Cicada Thermoregulation (Hemiptera, Cicadoidea)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Cicada Thermoregulation (Hemiptera, Cicadoidea)
A.F. SANBORN
Abstract
A review the mechanisms and ther-
moregulatory strategies used by cicadas is
presented. The behavioral and physiologi-
cal processes used to regulate body tempe-
rature are discussed. Behavioral strategies
include changes in body orientation to the
sun, basking, shade-seeking, microclimate
selection, vertical migration, using the
wings as a parasol, and suspension of
activity. The effect of temperature on the
biology of cicadas and the strategies used
to deal with temperature are discussed.
Physiological responses include thermal
adaptation, endothermy, and evaporative
cooling.
Key words: temperature regulation,
endothermy, evaporative cooling, thermal
responses.
Denisia 04,
zugleich Kataloge des OÖ. Landesmuseums,
Neue Folge Nr. 176 (2002), 455-470
455
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Temperature is a physical parameter of the Temperature has been shown to affect
environmental that affects all organisms at the cicadas at all levels, from controlling the rate
cellular level through the temperature depen- of neural firing (WAKABAYASHI & HAGIWARA
dent nature of chemical reactions. The daily 1953, WAKABAYASHI & IKEDA 1961) or the
fluctuations in ambient temperature (T a ) can contraction kinetics of muscles (AlDLEY &.
have a significant influence on the activity of WHITE 1969, JOSEPHSON 1981, JOSEPHSON &
YOUNG 1979, JOSEPHSON & YOUNG 1985,
any terrestrial animal. Animals have two opti-
SANBORN in press) to determining the rate of
ons with respect to body temperature (Tu,)
development (AZUMA 1976, HARTZELL 1954,
and the changing thermal environment: they
NAGAMINE and TERUYA 1976), when a popu-
lation emerges (HEATH 1968) or even the dis-
tribution of the species (OHGUSHI 1954,
SCHEPL 1986, TOOLSON 1998). Therefore,
temperature and the regulation of body tem-
perature (Tu,) are as essential in the lives of
cicadas as in any other terrestrial animal.
Previous studies (HEATH 1967, HEATH
1972, SANBORN et al. 1995a, SANBORN et al.
1995b, SANBORN 1997, SANBORN 2000, SAN-
RORN &. MATE 2000) have shown that cicadas
must maintain their Tu, within a small range
to coordinate reproductive activity, the main
purpose of the adult life stage. The daily varia-
tions of T a are large enough to prevent the
passive development of a constant Tu,, but it is
not always practical for an organism to delay
activity until ambient conditions are favorable
while spending the remainder of the day in a
thermal shelter. Cicadas, therefore, spend
time and energy thermoregulating to permit
activity for a significant portion of the day.
The strategies used by animals to regulate
Tu, can be divided into two major types of res-
ponses: behavioral and physiological. A com-
mon method of thermoregulation in cicadas is
to use behavioral mechanisms to alter the
uptake of solar radiation (Heath 1967, HEATH
& WILKIN 1970, HEATH et al. 1972, HEATH
Figs. 1: can be thermoconformers or thermoregula- 1972, HASTINGS 1989, HASTINGS & TOOLSON
Basking Tympanoterpes elegans BERG. tors. A thermoconformer is an animal whose 1991, TOOLSON 1998, SANBORN 2000,
Cicadas will bask at low body and/or
ambient temperature to elevate body Tu, changes with and is approximately equal to SANBORN & MATE 2000) (Fig. 1). This type of
temperature to a range necessary to T a . Any animal that is a thermoconformer is organism is termed ectothermic because it uses
coordinate activity. Cicadas maximize unable to function efficiently physiologically exogenous heat to regulate Tu,. However, the-
radiant energy uptake by orienting
the greatest body surface area to the over a wide temperature range because enzym- re have been studies (SANBORN et al. 1995a,
sun. The dorsal surface is perpendicu- es function most efficiently over a restricted SANBORN et al. 1995b, SANBORN 1997, SAN-
lar to the sun in this photograph. temperature range. The alternative strategy BORN 2000) showing cicadas using the physio-
that reduces the negative aspect of fluctuating logical mechanisms of endothermy (the gene-
Tu, is thermoregulation. An animal that ther- ration of endogenous heat for thermoregulati-
moregulates maintains Tu, within a "preferred" on) and evaporative cooling (KASER &
temperature range so its enzymes are functio- HASTINGS 1981, TOOLSON 1984, TOOLSON
ning near optimal levels while the animal is 1985, TOOLSON 1987, TOOLSON & HADLEY
active. 1987, HADLEY et al. 1989, HASTINGS 1989,
456
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
STANLEY-SAMUELSON et al. 1990, HADLEY et Similar patterns of movement and exposu-
al. 1991, HASTINGS & TOOLSON 1991, re to solar radiation occur in endothermic spe-
SANBORN et al. 1992, TOOLSON 1993, TOOLSON cies. Endothermic species use radiant heat
et al. 1994) in the field to regulate T^. when it is available to assist in the elevation of
Tjj. This behavioral strategy saves the animals
Behavioral Responses metabolic energy that can then be used for
activity at a time when solar radiation is not
All animals use behavioral thermoregula- available (SANBORN et al. 1995a, SANBORN et
tion as a first option to regulate T^. Behavi- al. 1995b, SANBORN 2000). This behavior may
oral thermoregulation is the quickest and most also potentially increase the life span of the
accessible mode of regulation since the effect individual adult since the adults do not recei-
is immediate and behavioral temperature ve significant amounts of energy from the
regulation does not require changes in cellular xylem fluid on which they feed (CHEUNG &
composition or activity to be effective. There- MARSHALL 1973a).
fore, it is not surprising that the majority of
We had the opportunity to see importance
cicadas regulate T^ through behavioral
of basking as a means to elevate T(j to a level
mechanisms (HEATH 1967, HEATH &. WlLKlN
necessary for activity during a partial (about
1970, HEATH et al. 1972, HEATH 1972,
70%) solar eclipse in 1991 (SANBORN & PHIL-
HASTINGS 1989, HASTING & TOOLSON 1991,
LIPS 1992). T a decreased by approximately
SANBORN et al. 1992, SANBORN et al. 1995a,
1°C during the eclipse but remained at a level
SANBORN et al. 1995b, SANBORN 1997,
where activity had been recorded previously
TOOLSON 1998, SANBORN 2000, SANBORN &
in the species active in the habitat (SANBORN
MATE 2000). et al. 1992). The cicadas were forced to sus-
As stated previously, the majority of cica- pend activity due to a loss of radiant input,
das use solar radiation as a mechanism to ele- which is required to maintain an elevated T(j.
vate Tjj. Cicadas move from their nocturnal A similar effect has also been reported several
feeding locations to perches where their expo- times with decreased radiation due to cloud
sure to the sun will be maximal early in the cover (see references in SANBORN & PHILLIPS
morning. Cicadas bask in order to keep T^ 199).
elevated to a range necessary for activity. A common mechanism used by ectother-
Cicadas have been shown to congregate in mic animals to decrease T^ is to decrease the
areas where exposure to the sun is greatest radiant heat load. A simple behavior animals
(HUDSON 1890, RAMSAY 1959, HEATH 1967, can employ to decrease radiant heat gain is to
HEATH & WILKIN 1970, HEATH et al. 1972, move into a shaded location. This strategy is
HEATH 1972, FLEMING 1975, YOUNG 1975, widely used by cicadas exposed to elevated T a
YOUNG 1980, YOUNG 1981, JOERMANN & and radiant energy from the sun to regulate TJL,
SCHNEIDER 1987, HASTINGS 1989, HASTINGS (HEATH 1967, HEATH & WILKIN 1970, HEATH
& TOOLSON 1991, SANBORN et al. 1992, et al. 1972, HASTINGS, 1989, HASTINGS &
TOOLSON 1998, SANBORN 2000, SANBORN & TOOLSON 1991, SANBORN et al. 1992, SAN-
MATE 2000). The concentration of animals BORN et al. 1995a, TOOLSON 1998, SANBORN
moves within the habitat to stay exposed to 2000, SANBORN & MATE 2000). Cicadas also
the sun as the sun moves through the day change the location of activity from the exter-
(HEATH 1967, SANBORN et al. 1995a, DURIN nal regions of plants to inner shaded locations
1981). These generalization are true of both as T a or T^ increase. This is an especially
ectothermic (HEATH 1967, HEATH & WlLKlN important strategy in active animals whose
1970, HEATH et al. 1972, HEATH 1972, metabolism would further increase T^.
