ANTARCTIC BIRDS (NEORNITHES) DURING THE CRETACEOUS-EOCENE TIMES
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
604 Revista de la Asociación Geológica Argentina 62 (4): 604-617 (2007)
ANTARCTIC BIRDS (NEORNITHES) DURING THE
CRETACEOUS-EOCENE TIMES
Claudia TAMBUSSI and Carolina ACOSTA HOSPITALECHE
Museo de La Plata, Paseo del Bosque s/nro, 1900 La Plata, and CONICET.
E-mails: tambussi@museo.fcnym.unlp.edu.ar, acostacaro@museo.fcnym.unlp.edu.ar
ABSTRACT:
Antarctic fossil birds can be confidently assigned to modern orders and families, such as a goose-like anseriform, two loon-like and a serie-
ma-like, all recorded before the K/T boundary at the López de Bertodano Fomation. Also, the discovery of a ratite and a phororhacids
from the uppermost levels of the Submeseta Allomember (Late Eocene), suggests that West Antarctica was functional to dispersal routes
obligate terrestrial birds. Representatives of Falconiformes Polyborinae, Ciconiiformes, Phoenicoteriformes, Charadriiformes, Pelagorni-
tidae and Diomedeidae constitute the non-penguin avian assemblages of the Eocene of La Meseta Formation. Fifthteen Antarctic spe-
cies of penguins have been described including the oldest penguin of West Antarctica, Croswallia unienwillia. The Anthropornis nordenskjoel-
di Biozone (36.13 and 34.2 Ma, Late Eocene) is characterized by bearing one of the highest frequencies of penguin bones and the phos-
patic brachiopod Lingula., together with remains of Gadiforms, sharks and primitive mysticete whales. Anthropornis nordenskjoeldi,
Delphinornis gracilis, D. arctowski, Archaeospheniscus lopdelli, and Palaeeudyptes antarcticus are exclusively of the La Meseta Formation. Anthropornis
nordenskjoeldi was evidently the largest penguin recorded at the James Ross Basin, whereas Delphinornis arctowski is the smallest, and include
one of the worldwide highest morphological and taxonomic penguin diversity living sympatrically. The progressive climate cooling of the
Eocene could have affected the penguin populations, because of climatic changes linked with habitat availability and food web processes.
However, there is not available evidence about Antarctic penguins' evolution after the end of the Eocene.
Keywords: Birds, Antarctica, Cretaceous, Paleogene.
RESUMEN: Aves antàrticas (Neornithes) durante el lapso cretácico - eoceno.
Las aves fósiles antárticas pueden ser asignadas a órdenes y familias vivientes, incluyendo restos de un Anseriformes que recuerda al ganso
overo, dos colimbos y una supuesta seriema, todos registrados en sedimentos cretácicos de la Formación López de Bertodano. El hallaz-
go de una ratites y un fororraco en los niveles más altos del Alomiembro Submeseta (Eoceno tardío) soporta la idea de que Antártida
Oeste fue utilizada como ruta de dispersión por aves terrestres. Representantes de los Falconiformes Polyborinae, Ciconiiformes,
Phoenicopteriformes, Charadriiformes, Pelagornitidae y Diomedeidae componen el conjunto de aves no-pingüinos registrados en los sedi-
mentos Eocenos de la Formación La Meseta. Hasta el momento se describieron quince especies de pingüinos, incluyendo el más antiguo
de los Sphenisciformes de Antártida Oeste, Croswallia unienwillia. Los pingüinos Anthropornis nordenskjoeldi, Delphinornis gracilis, D. arctowski,
Archaeospheniscus lopdelli, y Palaeeudyptes antarcticus asociados con restos de tiburones, misticetos primitivos y Gadiformes se encuentran en la
Biozona de Anthropornis nordenskjoeldi (36,13 and 34,2 Ma, Late Eocene). Estos niveles albergan una de las más grandes diversidades taxo-
nómicas de pingüinos hasta ahora conocida. Anthropornis nordenskjoeldi fue sin dudas el pingüino más grande del Eoceno de Antártida
mientras que en el otro extremo se ubica Delphinornis arctowski. Debido a que los cambios climáticos están ligados a la disponibilidad de
habitat y de recursos alimenticios, el progresivo enfriamiento climático acaecido durante el Eoceno podría haber afectado a las poblacio-
nes de pingüinos. Sin embargo, no tenemos evidencia acerca de la evolución de los pingüinos luego del Eoceno.
Palabras clave: Aves, Antártida, Cretácico, Paleógeno.
INTRODUCTION out in Seymour, James Ross and Vega Is- Recently, our understanding of the origins
land have resulted in the discovery of signi- and evolution of Neornithes - all modern
The James Ross Basin, at the Northern tip ficant vertebrate specimens that allow to birds-, has been dramatically influenced by
of the Antarctic Peninsula, is one of the improve our comprehension of the evolu- both molecular and fossil researches. In-
most important Early Cretaceous-early Pa- tionary history of Antarctic vertebrates, in deed, few neoavians from the end of the
laeogene sedimentary sequences in the particular the one that regards to birds. Mesozoic are known (Hope 2002), but so-
Southern Hemisphere (Francis et al. 2006a). However, despite intensive study of these me of them have been critical as factual evi-
Fossil floras and both invertebrate and ver- areas in the past decades, there is still much dences of the presence of modern lineages
tebrate faunas have provided clues to un- uncertainty about the exact composition of in the Cretaceous, and served as anchor
derstand past climate and paleoenviron- the Cretaceous-Paleogene Antarctic avifau- points for the molecular clocks. This is the
mental changes. Field expeditions carried na. case of the remarkable specimen of a mag-Antarctic birds (Neornithes) during the cretaceous-eocene times 605
pie-goose-like bird Vegavis iaai (Clarke et al. deposited in a back-arc setting relative to a and Cockburn iIslands, includes the Late
2005) to which we will refer below. volcanic arc through the Mid Mesozoic- Palaeocene Cross Valley Formation and the
By other hand, the most significant fossil early Cenozoic times (Hathway 2000), du- richly fossiliferous Eocene La Meseta For-
bird record from the James Ross Basin is ring subduction of the Pacific Ocean crust mation, both deposited in incised-valley
that of penguins. Currently, fifteen penguin beneath Gondwana (Hayes et al. 2006). The settings. At its type section, in the central
species have been described, and at least ten basin infilling consists of sandstones, silts- art of Seymour Island, the Cross Valley
of which would have coexisted. Most pro- tones and conglomerates, and comprises Formation (Elliot and Trautman 1982) fills
blematic is the assignment of many species three units: 1) the older Gustav Group (Ap- a steep-sided valley cut in the Lower Pa-
from the Eocene of Seymour that are based tian-Coniacian) that comprises the Peder- laeocene Sobral Formation and older beds
on non-comparable bones or different son, Lagrelius Point, Kotick Point, Whisky (Tambussi et al. 2005).
parts of the skeleton (Tambussi et al. 2006, Bay and Hidden Lake formations, all confi- The youngest bird fauna is from La Meseta
Tambussi et al. 2005). The recently publis- ned to the NW coast of James Ross Island Formation, which overlies the López de
hed catalogue by Myrcha and coauthors (Crame et al. 2006); 2) the Marambio Group Bertodano Formation. This unit was inter-
(2002) is a valuable source for the sphenis- (Coniacian-Maastrichtian), divided into preted as the filling of an incised-valley sys-
cids described up to date. Santa Marta, Snow Hill Island and López tem and is the topmost exposed sector of
The purpose of this paper is to review the de Bertodano formations (Pirrie et al. 1997) the sedimentary fill of the Late Jurassic-
current state of knowledge of Antarctic and is exposed over most of the James Ross Tertiary James Ross Basin (Del Valle et al.
Cretaceous-Paleogene avian fossils. Our ap- Basin. The latter group contains abundant 1992). It is composed of sandstones, muds-
proach has four parts: 1) we describe and microfossils, as well as fossil plants, inverte- tones and conglomerates deposited during
analyze the fossil continental birds; 2) we brates and vertebrates assemblages, profu- the Eocene in deltaic, estuarine and shallow
report and analyze the fossil marine birds; sely studied in the last years; and 3) the marine settings (Marenssi et al. 1998 a, b ).
