The jaw apparatus of the Late Cretaceous heteromorph ammonoid Turrilites costatus from central Hokkaido, Japan
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Bulletin of the Mikasa City Museum, 2021, (24), p. 1–8
ISSN 1344-2295
Article 原著
The jaw apparatus of the Late Cretaceous heteromorph ammonoid
Turrilites costatus from central Hokkaido, Japan
Kazushige Tanabe1*, Daisuke Aiba2 and Junya Abe3
1
The University Museum, The University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan. E-mail: kazu-tanabe@g.ecc.u-tokyo.ac.jp
2
Mikasa City Museum, 1-212-1 Ikushumbetsu Nishiki-cho, Mikasa City, Hokkaido 068-2111, Japan. E-mail: aiba698@city.mikasa.hokkaido.jp
3
1-1 Nijigaoka, Kitahiroshima City, Hokkaido 061-1103, Japan. E-mail: abejunya@gmail.com
北海道産後期白亜紀異常巻きアンモノイド Turrilites costatus の顎器
棚部一成.東京大学総合研究博物館 , 113-0033 東京都文京区本郷 7-3-1
相場大佑.三笠市立博物館 , 068-2111 北海道三笠市幾春別錦町 1 丁目 212-1
阿部純也.北海道北広島市虹ヶ丘 1-1
Abstract.—A specimen of the heteromorph ammonoid Turrilites costatus Lamarck, 1801 (Turrilitidae, Turrilitoidea)
with upper and lower jaws preserved within the body chamber in situ is newly described from the middle Cenomanian
(Upper Cretaceous) strata of the Yezo Group in the Ashibetsu area, central Hokkaido, Japan. The upper and lower jaws
of this species are almost equal-sized, both with a pointed rostrum. They are similar in overall morphology to the jaws of
nostoceratids and diplomoceratids of the Turrilitoidea, but differ from those of baculitids of the same superfamily by having
a more projected rostrum and thinner paired outer calcitic layers in the lower jaw. The beak-like upper and lower jaws of this
species suggest that they served for grasping and cutting up prey.
Key Words.—Turrilites costatus, Turrilitidae, jaw apparatus, Cenomanian, Hokkaido
Received.—2 October, 2020 / Accepted.—14 October, 2020
*Corresponding author
Cretaceous Ancyloceratina, we newly collected a specimen of
Introduction Turrilites costatus Lamarck, 1801 (Turrilitidae, Turrilitoidea)
with upper and lower jaws in situ from the Yezo Group in central
Modern and fossil cephalopod mollusks possess a well- Hokkaido, Japan. This is the first discovery of the jaw apparatus
developed jaw apparatus (synonymous with beaks and from this family. In this paper, we describe the jaw apparatus of
mandibles) as a primary feeding organ (Clarke, 1986; Nixon, this species and compare to those of other species of the Late
1988, 2015; Tanabe and Fukuda, 1999; Tanabe, 2012; Tanabe et Cretaceous Turrilitoidea with discussing its taxonomic and
al., 2015a). In modern cephalopods, the jaw apparatus consists paleoecological implications.
