Vertical Zonation of Some Crustose Lichens (Verrucariaceae) in Bay of Fundy Littoral Zones of Nova Scotia
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
2021
2021 Northeastern Naturalist
NORTHEASTERN NATURALIST Vol. 28, No. X
28(3):311–326
C.A. Vail and A.K. Walker
Vertical Zonation of Some Crustose Lichens
(Verrucariaceae) in Bay of Fundy Littoral Zones of Nova
Scotia
Cole A. Vail1 and Allison K. Walker1,*
Abstract - Despite harsh conditions provided by wave action, salinity, UV exposure, and
other pressures, lichens provide most of the substrate cover in the rocky littoral zone. We
documented the vertical zonation and general ecology of several littoral members of the
lichenized ascomycete family Verrucariaceae from 7 sites along Nova Scotia’s Bay of
Fundy coast. We identified lichens morphologically and confirmed species using ITS rDNA
barcoding to provide novel biodiversity data. We documented 8 lichens in Nova Scotia lit-
toral zones, with all transects showing low lichen diversity and moderate evenness. Vertical
zonation was not strongly correlated with mean low tide; a variety of biotic and abiotic
factors likely affected the distribution of the few species determining lichen community
composition in this region.
Introduction
Intertidal zones in the Bay of Fundy experience highly variable abiotic condi-
tions such as wind, wave action, salinity, and UV exposure (Higgins et al. 2015).
Though well-studied in the UK and Central Europe, the North American coasts
have underexplored littoral lichen biodiversity. The crustose and macro-algae,
accompanied by the lichens, make up the primary producers in these habitats;
lichens are a food source for some marine invertebrates (Higgins et al. 2015). In
cases where the rocky shore is too high to support substantial algal growth, li-
chens can serve as a large proportion of littorinid diet (Norton et al. 1990). High
lichen diversity can be observed mainly in the supralittoral zone (splash zone).
Abundance and diversity of lichens decreases approaching the sublittoral zone,
becoming limited to the members of the Verrucariaceae, Xanthopyreniaceae, and
Lichinaceae (Brodo et al. 2016). Within the Verrucariaceae, preference of habitat
in the intertidal zone can be clearly observed. Species such as Hydropunctaria
maura (Wahlenb.) Keller, Gueidan, and Thüs, and others in its genus prefer the
supra- to midlittoral zone. In comparison, members of the genus Wahlenber-
giella (Wahlenb.) Guiedan and Thüs, prefer the low-midlittoral to the lower
littoral zone (Higgins et al. 2015). Within the supra–sublittoral zone, most of
the lichen species belong to the Verrucariaceae (Thomas 1994). Shoreline lichen
species such as the Verrucariaceae or Lichinaceae prefer a siliceous substrate
such as granite but do occur in lower quantities and diversity on basic rock types
such as basalt (Gilbert 2000, Smith et al. 2017). For these reasons, the research
1
Department of Biology, Acadia University, Wolfville, NS B4P 2R6, Canada. *Correspond-
ing author - Allison.walker@acadiau.ca.
Manuscript Editor: David Richardson
311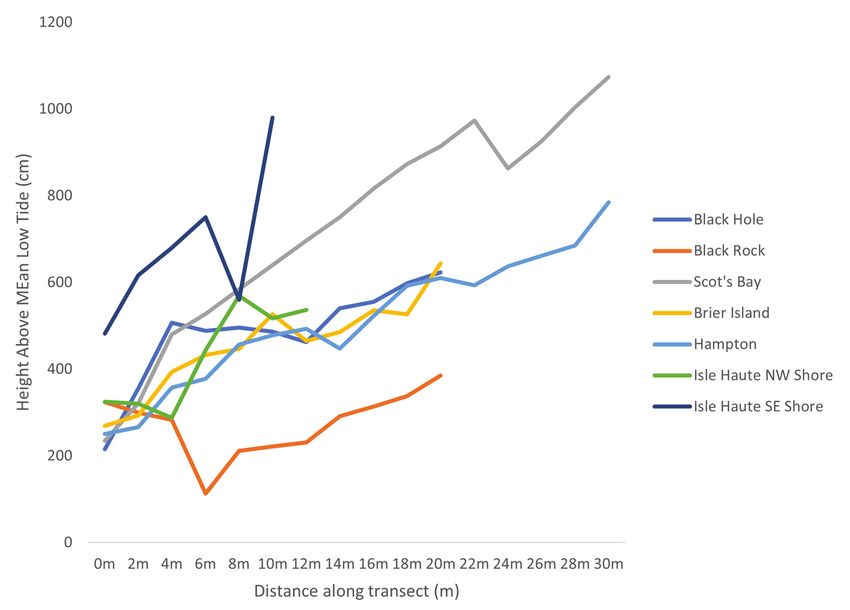
2021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
conducted here focused on the zonation of several Verrucariaceae species inhab-
iting the littoral zone of the Bay of Fundy.
The life habits of this family are versatile, with both endosubstratic and super-
ficial thalli observable. Their substrates can be biotic or abiotic; they may grow in
a lichenicolous manner, meaning on other lichen thalli as parasites (Knudsen et al.
2014). Members of the Verrucariaceae are known from freshwater and saltwater
habitats and are at times submerged, which is believed to be important for their
life cycle (Orange 2009) because lichen photosynthesis and dark respiration relies
on a relative thallus water content (WC) that is variable by species and in some
cases locality of those species. For example, in individuals of Lecanora muralis
(Schreb.) Rabenh., maximum net photosynthesis occurs around 0.55 mm WC and
levels off near 0.64 mm WC (Lange 2002). In freshwater habitats, especially acidic
waterways like streams or rivers with granitic substrate, regular dry conditions
can result in degenerated thalli, which in turn can cause aquatic lichens to die or
become dormant/nonreproductive (Krzewicka 2009). Most Verrucariaceae species
can withstand long periods of desiccation and UV exposure; an example of the more
desiccation-resilient genera is Hydropunctaria (Higgins et al. 2015).