3) we discuss the bioestratigraphic impor- Seymour Island Group (Early Paleocene- From the base to the top, six units are dis-
tance of the fossil penguin assemblage, and Late Eocene) that includes the Sobral, tinguished (Marenssi et al. 1998b): Valle de
4) we discuss the paleobiological significan- Cross Valley and La Meseta formations Las Focas, Acantilados, Campamento, Cu-
ce of the Antarctic fossil birds. (Francis et al. 2006b). cullaea I, Cucullaea II and Submeseta
Before developing each of these topics, The Late Cretaceous López de Bertodano Allomembers. The Valle de las Focas, Acan-
some geological characteristics of James Formation contains the oldest Antarctic tilados and Campamento Allomembers
Ross Basin will be considered. A more de- avian remains currently recorded (Case et al. constitute facies association I, composed by
tailed account can be found in Francis et al. 2006a, Chatterjee 1989, Chatterjee 2002, a fine-grained sequence with mudstones
(2006b). Chatterjee et al. 2006, Clarke et al. 2005, and very fine sandstones deposited in a del-
The following institutional abbreviations Noriega and Tambussi 1995, 1996). Among ta front plain environment. Facies associa-
are used in this paper: MLP Museo de La them, the anseriform Vegavis iaai was col- tion II includes the Cucullaea I, Cuccullaea
Plata, MACN Museo Argentino de Ciencias lected at Cape Lamb, southwestern Vega II and the lower part of the Submeseta
Naturales Bernardino Rivadavia, UCR Uni- Island (Western Antarctica), a well-known Allomembers, ranging from conglomeratic
versity of California Riverside, IB/P/B place because of its abundant and diverse beds to mudstones with diverse and abun-
Prof. A. Myrcha University Museum of Na- fossil record that includes conifers (Césari dant macrofauna (Marenssi et al. 1998b)
ture, University of Bialystok, Poland, TTU 2001), marine invertebrates, elasmosaurids, that corresponds to a valley-confined es-
P Museum of Texas Tech University. Ana- mosasaurids (Martin 2006) and a duck- tuary mouth to inner estuary complex. The
tomical nomenclature follows Nomina ana- billed dinosaur (Case et al. 1987). The sedi- base of the Cucullaea I Allomember has
tomica avian (Baumel and Witmer 1993) mentary sequence has been subdivided into produced a 87Sr/86Sr date of 49.5 Ma (Ma-
using English equivalents, with some modi- three informal units K1, K2 and K3 (Ma- renssi 2006). Finally, facies association III,
fications when necessary. Appendix I inclu- renssi et al. 2001), being the former two which includes the topmost sediments of
des the complete list of materials recovered Early Maastrichtian and the latter Mid-Late Submeseta Allomember, is characterized by
at Antarctic Peninsula and Islands. Maastrichtian. The unit K3 comprises the a more unvarying sandy lithology compo-
upper part of the Cape Lamb Member and sed mainly by fine to medium-grained sand-
GEOLOGICAL SETTING the Sandwich Bluff Member of the López stone and represents sedimentation on a
AND CLIMATIC CONDI- de Bertodano Formation (sensu Pirrie et al. sandy tidal shelf influenced by storms. The
TIONS 1991) or the Unit B (Olivero et al. 1992), three facies associations described above
which is has been dated in approximately suggest a major transgressive cycle. Dingle
Fossil birds are preserved within marine 66-68 million years old based on correla- and Lavelle (1998) reported a 87Sr/86Sr deri-
sediments in the James Ross Basin, which is tions of ammonites and palynological taxa ved age of 34.2 Ma (late Late Eocene) for
part of the larger Larsen Basin (Del Valle et (Crame et al. 1991, Pirrie et al. 1991). the topmost part of La Meseta Formation
al. 1992) on the East side of the Antarctic The Tertiary section (Seymour Island whereas Dutton et al. (2002) reported ages
Peninsula (Fig. 1). These sediments were Group), exposed mainly on Seymour Island of 36.13, 34.96 and 34.69 Ma (late Late606 C. TAMBUSSI AND C. ACOSTA HOSPITALECHE
Figure 1: a, Sketch geological map of the James Ross Island area. b. Cape Lamb, Vega Island, c. Seymour Island.
Eocene) for different levels within vents. The earliest studies upon fossil conti- cations, the ratites include two species of
Submeseta Allomember. nental birds in Antarctica were made by ostriches (Struthionidae) in Africa and Asia,
The climate in the Antarctic Peninsula du- Covacevich and Lamperein (1972) and Co- the Australian emu and three species of
ring the Late Cretaceous and Paleogene vacevich and Rich (1982) working at Fildes cassowaries (Casuariidae) in New Guinea
would have been relatively mild and moist, Peninsula in King George Island, the lar- and northeastern Australia, three species of
with no significant presence of ice at high gest of the South Shetland Islands. The forest-dwelling kiwis (Apterygidae) in New
latitudes (Francis 1996, Poole et al. 2001). A mid-Tertiary lacustrine sediments of King Zealand, and two rheas (Rheidae) in South
cooling event and a frostless climate charac- George Island preserved ichnofossils from America (Sibley et al. 1988). All the ratites
terized the environments between the Late four types of birds including the avian live currently in the Southern Hemisphere,
Cretaceous and the mid-Paleocene (Dingle tetradactyle footprint Antarctichnus fuenzali- and all of them lack a keel on the sternum,
and Lavelle 1998, Zachos et al. 1993). The dae Covacevich and Lamperein (1970) asso- a character associated with flightlessness.
fossil evidence suggests that during the Pa- ciated with shorebirds. One of the mor- The Antarctic material is a distal tarsometa-
leocene a cool to warm climate and high photypes apparently represents a non-vo- tarsus with a "large, narrow trochlea for di-
rainfall prevailed (Poole et al. 2001), where- lant ground bird that could belong to either git III, which is projected moderately
as paleotemperature data from the sea indi- ratites or gruiforms, and another probably beyond the trochlea for digit II with
cate that a peak occurred in the Early Eo- represents an anatid. In summary, the ich- straightend margins bordering a deep groo-
cene. Sedimentological (Coxall et al. 2005, nofossils from Fildes Peninsula include ve. Trochlea II has a wide articular surface
Ehrmann and Mackensen 1992), oxygen both solitary and group activities with their and extends posteriorly more than trochlea
isotopic (Dutton et al. 2002, Gadzicki et al. hypothetical avian tracemakers. III.The lateral margin of trochlea III allow
1992, Ivany et al. 2004, Kennett and Warnke Two different taxa of large flightless curso- us to infer that the intertrochlear space bet-
1993, Mackensen and Ehrmann 1992, Sa- rial birds from Antartica have been so far ween trochlea III and IV extends proxima-
lamy and Zachos 1999), floral (Francis described (Figs. 2 and 3), being a ratite tely beyond trochleae II and III" (Tambussi
1999, 2000) and faunal (Aronson and Blake (Tambussi et al. 1994) and a phororhacid et al. 1994). The estimated body mass of the
2001, Dzik and Gadzicki 2001, Feldmann bird (Case et al. 2006, Case et al. 1987). Both Antarctic specimen is approximately 60 kg
and Woodbourne 1988, Gadzicki 2004, forms were recovered from the topmost (Vizcaíno et al. 1998).
