per mouse) [88]. In a similar direction, both
bolism or transport (Fig. 1). For instance, ZnO NPs can maternal inhalation with 42 mg/m3 (GD 8–18) [77] and
reduce iron and nutrient uptake and transfer at the injection with 1000 μg/mouse (GD 9) [75] of TiO2 NPs
intestinal barrier [73, 74]. caused behavioral deficits in the offspring.. Finally, im-
munomodulatory effects were reported upon CuO inhal-
Evidence for indirect developmental toxicity ation [78]. Importantly, NM translocation to the fetus
To gather an overview on potential indirect pathways in was addressed but not observed in these studies, there-
developmental toxicity, we searched the open literature fore strongly supporting the presence of indirect toxicity
for studies reporting adverse effects of NMs on gestation pathways. However, it is conceivable that a small
and fetal development in the absence of detectable amount of particles might have crossed the placental
materno-fetal particle transfer (Table 1). However, since barrier, which were below the detection limit of the ap-
direct and indirect toxicity pathways may jointly contrib- plied analytical techniques (i.e. ICP-MS, gAAS, TEM,
ute to developmental toxicity, we also included studies histological and micro-Raman analysis), as for example
that provide hints for indirect toxicity pathways even if suggested by Hougaard et al., 2010 [77]. Moreover, for
placental transfer of NMs was detected (Table 2) or un- soluble NPs (e.g. CuO), placental translocation of small
known (Table 3). The studies are briefly described below quantities of dissolved ions might also partially account
alongside the provided evidence and forwarded hypoth- for developmental toxicity even in the absence of par-
eses for indirect mechanisms of toxicity. ticle transfer. Nevertheless, the adverse effects upon
CuO inhalation in mice observed by Adamcakova-Dodd
Studies without detectable placental particle transfer et al. [78] were not associated with detectable increase in
We identified a total of ten studies that reported devel- fetal or maternal blood Cu levels. Proposed pathways for
opmental toxicity in the absence of detectable NM indirect developmental toxicity included both placenta-
translocation across the placental barrier (Table 1). Most and maternally mediated secondary mechanisms. Mater-
used pregnant mice as the experimental model, but a nally mediated pathways comprised oxidative stress,
few studies used in vitro cell culture systems for more inflammatory, immune and endocrine responses [75, 77,
mechanistic studies. Gestational and litter parameters 78, 83, 84], whereas placental mediated pathways in-
were affected in four of the murine studies, including re- volved oxidative stress, inflammation, placental insuffi-
duced survival rate of offspring from dams inhaling cop- ciency, release of mediators (e.g., ATP, IL-6) and
per oxide (CuO) (3.5 mg/m3 at GD 3–19) [78], changes in placental transport of zinc, vitamin B12,
decreased fetal size and delayed neonatal growth from micronutrients or oxygen [76, 79–88]
cadmium oxide (CdO) NP inhalation (100 μg/m3 or
230 μg/m3 at GD 4.5–16.5) [83], and increased fetal re- Studies with placental particle transfer
sorption and malformations following maternal exposure Several publications suggested a role for indirect devel-
to SWCNTs by the oral (10 or 100 mg/kg body weight opmental toxicity of NMs even if particles in some cases
at GD 9) [84] and i.v. route (10 ng to 30 μg/mouse at were shown to cross the placental barrier and adverse
GD 5.5) [85]. Other studies described effects on placen- effects could have resulted from direct embryo-fetal
tal structure and function, offspring lung development exposure (Table 2). The gestational and litter parameters
and function and neurodevelopment. Regarding the pla- described in these studies include increased rate of mis-
centa, intratracheal instillation of TiO2 and cerium diox- carriage from quantum dot (QD) injection [89], smaller
ide (CeO2) NPs (total 300 μg/mouse: 100 μg on GD 2.5, fetuses and increased fetal resorption from silica dioxide
9.5 and 16.5, respectively) decreased placental efficiency (SiO2) and TiO2 NP injection [90], growth retardation
[76], injection of CdO NPs altered placental weight [83] from diesel exhaust particle (DEP) inhalation [93] or
and injection of SWCNTs induced vascular lesions and multi-walled carbon nanotube (MWCNT) injection [94]
increased placental level of ROS [85]. Gestational NM and fetal organ damage from QD [89] or SWCNT/
exposure can also affect maternal and fetal lungs as MWCNT injection [94, 95]. The maternal parameters
exemplified by maternal lung inflammation induced by reported were decreased maternal body weight upon
inhalation of TiO2 (42 mg/m3 on GD 8–18) [77] or CuO SiO2 and TiO2 NP injection (0.8 mg/mouse on GD 16
NPs (3.5 mg/m3 on GD 3–19) [78], or long-lasting and 17) [90] and hepatocellular injury from QD injection
impairment of lung development in the offspring result- (100 mg/kg body weight on GD 17 in mice and 25 mg/
ing from maternal intratracheal instillation of TiO2 or kg on GD 100 in monkeys) [89]. Paul et al. observed
CeO2 NP [76]. Neurodevelopmental abnormalities, like long-lasting impairment of lung development in off-
reactive astrogliosis and increased DNA damage in the spring of pregnant mice intratracheally instilled with sil-