of upper and lower elements that are made of chitin-protein
complex, in association with an anterior calcified covering for Material and geological settings
nautilids (Saunders et al., 1978; Hunt and Nixon, 1981). The
upper and lower jaws are accommodated together with a radula A specimen of Turrilites costatus with a jaw apparatus in
in a globular muscular organ called the buccal mass (see Tanabe situ was examined in this study. It was found in a calcareous
and Fukuda, 1987, fig. 1; Tanabe, 2012, fig. 1). siltstone slab, which was recovered by one of us (JA) as float at
Fossilized chitinous and calcareous remains of the jaw a locality in the western tributary of the Nokanan-gawa River on
apparatus are occasionally preserved in situ within the body the northern side of the Maruyama Dam, Ashibetsu area, central
chambers of ammonoids whose taxonomic relationships are Hokkaido (43°26′41.21″N, 142°13′40.02″E; Fig. 1A). In the area
known (Tanabe et al., 2015a). In Cretaceous heteromorph near the Maruyama Dam, siliciclastic deposits of the Cretaceous
ammonoids of the suborder Ancyloceratina, in situ upper and Yezo Group are widely distributed, and they have been
lower jaws have been reported from 32 species of 20 genera that lithostratigraphically divided into the Hikagenosawa and Saku
are distributed in seven families within the three superfamilies, Formations toward the upward sequence (Fig. 1B; Hashimoto
Ancyloceratoidea, Turrilitoidea, and Scaphitoidea (see Tanabe et et al., 1953; Takashima et al., 2004). The Hikagenosawa
al., 2015a, table 10.1). Formation consists mainly of massive and/or weakly laminated
In the process of searching additional jaw material from the siltstone, whereas the Saku Formation comprises siltstone and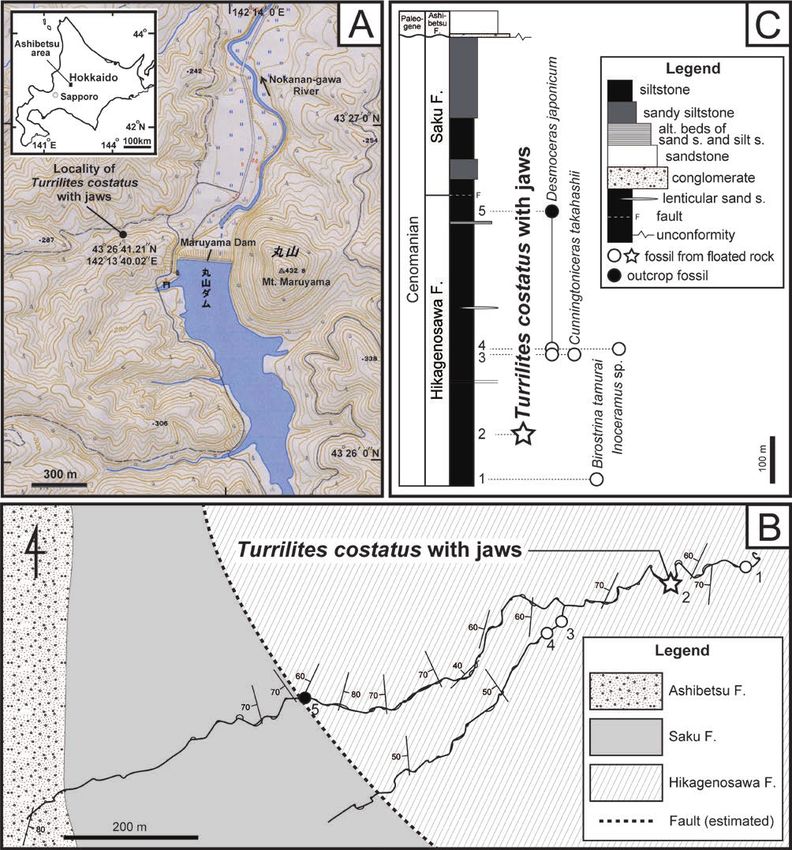
2 Kazushige Tanabe, Daisuke Aiba and Junya Abe
sandy siltstone in the lower part and sandy siltstone in the upper outcrops (Fig. 1B; see also Takashima et al., 2004, fig. 4). We
part (Fig. 1C; Motoyama et al., 1991; Kawabe et al., 2003; collected macrofossils identified as ammonites Desmoceras
Takashima et al., 2004). Geological investigations near the japonicum and Cunningtoniceras takahashii and inoceramid
fossil locality by one of us (DA) suggest that the siltstone slab bivalves Biostrina tamurai and Inoceramus sp. in situ or as float
yielded the T. costatus specimen came from a nearby outcrop from the Hikagenosawa Formation near the fossil locality (Fig.