Myrcha et al. 2002, Reguero et al. 2002) data levels of the Submeseta Allomember, part Phorusrhacids are a predominantly Neo-
indicate cooling, growth of terrestrial and of the near-shore deposits of the La Me- gene group of large predatory, terrestrial
marine ice sheets, and initiation of Ceno- seta Formation on Seymour Island, likely birds (Alvarenga and Höfling 2003) recor-
zoic glaciation at the end of the Eocene Late Eocene (ca 36 Ma Dutton et al. 2002, ded between the Late Paleocene (Brazil,
(Birkenmajer et al. 2004). Reguero et al. 2002). They are part of the Itaborian SALMA) and Late Pleistocene
few records of terrestrial biota recovered (USA) (MacFadden et al. 2006, Tambussi et
THE FOSSIL CONTINEN- from this predominantly marine formation. al. 1999). Classical studies on these birds
TAL BIRDS Strictly Late Eocene terrestrial birds of classified their diversity within five subfami-
Antarctica raise some interesting biogeo- lies (Brontornithinae, Phorusrhacinae, Pata-
The discovery and study of fossil continen- graphic issues that we will discuss below. gornithinae, Mesembriornithinae and Psi-
tal birds in Antarctica are relatively old e- According to current ornithological classifi- lopterinae) with a wide range of sizes andAntarctic birds (Neornithes) during the cretaceous-eocene times 607
morphotypes, since the sturdy non-flying cene of France. An incomplete right radius
brontornithines to the gracile and flying (MLP 87-II-1-2) of the La Meseta Forma-
psilopterines (Tambussi and Noriega 1996). tion was reported by Noriega and Tambussi
Phorusrhacid remains have been found in a (1996).
variety of sedimentary rocks in Uruguay, A probable Ciconiiforms was found at the
Brazil, Antarctica, United States, and Pa- upper level of La Meseta Formation (MLP
tagonia (Argentina), where they are best 90-I-20-9, which consists in a distal frag-
known Currently it is assumed that the Eu- ment of a right tarsometatarsus). Unfortu-
ropean "Phorusrhacidae" (Mourer-Chau- nately, the material is not preserved enough
viré 1981, Peters 1987) do not belong wi- to allow a more precise identification.
thin Phorusrhacidae but to Strigogyps (Mayr Recently, unquestionable remains of neor-
2005). nithines from the Maastrichtian of Antarc-
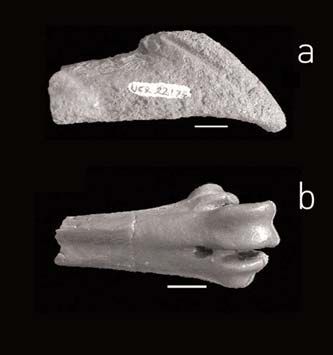
A distal end of bill (Fig.3) attributed to a gi- tica have bridged the disagreement between
gantic supposed phorusrhacid (Gruiformes Figure 2: Ratites. MLP 94-III-15-1, distal
molecular and palentological data about the
from Seymour Island, was described by Ca- fragment of right tarsometatarsus in poste- diversification history of Neornithes (Dyke
se and colleagues (1987). Additional mate- rior view. Scale: 10 mm. and Van Tuinen 2004). As mentioned pre-
rials assigned to phorusrhacids were recen- viously, the Anseriform Vegavis iaai Clarke et
tly described from the same levels (Case et al. 2005 was recovered from a southwestern
al. 2006). One of these specimens consists locality at Cape Lamb in Vega Island (Fig.
in a tarsometatarsus (Fig.3) with unquestio- 5). In a recent work, Clarke et al. (2005)
nable phorusrhacid affinities, similar in size point its importance out as one of only
to Patagornis marshii. The other two ele- handful specimen considered as a true Ne-
ments, a vertebra and a tibiotarsus, seem ornithinae, and whose phylogenetic posi-
not to be a Phorusrhacidae and we think tion has been established. Vegavis provides a
that their assignment should be revised. well-defined phylogenetic calibration point
In addition to phorusrhacids and ratites, for estimating the early divergence of mo-
other avian species have distributions that dern birds (see Slack et al. 2006).
span multiple continents. Current biogeo- By other hand, a fragment of femur recove-
graphic hypotheses based on the Gond- red near the base of Sandwich Bluff Mem-
wanan fragmentation or long distances mi- ber (Vega Island) at a level equivalent to
grations. Although the phylogenetic affini- that of Vegavis iaai, was identified as a serie-
ties of the Antarctic ratites and phororha- ma-like bird by Case et al. (2006). Spite se-
cids are not clear, their discovery strongly Figure 3: Phorurhacids cast UCR 22175, a) riemas have traditionally been considered as
Fragment of the bill, b) tarsometatarsus ante-
supports the idea that West Antarctica was rior view. Scale: 10 mm.
descendants of the phorusrhacids (Alva-
used as dispersal route for obligate terres- renga and Hofling 2003), further phyloge-
trial organisms. wider than the trochlea for the digit four, netic analysis between modern and fossil
The crown-group Falconiformes includes bearing a plantarly projection. This falconid Gruiformes birds are necessary, and the
the New World vultures (Cathartidae), the bird, together with the phorusrhacid, were monophyly of all the Phorusrhacidae is yet
secretary bird (Sagittaridae), the falcons the representatives of the carnivorous to be verified.
(Falconidae), and the hawks and allies (either scavenger or predator) role within Beyond this, all these avian records are cru-
(Accipitridae) (see discussions about the the late Eocene Antarctic fauna. cial for studies of biogeographic trends
monophyly of Accipitridae in Mayr et al. Unambiguous Charadriiform birds are during the final phases of the Gondwana
2003). Living Polyborines are vulture-like known from the late Eocene of the La Me- break-up.
falconids with scavenging habits that occur seta Formation, based on a right scapula
exclusively in the Americas, mainly in the (MLP 92-II-2-6). All Charadriiform, shore- THE FOSSIL MARINE
Neotropical regions. Polyborinae have been birds and waders are a heterogeneous and BIRDS
recorded upon a tarsometatarsus from the polymorphic group of birds of small to
La Meseta Formation (Tambussi et al. 1995) moderate size that frequent open inland Neogaeornis wetzeli Lambrecht, 1933 and
(Fig. 4). The animal would have reached a and marine wetlands. "Polarornis gregorii" have respectively been
body mass of about one kilogram and the Flamingos (Phoenicopteridae), are grega- described from the late Cretaceous of Chile
size of the living caracara Polyborus plancus. rious and invariably associated with warm and Antarctica (Chatterjee 1989, Chatterjee
This tarsometatarsus exhibits a morpholo- temperatures, brackish or salt-water lakes 2002). Both taxa have been considered as
gy similar to living polyborines in having and lagoons. The oldest record assigned to members of the crown gaviids or the stem
the trochlea for the second digit shorter and Phoenicopteridae, is from the lower Oligo- gaviiforms, and their phylogenetic affinities608 C. TA M BU S S I A N D C. AC O S TA H O S P I TA L E C H E
are still unknown (Mayr 2004). Living loons guin-likeplotopterids (González-Barbaa et
and grebes (Gaviiformes, Gaviidae) are al. 2002). Warheit (1992) has suggested that
foot-propelled diving birds. They show a such an assemblage for the Late Eocene
restricted North American distribution that could be the result of a worldwide oceanic
winter along sea coasts and breed at fres- cooling occurred at 50 Ma.
hwater sites. Procellariiformes include the modern alba-
Chaterjee (1997, 2002) described and figu- trosses, petrels and storm-petrels. Modern
red the skull of "Polarornis", but some skep- albatrosses (Diomedeidae) are worldwide
ticism about its assignment and anatomical pelagic and gliding sea-birds southern oce-
information arised. ans. However, its fossil record is fairly from Figure 4:
Gerald Mayr (2004) along with his descrip- the Northern Hemisphere, where they ap- Falconiformes
tion of the Paleogene Colymboides metzleri, pear since the Late Oligocene (Tambussi Polyborinae
MLP 95-I-10-8,
commented about Polarornis: "if correctly and Tonni 1988). A weathered tarsometa- distal fragment
assigned to the Gaviiformes, may be a sy- tarsus from the La Meseta Formation at of left tarsome-
nonym of Neogaeornis - a possibility already Seymour Island (Noriega and Tambussi tatarsus, anterior
proposed by Olson (1992) but not discus- 1996; Tambussi and Tonni 1988) can be view. Scale: 10
mm.
sed by Chatterjee 2002" (Mayr 2004: 285). unambiguously assigned to this family. Ad-
If this is the case, Polarornis should be con-
sidered junior synonym to Neogaeornis wetze-
li.
More recently, Chatterjee et al. (2006) pre-
sented a new species of "Polarornis" that
exhibit both aerial and aquatic locomotion
modes.
