Individual differences in prefrontal cortical activation on the Tower of London planning task: implication for effortful processing
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
European Journal of Neuroscience, Vol. 17, pp. 2219–2225, 2003 ß Federation of European Neuroscience Societies
Individual differences in prefrontal cortical activation on the
Tower of London planning task: implication for effortful
processing
F. Cazalis,1 R. Valabrègue,1 M. Pélégrini-Issac,1 S. Asloun,2 T. W. Robbins3 and S. Granon4
1
INSERM U.483, 9 quai Saint-Bernard, 75005, Paris, France
2
Service de Réhabilitation Neurologique, Hospital Raymond Poincaré, 92380 Garches, France
3
Department of Experimental Psychology, University of Cambridge, Downing Street, CB2 3EB, UK
4
Unité ‘Récepteurs et Cognition’, Département de Neuroscience, Institut Pasteur, 25 rue du Dr Roux, 75015 Paris, France
Keywords: cingulate cortex, fMRI, human subjects, planning
Abstract
Solving challenginxg (‘effortful’) problems is known to involve the dorsal and dorsolateral prefrontal cortex in normal volunteers,
although there is considerable individual variation. In this functional magnetic resonance imaging study, we show that healthy subjects
with different levels of performance in the Tower of London planning task exhibit different patterns of brain activation. All subjects
exhibited significant bilateral activation in the dorsolateral prefrontal cortex, the anterior and posterior cingulate areas and the parietal
cortex. However, ‘standard performers’ (performance 70% correct) differed in
the patterns of activation exhibited. Superior performers showed a significantly more spatially extended activation in the left dorsolateral
prefrontal cortex than did standard performers, whereas the latter group tended to show increased activation of the anterior cingulate
region.
Introduction
Planning processes engage multiple cerebral areas, such as the dorso- which subjects compute sequential displacements of coloured balls in
lateral prefrontal cortex (DLPFC, Brodmann areas 9/46), the anterior order to match a predetermined arrangement. For the most difficult
and posterior cingulate and the posterior parietal cortex (Baker et al., problems, intermediate states of planning must be maintained ‘on-line’
1996; Elliott et al., 1997; Dagher et al., 1999; Tanji & Hoshi, 2001). An and manipulated to achieve the final goal. Indeed, the major cognitive
extensive literature shows the particular involvement of the DLPFC in processes for solving the TOL task are mental manipulation, WM
executive and complex behaviours (Goldman-Rakic, 1995; Miller, monitoring, sequential organization and sustained attention (Shallice,
1999; Carpenter et al., 2000; Owen, 2000; Fuster, 2001) and supports 1982; Baker et al., 1996; Owen et al., 1996a). The ability to conceive
the notion that it is a critical structure for mediating the manipulation and execute plans while manipulating information in working memory
of information in working memory and the organization of different is the essence of efficient problem solving and gives rise to a gradation
steps in planning processes (Baker et al., 1996; Owen et al., 1996b; of cognitive effort. In addition to the involvement of various working
Owen, 1997). Furthermore, challenging (‘effortful’) processing has memory loads, effortful processes also implicate the ability to maintain
been associated with strong activation of the DLPFC and cingulate a high level of attention and, in most tasks, to choose appropriate
cortex, as these areas have been shown to be more active in unpractised cognitive strategies. These processes are likely to introduce much
tasks requiring high attentional control (Paus et al., 1993; Raichle et al., variability in individual performance during problem solution (Schnei-
1994; Schneider et al., 1994; Barch et al., 1997; Petersen et al., 1998; der et al., 1994; Rypma et al., 2002). Indeed, in a pilot experiment we
Adler et al., 2001). Recently, working memory (WM) performance has observed that healthy subjects segregated into two groups based on
been shown to correlate with DLPFC activation (Rypma et al., 2002) level of performance (Granon et al., 1998).
while previous studies have shown that the DLPFC is more activated in The present work tested the hypothesis that differential levels of
tasks involving higher WM load (Manoach et al., 1997; Petersen et al., activation would be associated with the level of cognitive performance
1998) and during the encoding phase of a working memory task in healthy subjects performing at different levels of difficulty of the
(Rypma & D’Esposito, 1999). TOL task. As our aim was to investigate the role of the prefrontal
To investigate the relationship between performance in effortful cortex in planning and manipulating a variable number of items, we
processes and prefrontal cortex activation, we utilized a test of plan- therefore focused on imaging the dorsal part of the prefrontal cortex.
ning of variable difficulty: the Tower of London (TOL) task, during Our MRI paradigm and customised statistical parametric mapping of
data allowed us to investigate the causes of differential patterns
between healthy subjects with different performance in this particular
task. For this purpose, we analysed behavioural data correlated with
Correspondence: Dr Sylvie Granon, as above. activation data. The major finding of our study was that specific and
E-mail: granon@pasteur.fr
differential patterns of activation were associated with different levels
Received 22 October 2002, revised 6 March 2003, accepted 10 March 2003 of performance.
doi:10.1046/j.1460-9568.2003.02633.x2220 F. Cazalis et al.
