Targeting the Limitless Replicative Potential of Cancer: The Telomerase/Telomere Pathway
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Molecular Pathways
Targeting the Limitless Replicative Potential of Cancer:
The Telomerase/Telomere Pathway
Lloyd Kelland
Abstract The maintenance of telomeric DNA underlies the ability of tumors to possess unlimited replicative
potential, one of the hallmarks of cancer. Telomere length and structure are maintained by the
reverse transcriptase telomerase and a multiprotein telomere complex termed shelterin. Telomer-
ase activity is elevated in the vast majority of tumors, and telomeres are critically shortened in
tumors versus normal tissues, thus providing a compelling rationale to target the telomerase/
telomere pathway for broad-spectrum cancer therapy. This strategy is supported by a variety of
genetic-based target validation studies. Both telomerase inhibitors and telomere interactive mol-
ecules have shown stand-alone antitumor activity at nontoxic doses against a variety of human
tumor xenografts in mice. These translational advances have resulted in the first antitelomerase
agent, the oligonucleotide-based GRN163L targeting the telomerase RNA template, entering
clinical evaluation. Additional translational approaches, such as targeting telomeres using
G-quadruplex ligands, should result in antitelomere agents, such as RHPS4, entering the clinic in
the near future. These prototype trials will be extremely informative in determining the role of the
telomerase/telomere pathway in clinical oncology and, moreover, whether drugs targeting the
unlimited replicative potential of cancer will find a place in cancer chemotherapy.
angiogenesis, migration, or invasion), and ultimately whether
Background
small-molecule or biological modulators provide a benefit to
Current cancer drug development is largely focused on cancer patients (2). Pioneering studies done over the past 2
combating the major phenotypical features of cancer [i.e., decades have shown that one of the above-described and to
uncontrolled growth (through inactivation of tumor suppres- date therapeutically untapped phenotypic hallmarks of cancer,
sors, activation of oncogenes, and evasion of apoptosis), unlimited replicative potential, is intimately related to the
unlimited replicative potential, formation of new blood vessels maintenance of telomeres, repetitive TTAGGG sequences of
(angiogenesis), and tissue invasion and metastatic spread; DNA located on the ends of human chromosomes. Whereas
ref. 1]. In recent years, this strategy has resulted in some notable mortal cells shorten their telomeres during each round of
success stories with new approved molecularly targeted drugs replication due to the ‘‘end-replication problem,’’ in tumor cells
that have made a significant effect in lengthening the survival of telomere length is stabilized mainly via reactivation of the
cancer sufferers (e.g., imatinib in chronic myelogenous reverse transcriptase termed telomerase, composed of a RNA
leukemia, trastuzumab in breast cancer, and bevacizumab in component (hTR or hTERC) and a catalytic protein (hTERT;
colorectal cancer; ref. 2). see ref. 3 for a recent review). Moreover, the introduction of
hTERT into normal human cells extends their life span (4) and
The Relevance of theTelomere Maintenance hTERT (along with two oncogenes, large T and H-ras)
expression results in the direct tumorigenic conversion of
Pathway to Cancer
normal human epithelial and fibroblast cells (5). Other studies,
Validation of a cancer target encompasses information on the however, suggest that telomerase may also play a role in
prevalence and role of the target or pathway in human cancer, maintaining telomeres in some normal human cells (6) and
whether modulation of the target or pathway in preclinical may possess additional roles to telomere maintenance, such
model systems results in a robust anticancer phenotype (e.g., as regulation of chromatin state and DNA damage responses,
growth inhibition, induction of apoptosis or prevention of as cells lacking hTERT possessed a diminished capacity for
DNA double-strand break repair and fragmented chromosomes
(7). In addition, in some tumors (particularly those of
neuroepithelial and mesenchymal origin), telomeres are main-
Author’s Affiliation: Head of Biology, Cancer ResearchTechnology Development
Laboratory, Wolfson Institute for Biomedical Research, University College London,
tained by an alternative lengthening of telomere mechanism
London, United Kingdom (8). In patients with glioblastoma multiforme tumors, alterna-
Received 2/20/07; revised 3/16/07; accepted 3/20/07. tive lengthening of telomere is associated with mutant TP53
Requests for reprints: Lloyd Kelland, Head of Biology, Cancer Research and a favorable prognosis (9); characteristics of alternative
Technology Development Laboratory, Wolfson Institute for Biomedical Research, lengthening of telomere cells are alternative lengthening of
University College London, London WC1E BBT, United Kingdom. Phone: 44-20-
76796949; E-mail: lkelland@ cancertechnology.com.