Fossil remains of the extinct bony-toothed
Pelagornithidae (Odontopterygiformes) we-
re found in the Late Eocene La Meseta For-
mation (Tonni and Tambussi 1985, Tonni,
1980). Remains of these enigmatic birds ha-
ve been also recovered from England, Eu-
rope, North America, Japan, New Zealand,
Africa, Chile and Peru (Harrison and Wal-
ker 1976, McKee 1985, Olson 1985, Walsh
and Hume 2001, Warheit 1992). Pseudo-
dontorns, supposedly related to pelicans
(Pelecaniforms) and tube-nosed birds (Pro-
cellariiformes), were large marine gliding
birds equipped with bony projections along
the edges of their robust bills (Fig. 6). An
alternative hypothesis about their phyloge-
netic affinities was proposed recently
(Bourdon 2005). This author proposes the
sibling relationships between the pseudo- Figure 5: Anseriformes
dontorns and waterfowl (Anseriformes), Vegavis iaai MLP 93-I-1-3
holotype. Above, larger
erecting the clade Odontoanserae to inclu- half concretion that pre-
de Odontopterygiformes plus Anserifor- serves most of the bones
mes. Regardless of their phylogenetic posi- of the holotype, Below, the
tion, pseudodontorns included taxa that second half of the same
concretion.
were among the largest known flying birds.
Noteworthy, the pelagornithids of the Late ditional fossil specimens housed at Museo nesting behaviour, near-shore aquatic habi-
Eocene of Seymour Island (as discussed de La Plata could be also assigned to Pro- tat and lack of skeletal pneumaticity (Triche
below) are associated with penguins, while cellariidae (Noriega and Tambussi 1996). 2006). They belong to a much derived clade
the pseudodontornitids from the Northern Thousands of bones are accumulated in of modern birds, Sphenisciformes (the cla-
Hemisphere were associated with the pen- some fossil sites, likely due their colonial de including all fossil and living penguins,Antarctic birds (Neornithes) during the cretaceous-eocene times 609
but see Clarke et al. 2003) with aquatic lifes-
tyle, non-pneumatic bones and wings trans-
formed into flippers.
The Late Paleocene Crossvallia unienwillia
Tambussi et al. 2005, together with the late
Eocene Anthropornis nordenskjoeldi Wiman
1905, Anthropornis grandis (Wiman 1905), Pa-
laeeudyptes antarcticus Huxley 1859, Palaeeu-
dyptes klekowskii Myrcha et al. 1990, Palaeeu-
dyptes gunnari (Wiman 1905), Archaeosphenis-
cus wimani (Marples 1953), Archaeospheniscus
lopdelli Marples 1952, Delphinornis larseni
Wiman 1905, Delphinornis gracilis Myrcha et
al., 2002, Delphinornis arctowskii Myrcha et al.
2002, Marambiornis exilis Myrcha et al. 2002,
Mesetaornis polaris Myrcha et al. 2002, Tonnior-
nis mesetaensis Tambussi et al. 2006 and Ton-
niornis minimum Tambussi et al. 2006, join to
the fifteen penguin species previously
known (Appendix I).
The Eocene species were primarily found
in sediments of the Submeseta Allomem-
ber, although four were recorded in the
Cucullaea I Allomember (Fig.7).
Due to the fragmentary nature of their re- Figure 6: Odontopterygi-
cord, the spheniscids' systematic is based formes Pelagornithidae,
on isolated bones, usually upon tarsometa- MLP 78-X-26-1, fragment
of the rostrum. Arrows
tarsi (Jadwiszczak 2001, 2003) and humeri show projections of the
(Simpson 1946). Indeed, most of the spe- tomia, a) lateral view, b)
cies are only known from one of those ele- transversal view. Scale: 10
ments. mm.
Regarding Antarctic fossil penguins, Myr-
cha et al. (2002) studied exclusively the tar- nisciformes (Tambussi et al. 2005; Slack et
sometatarsi and identified four new species, al. 2006), although molecular evidence sug-
whereas Tambussi et al. (2005, 2006) added gests a Late Cretaceous origin for the
three new ones based on humeral morpho- group.
logy. Considering that Crossvallia unienwillia, Ksepka and colleagues (2006) placed Wai-
Tonniornis minimum and T. mesetaensis are only manu outside of a clade that includes all
known by their humeri, and Palaeeudyptes kle- other penguins. Also, near the base, in a
kowskii, Delphinornis arctowskii, D. gracilis, Me- more basal position, Delphinornis larseni is
setaornis polaris and Marambiornis exilis were located as sister taxon of Mesetaornis polaris,
identified by their tarsometatarsi, compara- Marambiornis exilis and the remaining pen-
tive measurements and a deep anatomical guin species. Thus, most of the fossil pen-
descriptions by Kandefer (1994) and Tam- guins are nested in a largely pectinate arran-
bussi et al. (2006) allowed assigning some gement leading to the crown clade Sphe-
humeri to species previously known only by niscidae that includes all modern species of
the tarsometatarsi. Beyond these criteria, penguins (Ksepka et al. 2006 Figs. 2 and 3).
Jadwiszcak (2006) in his excellent work re- The pioneering work of Simpson (1946)
Figure 7: Sphenisciformes, representatives
cognizes several species upon elements o- provided the first systematic proposal at su-
bones of Anthropornis sp., a)right humerus in
ther than humeri and tarsometatarsi (see prageneric level (five subfamilies Palaeos- caudal view. Scale: 10 mm, b) left tibiatarsus
Appendix I). pheniscinae, Paraptenodytinae, Palaeeudyp- anterior view.
Crosswallia and the recently described tinae, Anthropornithinae, Spheniscinae),
Waimanu Jones, Ando and Fordyce 2006 and has remained the basis for all other lacking a cladistic framework (Clarke et al.
from the Paleocene are the earliest Sphe- analyses of penguin relationships, although 2003). Some of these subfamilies could be610 C. TA M BU S S I A N D C. AC O S TA H O S P I TA L E C H E
considered clades (Ksepka et al. 2006; that the penguins of La Meseta Formation record, diverse conjectures have been made
Acosta Hospitaleche et al. 2007) but some represent a high-quality record. We advan- about their biology. Southern South
modifications and further revisions are re- ced this idea in Tambussi et al. (2006). The America penguin colonies are formed
quired. According Ksepka et al. (2006), all upper part of the Submeseta Allomember exclusively by Spheniscus magellanicus, at both
Patagonian fossil species (more than six concentrates the bulk of the penguin-bea- Pacific and Atlantic coasts. Remarkably, o-
taxa of Palaeospheniscinae, Paraptenodyti- ring localities and documents the highest ther species of this genus also form exclu-
nae and Anthropornitinae in Simpson's morphological and taxonomical diversity of sive colonies, such as S. demersus in the
view ) fall outside the Spheniscidae (the less sympatric penguins worldwide. Five spe- South African coasts, S. humboldti in the Pe-
inclusive clade uniting all extant penguin), cies, Anthropornis nordenskjoeldi, Delphinornis ruvians and S. mendiculus in the Galapagos
refuting the monophyly of all the subfami- gracilis, D. arctowski, Archaeospheniscus lopdelli, archipelago. In contrast, the colonies that
lies excepting the clade composed by the and Palaeeudyptes antarcticus, are exclusive of occur in the Malvinas (Falklands) and South
modern taxa. According to our analysis (A- these upper levels in which their first and Georgias Islands comprise up to five sym-
costa Hospitaleche et al. 2007), Paraptenodytes last appearances took place. Because of patric species: Aptenodytes patagonicus,
from the Early Miocene (about 20 Ma) is these bioestratigraphic evidences, the An- Pygoscelis papua, P. antarctica, Eudyptes chrysoco-
located at the base of the Spheniscidae and, thropornis nordenskjoeldi Biozone was defined, me and E. chrysolophus. The islands situated
with some restrictions, we recognized some with an estimated age between 36.13 and south from South Africa are inhabited by
of the Simpson's clades (1946) such as Pa- 34.2 Ma, (Late Eocene, Tambussi et al. A. patagonica, P. papua and E. chrysocome, whe-
raptenodytinae and Palaeospheniscinae. 2006). This Biozone is characterized by ha- reas the islands south from New Zealand
However, our phylogenetic analysis was li- ving abundant penguin bones and the hold the most diverse colonies formed by
mited to twenty taxa (17 representative spe- phospatic brachiopod Lingula. Among pen- A. patagonica, P. papua, E. robustus, E. sclateri,
cies of all living genera and three fossils guins, Anthropornis nordenskjoeldi is numeri- E. chrysocome, E. schlegeli, Eudyptula minor and
species). cally predominant over the other species. Megadyptes antipodes. The coasts of the An-
One of the most peculiar quality of the An- Gadiforms, sharks and primitive mysticete tarctic Peninsula hold at present up to five
tarctic fossil fauna is the existence of giant whales are also part of the fossil assembla- species: A. forsteri, Pygoscelis papua, P. antarcti-
animals such us Anthropornis nordenskjoeldi in ge. Penguin bones are usually well preser- ca, P. adeliae and E. chrysolophus.