Many genera within the family are polyphyletic, still requiring extensive mo-
lecular research to separate species or genera. Recent work by Gueidan et al. (2007)
and Orange (2012) placed several species into separate genera including, among
others, both the genera Verrucaria and Hydropunctaria. However, many species
still await further study for correct taxonomic placement, with the potential of
undescribed species being quite high. Traditional species delimitation for the Ver-
rucariaceae has been conducted morphologically, examining key characters like
thallus composition and color, ascospore septation, and presence or absence of
algae in the hymenium of the ascocarp of the specimens (Gueidan et al. 2007). The
genera in this family are generally fissitunicate, meaning they have an inner and
outer ascus wall, with the outer ascus wall splitting to allow the inner ascus wall to
be projected out of the hymenium. In the Verrucariaceae, the apical portion of the
outer ascus wall is gelatinized. Recent study of the family has discovered that spore
septation and algal presence or absence is variable even among species, resulting in
inaccurate morphological identifications (Gueidan et al. 2007).
To further our understanding of this family, we studied the density and population
diversity of Verrucariaceae along the Bay of Fundy coastline at 7 sites extending
from Scot’s Bay south to Brier Island, NS, Canada. Studies examining the vertical
zonation of these species above sea-level have been conducted by Sheard (1968)
and Ryan (1988), and more recently in the Bay of Fundy by Thomas (1994). More
recent genetic work has distinguished species from these habitats (Gueidan et al.
2007). We documented the vertical zonation, abundance, and diversity of littoral
zone lichens from the rocky shores of the Bay of Fundy and provided DNA analysis
of some specimens. This contributes much needed molecular data to the growing
genetic database of the Verrucariaceae. With molecular work conducted largely in
Europe for this group, corresponding data are needed from North America, includ-
ing the maritime regions of both Canada and the United States, to fully characterize
New World biodiversity in this group.
3122021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
Field-Site Description
The 320-km-long Bay of Fundy, situated between Nova Scotia and New Bruns-
wick (Fig. 1), experiences the highest tides in the world, >15 m in some portions
(Coon and Becker 2012). Together with the Gulf of Maine, it covers around 93,000
km2 (Coon and Becker 2012). The Bay of Fundy is comparable to the Bristol
Channel, UK, where tidal ranges exceed 9 m with southeast exposures to North
Atlantic waters (Thomas 1994). Among the many unique ecosystems the Bay of
Fundy contains are rocky coasts (Coon and Becker 2012), which can be separated
into 4 distinct vertical zones: the spray (supralittoral) zone, the high tide zone, the
middle tide (mediolittoral) zone, and the low tide (lower littoral) zone (Gilbert
2000). These zones are defined by length of exposure or submersion. The spray
zone is submerged only during storms of extreme high tides, and is commonly only
wetted by spray. The high tide zone is submerged only at mean high tide, while the
middle tide zone is inundated twice a day by tides. The low tide zone is frequently
submerged, only being exposed with the lowest of low tides (Thomas 1994). The
7 sites surveyed in this study include Black Hole, Black Rock lighthouse, Scot’s
Figure 1. Map of littoral lichen collection sites sampled in Nova Scotia in 2019, created us-
ing Google Earth© satellite data in Rstudio version 1.3.1093 for MacOS.
3132021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
Bay, the southeast and northwest shores of Isle Haute, Brier Island, and Hampton
(Fig. 1). All these sites are rocky shores composed of exposed basalt bedrock. The
rocky shore on either side of the sampling transects at each of these sites were
exposed to the elements, as was the shore-facing end of each transect, none of the
which extended more than 5 m into the supralittoral zone. Each transect extended
from the low littoral zone to the high tide zone and occasionally into the spray zone.
Materials and Methods
Site selection and collection methods
We made collections at 7 randomly selected sites along the Nova Scotian side
of the Bay of Fundy (Fig. 1, Table 1), which mostly consists of cobbled beach and
exposed basalt bedrock. We used a random number generator to select the sites
from among a set of predetermined potential sampling areas observed from satel-
lite imagery. At each site, we laid out 1 transect varying from 10 to 30 m in length,
depending on available exposed bedrock shore. Within a 50-cm² square every 2
m along the transects, we took a vertical measurement and collection where thalli
were accessible (Fig. 2). We took vertical measurements using a surveyor method
outlined in Thomas (1983). For 3 of the 7 sites, we used a rudimentary method to
enable a single person to take vertical measurements between 2 points along the
transect line. This method involved lying the end of a 2-m measuring stick on a lev-
el tripod, while leveling that meter stick to a vertically placed 1-m measuring stick.
We also collected substrate conditions and exposure data to characterize lichen
habitat. We estimated lichen or barnacle cover and canopy cover within the 50-
cm2 sampling squares. If percentages did not sum to 100%, any missing coverage
percentages represent bare rock. We estimated levels of lichen cover in percentage
groups of 5 (i.e., 5%, 10%, 15%, etc.) due to the large number of contiguous thalli
of multiple species, wherein more precise quantification of species composition is
difficult to distinguish due to thallus morphology overlap.
Identification
Morphological identification employed a Nikon SMZ1000 stereomicroscope,
with fresh material from sites sectioned by hand. We measured and photographed
ascospores in distilled water or 5% KOH using an AmScope 50-60Hz compound
light microscope.
Table 1. Nova Scotia field site names, coordinates and sampling dates.