fetal hippocampus, have been observed after injection of ver (Ag) NPs (total 300 μg/mouse: 100 μg on GD 2.5, 9.5Table 1 Studies with evidences for indirect fetotoxicity pathways without placental transfer of NMs
NP type/ NP size exposure/model application placental developmental developmental toxicity (other hypothesis by authors on indirect publication
coating route/dose/ transfer toxicity (gestational parameters) toxicity pathways
exposure and litter parameters)
period
TiO2 5–6 nm mouse i.v./ 100 or not detected no overt fetal behavioral deficits relevant to ASD maternal mediated unknown [75]
1000 μg/ in fetus or malformations or and related neurodevelopmental pathways due to absence of
mouse/ GD9 placenta by changes in disorders in neonates particles in placenta and fetal
ICP-MS pregnancy tissues
outcomes/ no
impact on postnatal
growth
TiO2/CeO2 12.3 ± 0.1/ 22.4 ± mouse instillation/ Ti and Ce not evaluated long-lasting impairment of lung probably involves placental [76]
0.2 nm total 300 μg/ detected in development in offspring/ insufficiency secondary to the
mouse/ the placenta decreased placental efficiency presence of NPs in this organ with
Dugershaw et al. Particle and Fibre Toxicology
100 μg at GD but not in together with the presence of NPs ensuing down regulation of critical
2.5, GD 9.5 fetal tissues in the placenta/ no increase of mediators of lung development
and GD 16.5) by ICP-MS inflammatory mediators in amniotic without any amniotic fluid or fetal
fluid, placenta or offspring lungs/ lung inflammation/ not mediated
decreased pulmonary expression of via fetal or maternal lung
VEGF-α and MMP-9 at the fetal inflammation
stage (GD 17.5) and FGF-18 at the
(2020) 17:31
alveolarization stage (postnatal day
14.5)
UV-Titan 20.6 ± 0.3 nm mouse inhalation/ 1 not detected no impact on moderate neurobehavioural dissolution and translocation of [77]
L181/ h/day to 42 in fetal liver gestational and litter deficits/ persistent lung contaminating metal ions/
polyalcohols mg/m3/ GD by ICP-MS parameters inflammation in pregnant dams placental transfer of inflammatory
8–18 cytokines released from NP-
exposed maternal lung tissue
CuO 16 nm mouse inhalation/ not detected survival rate of 7 maternal pulmonary inflammation/ changes in maternal inflammatory [78]
3.5 mg/m3 for by ICP-MS week old pups no histopathological changes of and immune responses
4 h/day/ GD (similar Cu reduced/ no impact placenta tissue/
3–19 levels in pla- on litter size, male/ immunomodulatory effects in
centa and female ratio, body offspring (differential expression of
fetus as weight and lenght at several Th1/Th2 or other immune
controls) birth response genes in spleen)
CdO 11–15 nm mouse inhalation/ Cd detected decreased incidence altered placental weight disruption in placental oxygen [83]
100 μg/m3 in placenta of pregnancy/ transfer by Cd [79]/ decrease in
every other but not in decreased fetal fetal length could be due to
day or 230 fetus by gAAS length/ delayed alterations in the fetal and/or
μg/m3 daily and ICP-MS neonatal growth/ maternal IGF system [80, 81]/
for 2.5 h/ GD (Cd in delayed maternal changes in the placental transport
4.5–16.5 placenta) weight gain of zinc, vitamin B12, and other
micronutrients due to placental Cd
[82]
SWCNT /OH- 1–2 nm diameter mouse oral/ 10 mg/ not detected increased fetal none oxidative stress and inflammatory [84]
functionalized and 5–30 μm kg or 100 mg/ in placenta, resorption and fetal response in placenta/maternal
length kg/ GD 9 fetal liver and morphological and tissue
fetal kidney skeletal abnormalities
Page 6 of 22
by TEM at 10 mg/kg but notTable 1 Studies with evidences for indirect fetotoxicity pathways without placental transfer of NMs (Continued)
NP type/ NP size exposure/model application placental developmental developmental toxicity (other hypothesis by authors on indirect publication
coating route/dose/ transfer toxicity (gestational parameters) toxicity pathways
exposure and litter parameters)
period
at higher dose
SWCNT/non- 2.37 nm diameter, mouse i.v./ 10 ng to not detected high percentage of vascular lesions and increased ROS oxidative stress in placental tissue [85]
oxidized, 0.85 μm length/ 30 μg/mouse/ by early miscarriages in placenta/ increased ROS in
oxidized and 1.58 nm diameter, GD 5.5 histological and fetal malformed fetuses/ no increased
ultra-oxidized 0.76 μm length/ and micro- malformations; ROS or evident morphological
1.8 nm diameter, Raman lowest effective dose alterations in maternal tissues
0.37 μm length analyses 100 ng/mouse
CoCr 29 nm BeWo Transwell 40 μg/ml/ 24 not detected not applicable DNA damage to the fibroblasts fetal damage mediated by [86]
bilayer with h by ICP-MS without significant cell death/ placental tissue via release of
Dugershaw et al. Particle and Fibre Toxicology
underlying BJ (similar Co mechanism involving transmission mediators (e.g. ATP)
fibroblasts and Cr levels of purine nucleotides (e.g. ATP) and
in whole fetus intercellular signalling within the
as controls) placental barrier through connexin
gap junctions or hemichannels and
pannexin channels
CoCr 29 nm Bewo Transwell not detected not applicable DNA damage to fibroblasts or Oct4- indirect toxicity only across [87]
(2020) 17:31
40 μg/ml / 24
mono- or bilayers h [86] hES cells only with BeWo double bilayered (human)/multilayered
with underlying BJ layer (mice) placental barrier
fibroblasts or
Oct4-hES
mouse i.v./ 0.12 mg or not detected no pathological DNA damage in neonatal blood