horizons that are dated as latest Eocene ved, complete, dissarticulated and with var- Current available data indicate that the sym-
associated with other small and medium- ying degree of abrasion, suggesting quiet patric diversity in the colonies is no higher
sized penguins (Myrcha et al. 2002) such us and low-energy depositation conditions. than four species (Wilson 1983). This is im-
Tonniornis sp. To mention a single example, The underlying stratigraphic members of portant for the evaluation of colony com-
Delphinornis arctowski is the smallest penguin the sequence show reworked fossil mate- position during the Cenozoic. We have alre-
recorded from the James Ross Basin. rials (Tambussi et al. 2006). ady mentioned that 14 species are recogni-
Throughout this contribution, we have Knowing "who the members are, how ma- zed for the late Eocene of Seymour Island,
mentioned a wealth of literature dedicated ny of them there are, how they interact, and whereas a lower amount is recognized for
to the study of the Antarctic fauna. Pen- how they collectively forge a workable" the Late Eocene - Oligocene of New Zea-
guins are not the exception and have been (Vermeij and Herbert 2004: 1) is necessary land (Ando, pers. comm. to CAH). There
the basis for vary contributions (Myrcha et to understand how an ancient ecosystem are at least three possible interpretations for
al. 2002, Tambussi et al. 2005, 2006, Jadwis- functioned. The macrofauna of the An- this fact: 1) the Cenozoic taxonomical di-
zczak 2003, 2006 and the literature cited thropornis nordenskjoeldi biozone is adequate versity in Antarctica and New Zealand are
therein). For that reason, here we will not to improve our comprehension of Eocene the highest so far recorded, 2) this diversity
provide in-depth treatment of these as- ecosystems. is biased due to problems in species identi-
pects, although we will refer to some syste- fication, or 3) the deposits are the product
matic and paleobiological issues. PALEOBIOLOGICAL of an asynchronous accumulation of bo-
IMPLICATIONS OF THE nes.
THE BIOSTRATIGRAPHIC RECORD One of the most outstanding peculiarities
IMPORTANCE OF THE of the Antarctic fossil fauna is the co-exis-
FOSSIL PENGUIN The importance of the findings of terres- tence of giant animals such as Anthropornis
ASSEMBLAGES trial birds in the study of the distribution nordenskjoeldi with other small and medium-
and origin of the birds has been previously sized penguins (Myrcha et al. 2002) in hori-
Our depiction of the diversity and abun- mentioned, as well as the significance of zons that are dated as latest Eocene. An-
dance of avian species is potentially distor- the Antarctic findings as indisputable pro- thropornis nordenskjoeldi is considered the lar-
ted by the artifacts imposed by the tapho- ofs of the presence of Neornithes in the gest penguin known whereas Delphinornis
nomic conditions that determine the as- age of dinosaurs. Beyond these facts, pen- arctowski is the smallest penguin recorded
semblages. But after many palaeontological guins are the most recognizable hallmarks from the James Ross Basin. The hydrodyna-
investigations on Seymour Island, we deem of the Antarctic avifauna. Based on their mic constraints of A. nordenskjoeldi suggestAntarctic birds (Neornithes) during the cretaceous-eocene times 611
that it was a rather slow swimmer that could brations imply a radiation of modern ocene could have affected the penguin po-
reach speeds of perhaps 7-8 km per hour (crown-group) birds in the Late Cretaceous pulations, because climatic changes are lin-
with no diving specializations (Tambussi et and a divergence of the modern sea-birds ked with habitat availability, and food web
al. 2006). and shore-birds lineages at least by the Late process. However, there is not evidence
In turn, Crossvallia seems to provide eviden- Cretaceous about 74 ± 3 Ma (Campanian). about the evolution of the Antarctic pen-
ce of independent acquisition of large size The current knowledge of the fossil An- guin after the end of the Eocene.
during the Late Paleocene - Late Eocene ti- tarctic birds is based on fragmentary, but
me span, probably under different environ- very informative, evidence. ACKNOWLEDGEMENTS
mental conditions (Tambussi et al. 2005), a - Antarctic fossil birds can be confidently
point of view accepted by Ksepka et al. assigned to modern orders and families. We thank Marcelo Reguero, Alberto Cione
2006). However, the evolution of penguin - Anseriformes (Clarke et al. 2005), ?Gavii- and Eduardo Tonni for inspiring discus-
body size is still unknown (Ksepka et al. formes loon-like (Chatterjee et al. 2006), sions of Antarctic birds over the past years.
2006). ?Gruiformes seriema-like (Case et al. 2006) This work was partially funded by CONI-
Studies on recent marine systems suggest are recorded before the K/T boundary. CET PIP 5694 Project to the authors. We
that most seabird species are constrained by - The Anseriforms Vegavis iaai from the late specially thank Sergio Marenssi for the
specific physical environmental features, in Cretaceous of Vega Island provides a well- opportunity to participate in this special
juxtaposition with nesting habitats. It is rea- definded calibration point for estimating volume.
sonable to believe that the progressive cli- the early divergence times of modern birds.
mate cooling during the Eocene would have - Two cursorial birds, a ratite and a phoror- WORKS CITED IN THE TEXT
directly or indirectly affected penguin po- hacid were recovered from the topmost
pulations, because climatic changes are lin- levels of the Submeseta Allomember Late Acosta Hospitaleche C., Tambussi C., Donato
ked with habitat availability and food web Eocene in age. Their discovery strongly sup- M. and Cozzuol M. 2007. A new Miocene
phenomena. ports the idea that West Antarctica was penguin from Patagonia and a phylogenetic
There is a gap in regard to the evolution of used as dispersal route for obligate terres- analysis of living and fossil species. Acta
the Antarctic penguin after the end of the trial organisms. Paleontologica Polonica 52: 299-314.
Eocene until the Pleistocene. - Representative birds of Falconiformes Alvarenga, H. and Höfling, E. 2003. Systematic
Polyborinae, Ciconiiformes, Charadriifor- revison of the Phorusrhacidae (Aves: Ralli-
CONCLUSIONS mes (including flamingos), Pelagornithidae formes). Papéis avulsos Zoologia 43: 55-91,
and Diomedeidae constitute the non-pen- São Paulo.
Available evidence indicates the existence guin avian assemblages of the Eocene of Aronson, R. and Blake, D. 2001. Global climate
of climatic fluctuations since the mid-Cre- La Meseta Formation. change and the origin of modern benthic
taceous up to the Paleogene beginnings - Fifthteen species of penguins have been communities in Antarctica. American Zoolo-
characterized by a warming phase followed described including the oldest penguin of gist 41: 27-39, Seattle.
by a colder one, and a conspicuous Paleo- West Antarctica, Croswallia unienwillia (Tam- Baumel, J. and Witmer, L.M. 1993. Osteologia.
cene-Eocene thermal maximum and a pro- bussi et al. 2005). In Baumel, J., King, A., Breazile, J., Evans, H.
gressive cooling through the Cenozoic - The Anthropornis nordenskjoeldi Biozone and Vanden Berge, J. (eds.) Handbook of
(Francis et al. 2006a). (36.13 and 34.2 Ma, late Late Eocene, avian anatomy: Nomina Anatomica Avium.