Field site Latitude (°N) Longitude (°W) Sampling date
Black Hole 45.2366 64.4930 13 May 2019
Black Rock 45.1738 64.7641 16 May 2019
Brier Island 42.2866 66.3452 13 June 2019
Hampton 44.9052 65.3588 20 June 2019
Isle Haute SE Shore 45.2513 64.9908 25 June 2019
Isle Haute NW Shore 45.2516 65.0113 25 June 2019
Scot’s Bay 45.3141 65.3588 20 May 2019
3142021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
For molecular identifications, we extracted DNA from both thallus and ascoma
tissue from all fresh collections and purified it using the DNeasy Plant Mini Kit,
(Qiagen; Hilden, Germany). PCR reactions contained 12.5 μL Master Mix (Bio-
Rad), 9.5 μL distilled water, 1 μL of the reverse and forward primers (10 μM), and
1 μL of DNA. The Master Mix contained 0.075 units/μL Taq DNA polymerase,
4.0 mM MgCl2 pH 8.7, 0.4 mM dATP, 0.4 mM dCTP, 0.4 mM dGTP, and 0.4 mM
dTTP. We ran PCR reactions in a Biometra© T-Gradient 48 Thermal Cycler (Ana-
lytik Jena, Jena, Germany)for 2 hours and 50 minutes using the following PCR
parameters: 95 °C for 3 minutes, 56 °C for 45 seconds, 34 repetitions of 72 °C for
1 minute and 30 seconds, and a final round of 72 °C for 10 minutes. For the large
subunit (LSU) rDNA sequencing, we used the PCR primers LR0R (ACC-CGC-
TGA-ACT-TAA-GC), LR7 (TAC-TAC-CAC-CAA-GAT-CT), LR5 (TCC-TGA-
GGG-AAA-CTT-CG), and LR3R (GTC-TTG-AAA-CAC-GGA-CC) (Vilgalys and
Hester 1990). We used primers ITS-1F (CTT-GGT-CAT-TTA-GAG-GAA- GTA-A)
(Gardes and Bruns 1993) and ITS4 (TCC-TCC- GCT-TAT-TGA-TAT-GC) (White
et al. 1990) to amplify the internal transcribed spacer (ITS) rDNA region, which is
the accepted DNA barcode region for identification of fungi (Schoch et al. 2012).
To confirm successful PCR amplification, we conducted 1% agarose gel electro-
phoresis with ethidium bromide at 95 volts for 30 minutes.
We sent amplified DNA to the Genome Quebec Innovation Centre (McGill Uni-
versity, Montreal, QC, Canada) for Sanger sequencing in the forward and reverse
Figure 2. Vertical distances of each sampling point at each site in Nova Scotia, measured
every 2 m along transects 10–30 m in length.
3152021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
directions. We performed analysis and alignment of resulting consensus sequence
data using MEGA X: Molecular Evolutionary Genetics Analysis (Kumar et al.
2018). We searched consensus sequences against the NCBI GenBank online refer-
ence DNA sequence database using the BLASTn search algorithm.
Calculations and graphing
We performed graphing and calculations in Microsoft Excel (2018). We used the
Shannon–Wiener index to compare the relative diversity and abundance of species
in each transect, as there was large overlap in the relative species richness at each
transect.
Results
Substrate cover and distributions
We documented 8 lichens in Nova Scotia littoral zones, with all 7 transects
showing low lichen diversity and moderate evenness. Hydropunctaria species pre-
ferred a higher placement in all transects, and were negatively associated with the
Wahlenbergiella species W. mucosa (Wahlenb.), which extended further upwards
than W. striatula (Wahlenb.) Gueidan and Thüs, (Table 2). Wahlenbergiella mucosa
was more common than its sister species W. striatula and was the dominant species
in the Scot’s Bay transect and the southeastern shore of Isle Haute transect. In the
Scot’s Bay transect, for example, W. mucosa extended further up into the high lit-
toral region, despite the very large incline from the beginning of the transect. It did,
however, stay in relatively small numbers, and thalli were constrained to sheltered
cracks in the bedrock. In all other sites, Hydropunctaria species were the dominant
community, and never extended further down than 450 cm above the mean low
tide, (MLT), except at the Black Rock transect where it could be found as low as
221 cm above MLT. Verrucaria halizoa Leight. was the least common species we
encountered, and often blended in association with W. striatula thalli. Some in-field
misidentification is very likely for this species, due to its overlapping morphologies
with W. striatula, wherein it can sometimes appear as an underdeveloped thallus
of W. striatula. The respective sites were overall very similar in structure and ex-
posure. All sites were north-northeast facing, often with exposure to waves except
where sheltered by large boulders, for instance at the Black Hole transect. All sites
Table 2. Correlations of species occurrences across all transects and height above mean low tide (mlt).
Height Hydropunctaria
above mlt W. mucosa W. striatula sp. V. halizoa X. parietina
Height above mlt 1
W. mucosa -0.3378065 1
W. striatula -0.3082586 0.2608804 1
Hydropunctaria sp. 0.1376610 -0.3388282 -0.3100557 1
V. halizoa 0.0202090 0.0435655 -0.1006473 0.0276128 1
X. parietina 0.1397507 -0.1611741 -0.1209897 -0.0091313 -0.049988 1
X. elegans 0.0653408 -0.1088477 -0.0817095 -0.0188248 -0.017872 0.8394693
3162021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
were unshaded and exposed with the exception of those portions covered by Asco-
phyllum nodosum Scorpiodes canopies.
The Hydropunctaria species were the most abundant across all transects with
the exceptions of the Scots Bay Transect and the southeastern shore of Isle Haute,
where W. mucosa predominated. Verrucaria halizoa and W. striatula were the least
abundant depending on site; V. halizoa being least abundant at the Black Rock,
Scot’s Bay, and Hampton transects. Wahlenbergiella striatula was least abundant at
all remaining sites. All sites had a similar species diversity, though the southeastern
shore of Isle Haute was noted as the most diverse. The sites did, however, vary in
evenness, with values of 0.62–0.95, the highest being at Scot’s Bay and the least
at the Hampton transect (Table 3). The Wahlenbergiella taxa showed a moderate
negative correlation with increased vertical placement, while the rest of the species
showed weak positive correlations with increased height (Table 2).