of the Hikagenosawa Formation of the Yezo Group, because of 1C). The T. costatus specimen examined is dated as the middle
the lithological similarity between the siltstone slab and nearby Cenomanian, because C. takahashii and B. tamurai are zonal
Fig. 1. Topographic map (A) of the southern Ashibetsu area, central Hokkaido, and geological map (B) and geological column (C) of the Yezo
Group near the locality of the Turrilites costatus specimen. Localities and horizons of the T. costatus specimen and other float and in situ
mollusk fossils are respectively shown as a star mark, and white and solid circles with numbers in B and C. Topographic map around the
Maruyama Lake (A) is reproduced from the website of the Geographical Survey of Japan (https ://maps.gsi.go.jp).
Bulletin of the Mikasa City Museum, (24)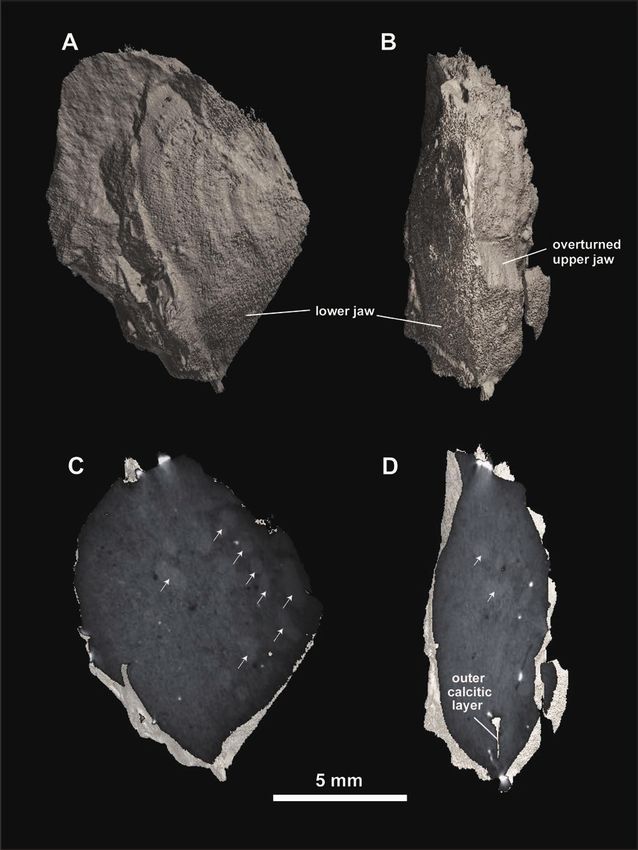
The Jaw Apparatus of Cretaceous Ammonoid Turrilites costatus 3
indices of this substage in Japan (Toshimitsu et al., 1995, table 1). wall of upper jaw, LHL: maximum length of hood (= length of
symphysis) of lower jaw, LWL: maximum length of wing of
Methods lower jaw.
The internal cast and external molds of the jaw remains Morphology of the jaw apparatus. – The upper jaw is
of the T. costatus specimen were photographed by a digital composed entirely of an undetermined, black material which
camera (Nikon model D3200) and measured by a slide caliper appears to have been primarily chitinous, as in the upper jaws
(accuracy + 0.1 mm). The external mold of the lower jaw was of extant cephalopods. It consists of a shorter hood portion and
also photographed by a digital microscope (KEYENCE model a pair of larger, posteriorly elongated wing-like lateral walls,
VHX-600) at the University Museum of the University of Tokyo the latter of which measured 10.6 mm in LLU and 5.7 mm in
(UMUT). Internal structure of the removed mouthpart portion HLU. The left lateral wall is mostly embedded in the mudstone
was analyzed by means of an X-ray CT scanning machine, matrix (Fig. 2C). The short hood portion is joined with the
ScanXmate–B100TSS110 (Comscan Co.) at UMUT. The tube paired lateral walls; accordingly, the upper jaw is composed of
voltage and tube current were 100 kV, and 98 μA respectively. A a single chitinous lamella. The paired lateral walls and hood
total of 2059 X-ray images sliced along the horizontal, sagittal become narrower anteriorly and join together, forming a sharply
and cross-sectional planes of the jaws were obtained. X-ray pointed rostral tip. They are sculptured by evenly spaced,
tomography of the jaws was reconstructed from the scanned files strong concentric undulations, in association with fine striations
by an image analyzing software Molcer Plus Ver. 1.35 (White between them (Fig. 2C, D). These concentric ornamentations
Rabbit Co.). are interpreted as growth lines, because their strength tends to
The specimen examined is housed in the Mikasa City increase posteriorly.