The Eocene represents a period of climate Tambussi et al. 2006) is characterized by the Publications of the Nuttall Ornithological
transition from global warmth to progressi- high frequency of penguin bones and the Club, 401 p., Massachusetts.
ve cooling, culminating in the initiation of phosphatic brachiopod Lingula. Five species Birkenmajer, K., Gadzicki, A., Krajewski, K.,
Antarctic glaciation. The incidence of these Anthropornis nordenskjoeldi, Delphinornis graci- Przybycin, A., Solecki, A., Tatur, A. and Yo-
climatic changes on the faunas produces di- lis, D. arctowski, Archaeospheniscus lopdelli, and on, H. II 2004. First Cenozoic glaciers in West
fferent consequences including both extinc- Palaeeudyptes antarcticus are exclusively for Antarctica. Polish Polar Research 26: 3-12.
tions and origin of groups. this unit. Bourdon, E. 2005. Osteological evidence for sis-
Several molecular phylogenetic studies are - Within the fossil penguins of the James ter group relationship between pseudo-too-
predicting Cretaceous or earlier origins of Ross Basin, Anthropornis nordenskjoeldi was thed birds (Aves: Odontopterygiformes) and
modern taxa, some of them occurred in evidently the largest, whereas Delphinornis waterfowls (Anseriformes). Naturwissen-
southern high latitudes. Unambiguous e- arctowski is the smallest. schaften 92: 586-591.
xamples of this are penguins whose fossil - One of the worldwide highest morpholo- Case, J., Reguero, M., Martin, J. and Cordes-Per-
record begins at the Late Paleocene (Slack et gical and taxonomic penguins diversity, son, A. 2006. A cursorial bird from the Maas-
al. 2006, Tambussi et al. 2005), which provi- including giant and tiny species, is docu- trictian of Antarctica, 2006 Society of verte-
des a lower estimate of 61-62 Ma for the mented at the topmost levels of the La brate paleontology. Journal of Vertebrate Pa-
divergence between penguins and related Meseta Formation. leontology 26: 48A.
flying birds (Slack et al. 2006). Penguin cali- - The progressive climate cooling of the E- Case, J., Woodbourne, M. and Chaney, D. 1987.612 C. TA M BU S S I A N D C. AC O S TA H O S P I TA L E C H E
A gigantic phororhacoid (?) bird from An- Ross Basin, Antarctica, The Geological So- of Eocene Fossiliferous Erratics, McMurdo
tartica. Journal of Paleontology 61: 1280-1284. ciety, Special Publications 7-19, London. Sound, East Antarctica, American Geophy-
Césari, S., Marenssi, S. A. and Santillana, S. 2001. Crame, J., Pirrie, D., Riding, J. and Thomson, M. sical Union, Antarctic Research Series 76: 253
Conifers from the Upper Cretaceous of Cape 1991. Campanian-Maastrichtian (Cretaceous) -260.
Lamb, Vega Island, Antarcctica. Cretaceous stratigraphy of the James Ross Island area, Francis, J., Crame, J. and Pirrie, D. 2006a. Cre-
Research 22: 309-319. Antarctica. Journal of the Geological Society taceous-Tertiary high latitude palaeonviron-
Chatterjee, S. 1989. The oldest Antarctic bird. 148: 1125-1140, London ments, James Ross Basin, Antarctica: Intro-
Journal of Vertebrate Paleontology 9: 16A. Del Valle, R.A., Elliot, D.H. and Thomson, duction. In: J.E. Francis, Crame, J.A. and Pi-
Chatterjee, S. 1997. The rise of birds. John Hop- M.R.A. 1992. Sedimentary basins on the east rrie, D. (eds.) Cretaceous-Tertiary high latitu-
kins University Press, Baltimor. flank of the Antarctic Peninsula: proposed de palaeonvironments, James Ross Basin,
Chatterjee, S. 2002. The Morphology and nomenclature. Science 4: 477-478. Antarctica. The Geological Society, Special
Systematics of Polarornis, a Cretaceous Loon Dingle, R., S. and Lavelle, M. 1998. Late Cre- Publication 258: 1-6, London.
(Aves: Gaviidae) from Antarctica). In Zhang, taceous-Cenozoic climatic variations of the Francis, J., Crame, J. and Pirrie, D. (eds.) 2006b.
Z.Z. (Editor), 5º Symposium of the Society northern Antarctic Peninsula: new geochemi- Cretaceous-Tertiary high latitude palaeonvi-
of Avian Paleontology and Evolution Beijing cal evidence and review. Palaeogeography Pa- ronments, James Ross Basin, Antarctica. Ge-
2000, Science Press 125-155, Beijing. laeoclimatology Palaeoecology 141: 215-232. ological Society London 258: 201 p., London.
Chatterjee, S., Martinioni, D., Novas, F., Mussel, Dutton, A.L., K., C Lohmann, K.C. and Zins- Gadzicki, A. 2004. Eocene-Pliocene ecosystems
F. and Templin, R. 2006. A new fossil loon meister, W.J. 2002. Stable isotope and minor of South Shetlands and Antarctic Peninsula.
from the late cretaceous of Antarctica and element proxies for Eocene climate of Sey- Bollettino di Geofisica Teorica ed Applicata
early radiation of foot-propelled diving birds. mour Island, Antarctica. Paleoceanography 45: 258-260.
Journal of Vertebrate Paleontology 26: 49A. 17: 1016-1029. Gadzicki, A., Gruszczynski, M., Hoffman, A.,
Clarke, J., Tambussi, C.P., Noriega, J., Erickson, Dzik, K. and Gadzicki, A. 2001. The Eocene Malkowski, K., Marenssi, S., Haias, S. and Ta-
G. and Ketcham, R. 2005. Definitive fossil expansion of nautilids to high latitudes. Pa- tur, A. 1992. Stable carbon and oxygen isoto-
evidence for the extant avian radiation in the laeogeography Palaeoclimatology Palaeoe- pe record in the Paleogene La Meseta For-
Cretaceous. Nature 433: 305-308. cology 17: 279-312. mation, Seymour Island, Antarctica. Antarc-
Clarke, J.A., Olivero, E.B. and Puerta, P. 2003. Ehrmann, W. and Mackensen, A. 1992. Sedi- tic Science 4: 461-468.
Description of the earliest fossil penguin mentological evidence for the formation of González-Barbaa, G., Schwennickea, T., Goe-
from South America and the first Paleogene an East Antarctic ice sheet in Eocene/ dertb, J. and Barnes, L. 2002. Earliest pacific
vertebrate Locality of Tierra del Fuego, Oligocene time. Palaeogeography Palaeocli- basin record of the pelagornithidae (Aves:
Argentina. American Museum Novitates matology Palaeoecology 93: 85-112. Pelecaniformes). Journal of Vertebrate
3423: 1-9. Elliot, D. and Trautman, T. 1982. Lower Tertiary Paleontology 22: 722-725.
Cooper, A., Penny D., 1997. Mass survival of strata on Seymour Island, Antarctic peninsu- Harrison, C. and Walker, C. 1976. A review of
birds across the Cretaceous-Tertiary boun- la. In Craddock, C. (ed.) Antarctic Geoscien- the bony-toothed birds (Odontopterygifor-
dary: molecular evidence. Science 275: 1109- ce, University of Wisconsin Press 287-297, mes) with descriptions of some new species.
1113. Madison Backhuys Publishers, Tertiary Research
Covacevich, V. and Lamperein, C. 1972. Hallaz- Feduccia, A. 1995. Explosive radiation in Tertia- Special Paper 2: 1-72.
gos de icnitas en Península Fildes, Isla Rey ry birds and mammals. Science 267: 637-638. Hathway, B. 2000. Continental rift to back-arc
Jorge, Archipiélago Shetland del Sur, Antár- Feduccia, A. 2003. "Big Bang" for Tertiary Birds? basin: Jurassic-Cretaceous stratigraphical and
tica. Serie Científica INACH (Instituto An- Trends in Ecology and Evolution 18: 172- structural evolution of the Larsen basin An-
tártico Chileno) 1: 55-74. 176. tarctic Peninsula. Journal of the Geological
Covacevich, V. and Rich, P.V. 1982. New birds Feldmann, R. and Woodbourne, M. 1988. Geo- Society 157: 417-432.
ichnites from Fildes Peninsula, King George logy and paleontology of Seymour Island, Hayes, P., Francis, J. and Crame, J.A. 2006. Pa-
Island, West Antarctica. In Craddock, C. (ed.) Antarctic Peninsula. The Geological Society laeoclimate analysis of Late Cretaceous an-
The University of Wisconsin Press: 245-254, of America, Memoir 169: 1-566. giosperm leaf floras, James Ross Island, An-
Madison. Francis, J. 1996. Antarctic palaeobotany: Clues to tarctica, Cretaceous-Tertiary high latitude pa-
Coxall, H., Wilson, P., Palike, H., Lear, C. and climate change. Terra Antartica 3: 135-140. laeonvironments, James Ross Basin, Antarc-
Beckman, J. 2005. Rapid stepwise onset of Francis, J.E. 1999. Evidence from fossil plants tica. In: J.E. Francis, Crame, J.A. and Pirrie,
Antarctic glaciation and deep calcite compen- for Antarctic palaeoclimates over the past D. (eds.) Cretaceous-Tertiary high latitude pa-
sation in the Pacific Ocean. Nature 422: 53-57. 100 million years. Terra Antartica Reports 3: laeonvironments, James Ross Basin, Antarc-
Crame, J., Pirrie, D. and Riding, J. 2006. Mid. 43-52. tica. The Geological Society, Special Publi-
Cretaceous stratigraphy of the James Ross Francis, J.E. 2000. Fossil wood from Eocene cation 258: 49-62. London.