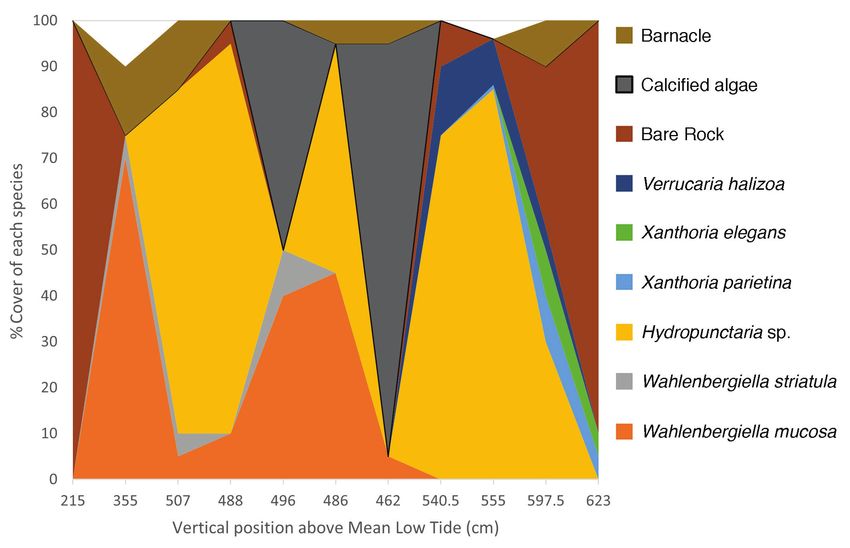
As an example of vertical zonation of lichen species, the transect in Black
Hole demonstrated the distribution trend that was present at all sites (Fig. 3). The
beginning (bottom) of the transect was marked by cobbled beach with no lichen
growth, mostly covered with Ascophyllum nodosum. It transitioned into bedrock
with large presence of W. mucosa and W. striatula; Ascophyllum nodosum cover
was still prevalent, but lichen growth was not hindered by the canopy cover, which
shaded 80% of the sampling square. Barnacles accounted for 15% of the substrate
cover within the 50-cm² sampling square. The 4-m mark had no canopy cover,
only barnacle and lichen cover. Barnacle cover was restricted to crevices in the
bedrock and accounted for 10% cover, and lichen cover was largely Hydropunc-
taria species, contributing to 75% of the substrate cover. Wahlenbergiella mucosa
and W. striatula thalli represented only 5% cover each. At the 6-m mark, the li-
chen cover consisted mostly of Hydropunctaria species (85%), with W. mucosa
thalli constrained to the underbelly of rocks within the sampling square. Evidence
of periwinkle herbivory was noticeable here, with contiguous thalli broken into
sectons 6–8 cm in width separated by feeding trails produced by herbivores. The
8-m mark had A. nodosum covering 90% of the sample square. The dominant spe-
cies of lichens were the Wahlenbergiella species, predominately W. mucosa with
small, interspersed thalli of W. striatula. Some Hydropunctaria species could be
observed on small, raised portions of rock within the sampling square, limited to
the apical portions of rock. The 10-m mark was much the same as the 8-m, with
Ascophyllum shading 70% of the substrate. Small amounts of barnacle species
could be observed, using 5% of the substrate, while the rest of the substrate (95%)
was covered with W. mucosa (45%) and Hydropunctaria sp. (50%). Accordingly,
the vertically lower 12-m mark was entirely covered with Ascophyllum and Fucus
Table 3. Shannon–Wiener index and evenness values for each study transect.
Black Black Brier Isle Haute Isle Haute Scot’s
Hole Rock Island Hampton SE NW Bay
Shannon 1.14 1.17 1.18 1.003 1.31 1.15 1.28
Evenness 0.63 0.84 0.73 0.62 0.94 0.83 0.95
3172021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
sp. with barnacles and calcified algae underneath, sometimes with dispersed thalli
of W. mucosa covering only about 5% of the substrate. Higher up, the 14-m mark
contained no barnacle cover, with about 20% Ascophyllum cover. Lichen species
consisted of Hydropunctaria maura thalli covering 75% of the substrate. Verru-
caria cf. halizoa thalli were now noticeable, though not exceeding more than 5%
of the substrate cover. The 16-m mark showed a transition into the splash zone in
this transect with less that 10% canopy cover from seaweed, and 5 individual thalli
of Xanthoria parietina (L.) Beltr., a dominant species in the supralittoral zone. Hy-
dropunctaria species covered 85% of the substrate. Verrucaria halizoa thalli could
be observed but comprised2021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
All 3 specimens identified morphologically as W. mucosa (AW3814, AW4018,
AW4022; Table 4) were supported as such in phylogenetic analyses, with boot-
strap values of >70% from 1000 bootstrap replications for each specimen yielding
strongly supported clades. Specimen AW4188, identified previously as a Hydro-
punctaria sp. was instead placed in V. degelii based on our DNA evidence, with
99% bootstrap support, making it the first record of this species in Nova Scotia.
Specimens AW4025 and AW4026 were identified as Collemopsidium halodytes
(Nyl.) Grube & B.D. Ryan based on morphology. Phylogenetic analysis revealed
them to be an unnamed or previously unsequenced species of Collemopsidium. All
voucher specimens can be found in Table 4, along with their E.C. Smith Herbarium
accession numbers and NCBI GenBank sequence accession numbers.
Discussion
This study highlights the lack of knowledge of lichen diversity in the littoral
zone in the Bay of Fundy. GBIF indicates known North American ranges for spe-
cies in the genus Wahlenbergiella as being from Newfoundland to Florida on the
East Coast, and from Vancouver to California on the West Coast. Well known from
Table 4. Taxa used for phylogenetic analyses. * indicates taxa obtained during this study; all other taxa
were obtained from NCBI GenBank.