Museum (MCM) with the registered number, MCM-A2153. The lower jaw is characterized by a widely open outer
lamella with a weakly pointed rostral tip, which is divided into
Description two wing portions by a prominent radial furrow (symphysis).
The left wing portion is mostly embedded in the mudstone
Recognition of upper and lower jaws. – Although the early matrix and could not be observed from the outside (Fig. 2B).
shell portion is missing in MCM-A2153, the preserved conch The outer lamella, 9.4 mm in LHL and 9.8 mm in LWL, is
measures 16.7 mm in shell height and 13.8 mm in maximum composed of an inner black-colored “chitinous” layer (Fig. 2E)
diameter. The jaw apparatus is preserved inside the body and approximately 0.1 mm thick, very thin outer paired calcitic
chamber (Fig. 2A) and consists of two elements whose lateral layers (aptychus sensu stricto) (Fig. 2G); the right half of the
sides are attached to each other (Fig. 2B). Both elements outer calcific layer is preserved in the external mold slab (Fig.
appear to have been secondarily deformed by compression 2F). A fragmented outer calcific layer on the left lateral side was
from the lateral side during sediment compaction. In spite of detected by a CT scan X-ray image taken from the dorsoventral
such incomplete preservation, upper and lower jaws can be side (Fig. 4D). A shortly reduced inner chitinous lamella,
recognized through comparative morphological examination which has been recognized in the lower jaws of Cretaceous
of the jaw apparatuses of modern and fossil cephalopods Ancyloceratina (e.g., Tanabe and Landman, 2002; Tanabe et
(Clarke, 1986; Nixon, 1988; Tanabe and Fukuda, 1999; Tanabe, al., 2015a), could not be observed from the outside, but it may
2012). The element having a slightly concave outer lamella present on the dorsal side of the outer lamella. The paired wing
with a prominent radial furrow (Fig. 2B, E–F) is identified as portions are ornamented with prominent concentric undulations,
the lower jaw, because these features are commonly observed in association with fine striations between them (Fig. 2E, F). The
in the aptychus-type lower jaws of Jurassic and Cretaceous concentric undulations and striations are interpreted as growth
Ammonitina and Cretaceous Ancyloceratina (Lehmann, 1990; lines, because their strength increases posteriorly.
Tanabe et al., 2015a). The other element, which consists of
posteriorly elongated, paired lamellae with a sharply pointed Biostratinomy of the jaw apparatus. – The mode of occurrence
anterior tip (Fig. 2A, C–D), is similar to the upper jaws of of the jaw apparatus shows that the upper jaw is preserved
previously known Mesozoic ammonoids; hence it is identified as upside down with respect to the orientation of the lower jaw (Figs.
an upper jaw. 2B, 4A, B). Our CT scan examination revealed that the space
between upper and lower jaws is filled with many elongated
Terminology and measurements. – Basic morphology and fecal pellets, each 1 to 2 mm in greater diameter (pointed by
measurements of the reconstructed jaw apparatus of T. costatus arrows in Fig. 4C, D). This evidence suggests that the upper and
are shown in Fig. 3, in which descriptive terms of modern lower jaws have been shifted from their life orientations by the
coleoid jaws proposed by Clarke (1986) are used. Morphological foraging activity of benthic organisms during the post-mortem
terms for the aptychus-type lower jaws proposed by Arkell (1957) decay and subsequent biostratinomic process of the ammonite
are also shown in parentheses in this figure. The following carcass.
abbreviations are used for measurements. LLU: maximum length
of lateral wall of upper jaw, HLU: maximum height of lateral
March 31, 20214 Kazushige Tanabe, Daisuke Aiba and Junya Abe Bulletin of the Mikasa City Museum, (24)
The Jaw Apparatus of Cretaceous Ammonoid Turrilites costatus 5
variation in the Cretaceous baculitids.