HASTINGS 1989, HASTINGS & TOOLSON 1991, Shade-seeking behavior has been measu-
SANBORN et al. 1992, TOOLSON 1998, red directly in the field. HEATH (1967) measu-
SANBORN 2000, SANBORN & MATE 2000) and red the Tjj when animals voluntarily moved to
endothermic species (SANBORN et al. 1995a, the shade. He demonstrated cicadas moved
SANBORN et al. 1995b, SANBORN 2000). from sun to shade over a very narrow range of
457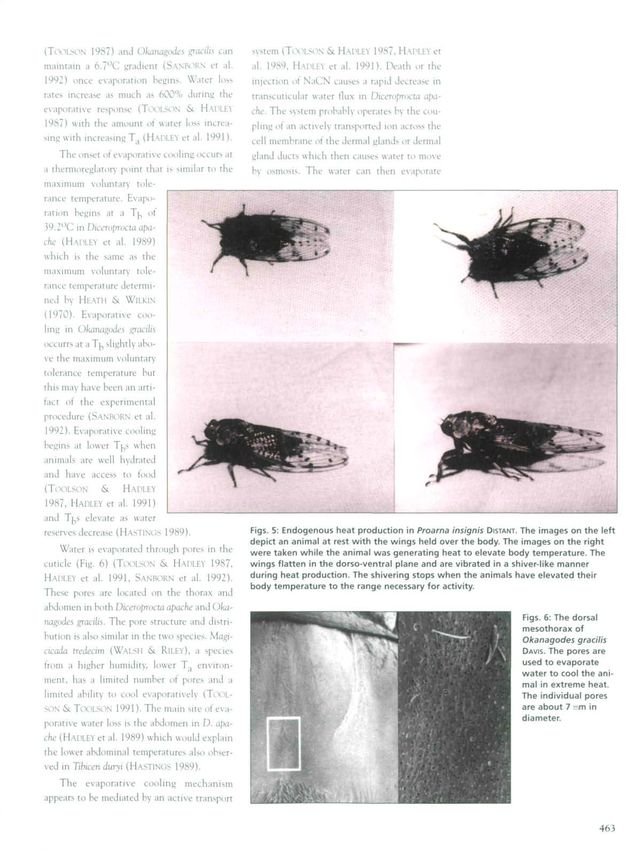
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Tj, (1.5-2.O°C). In addition, the T^ range of changes of Tj, in Cacama valvata (UHLER)
the laboratory shade-seeking response corre- (HEATH et al. 1972).
sponded to the maximum T^,s measured in the Changing the orientation of the body can
field suggesting a finely tuned response is con- also alter radiant heat uptake if the animal has
trolling behavioral changes that regulate T^ differences in the reflective pattern ot diffe-
(HEATH 1967). rent parts of the body. Animals such as Caca-
Animals can also regulate the amount of ma valvata (HEATH et al. 1972) and Okanagu-
radiant heating by changing the orientation o( des graci/is DAMS (SANBORN et al. 1992) have
their body to the sun. This will change the a morphological adaptation that assists in
decreasing radiant input at elevated Tj,. The
ventral surface of these, and many other desert
inhabiting species, is white in color (Fig. 3).
The difference in coloration between the dor-
sal and ventral surfaces facilitates reducing the
radiant heat gain when the animals become
negatively oriented to the sun as the white
ventral surface reflects a greater portion of the
radiant heat. Animals from less thermal rigo-
mus habitats do not have this type of reflec-
tive ventral surface (HEATH 1967, HEATH
1972).
The periodical cicada Ma^dcada cassinii
(FlSHER) has been shown to adopt an unusual
posture at low T., and when the sun is low on
the horizon (HEATH 1967). The wings are
spread away from the midline exposing a grea-
ter area of the black thorax and abdomen to
solar radiation. By the late morning, the wings
have returned to their usual position, peaked
over the midline of the abdomen. This stra-
tegy acts to increase the area of the body expo-
sed to radiant heating at low T a , thus increa-
sing the heating rate. Similarly when T^, T a
or the sun angle is elevated the wings act as a
parasol decreasing the radiant heat load and
the temperature excess of the animal.
There is also some evidence that cicadas
use the wings as a shade in a different mannet.
Active Magiacada cassinii orient their bodies
with the head facing directly away from the
Figs. 2: surface area of the body being exposed to the sun in the aftermxin. This behavior permits
A specimen of Cacama valvata (UHLER) the wings to "shade" the body and decrease
sun which will alter the heat transfer. As Tj,
that has partially rotated around its heat uptake (HEATH 1967).
perch. The radiant heat load decreases increases animals can decrease the total surfa-
when the animal exposes the lateral ce area exposed to the sun through three main Selection of a specific microhabitat can be
body surface to the sun permitting the used in addition to the gross body movements
cicada to maintain body temperature mechanisms: an animal can orient the side of
associated with shade-seeking behavior. This
in the range necessary for activity. Fur- the body to the sun, rotate the body around a
ther increases in body temperature was best illustrated by the description of
branch (Fig. 2, 3), or face the head into the
will stimulate movement into the movements in Diceroprocta apache (DAVIS)
shade. sun (HEATH & WILKIN 1970, HEATH et al.
associated with T a (HEATH & WlLKlN 1970).
1972, HEATH 1972, SANBORN et al. 1992, Diceroprocta apache is able to keep T^, below
SANBORN & MATE 2000). Changes in orienta- T a , in part, by selecting an appropriate
tion in the field have been correlated with microclimate.
458
© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Similarly, microhabitat choice is reported Another strategy for dealing with extreme
to be responsible for the difference in Tj, temperature is to suspend activity altogether.
recorded in two syntopic species (HASTINGS & Cicadas are torpid at low Tj, and low T a
TOOLSON 1991). Tibken chiricahua DAVIS is (HEATH 1967, HEATH & WILKIN 1970, HEATH
reported to exhibit a higher T^ because it is et al. 1971, HEATH et al. 1972, HEATH 1972,
active closer to the ground and in less shade. SANBORN et al. 1992, SANBORN 2000,
In contrast, T. duryi DAVIS uses more elevated SANBORN & MATE 2000). This torpidity can
perches in deeper shade and has an average T^ be a benefit to ectothermic animals in that
3.1°C lower than T. chiricahua. The differen- metabolic rate decreases as T^ decreases. The
ces are not due to biophysical factors as
T. duryi is the larger of the two species and
should, therefore, equilibrate at a higher T^.
These differences may be realted to distributi-
on of T. chiricahua which would require adap-
tation to warmer T a and the selection of ele-
vated T^s for activity since it lives in warmer
habitats (HASTINGS & TOOLSON 1991).
Selection of a proper microhabitat is also
important for one of the physiological respon-
ses to temperature to be effective. The decrea-
se in Tj, due to evaporative cooling disappears
when the animals are placed in high humidity
environments (TOOLSON 1985, TOOLSON
1987, HASTINGS 1989). Selection of an impro-
per microhabitat could lead to an ineffective
evaporative cooling response.
Related to the selection of a particular
microclimate is a vertical change in position
with changes in T a . This is especially well
exemplified by the vertical migrations of only behaviors exhibited at low T^ or T a are Figs. 3:
Okanagodes pracilis (SANBORN et al. 1992). A cicada that has moved to a negative
basking, in an effort to elevate Tjy and fee-
orientation. This specimen of Dicero-
When T a is less than© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
Physiological Responses All cicadas bask at low T^s. The endo-
thermic species augment solar heating with
Although behavioral thermoregulation metabolic heat production. At dusk or when
has several benefits, there are still limitations environmental conditions prevent the use of
placed on reproductive activity due to the solar radiation, the endothermic cicadas use
limits to changes in T^ through behavior. metabolic heat to become or to remain active.