Basin, Antarctica. In Francis, J. E., Pirrie, D., high latitude forests McMurdo Sound An- Hedges, B.S., Parker, P., Sibley, C., Kumar, S.
and Crame, J. A. (eds.). Cretaceous - Tertiary tarctica. In: Stilwell, J.D. and Feldmann, R.M. 1996. Continental breakup and the ordinal
High Latitude Palaeonvironments, James (eds) Palaeobiology and Palaeoenvironments diversification of birds and mammals. NatureAntarctic birds (Neornithes) during the cretaceous-eocene times 613
381: 226-229. Geología de cabo Lamb, isla Vega, Antártida. mes) from Vega Island, Antactic Peninsla:
Hope, S. 2002. The Mesozoic radiation of Neo- Contribuciones Científicas Instituto Antár- paleobiogeographic implications. Ameghi-
rnithes. In L.M. Chiappe and L.M. Witmer tico Argentino 530: 1-43. niana 32: 57-61.
(Editors), Mesozoic Birds: Above the Heads Marenssi, S., Santillana, S. and Rinaldi, C.A. Noriega, J. and Tambussi, C. 1996. The non pen-
of Dinosaurs. University California Press: 1998a. Paleoambientes sedimentarios de la A- guin avifauna from the Eocene (early Oli-
339-388, Berkeley loformación La Meseta (Eoceno), isla Ma- gocene?) of Seymour Island, Antarctic Pe-
Ivany, L., Blake, F., Lohmann, K. and Aronson, rambio (Seymour), Antártida, Instituto An- ninsula. Fourth International Meeting of the
R. 2004. Eocene cooling recorded in the che- tártico Argentino, Contribución 464, 51 p., Society of Avian Paleontology and evolution.
mistry of La Meseta Formation mollusks, 24 figs., Buenos Aires. Abstracts: 13, Washington.
Seymour Island, Antarctic Peninsula. Bolle- Marenssi, S., Santillana, S. and Rinaldi, C. 1998b. Olivero, E., Martinioni, D. and Mussel, F. 1992.
ttino di Geofisica Teorica ed Applicata 45: Stratigraphy of the La Meseta Formation Sedimentología y bioestratigrafía del cretáci-
242-245. (Eocene), Marambio (Seymour) Island, An- co superior del oeste de Cabo Lamb (Isla
Jadwiszcak, P. 2001. Body size of Eocene tarctica. In: S. Casadío (ed.), Paleógeno de A- Vega, Antartida). Implicancias sobre ciclos
Antarctic penguins. Polish Polar Research 22: mérica del Sur y de la Península Antártica. sedimentarios y evolución de la Cuenca. In
147-158. Asociación Paleontológica Argentina, Publi- Rinaldi, C.A. (ed.) Geología de la Isla James
Jadwiszczak, P. 2003. Eocene penguins of Sey- cación Especial: 137-146. Ross, Publicación Especial Instituto Antár-
mour Island: systematics, evolution and pale- Martin, J. 2006. Biostratigraphy of the Mosasau- tico Argentino: 125-145, Buenos Aires.
oecology, 9th International Symposium on ridae (Reptiles) from the Cretaceous of An- Olson, S. 1985. The fossil record of bird. In Far-
Antarctic Earth Sciences (ISAES IX) Abs- tarctica. In Francis, J. E., Pirrie, D., and Cra- ner, D.S., King, J.R., Parkes, K.C. (eds.) Avian
tracts: 169, Potsdam. me, J. A. (eds.) Cretaceous - Tertiary High La- Biology, Academic Press: 79-238, New York.
Jadwiszczak, P. 2006: Eocene penguins of titude Palaeonvironments, James Ross Basin, Olson, S. 1992. Neogaeornis wetzeli Lambrecht,
Seymour Island, Antarctica: Taxonomy. Po- Antarctica. Geological Society, Special Publi- a Cretaceous loon from Chile (Aves, Gavii-
lish Polar Research 27: 3-62. cations 258: 101-108, London. dae). Journal of Vertebrate Paleontology 12:
Kandefer, H.M. 1994. Róznorodnosc fauny Mayr, G. 2004. A partial skeleton of a new fossil 122-124.
pingwinów kopalnych antarktycznej Wyspy loon (Aves, Gaviiformes) from the early Oli- Peters, D. 1987. "Phorusrhacide" aus dem Mit-
Seymour w oparciu o analize humeri z ko- gocene of Germany with preserved stomach tel-Eoza¨n von Messel (Aves: Gruiformes:
lekcji. Magister Thesis, Warszawski Univer- content. Journal of Ornithology 45: 281-286. Cariamae). Documents des Laboratoires de
sity, 124 p., Warszaw. Mayr, G. 2005. ''Old World phorusrhacids'' (A- Geologie de Lyon 99: 71-87.
Kennett, J. and Warnke, D. 1993. The Antarctic ves, Phorusrhacidae) - a new look at Stri- Pirrie, D., Crame, J., Lomas, S. and Riding, J.
paleoenvironment: a perspective on global gogyps (''Aenigmavis'') sapea (Peters 1987). 1997. Late Cretaceous stratigraphy of the
change, Part Two, 60, 273 p. Paleobios 25: 11-16 Admiralty Sound Region, James Ross Basin,
Ksepka, D., Bertelli, S. and Giannini, N. 2006. Mayr, G., Manegold, A. and Johansson, U.S. Antarctica. Cretaceous Research 18: 109-137.
The phylogeny of the living and fossil Sphe- 2003. Monophyletic groups within "higher Pirrie, D., Crame, J. and Riding, J. 1991. Late Cre-
nisciformes (penguins). Cladistics 22: 412. land birds" - comparison of morphological taceous stratigraphy and sedimentology of
MacFadden, B., Hulbert, J. and Baskin, J. 2006. and molecular data. Journal Zoology and Cape Lamb, Vega Island, Antarctica. Cre-
Refinded age of the Late Neogene Terror Evolutionary Research 41: 233-248. taceous Research 12: 227-258.
bird (Titanis) from Florida and Texas using McKee, J. 1985. A pseudodontorn (Pelecanifor- Poole, I., Hunt, R.J. and Cantrill, D.J. 2001. A
rare earth elements. Journal of Vertebrate mes: Pelagornithidae) from the middle Pli- Fossil Wood Flora from King George Island:
Paleontology 26: 92A. ocene of Hawera, Taranaki, New Zealand. Ecological Implications for Antarctic Eocene
Mackensen, A. and Ehrmann, W. 1992. Middle New Zealand. Journal of Zoology 12: 181- Vegetation. Annales of Botany 88: 33-54.
Eocene through Early Oligocene climate his- 184, London. Reguero, M.A., Marenssi, A.M. and Santillana,
tory and paleoceanography in the Southern Mourer-Chauviré, C. 1981. Première indication S.N. 2002. Antarctic Peninsula and South A-
Ocean: stable oxygen and carbon isotopes de la présence de phorusrhacidés, famille merica (Patagonia) Paleogene terrestrial fau-
from ODP sites on Maud Rise and Ker-gue- d'oiseaux géants d'Amérique du Sud, dans le nas and environments: biogeographic rela-
len Pateau. Marine Geology 108: 1-27. Tertiaire européen: Ameghinornis nov. gen. tionships. Palaeogeography, Palaeoclima-
Marenssi, S.A. 2006. Eustatically controlled sedi- (Aves, Ralliformes) des phosphorites du tology, Palaeoecology 179: 189-210.
mentation recorded by Eocene strata of the Quercy, France. Geobios 14: 637-647. Salamy, K. and Zachos, J. 1999. Latest Eocene-
James Ross Basin, Antarctica. In Francis, J. Myrcha, A., Jadwiszczak, P. Tambussi, C., Early Oligocene climate change and Sou-
E., Pirrie, D., and Crame, J. A. (eds.) Cretace- Noriega, J. Gazdzicki, A., Tatur, A., del Valle, thern Ocean fertility: inferences from sedi-
ous - Tertiary High Latitude Palaeonviron- R. 2002. Taxonomic revision of Eocene ment accumulation and stable isotope data.
ments, James Ross Basin, Antarctica. Geolo- Antarctic penguins based on tarsometatarsal Palaeogeography, Palaeoclimatology, Palaeoe-
gical Society, Special Publications 258: 125- morphology. Polish Polar Research 23: 5-46. cology 145: 61-77.