0.012 mg/ by ICP-MS changes in neonatal and liver at GD 12.5 (placenta with
mouse/ GD (similar Co visceral organ three layers established) but not at
9.5 or 12.5 and Cr levels GD 9.5 (nutrient exchange via
in whole fetus uterus and yolk sac)/ no
as controls) pathological changes in placenta
CoCr 29 nm Bewo Transwell 40 μg/ml / 24 not detected not applicable altered differentiation of human exposure of the human placenta to [88]
bilayers and h [86] NPC and DNA damage in the CoCr NPs could initiate a singalling
conditioned derived neurons and astrocytes/ cascade that perturbs the
media transfer to importance of autophagy and IL-6 relationship between astrocytes
NPC or NPC- release from placental tissue in NP- and neurons during
derived astrocytes induced DNA-damaging singalling/ neurodevelopment
and neurons NPs can cause developmental
neurotoxicity across placental bar-
mouse i.v./ 0.12 mg / not detected see [87] riers/ astrocytes are key mediators
dpc 9 [87]
of this neurotoxicity/ fetal hippo-
campus is particularly affected in
mice
ASD autism spectrum disorders, gAAS graphite furnace atomic absorption spectroscopy, FGF-18 fibroblast growth factor 18, GD gestation day, ICP-OES inductively coupled plasma optical emission spectrometry, IGF
insulin growth factor, i.v. intravenous, MMP-9 matrix metalloproteinase 9, NP nanoparticles, NPC neural progenitor cells, ROS reactive oxygen species, TEM transmission electron microscopy. VEGF-α vascular endothelial
growth factor α
Page 7 of 22Table 2 Studies with evidences for indirect fetotoxicity pathways with placental transfer of NMs
NP type/ NP size Exposure/ application placental transfer developmental toxicity developmental toxicity (other hypothesis by authors on indirect publication
coating model route/dose/ (gestational and litter parameters) toxicity pathways
exposure parameters)
period
CdSe/CdS/ZnS 60 nm mouse i.v./ 100 mg/ increased Cd no gestational or fetal no significant abnormalities in acute hepatocellular injury and [89]
quantum kg/ GD 17 levels in umbilical abnormalities or complications maternal blood biomarkers, possible stress caused by the
dots/PEG- cord and fetuses histopathology or behavior injection did eventually
phospholipid by ICP-MS contribute to the high
micelle miscarriage rate in macaques
macaques i.v./ 25 mg/ slightly increased increased rate of miscarriage no pathological changes in the
kg/ GD 100 Cd levels in fetal placenta or major organs of the
organs by ICP-MS miscarried fetuses/ no
inflammatory response or injury
in maternal liver and lung tissues/
acute maternal hepatocellular
Dugershaw et al. Particle and Fibre Toxicology
injury
Si and TiO2 70 nm and 35 nm mouse i.v./ 0.8 mg/ Si and TiO2 NP in decrease of maternal body Si NP induced structural and adverse effects are linked to [90]
mouse /GD placenta, fetal weight at GD 17/18/ lower functional abnormalities in structural and functional
16 and 17 liver and brain by uterine weights/ higher fetal placenta (decreased sFlt-1)/ hep- abnormalities in the placenta/
TEM resorption rates/ smaller fetuses arin improved fetal weight and activation of coagulation,
sFlt-1 levels in Si NP exposed complement and oxidative stress
(2020) 17:31
mice in the placenta
Ag 12.3/ 22.4 / 10.4 mouse instillation/ Ag in placenta not evaluated long-lasting impairment of lung probably involves placental [76]
nm total and fetal lung by development in offspring/ insufficiency secondary to the
300 μg/ ICP-MS decreased placental efficiency presence of NPs in this organ
mouse/ together with the presence of with ensuing down regulation of
100 μg at NPs in the placenta/ no increase critical mediators of lung
GD 2.5, GD of inflammatory mediators in development without any
9.5 and GD amniotic fluid, placenta or amniotic fluid or fetal lung
16.5 offspring lungs/ decreased inflammation/ not mediated via
pulmonary expression of VEGF-α fetal or maternal lung
and MMP-9 at the fetal stage (GD inflammation/ combination of
17.5) and FGF-18 at the alveolari- direct and indirect pathways
zation stage (postnatal day 14.5) possible due to low placental
transfer of Ag
Ag 18–20 nm mouse inhalation/ Ag in maternal increased number of resorbed reduced oestrogen plasma levels adverse effects at least in part [91]
1 or 4 h/day tissues, placenta foetuses (in 4 h/day exposures)/ increased related to the release of
to 640 μg/ and fetus by expression of pregnancy-relevant inflammatory mediators by the
m3/ GD 0.5– TEM/ no particles inflammatory cytokines in the pla- placenta/ reduction of circulating
14.5 or ions detected centas/ no major pathological oestrogen levels could indicate
by spICP-MS changes in the lung of the an endocrine disrupting action of
mothers and only minor lesions Ag NPs
in maternal liver and kidney
Ag/ PEGylate 2–15 or 5–15 nm ex vivo 40 or 75 μg/ low levels of Ag not applicable low translocation of Ag ions and low translocation but comparably [92]
or carboxylate human ml / 6 h NPs > 25 nm in Ag NPs (below 0.02% of initial high accumulation of ionic Ag
placenta perfusion fetal circulation dose)/ considerable uptake of Ag and Ag NPs in placental tissue
perfusion by spICP-MS NPs in placental tissue (4.2% of may result in indirect placenta-
initial dose for AgCOONa; 0.75% mediated developmental toxicity
for AgPEG)
Page 8 of 22Table 2 Studies with evidences for indirect fetotoxicity pathways with placental transfer of NMs (Continued)