Physiological mechanisms assist behavioral Thus, the endothermic species do not rely on
strategies in regulating T^ to a more precise metabolic heat alone to regulate T^. They will
level, thus permitting activity over a wider use radiant solar energy to elevate T^ for
range of T g . There are three main physiologi- activity when it is available (SANBORN et al.
cal responses used by cicadas to survive the 1995a, SANBORN 2000). Restriction in the use
thermal environment: endothermy, evaporati- of metabolic heat saves energy stores in the
ve cooling and thermal responses. cicadas. The energetic expense of behavioral
thermoregulation is the cost of transporting
Endogenous heat production used specifi-
the animal's mass from one location to ano-
cally for thermoregulation constitutes the
ther or the cost of maintaining a particular
definition of endothermy (ANONYMOUS
posture which represents a small fraction of
1987). Endogenous heat production with the
the energy cost to maintain T^ using metabo-
potential for thermoregulation was first shown
lic heat (HEATH 1970).
in the tropical cicada Fidicina mannifera
(FABRICIUS) (BARTHOLOMEW & BARNHARDT The relationship between field temperatu-
1984). When BARTHOLOMEW & BARNHARDT res and the T^ when voluntary endogenous
(1984) described endothermy in F. mannifera, heat production ceased in Proarna bergi
they stated that the functional significance of (DISTANT) and P. insignis presents further evi-
the endogenous heat was problematic. My col- dence that the cicadas are warming to a level
leagues and I have begun to investigate endo- necessary for activity. Captive animals gene-
thermy in cicadas in order to determine its rally cease warm-up behavior in the same T^
biological significance (SANBORN et al. 1995a, range of those animals active in the field. The
SANBORN et al. 1995b, SANBORN 1997,
data also illustrate that the animals possess the
SANBORN 2000, SANBORN in press).
metabolic machinery necessary to raise their
T|j to a biologically significant range
Diel T|j distribution of endothermic cica- (SANBORN et al. 1995a, SANBORN et al. 1995b,
das is similar to that in ectothermic species in SANBORN 2000). Similarly, the mean T^ of
that T^ is elevated above T g during daylight endothermically active Tibicen winnemanna
hours. Endothermic cicadas differ, however, in does not differ significantly from the mean T^
that the T^ of cicadas "active" at dusk or during the day (SANBORN 2000) and the mean
when solar energy is unavailable is approxima- peak T^ measured by BARTHOLOMEW & BARN-
tely the same as the diurnal distribution of HARDT (1984) during warm-up behavior in
T[jS. A distinction between "active" and Fidicina mannifera is similar to the upper ther-
"inactive" individuals must be made because moregulatory temperature we determined for
the Tjj of "inactive" animals approximates T g the species F. torresi BOULARD & MARTINELLI
(SANBORN et al. 1995a, SANBORN 2000) as is (which was split out from F. mannifera)
found in ectothermic species under these con- (SANBORN et al. 1995a).
ditions (HEATH 1967, SANBORN 2000). T^s of
Female endothermic cicadas raise Tj^ dur-
Guyalna bonaerensis (BERG), Quesada gigas ing the chorusing activity of the males. Fema-
(OLIVIER), Proarna insignis DISTANT, and le Guyalna bonaerensis with elevated T^s have
Tibicen winnemanna (DAVIS) measured in the been observed ovipositing in the rain
field show that these cicadas are endothermic. (SANBORN et al. 1995a). Female Tibicen win-
Recorded Tj^s were respectively as much as nemanna captured during the species' evening
13.0°C, 12.0°C, 7.4°C, and 12.6°C greater activity period also had elevated Tus
than T a when solar heating was unavailable to (SANBORN 2000). The females may be requi-
the species (SANBORN et al. 1995a, SANBORN red to elevate Tjj for species recognition.
et al. 1995b, SANBORN 2000). Female planthoppers (VRIJER 1984) and
460© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
crickets (WALKER 1957, WALKER 1963, MOORE 1962, HAYASHI 1982, POPOV et al.
DOHERTY 1985) have been shown to respond 1985, DUFFELS 1988). These species may well
only to the calls produced by males at the be endothermic since the ectothermic dusk
same Tu, as the female. Some evidence to sup- singer Cystosoma saundersii (JOSEPHSON &.
port a similar temperature dependent recogni- YOUNG 1979) does not exhibit "sing and fly"
tion system in Tibicen winnemanna is that Tu,s behavior (DOOLAN & MACNALLY 1981).
of the mating females observed were within Fidicina mannifera (BARTHOLOMEW &
0.5°C of the males with which they were BARNHARDT 1984) and Tibicen winnemanna
copulating (SANBORN 2000). (SANBORN 2000) are capable of endothermi-
The heat for thermoregulation is produced cally warming without flight. Non-flapping
by the thoracic musculature. The relatively warm-up in F. mannifera was accompanied by
large percentage of body mass (35% in cicadas barely perceptible low frequency (1-2 s )
[BARTHOLOMEW & BARNHARDT 1984, wing movements and telescoping of the abdo-
SANBORN et al. 1995b]), the high metabolic men at 15-36 cycles min . Non-flapping
scope, and the inefficiency of flight all make warm-up was not observed in T. winnemanna
the flight musculature an ideal tissue to gene- in the laboratory, but individuals were obser-
rate heat for thermoregulation. The extremely ved to progress from a buzzing call to produ-
high metabolic rate of insect flight muscles cing a full mating call without changing sin-
makes them well suited for heat production ging perches in the field. Since the mean Tu,
(BARTHOLOMEW & EPTING 1975). Guyana of a cicada producing the fast song is signifi-
bonaerensis, Quesada gigas, Fidicina torresi cantly greater than one producing a buzz
(SANBORN et al. 1995a) and Tibicen winne- (SANBORN 1997), the cicadas were capable of
manna (SANBORN 2000) have been observed raising Tu, without flight. Species of Coleop-
to use the heat produced in flight to raise Tu, tera, Hymenoptera, Diptera, and Lepidoptera
endothermically. also have been shown to raise Tu, without
wing movements (MAY 1976, BARTHOLOMEW
Insect flight is a mechanically inefficient
& HEINRICH 1978, HEINRICH 1981, CHAPPELL
process. Flight is normally assumed to have an
& MORGAN 1987).
efficiency of 20% (WEIS-FOGH 1972, WEIS-
FOGH 1976, CASEY 1981a), but studies of Proama bergi and P. insignis generate meta-
several insect species demonstrate that flight bolic heat with shiver-like movements of the
efficiency ranges from 3.3-27.5% (WEIS-FOGH wings (SANBORN et al. 1995b) (Fig. 5). The
1972, WEIS-FOGH 1976, CASEY 1981b, amplitude of the wing vibrations is found to
ELLINGTON 1984, ELLINGTON 1985, CASEY et change with increasing Tu, in some moth spe-
al. 1985). The efficiency of insect flight is thus cies (DORSETT 1962, Kammer 1981) but this
much lower than the assumed value of 20% did not occur in these cicadas. The mecha-
and as a class insects have a mean flight effi- nism producing the wing vibrations in the
ciency between 10-15%. This means that a cicadas is probably similar to the near syn-
significant amount of the energy used in flight chronous activation of wing elevator and
(85-90%) is released as heat. depressor muscles described by KAMMER
Behavior similar to that observed in the (1970) in the butterfly Danaus plexippus (L.)
endothermic cicadas has been described in but this has yet to be shown with electrical
many dusk singing cicadas and endothermy recordings in the cicadas.
may be a wide spread phenomenon in the There have been diverse functions descri-
superfamily Cicadoidea. It has been reported bed for endogenous heat production of insects
that Tibicen auletes (GERMAR) and Tibicen reso- (HEINRICH 1993). It has been estimated that
nans (WALKER) are active for a greater period an endothermic cicada would use more than
of time than environmental conditions should five times as much energy during activity as an
permit (ALEXANDER 1960). "Sing and fly" ectothermic animal (SANBORN et al. 1995a).
