Tunicates Illuminate the Enigmatic Evolution of Chordate Metallothioneins by Gene Gains and Losses, Independent Modular Expansions, and Functional ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Tunicates Illuminate the Enigmatic Evolution of Chordate
Metallothioneins by Gene Gains and Losses, Independent
Modular Expansions, and Functional Convergences
Sara Calatayud,1 Mario Garcia-Risco,2 Oscar Palacios,2 Mercè Capdevila,2 Cristian Ca~
nestro,1 and
Ricard Albalat* ,1
1
Departament de Genètica, Microbiologia i Estadıstica and Institut de Recerca de la Biodiversitat (IRBio), Facultat de Biologia,
Universitat de Barcelona, Barcelona, Catalonia, Spain
2
Departament de Quımica, Facultat de Ciències, Universitat Autonoma de Barcelona, Cerdanyola del Vallès, Spain
*Corresponding author: E-mail: ralbalat@ub.edu.
Associate editor: Naruya Saitou
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
Abstract
To investigate novel patterns and processes of protein evolution, we have focused in the metallothioneins (MTs), a
singular group of metal-binding, cysteine-rich proteins that, due to their high degree of sequence diversity, still represents
a “black hole” in Evolutionary Biology. We have identified and analyzed more than 160 new MTs in nonvertebrate
chordates (especially in 37 species of ascidians, 4 thaliaceans, and 3 appendicularians) showing that prototypic tunicate
MTs are mono-modular proteins with a pervasive preference for cadmium ions, whereas vertebrate and cephalochordate
MTs are bimodular proteins with diverse metal preferences. These structural and functional differences imply a complex
evolutionary history of chordate MTs—including de novo emergence of genes and domains, processes of convergent
evolution, events of gene gains and losses, and recurrent amplifications of functional domains—that would stand for an
unprecedented case in the field of protein evolution.
Key words: metallothionein domains, modular proteins, Chordata, Tunicata, ascidians/thaliaceans/appendicularians,
metallothionein evolution.
Introduction domains, in which each domain enables the coordination of a
Heavy metals such as zinc (Zn) and copper (Cu) are integral number of metal ions through metal–thiolate bonds
components of numerous proteins involved in multiple cel- (reviewed in Blindauer [2014]). MTs have been classified in
lular functions (reviewed in Vallee and Falchuk [1993]; Maret a stepwise gradation, from extreme Zn/Cd-thioneins to ex-
Article
[2013]; Vest et al. [2013]), whereas other metals such as cad- treme Cu-thioneins depending on their metal-binding pref-
mium (Cd) have no known biological functions and are highly erence (Bofill et al. 2009; Palacios, Atrian, et al. 2011). There
toxic for living beings. Organisms have developed different have been attempts to predict the metal preference of a given
molecular mechanisms that regulate the homeostasis of the MT from the analysis of their non-Cys amino acids, and in the
physiological metals and counteract the harmful effects of the case of gastropod MTs, for instance, it has been proposed that
toxic ones. One of such mechanisms is based on an extremely the lysine-asparagine (K/N) ratio could be important for
diverse group of metal-binding proteins known as metallo- metal preference, so that a preponderance of K over N resi-
thioneins (MTs), which are engaged in the physiological con- dues would be a feature of Cd selectivity, whereas a predom-
trol of metals operating as ion reservoirs, metal transporters inance of N over K would be typical of Cu-MTs (Perez-Rafael
and/or metal deliverers to target metalloproteins, but also in et al. 2014; Pedrini-Martha et al. 2020). Due to the high se-
radical scavenging, oxidative stress protection, and antiapop- quence and structural diversity of MTs, however, it is unclear
totic defense (reviewed in Capdevila et al. [2012]; Ziller and whether the rule of K/N ratio applies to nongastropod MTs,
Fraissinet-Tachet [2018]). and thereby, metal-binding assays are needed to experimen-
Most MTs are low molecular weight (Calatayud et al. . doi:10.1093/molbev/msab184 MBE
molecular evolution of these genes and proteins. MT research planktonic and benthic species, as well as solitary and colonial
has exposed, for instance, pervasive events of internal se- forms. We have also functionally characterized the metal-
quence duplications (Tanguy and Moraga 2001; Palacios, binding properties of eight tunicate MTs, which were selected
Espart, et al. 2014; Iturbe-Espinoza et al. 2016; Jenny et al. on the basis of their taxon representativeness, sequence com-
2016; Calatayud et al. 2018), de novo emergence of domains position, degree of multiplicity, or modular organization.