NP type/ NP size Exposure/ application placental transfer developmental toxicity developmental toxicity (other hypothesis by authors on indirect publication
coating model route/dose/ (gestational and litter parameters) toxicity pathways
exposure parameters)
period
Diesel exhaust 69 nm rabbit inhalation/ non-aggregated growth retardation reduced placental efficiency/ adverse effects on placental [93]
1 mg/m3 for and “fingerprint” reduced placental vascularization/ structure and function and
2 h/day, 5 NP observed in reduced plasma insulin and IGF1 reduced plasma IGF-1 may con-
days/week/ maternal blood, concentrations/ in second tribute to the observed growth
GD 3–27 trophoblasts and generation, fetal metabolism was retardation/ effects could be due
fetal blood by modified to either NP or contaminants
TEM (e.g. PAHs)
MWCNT/ 1–2 μm length, mouse i.v./ 20 mg/ NPs in placental poor embryo development/ decreased progesterone levels fetal growth restriction due to [94]
oxidized and diameter 20–30 kg/ GD17 tissue and foetal fetal growth restriction/ and increased oestradiol levels in vascular reduction in the
99m
Tc nm liver, lung and embryonic death/ abortion/ serum/ decreased VEGF levels placenta/ toxicity higher in first
Dugershaw et al. Particle and Fibre Toxicology
heart by reduced fetal weight/ fetal heart and increased ROS amounts in time pregnancies as adaptations
radioactivity and brain damage placental tissue/ number of in the placenta may occur/
measurements placental blood vessels decreased oMWCNT affect secretion of
progestational hormones
SWCNT and SWCNT:1–2 nm mouse i.v./ 2 mg/kg all CNTs in larger sized MWCNT restricted MWCNTs directly triggered p53- placenta mediated toxicity [95]
MWCNT/ diameterMWCNT: (p53+/+; or 5 mg/kg/ placental tissue the development of fetuses and dependent apoptosis and cell thorugh interference with
(2020) 17:31
amine- < 8 nm, 20–30 nm p53 +/−; GD 10.5, and fetal liver by induced brain deformity (only at cycle arrest in response to DNA placental function
functionalized or 50 nm p53 −/−) 12.5 or 15.5/ positron emission GD 10.5 and only in p53−/− damage/ N-acetylcysteine (anti-
(PL-PEG-NH2)/ diameter, 500– single or tomography fetuses)/ SWCNTs and smaller oxidant) pevented CNT-induced
64
Cu for 2000 nm length repeated sized MWCNTs showed no or nuclear DNA damage andreduce
translocation doses less fetotoxicity brain development abnormalities
FGF-18 fibroblast growth factor 18, GD gestation day, ICP-OES/MS inductively coupled plasma optical emission spectrometry/mass spectroscopy, IGF insulin growth factor, i.v. intravenous, MMP-9 matrix
metalloproteinase 9, NP nanoparticles, PAH polycyclic aromatic hydrocarbons, ROS reactive oxygen species, spICP-MS single particle ICP-MS, TEM transmission electron microscopy; VEGF-α vascular endothelial growth
factor α
Page 9 of 22Table 3 Studies with evidences for indirect fetotoxicity pathways with unknown placental transfer of NMs
NP type/ NP size Exposure/ application route/ developmental toxicity developmental toxicity (other parameters) hypothesis by authors on indirect publication
coating model dose/exposure period (gestational and litter toxicity pathways
parameters)
TiO2 21 nm rat inhalation/ not evaluated increased placental vascular resistance and impaired fetoplacental vascular [96]
cummulative lung impaired umbilical vascular reactivity reactivity/ altered placental reactivity
burden of 525 μg/ and anatomy
GD 11–16
Si 70 nm mouse i.v. injection/ 0.025 or increased fetal resorption and particle uptake in placenta/ 0.04 mg/ml: placental inflammation [16]
0.04 mg/g/ GD 13–14 reduced fetal weight at 0.04 abnormalities in placental structure and
mg/ml reduced placental weight/ nanosilica
upregulated the inflammasome component
NLRP3 and induced placental inflammation
and ROS, resulting in pregnancy
complications/ pregnancy complications
Dugershaw et al. Particle and Fibre Toxicology
were dependent on the balance between
an inflammatory cytokine (IL-1a) and an
anti-inflammatory cytokine (IL-10)/ compli-
cations were completely prevented by ei-
ther inhibition of ROS generation or forced
expression of IL-10
(2020) 17:31
CdTe 2 nm rat i.p./ 5, 10 or 20 mg/ dose dependent placental tissue damage (decreased impeded embryogenesis due to the [97]
quantum kg/ GD 13 embryotoxicity/ reduced placental weight, abnormal morphological placental damage rather than QD
dots survival rate of fetuses/ features) penetration and accumulation in the
reduction of fetal body length fetuses/ distinct developmental toxicity
and mass/ disturbed effects than upon Cd2+ exposure
ossification of limbs
CdTe 3 nm/ 10–20 BeWo/ 0–25 μg/mL/ 24 h not applicable reduction of β-hCG secretion at sub-lethal interference with hormone release [98]
quantum nm HVMF concentrations
dots/ CuO placental
microtissues
Dendritic 5–7 nm first 10 nM and 1 μM/ 24 not applicable charge-dependent accumulation of potentially hazardous influences of [99]
polyglycerol/ trimester h particles/ no major acute toxicity but charged dendritic polygylcerol particles
sulfate, placental reduced secretion of β-hCG for charged on early placental physiology by
amine or explants particles at the lower concentration reduction of β-hCG hormone levels
neutral
MWCNT 13 μm length mouse i.p or intratracheally/ fetal malformations/ increased none inflammatory mechanism [100]
2,3,4 or 5 mg/kg/ GD leucocyte and related
9 hemocyte number and
increased weight of spleen in
dams