behavior has been described in these and The use of metabolic heat to raise Tu, for
many other species active at dusk (DAVIS activity will decrease the life span of animals
1894a, DAVIS 1894b, MATSUMURA 1898, that do not obtain large amounts of energy
ANNANDALE 1900, KERSHAW 1903, DAVIS from their food. Since cicadas feed on xylem
1922, DISTANT 1906, ALEXANDER 1956, fluid (CHEUNG & MARSHALL 1973a, WHITE &
461© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
STREHL 1978), the nutrients available for face, and the partial pressure gradient of water
energy conversion are limited. This raises the between the evaporating surface and the sur-
question as to why an animal would expend rounding air. The use of evaporative cooling
energy stores to become active when it could by insects is a relatively rare occurrence due to
use solar radiation during the day and save their relatively small body si:e. Their high sur-
metabolic energy. face to volume ratio, and the limitations of
Endothermic behavior in cicadas may ser- sufficient water reserves can quickly lead to
osmoregulatory problems for insects that
ve to increase reproductive fitness in several
attempt to use evaporative cooling. PRANGE
ways including: uncoupling of reproductive
(1996) has provided a fairly recent
review of evaporative cooling in
insects as a group.
The use of evaporative cooling
in cicadas was first described by
KASER and HASTINGS (1981) in
Tibicen duryi. It has since been
described in Cacama valvata
(TOOLSON 1993), Diceroprocta apa-
che (TOOLSON 1985, TOOLSON
1987, TOOLSON & HADLEY 1987,
HADLEY et al. 1989, HADLEY et al.
1991), Okanagodes gracilis (SAN-
RORN et al. 1992), T. chiricahua
(HASTINGS & TOOLSON 1991),
T. dealbatus (DAVIS) (TOOLSON
1984, STANLEY-SAMUELSON et al.
1990, TOOLSON et al. 1994), and
further descriptions for T. duryi
(TOOLSON 1984, HASTINGS 1989,
HASTINGS & TOOLSON 1991). All
species inhabit environments in
which T]., could elevate to lethal
levels. Significant evaporative coo-
Figs. 4: behavior from possible physiological con- ling has only been described for species that
Okanagodes gracilis DAVIS thermoregu- inhabit hot, dry environments even though
straints of the environment, avoidance of pos-
lating during the warmest portion of
the day. Ambient temeprature was sible thermoregulatory problems of midday by they have different phylogenetic histories
48°C when the image was taken. The restricting activity to the cooler portions of (HEATH 1978). An evaporative cooling
picture illustrates the combined use of the day, permitting the use of habitats unavai- mechanism was described in a species from a
behavioral and physiological strate-
lable to strictly ectothermic animals, helping warm, humid environment but it was found
gies to thermoregulate. The animal is
exhibiting a negative orientation species to avoid predators, optimizing broad- the species had limited evaporative capabili-
which shades the body with the cast coverage and sound transmission, or to ties (TOOLSON & TOOLSON 1991). The ability
branch and exposes a white ventral to decrease Tj, below T a disappears when ani-
surface to the sun to reflect more permit avoidance of acoustic interference
solar radiation and decrease heat load. through temporal separation of singing (SAN- mals are placed in a saturated environment
The animal has also abandoned the RORN et al. 1995a, SANBORN et al. 1995b, (TCXUSON 1985, TOOLSON 1987, HASTINGS
normal host plant for a higher perch 1989). It appears a significant selection pres-
SANBORN 1997, SANBORN 2000).
which places the animal in a cooler
sure in the evolution of evaporative cooling is
microclimate. The animal is feeding A second physiological mechanism to
to get the water necessary for evapo- the environment inhabited by the species.
regulate T^ is evaporative cooling. Evaporati-
rative cooling which depresses the
body temperature below ambient. on of water represents a potentially significant Cicadas will attempt to use behavioral
These activities combine with elevated mechanism for the dissipation of excess heat mechanisms to decrease T^ before evaporati-
thermal tolerances permit reproduc- on of water begins (HADLEY et al. 1991,
from any organism. The amount of energy lost
tive activity at a time when other ani-
is dependent upon the coefficient of evapora- SANBORN et al. 1992). However, Diceroprocta
mals are seeking shelter from the
heat. tion, the surface area of the evaporating sur- apache can maintain Tj, 5°C below T a
462© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
(TOOLSON 1987) and Okanagodes gracilis can system (TOOLSON &. HAPLEY 1987, HAPLEY et
maintain a 6.7°C gradient (SANBORN et al. al. 1989, HADLEY et al. 1991). Death or the
1992) once evaporation begins. Water Kiss injection of NaCN causes a rapid decrease in
rates increase as much as 600% during the transcuticular water flux in Diceroprocta apa-
evaporative response (TOOLSON & HAPLEY che. The system probably operates by the cou-
1987) with the amount of water loss increa- pling of an actively transported ion across the
sing with increasing T a (HAPLEY et al. 1991). cell membrane of the dermal glands or dermal
The onset of evaporative cooling occurs at gland ducts which then causes water to move
a thermoreglatory point that is similar to the by osmosis. The water can then evaporate
maximum voluntary tole-
rance temperature. Evapo-
ration begins at a T^ ot
39.2°C in Dkeroprocta apa-
che (HAPLEY et al. 1989)
which is the same as the
maximum voluntary tole-
rance temperature determi-
ned by HEATH & WILKIN
(1970). Evaporative coo-
ling in Okanagodes gracilis
occurrs at a T^ slightly abo-
ve the maximum voluntary'
tolerance temperature but
this may have been an arti-
fact of the experimental
procedure (SANBORN et al.
1992). Evaporative cooling
begins at lower T^s when
animals are well hydrated
and have access to food
(TOOLSON & HALUEY
1987, HAPLEY et al. 1991)
and T^s elevate as water
reserves decrease (HASTINGS 1989). Figs. 5: Endogenous heat production in Proarna insignis DISTANT. The images on the left
depict an animal at rest with the wings held over the body. The images on the right
Water is evaporated through pores in the were taken while the animal was generating heat to elevate body temperature. The
cuticle (Fig. 6) (TOOLSON & HAPLEY 1987, wings flatten in the dorso-ventral plane and are vibrated in a shiver-like manner
HADLEY et al. 1991, SANBORN et al. 1992). during heat production. The shivering stops when the animals have elevated their
body temperature to the range necessary for activity.
These pores are located on the thorax and
abdomen in both Dkeroprocta apache and Oka-
nagodes gracilis. The pore structure and distri- Figs. 6: The dorsal
mesothorax of
bution is also similar in the two species. Magi- Okanagodes gracilis
cicada tredecim (WALSH & RlLEY), a species DAVIS. The pores are
from a higher humidity, lower T a environ- used to evaporate
water to cool the ani-
ment, has a limited number of pores and a mal in extreme heat.
limited ability to cool evaporatively (TOOL- The individual pores
SON & TOOLSON 1991). The main site of eva- are about 7 mm in
diameter.
porative water loss is the abdomen in D. apa-
che (HAPLEY et al. 1989) which would explain
the lower abdominal temperatures also obser-
ved in Tibken duryi (HASTINGS 1989).
The evaporative cooling mechanism
appears to be mediated by an active transport
463© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
once it is outside the cuticle. There is a cycli- maintain Tj, less than T a (KASER & HASTINGS
cal rate of water loss at higher Ta> which is 1981, HASTINGS 1989) and appears to alter
similar to evaporative cooling mechanisms the T(j at which the evaporative response is
described for mammals (HADLEY et al. 1989). initiated (HADLEY et al. 1991, SANBORN et al.