telomere – associated promyelocytic leukemia bodies and cir-
F 2007 American Association for Cancer Research. cular extrachromosomal telomeric DNA (telomeric t circles,
doi:10.1158/1078-0432.CCR-07-0422 which require Xrcc3 and Nbs1 for their production; ref. 10).
Clin Cancer Res 2007;13(17) September 1, 2007 4960 www.aacrjournals.org
Downloaded from clincancerres.aacrjournals.org on October 12, 2015. © 2007 American Association for
Cancer Research.Targeting theTelomerase/Telomere Pathway
Finally, a group (six to date) of telomere-associated proteins, resulted in growth inhibition but this occurred with a different
collectively termed shelterin, comprising TRF1 and TRF2 that lag time dependent on this initial telomere length (15). Such
bind to double-stranded telomeric DNA, POT1 that binds the studies supported the view that, following inhibition of
single-stranded 3¶ G-rich overhang, and three interconnecting telomerase, telomeres needed to be gradually eroded down to
proteins (TIN2, TPP1, and RAP1), acts to shape and safeguard a critical length before any phenotypic effect occurred. Hence,
telomeres (Fig. 1; refs. 11, 12). small-molecule mimics of this effect would be best suited to the
The interest in telomerase and telomeres in a cancer treatment of tumors possessing very short telomeres.
therapeutics context has emerged from two main observations: Subsequent studies, however, targeting components of
first, telomerase activity, typically using a PCR-based assay, telomerase or telomeres/shelterin have shown that much more
telomeric repeat amplification protocol, is elevated in the great rapid apoptosis or induction of senescence effects can occur.
majority (f85%) of tumor types in comparison with normal For example, antisense oligonucleotide-mediated inhibition of
tissues, and second, telomeres are generally shorter in tumors hTERT, but not hTERC, caused a rapid reduction in cell growth
than in corresponding normal tissues (see refs. 13, 14 for and induction of apoptosis without telomere shortening in
reviews). Furthermore, several studies provide genetic valida- human prostate cancer cells (16). Second, the introduction of
tion of telomerase as an anticancer target in that inactivation of mutant template telomerase RNA into human tumor cells
hTR and/or hTERT by dominant-negative mutants or antisense rapidly decreased cell viability and increased apoptosis,
strategies resulted in an inhibition of tumor cell proliferation. possibly via a telomere uncapping mechanism and triggering