Genbank ACAD
accession Genetic herbarium
Species Country Voucher number locus accession #
Collemopsidium sp.* Canada AW4025 MW211112 LSU ECS047940
Collemopsidium sp.* Canada AW4026 MW211114 LSU ECS047941
Collemopsidium sp. Spain s1310 KU556901 LSU
Collemopsidium sp. Spain s464 KU556888 LSU
Collemopsidium sp. Spain s1339 KU556903 LSU
Collemopsidium sp. Spain s1411 KU556906 LSU
Collemopsidium sp. Spain s260 KU556880 LSU
Verrucaria degelii UK 16902 FJ664838 SSU/ITS
Verrucaria degelii UK 17125 FJ664840 SSU/ITS
Verrucaria degelii Iceland AMNH: KY697142 SSU/ITS
LA12811
Verrucaria degelii Iceland AMNH: KY697141 SSU/ITS
LA31913
Verrucaria degelii* Canada AW4188 MW204569 ITS/LSU ECS047942
Wahlenbergiella mucosa Iceland AMNH: KY773263 LSU
LA31918
Wahlenbergiella mucosa UK 16305 FJ6644875 SSU/ ITS
Wahlenbergiella mucosa USA AFTOL-ID EF643802 LSU
2264
Wahlenbergiella mucosa* Canada AW3814 MW211115 LSU ECS047937
Wahlenbergiella mucosa* Canada AW4018 MW211116 LSU ECS047938
Wahlenbergiella mucosa* Canada AW4022 MW211113 LSU ECS047939
3192021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
the UK, there are several records of Hydropunctaria amphibia (Clemente) Cl.
Roux from a single locality (Cape Ann) on the Massachusetts coast, northeastern
US (Flenniken and Gibson 2003); however overlap in ecologies and cryptic mor-
phologies allow little morphological differentiation of Hydropunctaria species, and
these specimens may actually be H. maura, which is known from both the east and
west coasts of North America. Molecular identification of additional North Ameri-
can specimens using multiple genetic loci is required for confirmation; currently
H. aractina (Wahlenb.) Orange, H. orae Orange, and H. oceanica Orange are also
principally known from Europe, where littoral zone lichens have been more exten-
sively documented.
Factors affecting littoral lichen distribution
Level of submersion is the main factor affecting the vertical zonation of littoral
zone lichens (Gilbert 2000). However, given recent data acquired through trans-
plant methods, the zonation of species susceptible to herbivory, such as H. maura,
is controlled more effectively by grazing (Higgins et al. 2015). Lichens are also
poikilohydric organisms and with no waxy cuticle to control the flow of water, they
are unable to control the gain or loss of moisture or inorganic nutrients (Brodo et
al. 2016). Given the relationship between thallus water content and rates of respi-
ration and photosynthesis, the occurrence of H. maura higher in the littoral zone
indicates more reliance on rainfall or spray and, in some cases, submersion at high
tides (Higgins et al. 2015). In cases where the photosystem activity of W. mucosa
has been measured, the recovery of photosynthetic rates was more pronounced upon
inundation than in thalli of H. maura, while photosynthetic rates were also more
reduced with desiccation in W. mucosa than in H. maura (Higgins et al. 2015).
In terms of exposure to solar radiation, species within both Hydropunctaria and
Wahlenbergiella prefer open spaces on the bedrock, with Wahlenbergiella spp. re-
stricted to crevices or depressions likely to retain moisture, as they would still be
exposed to light. This exposure could lead to photoinhibition, which is a process
catalyzed by the production of reactive species of oxygen due to excessive intake of
photosynthetically active radiation, as well as UV-B or UV-A absorption (Beckett
et al. 2019, Gauslaa and Solhaug 2004). This photoinhibition would result in a de-
creased photosynthetic capacity of the photobiont (Beckett et al. 2019). Desiccated
thalli are more susceptible to damage from these factors due to a longer exposure
to the effects of solar radiation during the course of drying (Gauslaa and Solhaug
2004). With desiccation, repair mechanisms in lichen thalli are also hindered, ex-
ponentially increasing the effects of solar radiation (Gauslaa and Solhaug 2004).
The mycobiont may respond to solar radiation by producing secondary metabolites
such as usnic acid or periatin, and melanized (darker) thalli of lichens are more
resistant to high light stress than paler species, due to cortical melanin (Beckett et
al. 2019). This is the case with Hydropunctaria species, which are all characterised
by their dark carbonaceous thallus color, as compared to the greens or browns of
the other Verrucariaceae genera inhabiting the littoral zone. Even in paler species,
lichen substances such as acids are still useful in reflecting excess PAR due to their
3202021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
crystalline structure (Beckett et al. 2019). This is likely the case with W. mucosa in
these environments, with polyphenol concentrations being recorded as 3 times as
high as in H. maura (Higgins et al. 2015).
Water and air temperatures also effect the distribution of lichen species and their
photosynthetic rates (Gilbert 2000). In the Bay of Fundy, the yearly average annual
water temperature is 7 °C, rising to 10–12 °C in the summer months and dropping
to an average of 1.8 °C in the winter months. The optimal temperatures for lichens
vary by species and genotype, and even further based on the microhabitat they in-
habit, with small adjustments in temperature or humidity resulting in dieback for
some more sensitive lichen species wherein part of the thallus may deteriorate but
the thallus is still living. (Brodo 2016). With the Verrucariaceae and Xanthopyrenia-
ceae that inhabit the littoral zone, a wide temperature preference might be expected
of these lichen species as the environment is in constant flux. This is not the case,
however, as photosynthetic activity in W. mucosa is optimal at 10–20°C, decreasing
rapidly near the extremes of this spectrum (Higgins et al. 2015). Hydropunctaria
maura is more tolerant of the aforementioned extremes, having maintained high
photosynthetic rates during a 6-h emersion period at 10 °C (Higgins et al. 2015).
Optimal temperatures in both water and air are crucial for the growth of lichen
species in the littoral zone, with the Hydropunctaria group likely more reliant on
ambient temperatures, while species pertaining to the low-midlittoral zones such as
the Wahlenbergiella genus would be entirely reliant on submersion for their photo-
synthetic activities (Higgins et al. 2015).
Other than photosynthetic activity, wave action, thallus–water interactions, and
ambient temperatures, few other abiotic factors are likely to affect the growth of
these lichens. Lower species diversity could be due to the mafic nature of the sub-
strate in these Bay of Fundy sites, combined with a general preference for siliceous
substrate by the Verrucariaceae family (Gilbert 2000). The effects of pollution on
these species have not been studied extensively; however, impacts of oil pollu-
tion on other coastal lichens have been documented in Scotland and the Baltic Sea
(Dalby 1993, 1995; Dalby and Dalby 2005; Ranwell 1968; Schiefelbein, 2009).