CB 14 nm mouse inhalation: 42 mg/m3/ neither inhalation nor DNA strand breaks in maternal and translocation across lung, GI tract and [17]
1 h/day/ GD 8–18 instillation affected gestation offspring liver after inhalation but not placenta expected to be very low for
instillation: 2.75, 13.5 and lactation instillation exposure/ persistent lung highly insoluble CB; changes in
or 67 μg/mouse/ GD inflammation in exposed mothers signalling cascades proposed e.g.
7, 10, 15 and 18 inflammatory molecules
CB 14 nm mouse intratracheal see (Jackson 2011) changes in the expression of several genes responses in newborns secondary to [101]
instillation/ 2.75, 13.5 and proteins associated with inflammation inflammation in dams
Page 10 of 22
or 67 μg/mouse/ GD in maternal lungs/ hepatic response inTable 3 Studies with evidences for indirect fetotoxicity pathways with unknown placental transfer of NMs (Continued)
NP type/ NP size Exposure/ application route/ developmental toxicity developmental toxicity (other parameters) hypothesis by authors on indirect publication
coating model dose/exposure period (gestational and litter toxicity pathways
parameters)
7, 10, 15 and 18 offspring at highest dose
CB/ TiO2/ not mouse intratracheal not evaluated increased allergic susceptibility in offspring components of DEP (especially PAHs) [102]
DEP determined instillation/ 50 μg/ could mediate pro-allergic effects by in-
mouse/ GD 14 creased production of Th2 cytokines
(e.g., IL- 4), known to be important me-
diators of allergy and asthma
graphene 4 different 2D BeWo or 0–40 μg/mL/ 6 h, 24 h not applicable particle uptake in BeWo cells/ no major interference with hormone release and [103]
oxide sizes (1– BeWo or 48 h acute toxicity but reduced secretion of β- barrier integrity
40 μm; 20 Transwell hCG and transient reduction in barrier
nm-1.4 μm; cultures integrity
Dugershaw et al. Particle and Fibre Toxicology
0.2–1 μm;
10–30 μm)
PM2.5 < 2.5 μm human ambient PM2.5 not evaluated positive relationship between PM2.5 intrauterine inflammation upon PM2.5 [49]
exposures over the exposure during preconception and exposure in pregnancy may influence
entire pregnancy pregnancy and intrauterine inflammation subsequent fetal growth, development,
from 5.54 to 29 μg/ and health outcomes
m3
(2020) 17:31
PM10 < 10 μm human mean exposure levels not evaluated short-term maternal PM10 exposure was exposure to air pollution during [104]
during pregnancy modestly associated with elevated maternal pregnancy may lead to maternal and
were 30.3 μg/m3 for CRP levels in early pregnancy and that fetal inflammatory responses
PM10 and 39.9 μg/m3 long-term maternal PM10 and NO2 expos-
for NO2 ure during pregnancy was associated with
elevated fetal CRP levels at delivery
PM10 < 10 μm human mean exposure levels not evaluated associations of PM10 and NO2 exposure maternal air pollution exposure may [104]
during pregnancy with changes in fetal sFlt-1 and PlGF levels influence markers of placental growth
were 30.3 μg/m3 for at delivery/ higher PM10 and NO2 expo- and function
PM10 and 39.9 μg/m3 sures were associated with lower placenta
for NO2 weight/ air pollution exposure was not con-
sistently associated with other markers of
placental growth and function
BC black carbon, CB carbon black, DEP diesel exhaust particles, GD gestation day, ICP-OES inductively coupled plasma optical emission spectrometry, hCG human chorionic gonadotropin, HVMF human villous
mesencyhmal fibroblasts, IGF insulin growth factor, i.p. intraperitoneally, i.v: intravenous, NP nanoparticles, PAHs polycyclic aromatic hydrocarbons, PM particulate matter, ROS reactive oxygen species, TEM transmission
electron microscopy
Page 11 of 22Dugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 12 of 22
and 16.5, respectively) and suggested that the underlying Candidate pathways for indirect developmental toxicity
mechanisms may involve placental insufficiency with en- Developmental toxicity is mostly assessed in experimen-
suing down-regulation of critical mediators of lung de- tal animals and often centers on classical gestational and
velopment [76]. Other proposed placenta mediated litter parameters. In light of the growing evidence for
indirect pathways involve adverse effects of NMs on pla- maternal and placenta mediated developmental toxicity
cental structure and function [90, 92, 93, 95], the release of NMs, it is crucial to perform more comprehensive
of placental inflammatory mediators [91], reduction in assessments of placental, maternal and fetal/offspring
placental vasculature [93, 94] and activation of coagula- tissue and organ functions. In this section, we will com-
tion, complement and oxidative stress in the placenta pile and discuss the different indirect pathways for-
[90] and disruption of endocrine signaling [91, 93]. warded in the reviewed literature, to outline how NMs
may adversely affect developmental outcomes without
Studies with unknown placental particle transfer direct exposure of the fetus to NMs. Although we will
In several studies, placental translocation was not mostly describe individual indirect pathways, these are
assessed, but the authors nevertheless postulated a role likely interlinked and jointly contribute to adverse fetal
for indirect pathways of developmental toxicity based on outcomes, potentially even in conjunction with direct
observed interference of NMs with maternal organs or toxicity pathways.