For example, transfection of a dominant-negative mutant of a DNA damage response (17, 18). Third, ribozymes targeting
hTERT into cancer cell lines of differing initial telomere length murine telomerase RNA induced antitumor effects, including
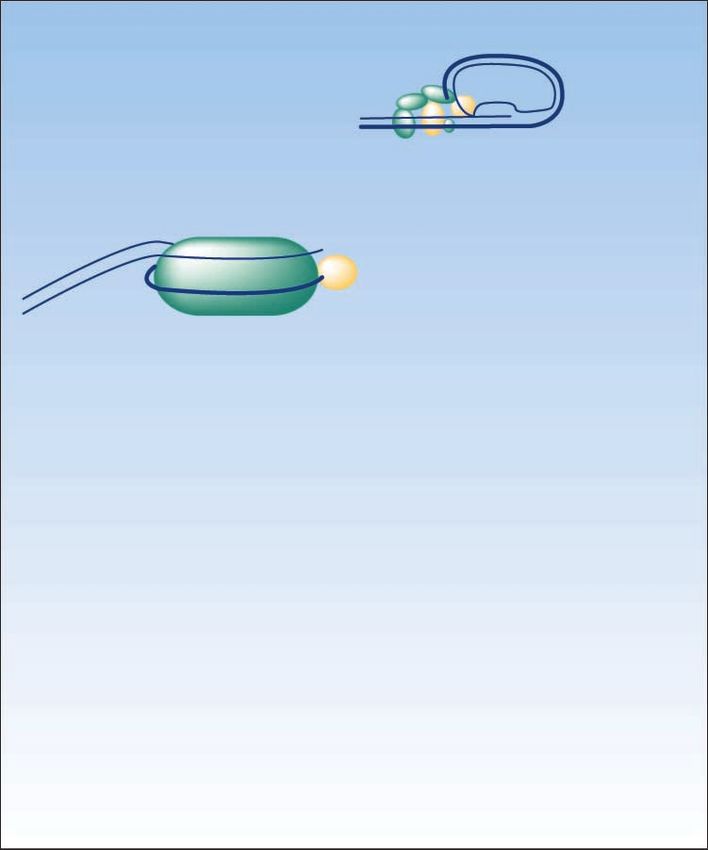
Fig. 1. The telomerase/telomere pathway
and main points of possible therapeutic
intervention. Pathway modulators may
result in either mainly telomere erosion
(e.g., BIBR1532) resulting in relatively
slow phenotypic anticancer effects or
predominantly telomere uncapping (e.g.,
RHPS4) producing more rapid anticancer
effects. Agents such as GRN163L probably
mainly affect telomere erosion but also seem
to induce telomere uncapping.
www.aacrjournals.org 4961 Clin Cancer Res 2007;13(17) September 1, 2007
Downloaded from clincancerres.aacrjournals.org on October 12, 2015. © 2007 American Association for
Cancer Research.Molecular Pathways
reducing melanoma tumor invasion and metastases in vivo; derivative of GRN163, a 13-mer thio-phosphoramidate oligo-
these studies also showed that telomerase activity may control nucleotide complementary to the RNA template of hTR (28).
the glycolytic pathway (19, 20). Finally, other genetic and Although cell-based studies with GRN163 generally showed a
biochemical studies suggest that targeting components of phenotypic lag of at least a few weeks between the onset of
shelterin, such as TRF2 (21) or POT1 (12, 22, 23), or exposing exposure and growth inhibition (e.g., ref. 28), the addition of a
the telomere 3¶ overhang (24) can induce apoptosis or 5¶ lipid (palmitate) moiety in GRN163L increased potency and
telomere-initiated senescence and thereby activate a DNA resulted in a more rapid loss of telomeres and inhibition of cell
damage checkpoint response (25). growth (29). Furthermore, in vivo inhibition of tumor growth
Because telomerase activity is relatively high in many tumors by GRN163L has been reported in mice in A549 human lung
and telomeres are generally shorter in tumors than normal cancer metastases treated with 15 mg/kg thrice weekly for 3
tissues, taken together with the above-described genetic-based consecutive weeks (30). Antitumor activity, observable from 2
validation studies, provide a compelling argument to suggest to 3 weeks of thrice-weekly dosing, has also been reported in
that the telomerase/telomere pathway is a well-validated target mice bearing orthotopic MDA-MB-231 human breast cancer
at the preclinical level. Therefore, appropriate modulators xenografts (31).