Prostaglandins have been suggested to 1992, TOOLSON et al. 1994). Okanagodes graci-
regulate the control of transcuticular water lis continually feeds during the day even while
flux in cicadas (STANLEY-SAMUELSON et al. performing reproductive activities such as sin-
1990, TOOLSON et al. 1994). Blocking the syn- ging and ovipositing (SANBORN et al. 1992) to
thesis of prostaglandins prevents the initiation maintain water balance (Fig. 4). Tibicen duryi
of the sweating response in Tibicen dealbatus. also feeds during the day (KASER & HASTINGS
The results of these experiments suggest a 1981, HASTINGS 1989) but this species sus-
complex interaction of chemicals in the pends other activities while it is feeding
20 carbon, polyunsaturated fatty acids and (HASTINGS 1989).
arachidonic acid synthesis pathway that regu- A final physiological mechanism cicadas
late the sweating response by regulating the employ is adaptation to particular thermal
set point for initiation (TOOLSON et al. 1994). environments. An analysis of the thermal res-
It appears cicadas that use evaporative ponses for a particular insect can give insights
cooling are adapted to survive the loss of a lar- into how the animal is adapted thermally to
ge percentage of their total body water during its environment. Early work (HEATH et al.
the evaporative cooling response. Mean water 1971, HEATH et al. 1972) suggested the ther-
loss rates of 141% h in Diceroprocta apache mal responses of a particular species were rela-
(HADLEY et al. 1991), 10.67% h ' 1 in Tibicen ted to the habitat type and the elevation of
duryi, 11.01% h ' l in T. chiricahua (HASTINGS the habitat. This relationship has proven to be
& TOOLSON 1991), and 9.73% h' 1 in Okana- an over simplification as the number of species
godes gracilis (SANBORN et al. 1992) with a investigated has increased and endothermy
maximum rate of 30-35% h ' ' in Diceroprocta was discovered. Our further analyses have sug-
apache (TOOLSON 1987) have been reported. gested that the thermal responses of cicadas
are related to the activity patterns, thermore-
Insects in general simply do not have suf-
gulatory strategies, and habitat of a particular
ficient body water reserves to employ evapora-
species.
tion as a main mechanism of thermoregulati-
on. However, cicadas are an exception in that Three measurements are required to deter-
they have access to a water source that other mine the thermal responses of a species
organisms in their environment do not. Cica- (HEATH 1967, HEATH & WILKIN 1970, HEATH
das feed on xylem fluid (CHEUNG & MARS- et al. 1971, HEATH et al. 1972, HEATH 1972,
HALL 1973a) which is mostly water. In fact, SANBORN et al. 1992, SANBORN et al. 1995a,
the digestive system of cicadas is modified to SANBORN et al. 1995b, SANBORN & PHILLIPS
eliminate the excess water load they get with 1996, SANBORN 1997, SANBORN 2000, SAN-
their food (CHEUNG & MARSHALL 1973b, BORN & MATE 2000, SANBORN &. PHILLIPS
MARSHALL & CHEUNG 1973, MARSHALL & 2001). The minimum flight temperature
CHEUNG 1974, MARSHALL & CHEUNG 1975, (MFT) represents the lowest T^ at which an
MARSHALL 1983). Thus, the use of a specific animal is considered fully coordinated. The
food source in a dry environment has permit- maximum voluntary tolerance (MVT) or sha-
ted certain cicadas to use evaporative cooling de-seeking temperature is a measure of an
as a means to regulate T^. This has allowed upper thermoregulatory set point (HEATH
the cicadas to remain active in a thermally 1970). The heat torpor temperature (HTT) is
stressed environment at times when other ani- the upper T^ limit of coordinated activity and
mals, especially their predators, have retreated can represent an ecologically lethal T^ since
to thermal shelters due to the excessive heat the animal is unable to prevent further increa-
of the environment. ses in T)j. The T^ range of full activity is the
Water uptake in evaporatively cooling difference between the MFT and HTT.
cicadas has become an integral daily activity. Table 1 is a summary of the reported cicada
Access to water is required for the cicadas to temperature responses.
464© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
The MFTs are the most difficult thermal MVT when it is endothermically active. On
response to analyze. There is no clear correla- the other extreme are Diceroprocta apache
tion between habitat type and MFT. For (HEATH & WILKIN 1970) and Okanagodes gra-
example, the desert inhabiting Diceroprocta cilis (SANBORN et al. 1992). These species
cinctifera (UHLER) has the second lowest repor- inhabit the Sonoran Desert where T a can
ted MFT (SANBORN &. PHILLIPS 1996). Simi- approach or exceed 50°C. The animals must
larly, OUanagana hesperia (UHLER) lives at alti- have elevated thermal tolerances to be active
tude (HEATH 1972) but has a higher MFT for a significant portion of the day in their
than desert dwelling species (HEATH & WlL- environment. In addition, the elevated MVT
Species M i n i m u m Flight Maximum Heat Torpor Range of Table 1:
Temperature Voluntary Full Activity M e a n temperature responses (°C)
Tolerance reported in t h e literature for cicadas.
Maqicicada cass/n/71 20.9 31.8 43.0 22.1 Endothermic species are marked w i t h
Okanaqana hesperia2 22.9 36.3 43.5 20.6 an asterisk(*).
Diceroprocta apache3 21.9 39.2 45.6 23.7
Okanaqodes qracilis* 22.7 41.2 48.7 26.0
Cacama valvata-'
CamD Verde 23.7 37.3 44.6 20.9
Aqua Fria 24.0 34.9 44.3 20.3
Diceroprocta cinctifera6 17.6 34.0 45.8 28.2
Diceroprocta cinctifera
var. limpia6 16.8 34.7 46.0 29.2
Diceroprocta cinctifera
var. viridicosta6 16.7 35.3 46.0 29.3
Diceroprocta aurantiaca7 17.7 36.3 46.0 28.3
Diceroprocta delicata7
Northern population 19.8 33.9 46.8 27.0
Southern population 21.4 35.3 46.5 25.1
Diceroprocta olvmpusa8 20.4 37.0 46.7 26.3
Tibicen chloromerus9 19.2 34.4 45.3 26.1
Tibicen winnemanna*9 16.4 32.7 45.6 29.2
1
Guyalna bonaerensis*™ 16.7 34.6 44.6 27.9 Heath 1967, 2 Heath 1972, 3 Heath and
Wilkin 1970, 4 Sanborn e t a l . 1992, 5 Heath
Fidicina torresi*^0 19.8 32.0 42.0 22.2
et al. 1972, 6 Sanborn and Phillips 1996,
Quesada qj/qas* 10 19.1 33.8 44.9 25.8 7
Sanborn and Phillips 2001, 8 Sanborn and
Proarna bero/* 1 1 20.7 37.6 46.3 25.6 Mate 2000, 9 Sanborn 2000, 10 Sanborn et
Proarna /ns/gn;'s*11 19.3 36.4 44.0 24.7 al. 1995a, 11 Sanborn e t a l . 1995b.
KIN 1970, SANBORN et al. 1992). It has been also delays the start of the evaporative cooling
suggested that the MFT probably relates more response in these species which could be
to the physical design of the cicada flight important in maintaining water balance.
motor system than to the origin of the cicada HTT is strictly related to the habitat of a
(HEATH et al. 1972). My laboratory has now
species. The HTT appears to have evolved to
found evidence to support this hypothesis. We
reflect the maximum thermal load a species
have been able to find several morphological
might face in a particular environment. It does
variables that influence lift that correlate with
not appear to be related to the thermoregula-
MFT. However, it appears that the environ-
tory strategy or behavior of a species (HEATH
ment still plays a role in determining what the
1967, HEATH & WILKIN 1970, HEATH et al.
MFT will be for a particular species. .