placental function (Table 3). Most of these studies did
not evaluate gestational and litter parameters, but alter- NM-induced oxidative stress and inflammatory responses
ations of these parameters have previously been de- The placenta has a very high turnover of oxygen and
scribed following maternal exposure to TiO2 NPs [90], ROS are generated continuously, with the main source
PM [2–4] and DEPs [93]. Injection of SiO2 NPs in preg- being the mitochondrial respiratory chain. Overall, the
nant mice (0.025 or 0.04 mg/g body weight on GD 13 balance between oxidants and antioxidants is vital for
and 14) resulted in increased fetal resorption and re- maintaining physiological homeostasis. During normal
duced fetal weight, possibly through particle-induced in- pregnancy conditions, ROS are eliminated by the
flammatory responses in the placental tissue [16]. These corresponding and abundant production of antioxidants
complications were entirely prevented by ROS inhibitors by the feto-placental unit. If this redox balance is dis-
or forced expression of IL-10 [16]. Maternal or intra- turbed pathological conditions may emerge [105]. Sev-
uterine inflammatory pathways were also proposed to eral types of NMs induce the generation of ROS, either
mediate developmental toxicity from exposure to air directly or via activation of oxidative enzymatic pathways
pollution [49, 104] carbon black (CB) [17, 101] and [106–109]. Excessive amounts of ROS may overwhelm
MWCNTs [100]. Besides inflammatory pathways, inter- the capacity of the intrinsic antioxidants and result in a
ference with the placenta (structure, growth or function/ condition of oxidative stress [110]. ROS can damage
reactivity) has been suggested to constitute another in- cells by interaction with lipids, proteins and DNA and
direct pathway for developmental toxicity of air by induction of inflammation [108, 111]. Placental in-
pollution particles [104], cadmium telluride (CdTe) QDs flammation is a well-established risk factor for preg-
[97], TiO2 NP [96] in vivo or graphene oxide (GO) nancy and fetal development [112]. If NMs are taken up
in vitro [103]. For prenatal exposure to CB, TiO2 and by placental cells, the subsequent generation of ROS,
CeO NPs (intratracheal instillation: 50 μg/mouse on GD oxidative stress and inflammation has been hypothesized
14), Fedulov et al. observed increased allergic suscepti- to represent one indirect mechanistic pathway by which
bility in the offspring that was proposed to occur due to NMs can interfere with placental development and
NM-induced production of Th2 cytokines in maternal function, and hence, with fetal development [36, 55].