Materials and methods be placed back into an available place before any other manipulation,
and that only one ball can be moved at a time. Subjects were asked to
Participants answer as soon as possible, with a maximum time allowed for respon-
Eleven healthy right-handed adults were studied (seven female, four ses of 30 s. A new problem was presented upon a response or if no
male; mean age 24.8, range 18–32 years; mean time in secondary response was made within 30 s. Subjects responded with their right
school education, 10.5 years, range 6–13 years; UK ‘A level’ equiva- hand on a 7-key pad (corresponding to how many moves were necess-
lent is 7 years). They all gave their informed written consent to ary for solution, ranging from 0 to 6).
participate in the experiment, which was approved (protocol 1664, The task was divided into three conditions: Control (0 or 1 move was
rbm 99017) by the local Ethics Committee at the Cochin Hospital in necessary to solve the problem), Easy (2 or 3 moves) and Difficult (4, 5
Paris, France (2000). Participants were divided into one group of five or 6 moves). The paradigm consisted of six 3-min runs. Runs were
‘standard performers’ (mean performance level for the difficult con- composed of two 1.5-min blocks combining two different conditions
dition 70% correct, mean age 27.5, range 24–32 years). As test condition at the beginning of each block.
subjects were naive for the task at the beginning of the experiment, Three behavioural variables were taken into account: total number
groups were defined on the basis of subjects’ performance during the of completed trials, accuracy of performance and response times.
functional magnetic resonance imaging (fMRI) experiment. The seg- Performance was calculated as the ratio (%) of correct responses over
regation criterion was chosen on the basis of previously observed the total number of completed trials (response given within 30 s).
individual performance in a pilot study, in which a larger population Response times were collected as the delay between the beginning of
exhibited a clear segregation in the difficult condition, with a cut-off at the presentation of the stimulus and the given response.
70% (Granon et al., 1998).
Cognitive data analysis
Cognitive paradigm The three behavioural variables (number of trials, performance and
The paradigm was a simplified TOL task (Baker et al., 1996). The task response times) were analysed using repeated measures of variance
was presented to the subjects just before the experiment to verify that with one between-subject factor with two levels (Group: Superior and
they understood the instructions. Two configurations of the TOL task Standard performers) and one within-subject factor with three levels
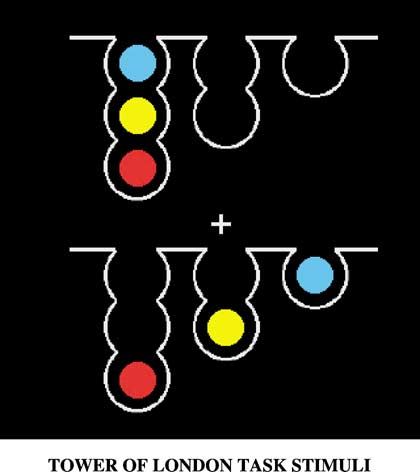
were presented on a screen (Fig. 1). Subjects were asked to compute (Condition: Control, Easy and Difficult).
mentally the minimum number of moves necessary to make the bottom
configuration resemble the top configuration and to give this response Imaging paradigm
on a special keyboard. Rules of the task are that each ball moved must The subjects were imaged using a standard 1.5-T scanner (Signa,
General Electric Medical Systems). The use of two neck pillows to
support subjects’ heads in the standard head coil avoided excessive
motion. A first (sagittal medial) T1-weighted run enabled location of
anatomical axial T1-weighted images (Spin-echo, TE 8 ms, TR
400 ms, voxel size 1.1 0.8 5.0 mm3) so that the eight contiguous
5-mm slices covered the superior part of the brain, including the
superior frontal sulcus and the rostral part of the cingulate gyrus.
Functional images were positioned at the same axial co-ordinates, i.e.
coregistered to anatomical images, so that an individual anatomical
study of blood oxygenated level-dependent (BOLD) measurement was
possible. Six runs were acquired, fitting the functional block design
(T2-weighted echo-planar images, TE 66 ms, TR 3000 ms, flip angle a
908, voxel size 2.2 1.6 5.0 mm3, 500 images per run). An anato-
mical 3-D scan was finally performed (T1-weighted Spoiled-Gradient
Recalled (SPGR), TE 6 ms, TR 35 ms, flip angle a 408, voxel size
0.9 1.4 1.5 mm3), in order to get unequivocal anatomical informa-
tion for each subject.
Imaging data analysis
The images were processed using SPM99 software (Wellcome Depart-
ment of Cognitive Neurology, UK; http://www.fil.ion.ucl.ac.uk/spm).
We chose to perform individual analyses based on individual anatomy.
Therefore, the data were not normalized on a common spatial brain.
Our procedure took individual anatomical variability into account and
aimed at avoiding loss of spatial resolution and uncertainty of the
position of individual sulci and anatomical structures due to normal-
ization (i.e. ‘fuzziness’; Postle & D’Esposito, 2000).