should be tested in patients. Over the past decade, many chemical classes of G-quadruplex
ligands have been described, the most potent of which,
Clinical Translational Advances telomestatin, inhibits telomerase in the telomeric repeat
amplification protocol assay at low nanomolar concentrations
As shown in Fig. 1, the burgeoning knowledge about the (32). Other compounds have been rationally designed based
structure of telomeres and the roles of various proteins involved on the crystal structure of parallel quadruplexes from human
in telomere maintenance provides several possible targets for telomeric DNA (33, 34). Several reduce the growth of various
modulation. Some of these, such as modulating the interaction cancer cell lines in vitro after 1 to 2 weeks of exposure to low
of TRF2 or POT1 with telomeres, may hold promise as cancer micromolar concentrations, often accompanied by cellular
drugs but, as yet, have not been described. To date, two areas senescence (34 – 37), apoptosis (32, 36), and/or chromosomal
have received major drug discovery attention: first, the targeting fusions/anaphase bridges (32, 35, 36). Several G-quadruplex
of components of telomerase (e.g., hTR RNA template or ligands have exhibited antitumor activity in vivo at nonacute
hTERT protein), and second, the targeting of telomeres (e.g., toxic doses in mice bearing various human tumor xenografts:
G-quadruplex ligands that induce the formation of guanine- the 3,6,9-trisubstituted acridine BRACO19 in combination with
rich telomeres to stably fold into four-stranded G-quadruplex paclitaxel against vulval carcinoma (34) or as monotherapy
structures, thereby adversely affecting telomere function). against DU145 prostate cancer (38) or UXF1138L uterine
Although, as described above, telomerase represents an cancer (39), the cationic porphyrin TMPyP4 against PC3
attractive cancer target, surprisingly few potent small-molecule prostate cancer (40), telomestatin against U937 acute myelo-
inhibitors have been discovered to date. It is unclear whether monocytic leukemia (41), and the pentacyclic acridine RHPS4
this is because of a general lack of interest in pursuing this against CG5 breast cancer (42). In general, antitumor effects
target within the pharmaceutical industry (perhaps because of were apparent relatively rapidly following the onset of drug
the phenotypic lag arguments described above) or because the treatment, typically after f10 days (34, 38 – 42). A consistent
enzyme is not particularly tractable to the main current drug mechanism of action schema is emerging involving displace-
discovery approach using high-throughput screening of large ment of POT1 and TRF2 from telomeres and erosion of 3¶
compound libraries resulting in few chemical hits or leads single-stranded telomeric overhangs (G-tails) with activation
emerging. One small-molecule nonnucleoside inhibitor of of a DNA damage response [e.g., reported with telomestatin
hTERT, BIBR1532, has been studied in preclinical models, (32, 43, 44) and RHPS4 (42)]. Such findings suggest that the
including in vivo (26, 27). BIBR1532 inhibited telomerase in G-quadruplex ligands can induce relatively rapid telomere
cell-free assays with an IC50 of 93 nmol/L and was shown to be uncapping and may provide an explanation as to why
a mixed-type noncompetitive inhibitor with a drug binding site reductions in telomere length are not always observed in cells
distinct from the sites for deoxyribonucleotides and the DNA exposed to this class of compound. The cell-based effects
primer. Cell-based studies with BIBR1532 revealed a significant (senescence, apoptosis, activation of a DNA damage response
phenotypic lag, typically of f100 days, between the onset of including gH2AX phosphorylation, etc.) are also consistent
telomerase inhibition and cellular growth arrest, hence with modulation of the currently known biological roles of
mimicking the above-described studies with dominant-negative POT1 (23) and TRF2 (21). Finally, measurements of POT1 or
hTERT. Antitumor effects were demonstrable in vivo in mice TRF2 displacement from telomeres or gH2AX phosphorylation
following inoculation of HT1080 fibrosarcoma cells that had may provide sensitive and appropriate biomarkers of target
been previously exposed to the compound for 100 days in vitro modulation in clinical trials. At least some of the G-quadruplex
(and leading to a reduction in telomere length to 1.6 kb). ligands, such as RHPS4, are expected to enter clinical trials in
An oligonucleotide-based molecule that acts as a telomerase the near future.