lungs [102]. Inhaled particles that deposit in the lung alveoli can
Overall, for all of the three study categories (studies also locally induce ROS and inflammation. This will
investigating but not detecting placental transfer often be accompanied by increased transcription of pro-
(Table 1), studies detecting placental transfer (Table 2) inflammatory genes and ultimately the production of
and studies with unknown placental transfer (Table 3)), inflammatory mediators, such as cytokines and acute-
indications of potential indirect toxicity pathways medi- phase proteins that can become systemically available
ated by maternal and/or placental tissue have been identi- [113, 114]. It is increasingly being described that mater-
fied. Moreover, considering that maternal and placental nal inflammation is a potent modulator of fetal develop-
tissues are probably exposed to NMs at earlier time points ment and that the developing nervous system may be
and higher dose levels compared to the fetal compart- especially sensitive. Maternal inflammation has been
ment, extending the focus from direct to indirect effects is proposed to constitute an immune challenge to the fetus
of key importance to advance our understanding of risks that could prime early alterations in the inflammatory
associated with NM exposure during pregnancy. response systems and, in turn, disrupt development andDugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 13 of 22 maturation of the central nervous system and enhance enhanced, astrogliosis in the offspring [117]. Another sensitivity to additional stress factors [115]. Maternal in- study investigated the developmental effects of flammation may not necessarily result in fetal inflamma- MWCNTs injected intravenously to pregnant p53+/− tion, but the placenta may act as a sensor of maternal mice (2 mg/kg or 5 mg/kg body weight on GD 10.5, 12.5 inflammation and subsequently adapt to the inflamma- or 15.5 as a single or repeated dosis). MWCNTs in- tory environment and may act both as a target and a creased the incidence of brain defects in the offspring producer of inflammatory mediators [116]. and decreased offspring survival rate after birth. The Shirasuna et al. (2015) elegantly aimed to investigate if underlying mechanism seemed to involve MWCNTs dir- NPs induce pregnancy complications through placental ectly triggering p53-dependent apoptosis and cell cycle inflammation [16]. Pregnant mice were injected i.v. with arrest in response to DNA damage. Co-injection of an 0.04 mg/kg body weight of 70 nm silica particles on GD antioxidant markedly decreased the number of fetuses 13 and 14. This exposure increased fetal resorptions, in- with brain defects, indicating that oxidative stress may duced placental dysfunction, ROS generation and infil- be implicated. In this study, MWCNTs were found to tration with neutrophil granulocytes (3-fold). Also, distribute to the placenta and fetal liver but were not ob- placental protein levels of several inflammatory cyto- served in the fetal brain [95]. Finally, intratracheal instil- kines were significantly increased (IL-1β, IL-6, TNF-α, lation of 4–5 mg MWCNTs/kg to pregnant mice on GD and CCL2). Pre-treatment with the antioxidant N-acetyl 9 was found to induce fetal malformations and to signifi- cysteine (NAC) almost completely reversed the placental cantly increase maternal leukocyte counts in peripheral and fetal effects of the injected NPs, reduced placental blood. At a lower dose of 3 mg/kg, no abnormality oc- ROS levels, cell infiltration and secretion of IL-1β and curred. This suggests that maternal inflammation may IL-1α. Findings in specific knock-out mouse strains indi- be contributing to fetal toxicity [100]. cated that the balance between the inflammatory cyto- Overall, these findings offer evidence of the involve- kine IL-1 and the anti-inflammatory IL-10 was pivotal ment of oxidative stress in developmental toxicity of for induction of adverse effects. Therefore, the study was NMs. It is, however, important to keep in mind that ob- repeated with forced expression of IL-10 by injection of servation of protection by antioxidants does not specify adeno-associated virus vectors encoding murine IL-10. whether the effects occurred due to oxidative stress- Again, the placental and fetal effects of the injected NPs induced directly by particles or indirectly via other were reversed. Of note, inhibition of placental phagocyt- mechanistic pathways. In some studies, particle exposure osis and hence uptake of NPs significantly blocked IL-1β also induced pregnancy complications, such as fetal and IL-1α secretion, indicating that uptake of NPs into death, that could be associated with apoptosis and hence the cells might has been involved in inducing inflamma- generation of increased levels of ROS. Therefore, it is tory pathways in placental cells [16]. not possible to deduct whether the increases in ROS The induction of placental ROS by NMs was also ad- levels occurred due to particle exposure or pregnancy dressed by Qi et al. (2014) [94]. Pregnant mice were complications. injected i.v. with 20 mg of oxidized (o-) MWCNTs/kg on GD 17. Exposure increased the ROS levels in placentas, NM interference with placental toll-like receptors but not in maternal plasma, indicating that the placenta Several cell types express receptors for recognition of may respond more vigorously or faster to o-MWCNTs pathogen-associated molecular patterns present on the than other maternal tissues. The observation of placental surface of microorganisms. Probably the best-described implication in developmental toxicity has some resem- group of pattern recognition receptors are the TLRs, a blance to reports on the effects of SWCNTs, TiO2 and group of evolutionary conserved transmembrane pro- silica NPs [85, 90]. teins [118]. Until now, 11 mammalian TLRs have been Also, other studies have attempted to investigate the defined. TLR 4 is crucial for response to lipopolysac- degree to which oxidative stress contributes to develop- charide (LPS) and, thereby, to gram-negative bacteria. mental effects by administering antioxidants alongside TLR 2 recognizes a broader array of molecular patterns the maternal exposure to NMs [55]. Onoda and co- from bacteria and fungi. Ligand recognition by the TLRs workers investigated the protective effects of antioxi- mostly results in the activation of the intracellular sig- dants on the development of reactive astrogliosis in the naling pathway of NFκB, ultimately increasing the pro- offspring that had been observed following maternal in- duction of cytokines and antimicrobial factors [119]. tranasal instillation of CB NM (95 μg/kg body weight) The human placenta expresses all of the TLRs, varying on GD 5 and 9 in several previous studies. N-acetyl cyst- in a temporal and spatial manner [120]. Activation of eine or ascorbic acid were administered intraperitoneally trophoblast TLRs enhances cytokine expression, which to pregnant mice prior to CB instillation. N-acetyl cyst- may be followed by significant recruitment of immune eine partly prevented, whereas ascorbic acid slightly cells (macrophages, NK cells) to the placenta. TLR-
Dugershaw et al. Particle and Fibre Toxicology (2020) 17:31 Page 14 of 22
activation is associated with negative pregnancy outcomes 137]. First evidence that NMs can have endocrine-
(preterm labor, fetal loss and preeclampsia), but also plays disrupting activity came from studies in non-pregnant
a role in long-term adverse outcomes in the offspring, individuals, where NMs have been reported to affect
such as the function of the immune and central nervous levels of both female and male sex hormones in vitro
systems [119]. Placental TLRs may, however, also be in- and in vivo (reviewed in [138, 139]). For example, expos-
volved in the protective effects hypothesized to occur in ure of female and male rats to nickel (Ni) NPs by gavage
case of “adequate” non-infectious microbial exposure as resulted in altered hormone regulation (FSH and LH
proposed by the hygiene hypothesis [121]. levels were elevated and estradiol lowered in females
Accumulating evidence indicate that TLRs might while testosterone and FSH levels were diminished in
recognize some NMs and activate similar pathways as males) and induced pathological changes in testes and
upon contact with LPS and bacteria [122, 123]. Hence, ovaries (reviewed in [140]). However, it largely remains
MWCNTs have been shown to induce DNA damage in to be established if NMs might act as endocrine disrup-
human lung epithelial cells due to the activation of TLR tors during pregnancy and how this could potentially
9 and subsequent generation of nitric oxide (NO) [124]. affect pregnancy and offspring health later in life.