(Calatayud, Garcia-Risco, Pedrini-Martha, et al. 2021), and a Comparative analyses of tunicate MTs with those of cepha-
high bias in the amino acid composition probably as a result lochordates and vertebrates have exposed a high degree of
of the 3D structural requirements for metal binding. MT re- structural diversity between lineages, which would not have
search has also provided useful information for many other been accompanied, however, by differences in their metal
research fields. In the field of Evolutionary Ecology and preferences. This diversity seems the result of numerous
Environmental Sciences, for instance, MTs are considered a and intricate evolutionary events, and therefore, chordate
good model to investigate the mechanisms by which different MTs would represent one of the most complex and enig-
organisms—from protists and fungi to insects, mollusks, and matic examples of protein evolution.
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
mammals—have adapted to diverse metal bioavailability and
other environmental stress conditions. These mechanisms Results
include duplications of MT genes (Maroni et al. 1987; The biological interest for tunicates in the last decades has
Adamo et al. 2012; Luo et al. 2020), increases in the levels given rise to many genomic and transcriptomic resources,
of MT expression (Timmermans et al. 2005; Janssens et al. facilitating the in silico identification of virtually any gene in
2008, 2009; Costa et al. 2012; Catalan et al. 2016; de Francisco a wide set of species. This advantageous situation allowed us
et al. 2018), and changes in metal specificity (Tio et al. 2004; to perform an extensive survey of the MT system in tunicate
Palacios, Pagani, et al. 2011; de Francisco et al. 2017; Dallinger databases.
et al. 2020; Garcıa-Risco et al. 2020). In the fields of
Bioinorganic Chemistry and Metallomics, MTs have been MTs in Ascidians
used to analyze metal–protein interactions, analyzing the Ascidians, commonly known as sea squirts, are the most
qualitative and quantitative characteristics of the metal- studied group of tunicates. Adults are sessile, either solitary
binding capacities of these proteins (Blindauer and or colonial, usually found in shallow water worldwide. We
Leszczyszyn 2010; Palacios, Pagani, et al. 2011; Beil et al. surveyed ascidian databases and identified 145 MT sequences
2019) and by extension, of a broad range of metalloproteins from 37 species distributed among Aplousobranchia (three
(Waldron et al. 2009). species), Phlebobranchia (eight species), and Stolidobranchia
In this work, we aim to explore the MTs found in a large (3 molgulidae þ 5 pyuridae þ 18 styelidae species) orders (fig.
number of species of the three tunicate classes—Ascidiacea, 1; see supplementary table S1, Supplementary Material on-
Thaliacea, and Appendicularia—in order to investigate the line, for the species list). Sequence comparison showed that
intriguing origin and evolution of MTs in chordates. most ascidian MTs are 38–45 amino acids long with 12 cys-
Tunicates (Tunicata subphylum, a.k.a. Urochordata) are the teines (31–27%) organized in a single functional domain. In
sister group of vertebrates, and together with basally branch- this domain, the C-motifs were distributed as
ing cephalochordates form the phylum Chordata. Tunicates [CxC]x5[CxC]x2Cx2Cx2[CxC]x4[CxC]x3-6CC, which would
have undergone a rapid and “liberal” pattern of evolution in represent the prototypical organization of the mono-
comparison with the “conservative” pattern of vertebrates modular ascidian MTs (supplementary fig. S1,
and cephalochordates (Somorjai et al. 2018; Ferrandez- Supplementary Material online, and table 1).