While air pollution does impact some sensitive terrestrial lichens, which serve as
bioindicators, other lichens can maintain a healthy abundance in urban settings or in
places of high nutrient inputs (Coffey and Fahrig 2012). Due to their lack of waxy
cuticles, as well as lack of roots, terrestrial lichens rely on nutrient absorption via
atmospheric deposition (Begu et al. 2014). Deposition in lichens occurs through 2
methods: dry and wet deposition. Wet deposition can be defined as both precipita-
tion or occult deposition (fog or mist), with occult deposition contributing the most
(Nash and Gries 1995b). The sensitivity of some species also relies on this process
of gaseous absorption. Because they possess no waxy cuticle, lichens will absorb
nutrients across the entirety of their thallus surface, and when dehydration occurs,
these contaminant levels can be potentially raised to toxic levels, (Nash and Gries
1995a). Lichens cannot as readily dehisce their vegetative portions like plants. Any
contaminants absorbed remain in the thallus tissue indefinitely, reaching concentra-
tions much higher than that in the air (Begu et al. 2014). One detrimental pollutant
3212021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
produced by the burning of fossil fuels as well as the smelting of various metals is
sulfur dioxide (SO2). Controlled fumigations of contaminants such as SO2 provide
concrete evidence for the effects of these compounds on morphological characters
of lichens, such as the overall degradation of alga and cyanobacterium cell walls
and mitochondria (Nash and Gries 1995a). Lichens are more sensitive to SO2 than
plants, and photosynthetic responses of lichens can be decreased to negligible
within close distance to a source of SO2 in laboratory conditions (Moser et al.
1980). With environmental atmospheric deposition of contaminants, production of
apothecia, the sexual fruiting structure of most lichens, showed a decrease in pol-
luted areas, with lichens favoring production of asexual conidia under such polluted
conditions (Nash and Gries 1995b). With the loss of sensitive species, less-sensitive
or tolerant species of lichens would have the opportunity to increase in numbers due
to decreased competition. A diverse community of lichens could in time become a
less diverse, albeit tolerant community (Osyczka et al. 2018). Lichens thrive in spe-
cific microhabitats; some are also very sensitive to canopy change, and as a result
are reliable as bioindicators of stage succession (Thomson 1972).
Some factors attributing to chemical effects on lichen growth could be provided
by nutrient inputs such as bird guano. For plants, bird guano can result in phos-
phate-enriched soils allowing for faster growth (Irick et al. 2015). Abrasive action
of waves is lessened both by the extensive Ascophyllum cover and the large tidal
range of the Bay of Fundy (Thomas 1994). As well, the crusts of these lichen spe-
cies lay closely adhered to their substrate, using hyphae and anchoring mechanisms
to hold them in place despite the erosion of their bedrock substrate (Gilbert 2000).
Salinity has no large effect on photosynthetic activity; Fletcher (1980) showed that
gas exchange in some littoral species was unaffected by submersion in saltwater
even after 35 days.
Biotic pressures, such as grazing, canopy cover, and space competition may also
impact littoral lichens but remain understudied. Herbivory within the littoral zones
of the Bay of Fundy is mainly attributed to the snail genus Littorina, particularly
L. saxatilis (Olivi) (Rough Periwinkle), L. obstutata (L.) (Yellow Periwinkle), and
L. littorea (L.) (Common Periwinkle), as well as sea urchins like Strongylocentro-
tus droebachiensis (O.F. Müller) (Thomas 1994). No sea urchins were observed
in any of the transects in this study, but Littorina species could be found in the
majority of the transects, mostly in the sublittoral through the midlittoral but never
extending past the Wahlenbergiella/Hydropunctaria border. In previous studies,
Littorina were observed grazing epiphytes rather than fucoid macroalgae (Thomas
1994). Despite this association, lichens can become an important staple of snail
diet given inhabitation on shorelines too high to support substantial algal growth
(Norton et al. 1990). The related Littorina angulifera (Lamarck) (Mangrove Peri-
winkle) showed regular consumption of lichens or fungal propagules in mangroves,
with fungal tissue showing in fecal pellets 80% of the time (Kohlmeyer and Bebout
1986). In cases such as that of W. mucosa in the littoral zone, phenolic concentra-
tions are linked to decreased herbivory (Higgins et al. 2015). Despite this, some
3222021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
indirect evidence showing increased grazing of H. maura transplanted to the lower
littoral zone suggests some preference by littorinids, but further evidence is needed
for confirmation (Higgins et al. 2015).
With regards to canopy cover, the littoral zone of the Bay of Fundy possesses a
unique zonation of organisms, differing from both the Atlantic coasts of the mari-
time provinces and those of the UK and Europe (Thomas 1994). The marine brown
macroalga A. nodosum dominates the midlittoral zone of Bay of Fundy shores,
occurring in more-sheltered localities elsewhere (Thomas 1994). This difference
may be due to the large tidal range of the Bay reducing the vertical zonation re-
sponse of fucoid species to exposure, and the persistent community of A. nodosum
may further reduce the effects of wave action, giving under-canopy communities
a slightly more stable environment (Thomas 1994). Our sites had a characteristic
A. nodosum-dominated midlittoral region, which became limited near the supralit-
toral. This extensive canopy cover in some portions of the transects may affect the
photosynthetic capability of some lichen species, particularly W. mucosa. The Ver-
rucariaceae inhabiting the intertidal zone prefer openly exposed substrate, with no
extensive shade (Gilbert 2000, Smith et al. 2017).
Competition for space is also common in this environment, with lichens in our
transects experiencing interspecific competition from both barnacle and red cor-
alline algae species, commonly Semibalanus balanoides (L.) and Hildenbrandia
rubra (Sommerfelt) Meneghini, respectively. However, any significant effects other
than limiting space have insufficient data to suggest a strong effect on lichens in
the littoral zone. Barnacle species could commonly be found covering some lichen
thalli in our study, which would reduce photosynthetic activity.
Challenges
Prior to molecular evidence, most lichens were thought to have a cosmopolitan
distribution, as 2 individuals from different continents with the same morphological
characters would have been identified as the same species (Gueidan et al. 2007).