Fig. 1. The Tower of London task. Example of screen with which subjects were Pre-processing
presented. The top represents the goal configuration and the bottom one the
starting configuration. In the Control condition, 0 or 1 move was necessary to For each subject, data were corrected for within-run subject motion
copy the top arrangement; in the Easy condition, 2 or 3 moves were necessary; and spatially smoothed using an 8-mm full-width-at-half-maximum
in the Difficult condition, 4, 5 or 6 moves were required. Gaussian kernel.
ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–2225Prefrontal cortex activation and cognitive effort 2221
Statistical estimation of BOLD effect
The general linear model was used, for each subject, to detect brain
areas in which the time course of the measured BOLD signal was
significantly correlated with that of the experimental paradigm. Boxcar
waveforms, convolved with the canonical haemodynamic response
function, synchronized on individual response time values, were used
to model the three cognitive conditions. The effect of fatigue during the
experiment was modelled by a derivative of time. Finally, statistical
parametric maps of t-values were obtained by contrasting Difficult vs.
Control and Easy vs. Control conditions.
Individual quantitative analysis of activated voxels were computed
for each subject. Seven volumes of interest (VOIs) were delineated
manually and individually on the basis of individual structural T1
images (Diamond et al., 1985; Talairach & Tournoux, 1988; McMinn
et al., 1990; Netter & Dalley, 1997; Truwit & Lempert, 1997).
Inconsistency of nomenclature within the literature was resolved on
the basis of the Latin nomenclature (Tamraz, 1983). Individual anato-
mical variability, for example sulcal interruption, was considered and
elucidated following the incidence rates indicated by Ono and col-
leagues (Ono et al., 1990). The validity of the VOIs was verified by a
neuroradiologist, based on anatomical criteria. VOIs, delineated
regardless of subgroup membership, behavioural results and imaging
results, were defined as follows: whole functional brain, right superior
frontal sulcus, left superior frontal sulcus, anterior cingulate, posterior
cingulate, right intraparietal sulcus and left intraparietal sulcus. There- Fig. 2. Performances in the Tower of London task are represented as (A) the
fore, VOIs did not strictly correspond to Brodmann areas. The spatial percentage of correct responses and (B) response time in seconds, in Control,
extent of the activated areas was then individually quantified for each Easy and Difficult conditions, for Standard performers and for Superior
VOI as the number of suprathreshold voxels in each individual t-map, performers. Bars represent SEM.
for two contrasts: Difficult vs. Control and Easy vs. Control conditions,
yielding 14 comparisons per subject. We defined individual uncor-
rected thresholds so that the total number of activated voxels within the Response time
whole brain was the same for all subjects (Feydy et al., 2002). Statistical analyses showed no significant effect of Group (F1,9 < 1
Therefore, equivalent activation patterns were obtained for all subjects, (NS), a significant effect of Condition (F2,18 ¼ 26.12, P < 0.0001) and
i.e. quantification of thresholded voxels inside the whole functional no significant Group Condition interaction (F2,18 < 1, NS). Bonfer-
brain resulted in equivalent values for all subjects, thus enabling roni/Dunn post hoc tests showed that the two groups did not differ,
statistical comparisons between individuals. As the total number of whatever the condition (all P > 0.25). Post hoc t-tests showed that
voxels was the same for all subjects, we could calculate and compare response times differed significantly (P < 0.0001) for Control vs. Easy,
for each individual the spatial extent of activation in each VOI as a Easy vs. Difficult and Control vs. Difficult problems. As illustrated in
percentage of the total spatial extent of activation, i.e. 100 (spatial Fig. 2B, both groups of subjects showed increased response times with
extent for considered VOI)/(spatial extent for whole-volume VOI). task difficulty.
Results were collected as a matrix of two contrasts 7 VOI activa-
tions, for each subjects. Number of trials
Student’s t-tests were performed on the total number of completed
Group effect analysis trials per condition. Statistical analyses showed no significant effect of
A between-groups comparison of the spatial extent of activation was Group for either Control (P ¼ 0.34), Easy (P ¼ 0.088) or Difficult
performed using Student’s t-tests (StatView1 software) between (P ¼ 0.46) conditions, indicating that both groups completed a similar
Superior and Standard performers, for each VOI separately. This number of trials for each experimental condition.
analysis helped to determine which areas were significantly more
spatially activated in one group than in the other. Global spatial extent of activation
The global spatial extent of activation was represented by the sum of
Results spatial extents for all VOIs. Statistical analyses showed no significant
Group or Condition effect, nor a Group Condition interaction (all
Performance F < 1, NS). As illustrated in Fig. 3, ‘Superior’ and ‘Standard’ perfor-
There was a significant Group effect (F1,9 ¼ 14.4, P < 0.01), a mers exhibited comparable cerebral activity, whatever the difficulty of
significant effect of Condition (F2,18 ¼ 146.2, P < 0.0001) and a the task.
significant Group Condition interaction (F2,18 ¼ 16.8, P < 0.001).