RNA template antagonist, GRN163L, represents the most
advanced antitelomerase therapeutic. Phase I clinical trials
Concluding Remarks
have been initiated in patients with either chronic lymphocytic
leukemia or solid tumors.1 GRN163L is a more potent The telomerase/telomere field has come a long way in a
relatively short time and is now at a particularly exciting
juncture, as the first telomerase inhibitor, GRN163L, undergoes
1
http://www.clinicaltrials.gov/ early clinical development. Other small-molecule modulators
Clin Cancer Res 2007;13(17) September 1, 2007 4962 www.aacrjournals.org
Downloaded from clincancerres.aacrjournals.org on October 12, 2015. © 2007 American Association for
Cancer Research.Targeting theTelomerase/Telomere Pathway
of telomerase/telomere biology, notably G-quadruplex ligands a broad-spectrum of cancers, most likely in combination with
such as RHPS4, are in preclinical development and are existing chemotherapeutic drugs. Although there is relatively
anticipated to enter the clinic in the near future. These clinical little to guide rational clinical combinations of antitelomerase/
studies will be extremely informative in determining whether telomere therapeutics with other drugs at present (and
the positive anticancer findings from various genetic-based although dose-limiting toxicities remain unknown), the asso-
preclinical validation studies are borne out in cancer patients. ciation of telomerase/telomeres with DNA repair, particularly
Furthermore, based on the high prevalence of elevated double-strand break repair, pathways is suggestive of this new
telomerase and/or shortened telomeres in multiple tumor types class of agent being usefully combined with cytotoxics that
in comparison with normal tissue counterparts, targeting induce DNA damage (e.g., platins and topoisomerase inhib-
telomerase/telomeres could be applicable to the treatment of itors) or ionizing radiation.
References
1. Hanahan D, Weinberg RA. The hallmarks of cancer. tumor cell proliferation. Proc Natl Acad Sci U S A phase bridge formation accompanied by loss of the 3¶
Cell 2000;100:57 ^ 70. 2001;98:7982 ^ 7. telomeric overhang in cancer cells. Oncogene 2006;
2. Benson JD, ChenY-NP, Cornell-Kennon SA, et al. Val- 18. Goldkorn A, Blackburn EH. Assembly of mutant- 25:1955 ^ 66.
idating cancer drug targets. Nature 2006;441:451 ^ 6. template telomerase RNA into catalytically active telo- 33. Parkinson GN, Lee MPH, Neidle S. Crystal structure
3. Blackburn EH, Greider CW, Szostak JW. Telomeres merase ribonucleoprotein that can act on telomeres is of parallel quadruplexes from human telomeric DNA.
and telomerase: the path from maize, tetrahymena required for apoptosis and cell cycle arrest in human Nature 2002;417:876 ^ 80.
and yeast to human cancer and aging. Nat Med cancer cells. Cancer Res 2006;66:5763 ^ 71. 34. Gowan SM, Harrison JR, Patterson L, et al. A
2006;12:1133 ^ 8. 19. Nosrati M, Li S, Bagheri S, et al. Antitumor activity G-quadruplex-interactive potent small-molecule inhib-
4. Bodnar AG, Ouellette M, Frolkis M, et al. Extension of of systemically delivered ribozymes targeting murine itor of telomerase exhibiting in vitro and in vivo anti-
life-span by introduction of telomerase into normal telomerase RNA. Clin Cancer Res 2004;10:4983 ^ 90. tumour activity. Mol Pharmacol 2002;61:1154 ^ 62.
human cells. Science 1998;279:349 ^ 52. 20. Bagheri S, Nosrati M, Li S, et al. Genes and path- 35. Incles CM, Schultes CM, Kempski H, Koehler H,
5. Hahn WC, Counter CM, Lundberg AS, Beijersbergen ways downstream of telomerase in melanoma metas- Kelland LR, Neidle S. A G-quadruplex telomere target-
RL, Brooks MW, Weinberg RA. Creation of human tasis. Proc Natl Acad Sci U S A 2006;103:11306 ^ 11. ing agent produces p16-associated senescence and
tumour cells with defined genetic elements. Nature 21.Van Steensel B, Smogorzewska A, de Lange T.TRF2 chromosomal fusions in human prostate cancer cells.
1999;400:464 ^ 8. protects human telomeres from end to end fusions. Mol CancerTher 2004;3:1201 ^ 6.