Discussion
Functional morphology of the jaws of Turrilites costatus.
Comparison with the jaws of other Late Cretaceous – The almost equal-sized upper and lower jaws with a beak-
Turrilitoidea. – As mentioned above, the jaw apparatus of like rostral tip in T. costatus might be effective for biting and
Turrilites costatus is made of almost equal-sized upper and cutting up prey, as postulated for the jaws of the Late Cretaceous
lower jaws, and the lower jaw is characterized by a posteriorly diplomoceratids (Tanabe et al., 1980; Tanabe and Landman,
elongated outer lamella with a weakly pointed rostral tip, 2002; Tanabe, 2011) and nostoceratids (Kruta et al., 2010;
consisting of an inner chitinous layer and thin outer bivalved Tanabe et al., 2015a). The presence of a radial furrow (symphysis)
calcareous layers, divided by a radial furrow (symphysis) on the chitinous outer lamella suggests that this species possibly
along the hood portion. These features are commonly observed could temporally open and expose the outer lamella of the lower
in the aptychus-type jaw apparatuses of other species of the jaw outside of a buccal mass for feeding (Tanabe et al., 2015a,
Late Cretaceous superfamily Turrilitoidea, such as Scalarites fig. 10.11c). However, an operculum-like secondary function
mihoensis Wright and Matsumoto, 1954 (Tanabe et al., 1980, hypothesized by Lehmann and Kulicki (1990) and Seilacher
fig. 1; pl. 1, figs. a–f), Polyptychoceras cf. pseudogaultinum (1993) in the aptychus-type lower jaws with thick and strongly
(Yokoyama, 1890) (Tanabe and Landman, 2002, pl. 1, fig. ornamented paired calcitic plates in some Jurassic Ammonitina
6), P. sp. (Tanabe, 2011), and Subptychoceras sp. (Tanabe is unlikely for the lower jaw of the present species, because the
and Landman, 2002, text-fig. 2–6a, b; pl. 1, figs. 7a–c) of outer paired calcified layers are so thin to protect the soft body
the family Diplomoceratidae, and Didymoceras nebrascense from predator’s attack.
(Meek and Hayden, 1856) (Kruta et al., 2010, figs. 1–5, 7–9), On the other hand, the posteriorly elongated lower jaws
Nostoceras sp. (lower jaw only; Tanabe et al., 2015a, fig. of Baculites without a beak-like posterior tip are supposed to
10.7e) and Pravitoceras sigmoidale Yabe, 1902 (Tanabe et al., had a secondary function as opercula. However, Larson and
2015b, figs. 3-4) of the family Nostoceratidae. Accordingly, the Landman (2017) rejected this possibility based on the misfit of
aforementioned jaw features are shared by the Diplomoceratidae, the measurements of the jaws and the shape of the corresponding
Nostoceratidae, and Turrilitidae, and can be used as diagnostic shells. Instead, these authors suggested that the enlarged lower
taxonomic characters of these families. jaws of Baculites have been developed to collect and convey
In the family Baculitidae of the Turrilitoidea, an aptychus- food, in combination of the upper jaws and radula. Their
type jaw apparatus has been found in the species of Sciponoceras interpretation is consistent with the discovery of zooplankton
(Kaplan et al., 1998; Tanabe and Landman, 2002) and Baculites (isopods and larval gastropods) prey remains in the buccal cavity
(Landman et al., 2007; Kruta et al., 2009, 2011; Klug et al., of the body chamber of a Baculites shell, in combination with
2012; Larson and Landman, 2017). The lower jaws of these an upper jaw and radula by means of synchrotron X-ray analysis
baculitids differ from those of T. costatus, diplomoceratids, (Kruta et al., 2011, figs. 1–3). In summary, this and previous
and nostoceratids of the same superfamily by having a pair of works suggest that the wide variation in the overall shape,
posteriorly elongated calcitic layers without a rostral tip, which relative size and the degree of development of paired calcific
cover the underlying chitinous outer lamella. The upper jaws layers observed in the jaws of the Turrilitoidea presumably
of baculitids are rare in occurrence, and only two examples reflect the variety of dietary habits among different taxa.