Pairwise comparisons showed that the two groups did not differ for Between-subjects analyses: activation of the VOIs
either Control (F < 1, NS) or Easy conditions (F1,9 ¼ 1.1, NS), but As we hypothesized that one group would exhibit a more extended
were clearly different for the Difficult condition (F2,9 ¼ 28.5, BOLD signal than the other, we conducted one-tailed Student’s t-tests
P < 0.001). Figure 2A shows the performance of both groups in each to assess statistical differences between Groups for each VOI in both
condition. conditions. Results showed that Superior performers activated the left
ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–22252222 F. Cazalis et al.
exhibited larger activations in the left DLPFC than Standard perfor-
mers. Standard performers, conversely, exhibited a tendency to show
greater activations in the anterior cingulate than Superior performers.
This suggests that efficient problem solving elicits specific patterns of
brain activation involving the DLPFC and the anterior cingulate.
Previous functional imaging studies have identified the DLPFC as
being the neural substrate for different steps of planning processes
(Baker et al., 1996) and as being sensitive to memory load (Manoach
et al., 1997; Petersen et al., 1998; Rypma & D’Esposito, 1999), to the
type of memory processes (D’Esposito et al., 1999) and, more recently,
to working memory performance (Rypma et al., 2002). In this paper,
Fig. 3. Sum of spatial extent of activation of all volumes of interest, in Easy vs. we further explored the latter relationship in a complex planning task.
Control contrast and Difficult vs. Control contrast, for Standard performers and
for Superior performers. No significant difference was observed either between Behavioural data
conditions or between groups.
The present behavioural data confirmed the results of our pilot
experiment (Granon et al., 1998) suggesting that subjects mentally
frontal VOI to a significantly greater level than Standard performers in solving the TOL task segregated into two groups (Standard and
the Easy condition (P < 0.05) as well as in the Difficult condition Superior performers) with significantly different performance speci-
(P < 0.05). Standard performers showed a tendency to activate more fically for the Difficult condition. The groups did not differ for the
the anterior cingulate area than did Superior performers in the Easy Control and Easy conditions. Response times and number of com-
(P ¼ 0.063) and the Difficult (P ¼ 0.056) conditions (see Fig. 4A and pleted trials were also identical in the two groups, whatever the
B). There was no significant Group effect for any other VOI, in either difficulty. This suggests that all subjects exhibited similar levels of
condition. motivation to solve the task. The difference in performance cannot be
interpreted in terms of different social education between groups, as
we showed no significant difference between groups for this measure.
Discussion
We observed a significant effect of age, Standard performers being
We analysed the activation pattern of subjects performing the Tower of slightly younger than Superior performers. Though unexplained, such
London (TOL) task, a complex test of planning which offers different an increase of performance with age has been reported in children and
levels of difficulty. Our major findings were that Superior performers young adults by Krikorian et al. (1994).
Fig. 4. BOLD effect in all volumes of interest, in (A) Easy vs. Control contrast and (B) ‘Difficult vs. Control’ contrast, for Standard performers and for Superior
performers. The star represents a significant difference between the two groups for the left dorsolateral prefrontal cortex. The cross represents a marginally significant
difference between the two groups for the anterior cingulate area. Bars represent SEM.
ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–2225Prefrontal cortex activation and cognitive effort 2223
Table 1. Studies employing the Tower of London task. Therefore, despite the fact that the Easy condition did not require the
Imaging Effect of difficulty manipulation of a large number of items in working memory (e.g.
Reference methods Laterality on PFC activation moves), it can nevertheless be considered an effortful cognitive task
(Schneider & Shiffrin, 1977; Hasher & Zacks, 1979). D’Esposito and
Morris et al. (1993) SPECT Left Increased
collaborators showed that dual-task solving (D’Esposito et al., 1995)
Baker et al. (1996) PET Bilateral Increased
Owen et al. (1996a) PET Left Increased and manipulation rather than maintenance in memory (D’Esposito
Elliot et al. (1997) PET Bilateral Increased et al., 1999) produced increased DLPFC activation. Thus, the DLPFC
Owen et al. (1998) PET Left No effect activation we observed in all subjects in both conditions may be
Dagher et al. (1999) PET Bilateral Increased elicited by our particular paradigm and related to effortful processing.