6. Masutomi K, Yu EY, Khurts S, et al. Telomerase Cell 1998;92:401 ^ 13. 36. Leonetti C, Amodei S, D’Angelo C, et al. Biological
maintains telomere structure in normal human cells. 22. Zaug AJ, Podell ER, CechTR. Human POT1disrupts activity of the G-quadruplex ligand RHPS4 (3,11-
Cell 2003;114:241 ^ 53. telomeric G-quadruplexes allowing telomerase exten- difluoro-6,8,13-trimethyl-8H-quino[4,3,2-kl] acridi-
7. Masutomi K, Possemato R,Wong JMY, et al.The telo- sion in vitro. Proc Natl Acad Sci U S A 2005;102: nium methosulfate) is associated with telomere capping
merase reverse transcriptase regulates chromatin state 10864 ^ 9. alteration. Mol Pharmacol 2004;66:1138 ^ 46.
and DNA damage responses. Proc Natl Acad Sci 23. Churikov D, Wei C, Price CM. Vertebrate POT1 37. Cookson JC, Dai F, SmithV, et al. Pharmacodynam-
U S A 2005;102:8222 ^ 7. restricts G-overhang length and prevents activation ics of the G-quadruplex-stabilizing telomerase inhibi-
8. Dunham MA, Neumann AA, Fasching CL, Reddel of a telomeric DNA damage checkpoint but is dis- tor 3,11-difluoro-6,8,13-trimethyl-8H-quino[4,3,2-kl]
RR. Telomere maintenance by recombination in human pensable for overhang protection. Mol Cell Biol acridinium methosulfate (RHPS4) in vitro : activity in
cells. Nat Genet 2000;26:447 ^ 50. 2006;26:6971 ^ 82. human tumor cells correlates with telomere length
9. Chen Y-J, Hakin-Smith V,Teo M, et al. Association of 24. Li G-Z, Eller MS, Firoozabadi R, Gilchrest BA. and can be enhanced, or antagonized, with cytotoxic
mutantTP53 with alternative lengthening of telomeres Evidence that exposure of the telomere 3¶ overhang se- agents. Mol Pharmacol 2005;68:1551 ^ 8.
and favorable prognosis in glioma. Cancer Res 2006; quence induces senescence. Proc Natl Acad Sci U S A 38. Kelland LR. Overcoming the immortality of tumour
66:6473 ^ 6. 2003;100:527 ^ 31. cells by telomere and telomerase based cancer thera-
10. Compton SA, Choi J-H, Cesare AJ, Ozgur S, 25. di Fagagna FD, Reaper PM, Clay-Farrace L, et al. peuticsJcurrent status and future prospects. Eur J
Griffith JD. Xrcc3 and Nbs1 are required for the pro- A DNA damage checkpoint response in telomere- Cancer 2005;41:971 ^ 9.
duction of extrachromosomal telomeric circles in initiated senescence. Nature 2003;426:194 ^ 8. 39. Burger AM, Dai F, Schultes CM, et al. The G-quad-
human alternative lengthening of telomere cells. 26. Damm K, Hemmann U, Garin-Chesa P, et al. A ruplex-interactive molecule BRACO-19 inhibits tumor
Cancer Res 2007;67:1513 ^ 9. highly selective telomerase inhibitor limiting human growth, consistent with telomere targeting and inter-
11. de lange T. Shelterin: the protein complex that cancer cell proliferation. EMBO J 2001;20:6958 ^ 68. ference with telomerase function. Cancer Res 2005;
shapes and safeguards human telomeres. Genes Dev 27. Pascolo E, Wenz C, Lingner J, et al. Mechanism of 65:1489 ^ 96.
2005;19:2100 ^ 10. human telomerase inhibition by BIBR1532, a synthetic, 40. Grand CL, Han H, Munoz RM, et al. The cationic
12. Wang F, Podell ER, Zaug AJ, et al. The POT1-TPP1 non-nucleoside drug candidate. J Biol Chem 2002; porphyrin TMPyP4 down-regulates c-MYC and
telomere complex is a telomerase processivity factor. 277:15566 ^ 72. human telomerase reverse transcriptase expression
Nature 2007;445:506 ^ 10. 28. Akiyama M, HideshimaT, Shammas MA, et al. Effects and inhibits tumor growth in vivo. Mol Cancer Ther
13. Shay JW. Meeting report: the role of telomeres and of oligonucleotide N3¶-P5¶ thio-phosphoramidate 2002;1:565 ^ 73.
telomerase in cancer. Cancer Res 2005;65:3513 ^ 7. (GRN163) targeting telomerase RNA in human multiple 41. Tauchi T, Shin-ya K, Sashida G, et al. Telomerase inhi-
14. Shay JW, Wright WE. Telomerase therapeutics for myeloma cells. Cancer Res 2003;63:6187 ^ 94. bition with a novel G-quadruplex interactive agent,
cancer: challenges and new directions. Nat Rev Drug 29. Herbert B-S, Gellert GC, Hochreiter A, et al. Lipid telomestatin: in vitro and in vivo studies in acute
Discov 2006;5:577 ^ 83. modification of GRN163, an N3¶-P5¶ thio-phosphora- leukemia. Oncogene 2006;25:5719 ^ 25.