have been reported to date; one is found in a specimen from the
Campanian Pierre Shale in South Dakota, USA (Kruta et al., Acknowledgments
2011, figs. 1C, 3A) and the other is preserved in specimens from
the Cenomanian–Turonian boundary horizon in northwestern We thank Yu Maekawa and Takenori Sasaki (both UMUT)
Germany (Klug et al., 2012, figs. 4A, B, 7–8). The upper jaw of for facilitating the use of an X-ray CT machine, and Neil H.
the former example is extremely reduced as compared with the Landman (American Museum of Natural History) for critical
lower jaw of a baculitid shell from the same locality (Kruta et comments to improve this manuscript.
al., 2011, figs. 1C), whereas those of the latter example are as
large as the co-occurred lower jaws (Klug et al., 2012, figs. 4A,
B, 7–8). These lines of evidence indicate that the relative size
of an upper jaw versus a lower jaw exhibits a wide interspecific
Fig. 2. Turrilites costatus, with upper and lower jaws preserved in the body chamber in situ. MCM-A2153. A. Mode of occurrence of the jaw
apparatus (only the upper jaw is exposed) in the body chamber, B. Frontal view of the jaw apparatus, C, D. Internal cast (C) and external mold
(D) of the upper jaw (right lateral view for C), E, F. Internal cast (E) and external mold (F) of the lower jaw (right lateral view for E). G. Cross
section of the outer calcitic layer (ocl) of the lower jaw at the hood portion. Outer surface of the inner chitinous lamella and inner surface of the
outer calcitic layer are respectively seen in E and F. White arrows in B and E point to the radial furrow (symphysis).
March 31, 20216 Kazushige Tanabe, Daisuke Aiba and Junya Abe
Fig. 3. Three-dimensional reconstruction and measurements of the jaw apparatus of T. costatus (anterolateral view). Descriptive terms are based
on those of modern coleoid jaws by Clarke (1986). Morphological terms of the aptychus-type lower jaw by Arkell (1957) are also given in
parentheses in this figure. See the text for abbreviations of measurements.
cephalopods. Comparative Biochemistry and Physiology,
References vol. 68B, p. 535–546.
Kaplan, U., Kennedy, W. J., Lehmann, J. and Marcinowski,
Arkell, W.J., 1957: Aptychi. In Moore, R. C. ed. Treatise R., 1998: Stratigraphie und Ammonitenfaunen des
on Invertebrate Paleontology, Part L, Mollusca 4, westfälischen Cenoman. Geologie und Paläontologie in
Cephalopoda, Ammonoidea. p. L437–L441.Geological Westfalen, vol. 51, p. 5–237.
Society of America and University of Kansas, Kansas. Kawabe, F., Takashima, R., Wani, R., Nishi, H. and Moriya, K.,
Clarke, M. R., 1986: A Handbook for the Identification of 2003: Upper Albian to Lower Cenomanian biostratigraphy
Cephalopod Beaks. 273 p. Clarendon Press, Oxford. in the Oyubari area, Hokkaido, Japan: toward a Cretaceous
Hashimoto, I., Tanaka, K. and Imai, I., 1953: Geological biochronology for the North Pacific. Acta Geologica
Sheet Map “Kami-Ashibetsu”, Scale 1:50,000 and its Polonica, vol. 53, p. 81–91.