Lazeron et al. (2000) fMRI Bilateral No effect
Rowe et al. (2001) PET Left Not tested This view is supported by data showing increased DLPFC activation
specifically during effortful encoding (Rypma & D’Esposito, 1999;
Adler et al., 2001; Rypma et al., 2002). Therefore, it is likely that our
extended DLPFC activation in Superior performers reflects effortful
Planning activation and differential patterns related to processes associated with efficient planning.
performance level Moreover, Callicott et al. (1999) used an n-back task (in which the
Suprathreshold activation was found in all anatomical VOIs, chosen subject must respond when seeing a stimulus that appeared ‘‘n-back’’
according to previously reported data related to TOL solutions: in a sequence) to show that the amount of activation in the DLPFC
bilateral dorsolateral prefrontal cortex, anterior cingulate, posterior increases with n up to two items and then decreases, reflecting the
cingulate and bilateral posterior parietal cortices. This pattern confirms behavioural and concomitant neural inflection point at the upper limit
previous results obtained with the TOL task (Baker et al., 1996; Owen of performance. This result is particularly interesting because it shows
et al., 1996a; Dagher et al., 1999; Lazeron et al., 2000; Cools et al., that extended DLPFC activation may not be linearly related to working
2002). However, in our study the two groups of subjects with different memory span. Following this view, our Standard performers might use
levels of performance exhibited differential activation in terms of the a larger working memory span whereas Superior performers might
spatial extent of activation. Activation in the left DLPFC was found to search for the critical subgoal correctly organized in time, therefore
be significantly more spatially extended in Superior performers than in reducing working memory span (Ward & Allport, 1997) and thus use a
Standard performers, in both the Easy and Difficult conditions. Con- more efficient cognitive strategy. This hypothesis is supported by
versely, the anterior cingulate tended to show more spatially extended recent results which show that efficient planners for the mental solving
activation in Standard performers than in Superior performers, in both of the TOL task are able to select visually the critical intermediate
the Easy and Difficult conditions. steps necessary to reach the ultimate goal whereas lower performers
The role of the DLPFC in working memory and planning processes have difficulty in avoiding the gaze toward irrelevant items (Hodgson
involved in the solving of the Tower of London task is well-estab- et al., 2000). Several computational modelling approaches for the TOL
lished, although the laterality of effect is still controversial (see Table 1 task have been designed (Dehaene & Changeux, 1997; Ward &
for a summary). Our results showed a significant BOLD signal in the Allport, 1997; Gilhooly et al., 1999). These models either examine
DLPFC in both groups of subjects for both experimental conditions. the performance level (Ward & Allport, 1997; Gilhooly et al., 1999) or
We furthermore observed that Superior performers exhibited more the role of the prefrontal cortex in this task (Dehaene & Changeux,
spatially extended activation in the left DLPFC than did Standard 1997). The latter model illustrates particularly that complex planning
performers in both conditions, suggesting that spatial extent of activa- tasks can be solved using a step-by-step process, involving a functional
tion in this area may not be related to increased difficulty per se but to architecture comprising reward, plans, operation and move units, as
other factors influencing performance, such as strategy selection and well as residual goal and working memory units. The authors showed
attentional capacity. Several studies have indeed reported a lack of that increasing task difficulty necessarily results in increased working
influence of task difficulty in DLPFC activation (Owen et al., 1998; memory load and therefore increased error rate. This is consistent with
Lazeron et al., 2000), whereas others have emphasized such an the model of Gilhooly et al. (1999) which further emphasized the
influence (Manoach et al., 1997; Dagher et al., 1999; see also Table 1). importance of temporal organization both within and between plan-
Our results may therefore appear to contradict some earlier results ning steps. It should be noted that the temporal organization of a
obtained with the TOL task showing that regional cerebral blood flow sequence of action is a function which is crucially dependent upon the
increases with difficulty in the DLPFC. It should be noted, however, DLPFC activation (Fuster, 2001). Consequently, it is plausible that the
that a more effortful task does not necessarily lead to a greater spatial step-by-step model illustrates the usual way of solving the TOL task,
extent of BOLD activation, in terms of number of activated voxels, but such as the one used by untrained subjects. On the other hand, the
could result in a higher BOLD intensity signal reflecting a higher ‘move-selection’ model of Ward and Allport (1997) suggested that
frequency of neuronal firing (Logothetis et al., 2001; Kim & Ogawa, performance on the TOL task depends on the selection of action
2002). It has also been reported that enhanced activity in the DLPFC between concurrent moves. They defined superior performers as being
may correlate with a less effective memory encoding (Otten & Rugg, better at selecting subgoal moves, defined as ‘moves that are essential
2001). Overall, these observations lead to the suggestion that BOLD to the optimum solution, but which do not place a disc into its goal
signal intensity measures and quantification of activated voxels may position’, thus predicting the behavioural results of Hodgson et al.
relate to different neurophysiological processes and thus give com- (2000). Subjects with better performance, as compared to standard
plementary information. Furthermore, the TOL task was previously performers, would be able to recognize and then achieve the critical
performed on a touch screen and did not require mentally imagining step, as defined above. They are able to break down one difficult
the moves, as in the present study. These differences in procedure may problem into several subproblems of lower difficulty. Therefore, this
have induced either the use of different strategies as compared to our move-selection strategy would place much less load on working
study, or a major difference in the method of solving the Easy task. memory and so would be associated with fewer errors. In our study,
Indeed, in our case, solving the Easy condition required subjects to Superior performers may use such a move-selection strategy instead of
mentally move balls while counting them, thus constituting a dual task. the step-by-step strategy used by Standard performers. It is noteworthy
ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–22252224 F. Cazalis et al.