15. Hahn WC, Stewart SA, Brooks MW, et al. Inhibition midate oligonucleotide, enhances the potency of telo- 42. Salvati E, Leonetti C, Rizzo A, et al. Telomere damage
of telomerase limits the growth of human cancer cells. merase inhibition. Oncogene 2005;24:5262 ^ 8. promotes antitumoral activity of the G-Quadruplex
Nat Med 1999;5:1164 ^ 70. 30. Dikmen ZG, Gellert GC, Jackson S, et al. In vivo ligand RHPS4. J Clin Invest. In press 2007.
16. Folini M, Brambilla C, Villa R, et al. Antisense inhibition of lung cancer by GRN163L: a novel human 43. Gomez D, Wenner T, Brassart B, et al. Telomestatin-
oligonucleotide-mediated inhibition of hTERT, but telomerase inhibitor. Cancer Res 2005;65:7866 ^ 73. induced telomere uncapping is modulated by POT1
not hTERC, induces rapid cell growth decline and 31. Hochreiter AE, Xiao H, Goldblatt EM, et al. Telomer- through G-overhang extension in HT1080 human tumor
apoptosis in the absence of telomere shortening in ase template antagonist GRN163L disrupts telomere cells. J Biol Chem 2006;281:38721 ^ 9.
human prostate cancer cells. Eur J Cancer 2005;41: maintenance, tumor growth, and metastasis of breast 44. Gomez D, O’Donohue M-F, Wenner T, et al. The
624 ^ 34. cancer. Clin Cancer Res 2006;12:3184 ^ 92. G-quadruplex ligand telomestatin inhibits POT1 bind-
17. Kim MM, Rivera MA, Botchkina IL, Shalaby R, Thor 32. Tahara H, Shin-ya K, Seimiya H,Yamada H,TsuruoT, ing to telomeric sequences in vitro and induces GFP-
AD, Blackburn EH. A low threshold level of expression IdeT. G-quadruplex stabilization by telomestatin indu- POT1 dissociation from telomeres in human cells.
of mutant-template telomerase RNA inhibits human cesTRF2 protein dissociation from telomeres and ana- Cancer Res 2006;66:6908 ^ 12.
www.aacrjournals.org 4963 Clin Cancer Res 2007;13(17) September 1, 2007
Downloaded from clincancerres.aacrjournals.org on October 12, 2015. © 2007 American Association for
Cancer Research.Targeting the Limitless Replicative Potential of Cancer: The
Telomerase/Telomere Pathway
Lloyd Kelland
Clin Cancer Res 2007;13:4960-4963.
Updated version Access the most recent version of this article at:
http://clincancerres.aacrjournals.org/content/13/17/4960
Cited articles This article cites 43 articles, 26 of which you can access for free at:
http://clincancerres.aacrjournals.org/content/13/17/4960.full.html#ref-list-1
Citing articles This article has been cited by 14 HighWire-hosted articles. Access the articles at:
http://clincancerres.aacrjournals.org/content/13/17/4960.full.html#related-urls
E-mail alerts Sign up to receive free email-alerts related to this article or journal.
Reprints and To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Subscriptions Department at pubs@aacr.org.
Permissions To request permission to re-use all or part of this article, contact the AACR Publications
Department at permissions@aacr.org.
Downloaded from clincancerres.aacrjournals.org on October 12, 2015. © 2007 American Association for
Cancer Research.You can also read

















