Explanatory Text, 78 p. Geological Survey of Japan, Klug, C., Riegraf, W. and Lehmann, J., 2012: Soft-part
Tsukuba. (in Japanese with English summary) preservation in heteromorph ammonites from the
Hunt, S. and Nixon, M., 1981: A comparative study of protein Cenomanian-Turonian boundary event (OAE 2) in north-
composition in the chitin-protein complexes of the beak, west Germany. Palaeontology, vol. 55, p. 1307–1331.
pen, sucker disc, radula and oesophageal cuticle of Kruta, I., Landman, N. H., Rouget, I., Cecca, F. and Larson, N. L.,
Bulletin of the Mikasa City Museum, (24)The Jaw Apparatus of Cretaceous Ammonoid Turrilites costatus 7
Fig. 4. X-ray CT scan images of the jaw apparatus of T. costatus. A, B. X-ray tomographies of the jaw apparatus viewed from right lateral (A) and
frontal (B) sides, C, D. X-ray CT images of the inner portion of the jaw apparatus taken from right lateral (C) and frontal (D) sides. A piece
of the outer calcitic layer of the outer lamella in the lower jaw on the left lateral side is visible in D. Small fecal pellets produced by benthic
organisms in the silty mudstone matrix are pointed by white arrows in C and D.
March 31, 20218 Kazushige Tanabe, Daisuke Aiba and Junya Abe
2010: The Jaw apparatus of the Late Cretaceous ammonite Ando, H., 2004: Geology and stratigraphy of fore-arc basin
Didymoceras. Journal of Paleontology, vol. 84, p. 556– sediments in Hokkaido, Japan: Cretaceous environmental
560. events on the north-west Pacificmargin. Cretaceous
Kruta, I., Landman, N., Rouge,t I., Cecca, F. and Tafforeau, P., Research, vol. 25, p. 365–390.
2011: The role of ammonites in the Mesozoic marine food Tanabe, K., 2011: The feeding habits of ammonites. Science, vol.
web revealed by jaw preservation. Science, vol. 331, p. 331, p. 37–38.
70–72. Tanabe, K., 2012: Comparative morphology of modern and fossil
Kruta, I., Rouget, I., Landman, N. H., Tanabe, K. and Cecca, coleoid jaw apparatuses. Neues Jahrbuch für Geologie und
F., 2009: Aptychi microstructure in Late Cretaceous Paläontologie, Abhandlungen, vol. 266, p. 9–18.
Ancyloceratina (Ammonoidea). Lethaia, vol. 42, p. 312– Tanabe, K. and Fukuda, Y., 1987: Mouth part histology and
321 morphology. In Saunders, W. B. and Landman, N. H. eds.,
Lamarck, J. B. P. A. de M. de., 1801: Systéme des animaux sans Nautilus, the Biology and Paleobiology of a Living Fossil. p.
vertèbres. 432 p. Deterville, Paris. 313–322, Plenum Press, New York.
Landman, N. H., Larson, N. L. and Cobban, W. A., 2007: Tanabe, K. and Fukuda, Y., 1999: Morphology and function of
Jaws and radula of Baculites from the Upper Cretaceous cephalopod buccal mass. In Savazzi, E. ed., Functional
(Campanian) of North America. In Landman, N., Davis, Morphology of the Invertebrate Skeleton. p. 245–262,
R. and Mapes, R. H. eds., Cephalopods – Present and Wiley, London.
Past, New Insights and Fresh Perspectives, p. 257–298, Tanabe, K., Hirano, H. and Kanie, Y., 1980: The jaw apparatus
Springer, Dordrecht. of Scalarites mihoensis, a Late Cretaceous ammonite.
Larson, N. L. and Landman, N. H., 2017: Description of the In Igo, H. and Noda, H. eds., Professor Saburo Kanno
lower jaws of Baculites from the Upper Cretaceous U.S. Memorial Volume, p. 159–165, The University of Tsukuba,
Western Interior. Acta Geologica Polonoica, vol. 67, p. Tsukuba.