that different strategies would not necessarily lead to different res- Baker, S.C., Rogers, R.D., Owen, A.M., Frith, C.D., Dolan, R.J., Frackowiak,
ponse times. Indeed, Ward & Allport (1997) showed that the planning R.S.J. & Robbins, T.W. (1996) Neural systems engaged by planning: a PET
study of the Tower of London task. Neuropsychologia, 34, 515–526.
time in the TOL task does not vary with WM load. It is plausible that Barbas, H. (2000) Connections underlying the synthesis of cognition, memory,
the relationships between performance level and strategy described in and emotion in primate prefrontal cortices. Brain Res. Bull., 52, 319–330.
those models, although not explicitly tested in our experiment, can Barch, D.M., Braver, T.S., Nystrom, L.E., Forman, S.D., Noll, D.C. & Cohen,
account for the differential pattern of activation between our two J.D. (1997) Dissociating working memory from task difficulty in human
prefrontal cortex. Neuropsychologia, 35, 1373–1380.
groups of subjects. This would be consistent with the recent findings of Barch, D.M., Braver, T.S., Sabb, F.W. & Noll, D.C. (2000) Anterior cingulate
Rypma et al. (2002). and the monitoring of response conflict: evidence from an fMRI study of
In summary, while we have shown here that all healthy subjects overt verb generation. J. Cogn. Neurosci., 12, 298–309.
activated areas commonly implicated in spatial planning tasks (bilat- Callicott, J.H., Mattay, V.S., Bertolino, A., Finn, K., Coppola, R., Frank, J.A.,
Goldberg, T.E. & Weinberger, D.R. (1999) Physiological characteristics of
erally the DLPFC, the anterior cingulate, posterior cingulate and
capacity constraints in working memory as revealed by functional MRI.
bilateral posterior parietal cortex) we also provide some new insight Cereb. Cortex, 9, 20–26.
into the individual differences in activation of the network mainly Carpenter, P.A., Just, M.A. & Reichle, E.D. (2000) Working memory and
consisting of the left DLPFC and the anterior cingulate area. Our executive function: evidence from neuroimaging. Curr. Opin. Neurobiol., 10,
results for effortful planning are in agreement with recent data 195–199.
Carter, C.S., Braver, T.S., Barch, D.M., Botvinick, M.M., Noll, D. & Cohen,
suggesting that these brain areas are differentially activated depending J.D. (1998) Anterior cingulate cortex, error detection, and the online mon-
upon cognitive performance (Rypma et al., 2002), with the cingulate itoring of performance. Science, 280, 747–749.
activation being dependent upon task difficulty (Barch et al., 1997, Carter, C.S., Macdonald, A.M., Botvinick, M., Ross, L.L., Stenger, A., Noll, D.
2000), conflict resolution or error detection (Carter et al., 1998, 2000). & Cohen, J.D. (2000) Parsing executive processes: strategic vs. evaluative
Indeed, the anterior cingulate area is mostly engaged in Standard functions of the anterior cingulate cortex. Proc. Natl Acad. Sci. USA, 97,
1944–1948.
performers whereas Superior performers recruit markedly the left Cools, R., Stefanova, E., Barker, R.A., Robbins, T.W. & Owen, A.M. (2002)
DLPFC. As subjects were naı̈ve to the task at the beginning of the Dopaminergic modulation of high-level cognition in Parkinson’s disease: the
experiment, it is likely that Superior performers were quickly able to role of the prefrontal cortex revealed by PET. Brain, 125, 584–594.
adapt and generate efficient strategy. This form of rapid mental D’Esposito, M., Detre, J.A., Alsop, D.C., Shin, R.K., Atlas, S. & Grossman, M.
(1995) The neural basis of the central executive system of working memory.
flexibility for effortful cognitive tasks is well described by the work- Nature, 378, 279–281.
space model (Dehaene et al., 1998). In this model, the anterior D’Esposito, M., Postle, B.R., Ballard, D. & Lease, J. (1999) Maintenance versus
cingulate and the DLPFC are represented as strongly correlated areas manipulation of information held in working memory: an event-related fMRI
(Barbas, 2000) active in effortful tasks because of their long distance study. Brain Cogn., 41, 66–86.
cortico-cortical connectivity. This theoretical reflection is supported by Dagher, A., Owen, A.M., Boecker, H. & Brooks, D.J. (1999) Mapping the
network for planning: a correlational PET activation study with the Tower of
recent experimental studies, suggesting that the left DLPFC and the London task. Brain, 122, 1973–1987.
anterior cingulate cortex are both engaged and communicate for Dehaene, S. & Changeux, J.P. (1997) A hierarchical neuronal network for
effortful conflict resolution (Carter et al., 2000; MacDonald et al., planning behavior. Proc. Natl Acad. Sci. USA, 94, 13293–13298.