1971, HEATH et al. 1972, HEATH 1972, SAN-
The MVT is easier to relate to the habitat
BORN et al. 1992, SANBORN et al. 1995a, SAN-
than the MFT. There is an increase in MVT as
BORN et al. 1995b, SANBORN & PHILLIPS 1996,
habitats become warmer. Fidicina wrresi is
SANBORN 2000, SANBORN & MATE 2000, SAN-
active on the trunks of primary forest trees in
BORN & PHILLIPS 2001).
the tropics and has the lowest reported MVT
(SANBORN et al. 1995a). The limited exposure There are some generalizations that can be
to solar radiation due to the canopy means made in the analysis of the thermal responses
F. torresi will save energy with a depressed of endothermic species. The endothermic
465© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
cicadas generally have a lower MFT and a CADE 1975, SOPER et al. 1976, MANGOLD
greater T^ range of full activity when compa- 1978, BELL 1979, BUCHLER & CHILDS 1981,
red to ectothermic species. The low MFT of SAKALUK & BELWOOD 1984, FOWLER &
the endothermic cicadas may be necessary KOCHALKA 1985, TUTTLE et al. 1985, BEL-
because of their endothermic activity'. A low WOOD & MORRIS 1987). An increase in cica-
MFT permits the cicada to utilize heat produ- da activity with a decrease in bird activity was
ced in flight to raise Tj,. If the MFT were ele- also observed in tropical Brazil (Jones 1884).
vated, low T a could possibly inhibit activity The study of cicada thermal biology has
(SANBORN et al. 1995a, SANBORN et al. 1995b,
led to some interesting insights into tempera-
SANBORN 2000).
ture effects and thermoregulation in insects.
The MVTs of the endothermic species are However, there is always more to learn about
variable but generally are lower than those how temperature influences activity in ani-
found in ectothermic species even if the habi- mals. We have even applied thermal biology
tats are similar (SANBORN et al. 1995a, SAN- to systematics in helping to separate Dicero-
BORN et al. 1995b, SANBORN 2000). The MVT procta aurantiaca DAVIS as a distinct species
values obtained probably represent a balanced (SANBORN & PHILLIPS 2001). Continued rese-
response to the T a s encountered during their arch using integrated methods should attempt
activity period and the maximum T a encoun- to answer more questions about the thermal
tered during the day. If the upper ther- biology of this interesting group of insects.
moregulatory point were elevated too high,
the cicada might be unable to produce suffi-
cient heat to become active at dusk. Similarly
Zusammenfassung
high T a s could prohibit activity if the MVT
was depressed any further. Mechanismen und Strategien der Tempe-
The HTTs of endothermic species show raturregelung bei Singzikaden werden im
the same dependency to the environment as Überblick dargestellt, Verhaltensweisen und
the HTT of ectothermic species. This is best physiologische Vorgänge zur Regulation der
illustrated in the HTT of the endothermic Körpertemperatur diskutiert. Verhaltenswei-
Tibicen uiinnemanna and the syntopic ecto- sen umfassen Lageveränderung des Körpers zur
thermic T. chloromerus (WALKER). The HTT Sonne, sonnenbaden, Schatten aufsuchen,
of both species are approximately equal even Auswahl günstiger mikroklimatischer Verhält-
though the two species have vastly different nisse, Vertikalwanderung, Verwendung der
behavioral strategies and thermoregulatory Flügel als "Sonnenschirm" und die Einstellung
points (SANBORN 2000). der Aktivität. Die Auswirkungen der Tempe-
Elevated thermal tolerances may also be ratur auf die Biologie der Singzikaden und
an adaptation to predator avoidance. Dicero- deren Reaktionen auf bestimmte Temperatur-
procta apache (HEATH &. WlLKiN 1970) and the verhältnisse werden diskutiert. Physiologi-
syntopic species Okanagodes gracilis (SANBORN sche Reaktionen umfassen Temperaturanpas-
et al. 1992) have the highest reported thermal sung, Endothermie und Kühlung durch Eva-
tolerances in cicadas. Their elevated thermal poration.
tolerances permit the species to be active in a
habitat when their potential predators must
References
retreat to a thermal shelter due to the extreme
heat. This is an advantage to the cicadas since AIDLEY DJ. & D.C.S. WHITE (1969): Mechanical pro-
perties of glycerinated fibers from the tymbal
the males produce conspicuous acoustic sig- muscles of a Brazilian cicada. — Jour. Physiol.
nals to attract females. By calling when preda- 205: 179.
tors are not active, the cicadas should decrea- ALEXANDER R.D. (1956): A comparative study of sound
se their predation pressure since several types production in insects with special reference to
the singing Orthoptera and Cicadidae of the
of predators have been shown to orient to the
eastern United States. — Ph. D. dissertation. The
acoustic signals of insects (WALKER 1964, Ohio State University, Columbus, Ohio, 529 pp.
466© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
ALEXANDER R.D. (1960): Sound communication in CHEUNG W.W.K. S A.T. MARSHALL (1973b): Studies on
Orthoptera and Cicadidae. — In: LANYON W.E. & water and ion transport in homopteran inserts:
W.N. TAVOLGA, Animal sounds and communica- Ultrastrurture and cytochemistry of the cicado-
tion. — Amer. Inst. Biol. Sei. Symp. Ser. Publ. 7: id and cercopoid midgut. — Tissue and Cell 5:
pp. 38-92. 651.
ANNANDALE N. (1900): Observations on the habits and DAVIS W.T. (1894): Staten Island harvest flies. —
natural surroundings of insects made during the Amer. Nat. 28: 363.
"Skeat Expedition" to the Malay Peninsula,
DAVIS W.T. (1894): Staten Island harvest flies. — Proc.
1899-1900. V. Sounds produced by insects. —
Nat. Sei. Assoc. Staten Island 4: 9.
Proc. Zool. Soc. London 1900: 859.
DAVIS W.T. (1922): An annotated list of the cicadas of
ANONYMOUS (1987): Glossary of terms for thermal Virginia with description of a new species. —
physiology, second edition. Revised by the com- Jour. N.Y. Ent. Soc. 30: 36.
mission for thermal phsiology of the internatio-
nal union of physiological sciences. — Pflüg. DISTANT W.L. (1906): Order Rhynchota. Suborder
Archiv. Eur. Jour. Physiol. 410: 567. Homoptera. Fam. Cicadidae. — Insecta Trans-
vaaliensia 7: 167.
AZUMA S. (1976): Biological studies of the sugar cane
DOHERTY J.A. (1985): Temperature coupling and 'tra-
cicada, Mogannia minuta MATSUMURA, w i t h
de-off' phenomena in the acoustic communica-
special reference to its occurence in Okinawa. —
tion system of the cricket, Gryllus bimaculatus
Bull. Coll. Agric. Ryukyus Univ. 23: 125.
DE GEER (Gryllidae). — Jour. Exp. Biol. 114: 17.
BARTHOLOMEW G.A. & M.C. BARNHARDT (1984): Tracheal
DOOLAN J.M. & R.C. MACNALLY (1981): Spatial dyna-
gases, respiratory gas exchange, body tempera-
mics and breeding ecology in the cicada, Cysfo-
ture and flight in some tropical cicadas. — Jour.
soma saundersii: The interaction between distri-
Exp. Biol. 111: 131.
butions of resources and interspecific behavior.
BARTHOLOMEW G.A. & R.G. EPTING (1975): Allometry of — Jour. Anim. Ecol. 50: 925.
post-flight cooling rates in moths: A comparison
DORSETT D.A. (1962): Preparation for flight by hawk-
with vertebrate homeotherms. — Jour. Exp.
moths. — Jour. Exp. Biol. 39: 579.
Biol. 63: 603.
DUFFELS J.P. (1988): The cicadas of the Fiji, Samoa, and
BARTHOLOMEW G.A. & B. HEINRICH (1978): Endothermy
Tonga Islands, their taxonomy and biogeogra-
in African dung beetles during flight, ball
phy (Homoptera, Cicadoidea) with a chapter on
making, and ball rolling. — Jour. Exp. Biol. 73:
the geological history of the area by A. Ewart.
65.
— Entomonograph 10: 1.