Also, SWCNTs have been reported to provoke In pregnancy, one of the critical hormones secreted by
chemokine secretion in macrophages via the TLR 2/4- the human placenta is human chorionic gonadotropin
MyD88-NFκB signaling pathway [125]. Interestingly, (hCG) [137]. It supports the function of the corpus
when graphene oxide was tested in the same setup, no luteum, a transient ovarian structure particularly import-
such response was elicited, indicating that TLRs may ant in the early gestational phase, which secretes ovarian
have a differential preference for subgroups of NMs progesterone and estrogens to maintain a successful
[125]. In silico investigations show that the internal pregnancy [141]. hCG also regulates the formation of
hydrophobic pockets of TLR 4 might be able to bind the ST [142, 143], modulates immune responses [143],
small-sized carbon nanostructures such as fullerenes and ensures uterine quiescence [143], promotes angiogenesis
CNTs [126]. TLR 4 has, however, been shown to also of the endometrial spiral arteries [143, 144], and dilates
recognize non-carbonaceous NMs, such as iron and TiO2 these vessels to enhance maternal blood flow [145]. Due
NPs, to promote inflammatory responses [127–129]. to these various crucial functions of hCG, disturbances
In the human placenta, TLR 2 and TLR 4 have been ob- in the tightly regulated levels of this hormone could,
served to lack in the ST but to be expressed in villous and therefore, increase the risk of adverse pregnancy out-
extravillous trophoblasts, at least during early pregnancy comes [146]. A few in vitro studies using BeWo tropho-
[119]. This could indicate that, at this stage, the placenta blast monocultures [103], 3D placental co-culture
responds primarily to pathogen-associated molecular pat- microtissues (BeWo cells/primary human villous mesen-
terns if the ligand has broken through the outer layer chymal fibroblasts) [98] or first trimester human placen-
[119]. Therefore, NMs would need to be internalized by tal explants [99] showed a significant reduction of hCG
the trophoblast for TLR activation. Interestingly, several release after exposure to GO, CdTe and CuO NPs or
studies in the ex vivo human placenta model and experi- dendritic polyglycerol NPs, respectively. This emphasizes
mental animals report that nanosized particles accumulate that disturbances in hCG release should be considered
in placental tissue [130–132] and that particles can be in developmental toxicity studies.
visualized in trophoblasts [90, 91, 133–135]. Also, the steroid hormones estrogen (reviewed in
Activation of placental TLRs by NMs would implicate [147]) and progesterone (reviewed in [148]) are indis-
the presence of NMs in maternal blood and their pensable to maintain human pregnancy. Estrogens are
uptake/penetration into the ST. Some TLRs do also essential for vasodilation and local angiogenesis due to
respond to endogenous molecules via so-called danger- their close interaction with angiogenic factors like vascu-
associated molecular patterns, including, but not lar endothelial growth factor (VEGF) and placental
restricted to, ROS and proteins released from dead or growth factor (PLGF) (reviewed in [147]). Dysregulation
dying cells [119]. Hence induction of ROS or inflamma- of estrogen secretion could, therefore, play a major role
tion by NMs in placental tissue, via direct or indirect in the development of preeclampsia and other adverse
pathways, may indirectly activate TLRs. conditions during pregnancy. Progesterone is essential
for the reproductive process. Altered progesterone secre-
NM interference with endocrine signaling tion has been associated with miscarriage and preterm
Endocrine signaling pathways are central in mediating birth [148]. So far, only a few descriptive studies report-
physiological and metabolic adaptations required for a ing NM effects on steroid hormone levels in pregnant
successful pregnancy and are orchestrated by the pla- animals are available. Inhalation of Ag NPs decreased es-
centa and the maternal endocrine organs (e.g., the pituit- trogen plasma levels in pregnant mice, but it was unclear
ary, thyroid and adrenal glands, and the ovaries) [136, if the Ag NP exposure caused the increase in observedYou can also read

















