109–120. Tanabe, K., Kruta, I. and Landman, N. H., 2015a: Chapter 10.
Lehmann, U., 1990: Ammonoideen. 257 p. Enke, Stuttgart. Ammonoid buccal mass and jaw apparatus. In, Klug, C.
Lehmann, U. and Kulicki, C., 1990: Double function of aptychi Korn, D., De Beats, K. Kruta, I. and Mapes, R. H. eds.,
(Ammonoidea) as jaw elements and opercula. Lethaia, vol. Ammonoid Paleobiology: From Anatomy to Ecology.
23, p. 325–331 Topics in Geobiology 43. p. 429–484, Springer, Dordrecht.
Meek, F. B. and Hayden, F. V., 1856: Description of new fossil Tanabe, K. and Landman, N. H., 2002: Morphological diversity
species of Mollusca collected by Dr. F. V. Hayden, in of the jaws of Cretaceous Ammonoidea. In Summesberger,
Nebraska Territory; together with a complete catalogue H., Histon, K. and Daurer, A. eds., Proceedings of the 5th
of all the remains of Invertebrata hitherto described and International Symposium, Cephalopods – Present and
identified from the Cretaceous and Tertiary formations of Past. Abhandlungen der Geologischen Bundesanstalt , 57,
that region. Proceedings of Academy of . Natural Science p. 157–165.
of Philadelphia, vol. 8, p. 265–286. Tanabe, K., Tsujino, Y., Okuhira, K. and Misaki, A., 2015b:
Motoyama, I., Fujiwara, O., Kaiho K. and Murota, T., 1991: The jaw apparatus of the Late Cretaceous heteromorph
Lithostratigraphy and calcareous microfossil biochronology ammonoid Pravitoceras. Journal of Paleontology, vol. 89,
of the Cretaceous strata in the Oyubari area, Hokkaido, p. 611–616.
Japan. Journal of the Geological Society of Japan, vol. 97, p. Toshimitsu, S., Matsumoto, T., Noda, Y., Nishida, T., Maiya, S.,
507–527. (in Japanese with English abstract) 1995: Towards an integrated mega-, micro- and magneto-
Nixon, M., 1988: The buccal mass of fossil and Recent stratigraphy of the Upper Cretaceous in Japan. Journal of
Cephalopoda. In Clarke, M. R. and Trueman, E. R. eds., the Geological Society of Japan, vol. 101, p. 19–29. (in
The Mollusca, Vol. 12, Paleontology and Neontology of Japanese with English abstract)
cephalopods, p. 103–122, Academic Press, San Diego. Wright, C. W. and Matsumoto, T., 1954: Some doubtful
Nixon, M., 2015: Part M, Chapter 12: The buccal apparatus Cretaceous ammonite genera from Japan and Saghalien.
of Recent and fossil forms. Treatise Online, (69), 29 p. Memoirs of the Faculty of Science, Kyushu University,
The University of Kansas and Paleontological Institution, Series D (Geology), vol. 9, p. 107–134.
Lawrence, Kansas. Yabe, H., 1902: Note on three Cretaceous ammonites from
Saunders, W. B., Spinosa, C., Teichert, C. and Banks, R. C., Japan, outside of Hokkaido (continued). Journal of
1978: The jaw apparatus of Recent Nautilus and its Geological Society of Tokyo, vol. 9, p. 1–7.
palaeontological implications. Palaeontology, vol. 21, p. Yokoyama, M., 1890: Versteinerungen aus der japanischen
129–141. Kreide. Palaeontographica, vol. 36, p. 159–202.
Seilacher, A., 1993: Ammonite aptychi: how to transform a jaw
into an operculum?. American Journal of Science, vol.
293A, p. 20–32.
Takashima, R., Kawabe, F., Nishi, H., Moriya, K., Wani, R. and
Bulletin of the Mikasa City Museum, (24)You can also read

















