Species delimitation is important for lichen taxonomy, as well as the accuracy of
ecological studies; the phenotypic speciation of individuals has underestimated
the true diversity of lichenized ascomycetes (Lumbsch and Leavitt 2011). Cryptic
species such as those found in the Verrucariaceae reveal phenotypic delimitation
of species cannot provide an accurate estimate of the true diversity of lichenized
fungi and also indicate a lack of evidence of unique morphological traits serving
as species-level characters. The latest global estimate of lichenized fungi is 16,000
species (Lücking et al. 2017). With more groups having their molecular phylogenies
examined, the number of species can only be expected to steadily increase. The
recent debate of the importance of morphological characters when compared to
molecular phylogenies has bearing on our study (Lumbsch and Leavitt 2011). Mor-
phological characteristics were quite variable, and cryptic parasitic species were
not detectable by morphological investigation alone. It is also worth noting that
the number of Canadian sequences of these particular genera are lacking in public
genetic databases. Genetic databases reliant on voucher specimens from Europe or
the UK can lead to inaccurate identifications of North American taxa.
3232021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
Characterizing intertidal lichen communities and calculating the likelihood of
the occurrences of rare littoral lichens will boost accuracy and efficiency in find-
ing the associated symbionts, as well as improve the documented species diversity
present in understudied coastal areas. Crustose lichens often lack georeferenced
location data to support abundance or rarity rankings, and studies such as ours can
provide a foundation for future conservation management and diversity studies.
The Consortium of North American Lichen Herbaria shows several collections of 3
species identified in our study only at Peggy’s Cove, near Halifax, NS. Describing
species diversity and using DNA barcoding to confirm identifications will provide a
stronger foundation for future work on intertidal lichen communities within the Bay
of Fundy. M.L.H Thomas (1983, 1994) described the zonation of intertidal species
at Musquash Head and the Quoddy region of New Brunswick. He examined algae,
lichens, and invertebrate populations but did not extensively outline the ecologi-
cal roles or niches of lichens present in these studies; presence of a few species was
noted. New littoral lichens are being discovered in Europe, which has had extensive
molecular work done on littoral lichen communities (Pykälä et al. 2017). Continued
biodiversity studies in the Bay of Fundy will likely reveal additional new species re-
cords, as these important primary producers are understudied in this region.
Acknowledgments
C.A. Vail acknowledges an NSERC USRA Undergraduate Honours Research Award
through Acadia University. A.K. Walker acknowledges an NSERC Discovery Grant (No.
NSERC—2017-04325). We thank Dr. Stephen Clayden (New Brunswick Museum) and
Alain Belliveau (E.C. Smith Herbarium, Acadia University) for encouragement and exper-
tise. We also thank Sarah Adams (Acadia University, Department of Biology) for her labo-
ratory assistance; Génome Québec Innovation Centre, McGill University, for sequencing
services; and Brent Robicheau (Dalhousie University) for map-making assistance.
Literature Cited
Beckett, R., K. Solhaug, Y. Gauslaa, and F. Minibayeva. 2019. Improved photoprotection in
melanized lichens is a result of fungal solar radiation screening rather than photobiont
acclimation. The Lichenologist 51(5):483–491.
Begu, A., V. Brasoveanu, L. Nina, and V. Brega. 2014. Mosses and lichens: Bioindicators
of heavy metal pollutions of forest ecosystems. Aerul Si Apa. Componente Ale Mediului
2104:56–61.
Brodo, I. M., S.D. Sharnoff, and S. Sharnoff. 2016. Lichens of North America. Yale Uni-
versity Press, New Haven, CT. 795 pp.
Coffey, H., and L. Fahrig. 2012. Relative effects of vehicle pollution, moisture, and coloni-
zation sources on urban lichens. Journal of Applied Ecology 49(6):1467–1474.
Coon, J., and M.L. Becker. 2012. Can we get there from here? Ecosystem-based governance
in the Bay of Fundy/Gulf of Maine region. Ph.D. Dissertation. University of Newhamp-
shire, Durham, NH. 621 pp.
Dalby, D.H. 1993. Survey of lichen damage from the M.V. Braer. Unpublished report to
Scottish Natural Heritage, Edinburgh, Scotland, UK. 43 pp.
Dalby, D.H. 1995. Coastal lichens studies of areas affected by oil from the Braer. Northeast
Region Report 94. Scottish Natural Heritage, Edinburgh, Scotland, UK. 29 pp.
3242021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
Dalby, K., and C. Dalby. 2005. Shetland Lichens. Shetland Amenity Trust, Lerwick, Shet-
land, UK. 120 pp.
Flenniken, D.G., and E.S. Gibson. 2003. The littoral species of Verrucaria of Cape Ann,
MA. Evansia 20:31–48.
Fletcher, A., 1980. Marine and maritime lichens of rocky shores: Their ecology, physiol-
ogy, and biological interactions. Pp. 789–842, In J.H. Price, D.E.G. Irvine and W.F.
Farnham (Eds.). The Shore Environment, Vol. 2: Ecosystems. Academic Press, Lon-
don, UK. 622 pp.
Gardes M., and T.D. Bruns. 1993. ITS primers with enhanced specificity for basidiomy-
cetes: Application to the identification of mycorrhizae and rusts. Molecular Ecology
2:113–118.
Gauslaa, Y., and K. Solhaug. 2004. Photoinhibition in lichens depends on cortical charac-
teristics and hydration. The Lichenologist 39(2):133–143.
Gilbert, O. 2000. Lichens. HarperCollins, London, UK. 288 pp.
Gueidan, C., C. Roux, and F. Lutzoni. 2007. Using a multigene phylogenetic analysis to
assess generic delineation and character evolution in Verrucariaceae (Verrucariales, As-
comycota). Mycological Research 111(10):1145–1168.
Higgins, N., S. Connan and D. Strengel. 2015. Factors influencing the distribution of
coastal lichens Hydropunctaria maura and Wahlenbergiella mucosa. Marine Ecology
36(4):1400–1414.