133, London. Noriega, J. and Tambussi, C. 1995. A late Cre- Sibley, C.G., Ahlquist, J.E. and Monroe, B.L.
Marenssi, S., Salani, S. and Santillana, S. 2001. taceous Presbyornithidae (Aves Anserifor- 1988. A classification of the living birds of614 C. TA M BU S S I A N D C. AC O S TA H O S P I TA L E C H E
the world based on DNA- DNA hybridiza- Science 330: 105-114, Los Angeles. Falconiformes Polyborinae
tion studies. Auk 105: 409-423. Tonni, E. and Tambussi, C. 1985. Nuevos restos MLP 95-I-10-8 (distal fragment of tarsometatar-
Simpson, G.G., 1946. Fossil penguins. Bulletin A- de Odontopterygia (Aves: Pelecaniformes) en sus)
merican Museum Natural History 87: 1-100. el Terciario temprano de Antártida. Ameghi- Ocurrence Cucullaea I Allomember (Noriega
Slack, K., Jones, C., Tatsuro, A., Harrison, G. niana 21: 121-124. and Tambussi, 1996)
Fordyce, E., Arnason, U. and Penny, D. 2006. Triche, A. 2006. Origin and completeness of the Gruiformes ? Phorusrhacidae
Early Penguin Fossils, plus Mitochondrial penguin fossil record. Journal of Vertebrate UCR 22175 Cast, distal end of bill; distal half of
Genomes, Calibrate Avian Evolution. Mo- paleontology 26: 132A. tarsometatarsus
lecular Biology and evolution 23: 1144-1155. Van Tuinen, M. 2002. Relationships of birds - Ocurrence Submeseta Allomember (Case et al.
Tambussi, C., Acosta Hospitaleche, C., Reguero, molecules versus morphology. In Encyclo- 1987, 2006)
M. and Marenssi, S. 2006. Late Eocene pen- pedia of Life Sciences, Special Essay Article. Charadriiformes indet.
guins from West Antarctica: Systematic and Macmillan Publishers Ltd., Nature Publi- MLP 92-II-2-6 (right scapula)
biostratigraphy. Cretaceous Tertiary High-la- shing Group: 1-7. Ocurrence Cucullaea I Allomember (Noriega
titude palaeoenvironments, James Ross Ba- VanTuinen, M. 2006. Tempo and mode of mo- and Tambussi, 1996)
sin, Antarctica. In Francis, J. E., Pirrie, D., dern bird evolution observed with large-scale ?Phoenicopteridae
and Crame, J. A. (eds.) Cretaceous - Tertiary taxonomic sampling. Historical Biology 18: MLP 87-II-1-2 (incomplete right radius)
High Latitude Palaeonvironments, James 205-221. Ocurrence Cucullaea I Allomember (Noriega
Ross Basin, Antarctica. Geological Society, Vermeij, G. and Herbert, G. 2004. Measuring re- and Tambussi, 1996)
Special Publications 258: 145-162. lative abundance in fossil and living assem- Ciconiiformes indet.
Tambussi, C.P. and Noriega, J.I., 1996. Summary blages. Paleobiology 30: 1-4. MLP 90-I-20-9 distal fragment of right tarsome-
of the Avian Fossil Record from Southern Vizcaíno, S., Reguero, M., Goin, F., Tambussi, C. tatarsus.
South America. In Arratia, G. (ed.) Contri- and Noriega, J. 1998. Community structure Occurrence Submeseta Allomember (Noriega
butions of southern South America to verte- of Eocene terrestrial vertebrates from An- and Tambussi, 1996)
brate paleontology. Müncher Geowissens- tarctic Peninsula. Ameghiniana, Publicacion Procellariidae indet.
chaftliche Abhandlungen: 245-264, Munich. Especial 5: 177-183. MLP 88-I-1-5 (incomplete tarsometatarsus),
Tambussi, C., Noriega, J., Gazdzicki, A., Tatur, Walsh, S.A. and Hume, J. 2001. A new Neogene MLP 95-I-10-14 (left coracoid), MLP 96-I-5-8
A., Reguero, M. and Vizcaino, S. marine avian assemblage from North-Central (distal end of rostrum). MLP 91-II-4-6 (distal
1994. The first occurrence of a Ratite bird Chile. Journal of Vertebrate Paleontology 21: fragment of ulna).
(Pycraft) in Seymour Island, Antarctica. 484-491. Occurrence Submeseta Allomember (Noriega
Polish Polar Research 15: 15-20. Warheit, K. 1992. A review of the fossil seabirds and Tambussi, 1996)
Tambussi, C., Noriega, J., Santillana, S. and Ma- from the Tertiary of the North Pacific: plate Diomedeidae
renssi, S. 1995. Falconid bird from the middle tectonics, paleoceanography, and faunal MLP 88-I-1-6 (distal end of rostrum)
Eocene La Meseta Formation, Seymour Is- change. Paleobiology 18: 401-424. Occurrence Submeseta Allomember (Noriega
land, West Antarctica. Journal of Vertebrate Wilson, G.J. 1983. Distribution an abundance of and Tambussi, 1996)
Paleontotogy 15: 55A. antarctic and sub-antarctic pinguins: a Anseriformes ?Presbyornithidae
Tambussi, C., Reguero, M., Marenssi, S. and synthesis of current knowledge. SCAR and MLP 96-I-5-19 (proximal end of scapula), MLP
Santillana, S. 2005. Crossvallia unienwillia, a SCOR, Biomass Scientific Series: 1-43, 95-I-10-9 (proximal fragment of scapula), MLP
new Spheniscidae (Sphenisciformes, Aves) Cambridge. 96-I-5-7 (ulna)
from the Late Paleocene of Antarctica. Geo- Zachos, J., Lohmann, K., Walker, J. and Wise, J. Ocurrence Cucullaea I Allomember (Noriega
bios 38: 557-675. 1993. Abrupt climate change and transient and Tambussi, 1996)
Tambussi, C. and Tonni, E. 1988. Un Diome- climates during the Paleogene: A marine Vegavis iaai Clarke, Tambussi, Noriega,
deidae (Aves:Procellariiformes) del Eoceno perspective. Journal of Geology 101: 191- Erickson & Ketcham, 2004
tardío de Antártida. In Quiroga, J. and Cione, 213. MLP 93-I-1-3 (disarticulated partial postcranial
A. (eds.) 5º Jornadas Argentinas de Paleon- skeleton preserved in two halves of a concretion:
tología Vertebrados, Resúmenes 4, La Plata. APPENDIX I. five thoracic vertebrae, two cervical vertebrae,
Tambussi, C., Ubilla, M. and Perea, D. 1999. The left scapula, right ulna, pelvic bones, right and
youngest large carnassial bird (Phorusrhaci- Taxonomical and anatomical identification of left fibulae, right humerus, proximal left hume-
dae, Phorusrhacinae) from South America fossil birds materials from Antarctica. rus, right coracoid, femora, left tibiotarsus, distal
(Pliocene-early Pleistocene of Uruguay). right radius, sacrum, left tarsometatarsus, proxi-
Jour-nal of Vertebrate Paleontology 19: 406- Ratitae indet. mal right tarsometarsus and more than six dorsal
408. MLP 94-III-15-1 (distal fragment of right tarso- ribs).
Tonni, E.P. 1980. The present state of knowled- metatarsus) Ocurrence Unit K3 (upper part of the Cape
ge of the Cenozoic birds of Argentina. Na- Occurrence Submeseta Allomember (Tambussi Lamb Member and the Sandwich Bluff Member
tural History Museum, Contributions in et al., 1994) of the López de Bertodano Formation, of PirrieYou can also read

















