2000), and the fact that subjects with lower performance in the TOL Dehaene, S., Kerszberg, M. & Changeux, J.P. (1998) A neuronal model of a
global workspace in effortful cognitive tasks. Proc. Natl Acad. Sci. USA, 95,
task may have difficulty in solving conflict resolution between relevant 14529–14534.
and irrelevant items useful for the intermediate planning steps (Hodg- Diamond, M.C., Scheibel, A.B. & Elson, L.M. (1985) The Human Brain
son et al., 2000). Methodologies such as the ones developed in the Coloring Book. Harper Collins, New York.
latter studies and that enable the analysis of the temporal dynamics of Elliott, R., Baker, S.C., Rogers, R.D., O’Leary, D.A., Paykel, E.S., Frith, C.D.,
Dolan, R.J. & Sahakian, B.J. (1997) Prefrontal dysfunction in depressed
signal change should allow further evaluation of the respective role of
patients performing a complex planning task: a study using positron emission
the areas contributing to the workspace model. tomography. Psychol. Med., 27, 931–942.
Feydy, A., Carlier, R., Roby-Brami, A., Bussel, B., Cazalis, F., Pierrot, L.,
Burnod, Y. & Mayer, M.A. (2002) Longitudinal study of motor recovery after
Acknowledgements stroke: recruitment and focusing of brain activation. Stroke, 33, 1610–1617.
We are grateful to R. Toro and B. Mendiburu for programming the task. We Fuster, J.M. (2001) The prefrontal cortex – An update: time is of the essence.
acknowledge Pr. J-P. Changeux, Pr. P. Azouvi, Dr Y. Burnod, Dr B. Gutkin, Dr Neuron, 30, 319–333.
A. Cressant and Dr N. Mechawar and two anonymous referees for valuable Gilhooly, K.J., Phillips, L.H., Wynn, V., Logie, R.H. & Della Sala, S. (1999)
comments on a previous version of the manuscript. We thank the volunteers who Planning processes and age in the five-disc Tower of London task. Thinking
took part in the study. F.C and R.V. were supported by the French Ministry of Reasoning, 5, 339–361.
Research. Part of this work was supported by the French Foundation for Goldman-Rakic, P. (1995) Architecture of the prefrontal cortex and the central
Medical Research (to S.G.) T.W.R. was supported by the Wellcome Trust. executive. Proc. Natl Acad. Sci. USA, 769, 71–83.
Granon, S., Anton, J.L., Dauchot, K., Kouneiher, F., Cazalis, F., Jolivet, O.,
Bittoun, J., Pierrot, L., Azouvi, P., Rogers, R., Robbins, T.W., Vidal, C. &
Burnod, Y. (1998) A functional MRI study of the human prefrontal cortex
activity as afunction of WM load using the new tower of london task. Soc.
Abbreviations Neurosci. Abstr., 24, 1896.
BOLD, blood oxygenated level-dependent; DLPFC, dorsolateral prefrontal Hasher, L. & Zacks, R.T. (1979) Automatic and effortful processes in memory.
cortex; fMRI, functional magnetic resonance imaging; SPGR, Spoiled-Gradient J. Exp. Psychol. Gen., 108, 356–388.
Recalled; TOL, Tower of London; VOI, volume of interest; WM, working Hodgson, T.L., Bajwa, A., Owen, A.M. & Kennard, C. (2000) The strategic
memory. control of gaze direction in the Tower-of-London task. J. Cogn. Neurosci.,
12, 894–907.
Kim, S.-G. & Ogawa, S. (2002) Insights into new techniques for high resolution
References functional MRI. Curr. Opin. Neurobiol., 12, 607–615.
Krikorian, R., Bartok, J. & Gay, N. (1994) Tower of London procedure: a
Adler, C.M., Sax, K.W., Holland, S.K., Schithorst, V., Rosenberg, L. & standard method and developmental data. J. Clin. Exp. Neuropsychol., 16,
Strakowski, S.M. (2001) Changes in neuronal activation with increasing 840–850.