Irick, D., B. Gu, Y. Li, P. Inglett, P. Frederick, M. Ross, A. Wright, and S. Ewe. 2015. Wad-
ing bird guano enrichment of soil nutrients in tree islands of the Florida Everglades.
Science of the Total Environment 532:40–47.
Knudsen, K., O. Breuss. and J. Kocourková. 2014. A new lichenicolous Heteroplacidium
(Verrucariaceae) from the deserts of southern California. Opuscula Philolichenum
13:26–33.
Kohlmeyer, J., and B. Bebout. 1986. On the occurrence for marine fungi in the diet of Lit-
torina angulifera and observations on the behaviour of the periwinkle. Marine Ecology
7(4):333–343.
Krzewicka, B. 2009. Some new records of Verrucaria from Beskid Niski mountains. Acta
Mycologica 44(2):265–273.
Lange, O. 2002. Photosynthetic productivity of the epilithic lichen Lecanora muralis:
Long-term field monitoring of CO2 exchange and its physiological interpretation. I. De-
pendence of photosynthesis on water content, light, temperature, and CO2 concentration
from laboratory measurements. Flora 197(4):233–249.
Lücking, R., Hodkinson, B. P., and S. D. Leavitt. 2017. The 2016 classification of lichen-
ized fungi in the Ascomycota and Basidiomycota: Approaching one thousand genera.
The Bryologist 119(4):361–416.
Lumbsch, H.T., and S.D. Leavitt. 2011. Goodbye morphology? A paradigm shift in the de-
limitation of species in lichenized fungi. Fungal Diversity 50(1):59–72.
Microsoft Corporation. 2018. Microsoft Excel. Version 16.50. Available online at https://
office.microsoft.com/excel.
Moser, T.J., T. H. Nash III, and W.D. Clark. 1980. Effects of a long-term field sulfur dioxide
fumigation on Arctic Caribou forage lichens. Canadian Journal of Botany. 58(21):2235–
2240. https://doi.org/10.1139/b80-258.
Nash, T., and C. Gries. 1995a. The use of lichens in atmospheric deposition studies with an
emphasis on the Arctic. Science of the Total Environment 160(C):729–736.
Nash, T., and C. Gries. 1995b. The response of lichens to atmospheric deposition with an
emphasis on the Arctic. Science of the Total Environment 160(C):737–747.
3252021 Northeastern Naturalist Vol. 28, No. X
C.A. Vail and A.K. Walker
Norton, T., A. Hawkins, S. Manley, J. Williams, and N. Watson. 1990. Scraping a living: A
review of littorinid grazing. Hydrobiologia 193(1):117–138.
Orange, A. 2009. Two parasitic species of Placopyrenium (Verrucariaceae) from freshwater
habitats in northwest Europe. The Lichenologist 41(2):131–139.
Orange, A. 2012. Semi-cryptic marine species of Hydropunctaria (Verrucariaceae, lichen-
ized Ascomycota) from northwest Europe. The Lichenologist 44(3):299–320.
Osyczka, P., P. Boron, A. Lenaer-Boro, and K. Rola. 2018. Modifications in the structure
of the lichen Cladonia thallus in the aftermath of habitat contamination and implica-
tions for its heavy-metal accumulation capacity. Environmental Science and Pollution
Research 25(2):1950–1961.
Pykälä, J., A. Launis, and L. Myllys. 2017. Four new species of Verrucaria from calcareous
rocks in Finland. The Lichenologist 49(1):27–37.
Ranwell, D.S. 1968. Lichen mortality due to “Torrey Canyon’” oil and decontamination
measures. The Lichenologist 4:55–56.
Ryan, B. 1988. Zonation of lichens on a rocky seashore on Fidalgo Island, Washington. The
Bryologist 91(3):167–180.
Schiefelbein, U. 2009. The marine and maritime lichens of the Baltic Sea: An overview.
Bibliotheca Lichenologica 100:463–494.
Schoch, C.L., Seifert, K.A., Huhndorf, S., Robert, V., Spouge, J.L., Levesque, C.A.
Levesque, W. Chen, and Fungal Barcoding Consortium. 2012. Nuclear ribosomal in-
ternal transcribed spacer (ITS) region as a universal DNA barcode marker for Fungi.
Proceedings of the National Academy of Sciences 109(16):6241–6246.
Sheard, J. 1968. The zonation of lichens on three rocky shores of Inishowen, Co. Donegal.
Proceedings of the Royal Irish Academy. Section B: Biological, Geological, and Chemi-
cal Science 66:101–112.
Smith, C.W., A. Aptroot, B.J. Coppins, A. Fletcher, O.L. Gilbert, P.W. James, and P.A.
Wolseley, 2017. Lichens of Greater Britain and Ireland. British Lichen Society, London,
UK. 1046 pp.
Thomas, M.L.H. 1983. Marine and coastal systems of the Quoddy Region, New Brunswick.
Canadian Special Publication of Fisheries and Aquatic Sciences 64. Department of Fish-
eries and Oceans, Ottawa, ON, Canada. 319 pp.
Thomas, M.L.H. 1994. Littoral communities and zonation on rocky shores in the Bay of
Fundy, Canada: An area of high tidal range. Biological Journal of the Linnaean Society
51:149–168.
Thomson, J. 1972. Distribution patterns of American Arctic lichens. Canadian Journal of
Botany 50(5):1135–1156.
Vilgalys, R., and M. Hester. 1990. Rapid genetic identification and mapping of enzymati-
cally amplified ribosomal DNA from several Cryptococcus species. Journal of Bacteriol-
ogy 172:4239–4246.
White, T.J., T. Bruns, S. Lee, and J. Taylor. 1990. Amplification and direct sequencing
of fungal ribosomal RNA genes for phylogenetics. Pp. 315–322, In M.A. Innis, D.H.
Gelfand, J.J. Sninsky, and T.J. White (Eds.). PCR Protocols: A Guide to Methods and
Applications. Academic Press, New York, NY. 482 pp.
326You can also read

















