BELL P.D. (1979): Acoustic attraction of herons by
DURIN B. (1981): Inserts Etc.: An anthology of Arthro-
crickets. — Jour. N.Y. Ent. Soc. 87: 126. pods featuring a bounty of beetles. — Hudson
BELWOOD J.J. & G.K. MORRIS (1987): Bat predation and Hills Press, New York, 108 pp.
its influence on calling behavior in neotropical ELLINGTON C.P. (1984): The aerodynamics of hovering
katydids. — Science 238: 64. insert flight. VI. Lift and power requirements. —
BUCHLER E.R. S S.B. CHILDS (1981): Orientation t o Phil. Trans. Roy. Soc. Lond. Ser. B. 305: 145.
distant sounds by big brown bats (Eptescus ELLINGTON C.P. (1985): Power and efficiency of insert
fuscus). — Anim. Behav. 29: 428. flight muscle. —Jour. Exp. Biol. 115: 293.
CADE W. (1975): Acoustically orienting parasitoids: fly FLEMING C.A. (1975): Adaptive radiation in New Zea-
phonotaxis to cricket song. — Science 190: land cicadas. — Proc. Amer. Phil. Soc. 119: 298.
1312. FOWLER H.G. & J.N. KOCHALKA (1985): New record of
CASEY T.M. (1981a): Energetics and thermoregulation Euphasiopteryx depleta (Diptera: Tachinidae)
of Malacosoma americanum (Lepidoptera: from Paraguay: Attraction t o broadcast calls of
Lasiocampidae) during hovering flight. — Scapteriscus acletus (Orthoptera: Gryllotalpi-
Physiol. Zool. 54: 362. dae). — Florida Entomol. 68: 225.
HADLEY N.F., QUINLAN M.C. & M.L. KENNEDY (1991): Eva-
CASEY T.M. (1981b): A comparison of mechanical and
porative cooling in the desert cicada: thermal
energetics estimates of flight cost for hovering
efficiency and water/metabolic costs. — Jour.
sphinx moths. — Jour. Exp. Biol. 91: 117.
Exp. Biol. 159: 269.
CASEY T.M., MAY M.L. & K.R. MORGAN (1985): Flight
HADLEY, N.F., TOOLSON E.C. & M.C. QUINLAN (1989):
energetics of Euglossine bees in relation to mor-
Regional differences in cuticular permeability in
phology and wing stroke frequency. — Jour.
the desert cicada Diceroprocta apache: Implica-
Exp. Biol. 116:271.
tions for evaporative cooling. — Jour. Exp. Biol.
CHAPPELL M.A. & K.R. MORGAN (1987): Temperature 141: 219.
regulation, endothermy, resting metabolism, HARTZELL A. (1954): Periodical cicada. — Contrib.
and flight energetics of Tachnid flies {Nowickia Boyce Thompson Inst. 17: 375.
sp.). — Physiol. Zool. 60: 550.
HASTINGS J.M. (1989): Thermoregulation in the
CHEUNG W.W.K. S A.T. MARSHALL (1973a): Water and dog-day cicada Tibicen duryi (Homoptera:
ion regulation in cicadas in relation t o xylem Cicadidae). — Trans. Kentucky Acad. Sei. 50:
feeding. — Jour. Insect Physiol. 19: 1801. 145.
467© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
HASTINGS J.M. & E.C. TOOLSON (1991): Thermoregulati- KAMMER A.E. (1970): Thoracic temperature, shivering,
on and activity patterns of 2 syntopic cicadas, and flight in the monarch butterfly, Danaus
Tibicen chiricahua and T. duryi (Homoptera, plexippus (L). — Zeit. f. Vergl. Physiol. 68: 334.
Cicadidae), in central New Mexico. — Oecologia
KAMMER A.E. (1981): Physiological mechanisms of
85: 513.
thermoregulation. — In: HEINRICH B., Insect
HAYASHI M. (1982): Notes on the distribution of Cica- thermoregulation, John Wiley & Sons, New
didae (Insecta, Homoptera) in the Fuji-Hakone- York, pp. 115-158.
Izu area. — Mem. Nat. Sei. Mus., Tokyo 15: 187.
KASER S.A. & J. HASTINGS (1981): Thermal physiology of
HEATH J.E. (1967): Temperature responses of the the cicada Tibicen duryi. — Amer. Zool. 2 1 :
periodical "17-year" cicada, Magicicada Cassini 1016.
(Homoptera, Cicadidae). — Amer. Midi. Nat. 77:
KERSHAW J.C. (1903): A naturalist's notes from China.
64.
— Field Nat. Quart. 2: 233.
HEATH J.E. (1968): Thermal synchronization of emer-
MANGOLD J.R. (1978): Attraction of Euphasiopteryx
gence in periodical "17-year" cicadas (Homop-
ochracea, Corethrella sp. and gryllids to broad-
tera, Cicadidae, Magicicada). — Amer. Midi.
cast songs of the southern mole cricket. —
Nat. 80: 440.
Florida Entomol. 6 1 : 57.
HEATH J.E. (1970): Behavioral regulation of body tem-
perature in poikilotherms. — Physiologist 13: MARSHALL A.T. (1983): X-ray microanalysis of the filter
399. chamber of the cicada, Cyclochila australasiae
DON.: A water-shunting epithelial complex. —
HEATH J.E., HANAGAN J.L, WILKIN P.J. & M.S. HEATH
Cell and Tissue Res. 231: 215.
(1971): Adaptation of the thermal responses of
insects. — Amer. Zool. 11: 147. MARSHALL A.T. & W.W.K. CHEUNG (1973): Studies on
water and ion transport in homopteran insects:
HEATH J.E. & P.J. WILKIN (1970): Temperature respon-
infrastructure and cytochemistry of the cicado-
ses of the desert cicada, Diceroprocta apache
id and cercopoid malpighian tubules and filter
(Homoptera, Cicadidae). — Physiol. Zool. 43: chamber. — Tissue & Cell 6: 153.
145.
MARSHALL A.T. & W.W.K. CHEUNG (1974): Studies on
HEATH J.E., WILKIN P.J. & M.S. HEATH (1972): Tempera-
water and ion transport in homopteran insects:
ture responses of the cactus dodger, Cacama
Ultrastructure and cytochemistry of the cicado-
valvata (Homoptera, Cicadidae). — Physiol.
id and cercopoid hindgut. — Tissue 8 Cell 5:
Zool. 45: 238.
671.
HEATH M.S. (1972): Temperature requirements of the
MARSHALL A.T. & W.W.K. CHEUNG (1975): Ionic balance
cicada Okanagana striatipes beameri: A study
of Homoptera in relation t o feeding site and
from Flagstaff, Arizona. — Plateau 45: 31.
plant sap composition. — Entomol. Exp. Appl.
HEATH M.S. (1978): Genera of American cicadas north 18: 117.
of Mexico. — Ph. D. dissertation, University of MATSUMURA S. (1898): A summary of Japanese Cicadi-
Florida, Gainesville, 231 pp. dae with description of a new species. — Ann.
Zool. Jap. 2: 1.
HEINRICH B. (1981): Temperature regulation during
locomotion in insects. — In: HERRIED C.F. & C.R. MAY M.L. (1976): Warming rates as a function of
FOURTNER, Locomotion and energetics in Arthro- body size in periodic endotherms. — Jour.
pods, Plenum Press, New York, pp. 391-417. Comp. Physiol. 111B: 55.
HEINRICH B. (1993): The hot-blooded insects. — MOORE T.E. (1962): Acoustical behavior of the cicada
Harvard University Press, Cambridge, 601 pp. Fidicina pronoe (WALKER) (Homoptera: Cicadi-
dae). — Ohio Jour. Sei. 62: 113.
HUDSON G.V. (1890): On the New Zealand Cicadidae.
— Trans. Proc. New Zealand Inst. 23: 49. NAGAMINE M. & R. TERUYA (1976): Life history of
Mogannia iwasakii MATSUMURA. — Bull. Okinawa
JOERMANN G. S H. SCHNEIDER (1987): The songs of four Agric. Exp. Sta. 2: 15.
species of cicada in Yugoslavia (Homoptera:
OHGUSHI R. (1954): Preliminary study on a cicada com-
Cicadidae). — Zool. Anz. 219: 283.
munity at Mt. Nakatsumine. — KontyCl 21: 10.