attention demand in healthy volunteers: An fMRI study. Synapse, 42, Lazeron, R.H., Rombouts, S.A., Machielsen, W.C., Scheltens, P., Witter, M.P.,
266–272. Uylings, H.B. & Barkhof, F. (2000) Visualizing brain activation during
ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–2225Prefrontal cortex activation and cognitive effort 2225 planning: the Tower of London test adapted for functional MR imaging. Am. Paus, T., Petrides, M., Evans, A.C. & Meyer, E. (1993) Role of the human J. Neuroradiol., 21, 1407–1414. anterior cingulate cortex in the control of occulomotor, manual, and speech Logothetis, N.K., Pauls, J., Augath, M., Trinath, T. & Oeltermann, A. (2001) responses: a positron emission tomography study. J. Neurophysiol., 70, Neurophysiological investigation if the basis of the fMRI signal. Nature, 412, 453–469. 150–157. Petersen, S.E., Van Mier, H., Fiez, J.A. & Raichle, M.E. (1998) The effects of MacDonald, A.W., Cohen, J.D., Stenger, V.A. & Carter, C.S. (2000) Dissociat- practice on the functional anatomy of task performance. Proc. Natl Acad. Sci. ing the role of the dorsolateral prefrontal and anterior cingulate cortex in USA, 95, 853–860. cognitive control. Science, 288, 1835–1838. Postle, B.R. & D’Esposito, M. (2000) Evaluating models of the topographical Manoach, D.S., Schlaug, G., Siewert, B., Darby, D.G., Bly, B.M., Benfield, A., organization of working memory function in frontal cortex with event-related Edelman, R.R. & Warach, S. (1997) Prefrontal cortex fMRI signal changes fMRI. Psychobiology, 28, 132–145. are correlated with working memory load. Neuroreport, 8, 545–549. Raichle, M.E., Fiez, J.A. & Videen, H. (1994) Practice-related changes in McMinn, R.M., Hutchings, H., R.T. & Logan, B.M. (1990) Anatomie de la tête human brain functional anatomy during nonmotor learning. Cereb. Cortex, 4, et du cou. Editions André Delcourt, Paris. 8–26. Miller, E.K. (1999) The prefrontal cortex: complex neural properties for Rowe, J.B., Owen, A.M., Johnsrude, I.S. & Passingham, R.E. (2001) Imag- complex behavior. Neuron, 22, 15–17. ing the mental components of a planning task. Neuropsychologia, 39, Morris, R.G., Ahmed, S., Syed, G.M. & Toone, B.K. (1993) Neural correlates of 315–327. planning ability: frontal lobe activation during the Tower of London test. Rypma, B., Berger, J.S. & D’Esposito, M. (2002) The influence of working- Neuropsychologia, 31, 1367–1378. memory demand and subject performance on prefrontal cortical activity. J. Netter, F.H. & Dalley, A.F.I.I. (1997) Atlas of Human Anatomy. Novartis, East Cogn. Neurosci., 14, 721–731. Hanover. Rypma, B. & D’Esposito, M. (1999) The roles of prefrontal brain regions in Ono, M., Kubik, S., Abernathey, C.D. & Yasargil, M.G. (1990) Atlas of the components of working memory: effects of memory load and individual Cerebral Sulci. Georg Thieme Verlag, Stuttgart, New York. differences. Proc. Natl Acad. Sci. USA, 96, 6558–6563. Otten, L.J. & Rugg, M.D. (2001) When more means less: neural activity related Schneider, W., Pimm-Smith, M. & Worden, M. (1994) Neurobiology of to unsuccessful memory encoding. Curr. Biol., 11, 1528–1530. attention and automaticity. Curr. Opin. Neurobiol., 4, 177–182. Owen, A.M. (1997) Tuning in to the temporal dynamics of brain activation Schneider, W. & Shiffrin, R.M. (1977) Controlled and automatic information using functional magnetic resonance imaging (fMRI). Trends Cogn. Sci., 1, processing. I. Detection, search, and attention. Psychol. Rev., 84, 1–53. 123–125. Shallice, T. (1982) Specific impairments of planning. Phil. Trans. Roy. Soc. Owen, A.M. (2000) The role of the lateral frontal cortex in mnemonic Lond. B: Biol. Sci., 298, 199–209. processing: the contribution of functional neuroimaging. Exp. Brain Res., Talairach, J. & Tournoux, P. (1988) Co-Planar stereotaxis atlas of the human 133, 33–43. brain. George Thieme Verlag, Stuttgart. Owen, A.M., Doyon, J., Dagher, A., Sadikot, A. & Evans, A.C. (1998) Tamraz, J. (1983) Atlas D’anatomie Céphalique Dans le Plan Neuro-Oculaire Abnormal basal ganglia outflow in Parkinson’s disease identified with (P.N.O). University of Paris 6, Paris. PET implications for higher cortical functions. Brain, 121, 949–965. Tanji, J. & Hoshi, E. (2001) Behavioral planning in the prefrontal cortex. Curr. Owen, A.M., Doyon, J., Petrides, M. & Evans, A.C. (1996a) Planning and Opin. Neurobiol., 11, 164–170. spatial working memory: a positron emission tomography study in humans. Truwit, C.L. & Lempert, T.E. (1997) High Resolution Atlas of Cranial Eur. J. Neurosci., 8, 353–364. Neuroanatomy. Hanil Medical Book Center, DPS Press-Williams & Wilkins, Owen, A.M., Evans, A.C. & Petrides, M. (1996b) Evidence for a two-stage Baltimore, USA. model of spatial working memory processing within the lateral frontal cortex: Ward, G. & Allport, A. (1997) Planning and problem-solving in the five-disc a positron emission tomography study. Cereb. Cortex, 6, 31–38. tower of london task. Quart. J. Exp. Psychol. A, 50, 49–78. ß 2003 Federation of European Neuroscience Societies, European Journal of Neuroscience, 17, 2219–2225
You can also read

















