JONES E.D. (1884): In the tropics. — Naturalist (2)9:
POPOV A.V., ARONOV I.B. & M.V. SERGEEVA (1985):
125.
Calling songs and hearing in cicadas from Sovi-
JOSEPHSON R.K. (1981): Temperature and the mecha- et Central Asia. — Jour. Evol. Biochem. Physiol.
nical performance of insect muscle. — In: 21: 288.
Heinrich B., Insect thermoregulation, John PRANGE H.D. (1996): Evaporative cooling in insects. —
Wiley & Sons, New York, pp. 19-44. Jour. Insect Physiol. 42: 493.
JOSEPHSON R.K. & D. YOUNG (1979): Body temperature RAMSAY G.W. (1959): Notes on the ecology of some
and singing in the bladder cicada, Cystosoma cicadas in the Wellington District. — New Zea-
saundersii. — Jour. Exp. Biol. 80: 69. land Entomol. 2: 29.
JOSEPHSON R.K. & D. YOUNG (1985): A synchronous ins- SAKALUK S.K. & J.J. BELWOOD (1984): Gecko phonotaxis
ect muscle with an operating frequency greater to cricket song: a case of satellite predation. —
than 500 Hz. — Jour. Exp. Biol. 118: 185. Anim. Behav. 32: 659.
468© Biologiezentrum Linz/Austria; download unter www.biologiezentrum.at
SANBORM A.F. (1997): Body temperature and the acou- TALHOUK A.S. (1959): The grapevine cicada
stic behavior of the cicada Tibicen winnemanna Chloropsalta viridissima (WALKER) (Homoptera-
(Homoptera: Cicadidae). — Jour. Insect Behav. Cicadidae). — Proc. Int. Cong. Crop Protect.,
10: 257. Hamburg 1: 799.
SANBORN A.F. (2000): Comparative thermoregulation TOOLSON E.C. (1984): Interindividual variation in epi-
of sympatric endothermic and ectothermic cica- cuticular hydrocarbon composition and water
das (Homoptera: Cicadidae: Tibicen winneman- loss rates of the cicada, Tibicen dealbatus
na and Tibicen chloromerus). — Jour. Comp. (Homoptera: Cicadidae). — Physiol. Zool. 57:
Physiol. 186A: 551. 550.
SANBORN A.F. (2001): Timbal muscle physiology in the TOOLSON E.C. (1985): Evaporative cooling in the
endothermic cicada Tibicen winnemanna desert cicada, Diceroprocta apache. — Amer.
(Homoptera: Cicadidae). — Comp. Biochem. Zool. 25: 144A.
Physiol. : in press.
TOOLSON E.C. (1987): Water profligacy as an adap-
SANBORN A.F., HEATH J.E. & M.S. HEATH (1992): Ther- tation t o hot deserts. Water loss rates and eva-
moregulation and evaporative cooling in the porative cooling in the Sonoran Desert cicada,
cicada Okanagodes gracilis (Homoptera: Cicadi- Diceroprocta apache (Homoptera: Cicadidae).
dae). — Comp. Biochem. Physiol. 102A: 751. — Physiol. Zool. 60: 379.
SANBORN A.F., HEATH J.E., HEATH M.S. & F.G. NORIEGA
TOOLSON E.C. (1993): In the Sonoran Desert cicadas
(1995b): Thermoregulation by endogenous heat court, mate, and waste water. — Nat. Hist. 102:
production in two South American grass dwel- 37.
ling cicadas (Homoptera: Cicadidae: Proarna). —
Florida Entomol. 78: 319. Toolson E.C. (1998): Comparative thermal physiolo-
gical ecology of syntopic populations of Cacama
SANBORN A.F., HEATH M.S., HEATH J.E. & F.G. NORIEGA
valvata and Tibicen bifidus (Homoptera: Cicadi-
(1995a): Diurnal activity, temperature responses
dae): Modeling fitness consequences of tempe-
and endothermy in three South American cica-
rature variation. — Amer. Zool. 38: 568.
das (Homoptera: Cicadidae: Dorisiana bonae-
rensis, Quesada gigas, and Fidicina mannifera). TOOLSON E.C, ASHBY P.D., HOWARD R.W. & D.W. STANLEY-
— Jour. Thermal Biol. 20: 451. SAMUELSON (1994): Eicosanoids mediate control
of thermoregulatory sweating in the cicada,
SANBORN A.F. S S. MATE (2000): Thermoregulation and
Tibicen dealbatus (Insecta: Homoptera). — Jour.
the effect of body temperature on call temporal
Comp. Physiol. 164B: 278.
parameters in the cicada Diceroprocta olympu-
sa (Homoptera: Cicadidae). — Comp. Biochem. TOOLSON E.C. & N.F. HADLEY (1987): Energy-dependent
Physiol. 125A: 141. facilitation of transcuticular water flux contri-
butes to evaporative cooling in the Sonoran
SANBORN A.F. & P.K. PHILLIPS (1992): Observations on
Desert cicada, Diceroprocta apache (Homop-
the effect of a partial solar eclipse on calling in
tera: Cicadidae). — Jour. Exp. Biol. 131: 439.
some desert cicadas (Homoptera: Cicadidae). —
Florida Entomol. 75: 285. TOOLSON E.C. & E.K. TOOLSON (1991): Evaporative
cooling and endothermy in the 13-year periodi-
SANBORN A.F. & P.K. PHILLIPS (1996): Thermal responses
cal cicada, Magicicada tredecim (Homoptera,
of the Diceroprocta cinctifera species group
Cicadidae). — Jour. Comp. Physiol. 161B: 109.
(Homoptera: Cicadidae). — Southwestern Nat.
41: 136. TUTTLE M.D., RYAN M.J. & J. BELWOOD (1985): Acousti-
cal resource partitioning by t w o species of
SANBORN A.F. & P.K. PHILLIPS (2001): Re-evaluation of
phyllostomatid bats (Trachops cirrhosus and
the Diceroprocta delicata species complex
Tonatia silvicola). — Anim. Behav. 33: 1369.
(Homoptera: Cicadidae). — Ann. Ent. Soc. Amer.
94: in press. VRIJER P.W.F. de (1984): Variability in calling signals of
SCHEDL V.W. (1986): Zur Verbreitung, Biologie und the planthopper Javesella pellucida (F.) (Homop-
Ökologie der Singzikaden von Istrien und dem tera: Delphacidae) in relation to temperature,
angrenzenden Küstenland (Homoptera: and consequences for species recognition
Cicadidae und Tibicinidae). — Zool. Jahr. Abt. f. during distant communication. — Netherlands
Syst. Ökol. Geog. Tiere 113: 1. Jour. Zool. 34: 388.
SOPER R.S., SMITH L.F.R. & A.J. DELYZER (1976): Epizoo- WAKABAYASHI T. & S. HAGIWARA (1953): Mechanical and
tiology of Massospora levispora in an isolated electrical events in the main sound muscle of
population of Okanagana rimosa. — Ann. Ent. cicada. — Jap. Jour. Physiol. 3: 249.
Soc. Amer. 69: 275.
WAKABAYASHI T. & K. IKEDA (1961): Interrelation
STANLEY-SAMUELSON D.W., HOWARD R.W. & E.C. TOOLSON between action potential and miniture electri-
(1990): Phospholipid fatty acid composition and cal oscillation in the tymbal muscle of the
arachidonic acid uptake and metabolism by the cicada. — Jap. Jour. Physiol. 11: 585.
cicada Tibicen dealbatus (Homoptera: Cicadi-
WALKER T.J. (1957): Specificity in the response of
dae). — Comp. Biochem. Physiol. 97B: 285.
female tree crickets (Orthoptera, Gryllidae,
SWINTON A.H. (1908): The vocal and instrumental Oecanthinae) to calling songs of the males. —
music of insects. — Zoologist (4th Ser.) 12: 376. Ann. Ent. Soc. Amer. 50: 626.
469You can also read

















