Roldan et al. 2019), yielding a group of ecologically diverse Twenty-seven out of the 37 analyzed species showed mul-
filter-feeding marine animals—including planktonic and ben- tiple MT genes: seven species with two sequences, six with
thic specimens as well as solitary and colonial forms–, which three, two with four, three with six, two with seven, four with
have adapted to different conditions of metal bioavailability. eight, one with nine, and two species with up to ten sequen-
In addition, the filtering lifestyle makes tunicates prone to ces. Despite in some cases, due to the nature of the data
accumulate metals from the seawater (Papadopoulou and source—mainly sequence read archive (SRA) projects–, allelic
Kanias 1977; Tzafriri-Milo et al. 2019; and references therein), variants of a particular gene could not be discriminated from
and thereby, molecular mechanisms such as the MTs that gene duplicates, overestimating thereby the degree of MT
regulate metal homeostasis and detoxification are physiolog- multiplicity in our catalog, our results suggested that, overall,
ically relevant to them. the MT multiplicity in ascidians is pervasive, although a de-
Our understanding of the MTs in tunicates is, however, finitive evaluation of the degree of multiplicity must await
scarce, limited to two MT sequences from the ascidian Ciona more complete and deeper genomic databases.
robusta (formerly C. intestinalis) and Herdmania curvata Interestingly, there were some exceptions to the prototyp-
(Franchi et al. 2011), and two sequences from the appendi- ical organization, ranging from MTs that have lost or gained
cularian Oikopleura dioica recently described by our group some cysteines or small protein fragments (supplementary fig.
(Calatayud et al. 2018). To overcome this situation, we have S1, Supplementary Material online) to multimodular forms
conducted an exhaustive survey of the MTs in tunicate data- containing tandem repeats of the prototypical domain (sup-
bases, identifying 168 MTs in 37 species of ascidians, 4 of plementary figs. S2 and S3, Supplementary Material online).
thaliaceans, and 3 of appendicularians, representing We were able to reconstruct 12 multimodular MTs (five partial
4436Enigmatic Evolution of Chordate Metallothioneins . doi:10.1093/molbev/msab184 MBE
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
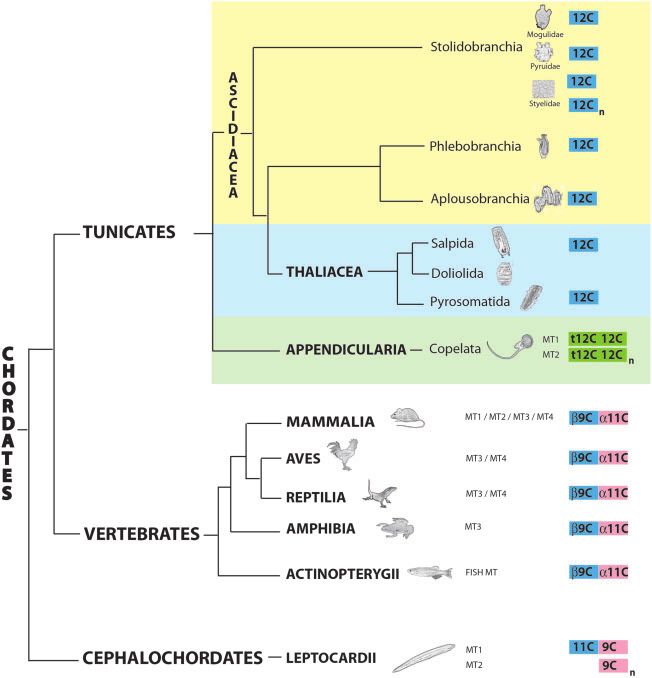
FIG. 1. Phylogenetic relationships in the Chordata phylum. Regarding tunicates, traditional classifications divided this subphylum in three classes,
Appendicularia (green background), Ascidiacea (yellow background), and Thaliacea (blue background), but recent analyses propose Thaliacea
species (salps, doliolids, and pyrosomes) are nested within the Ascidiacea class, closer to the Aplousobranchia and Phlebobranchia orders than to
the Stolidobranchia order (Delsuc et al. 2018; Kocot et al. 2018). A schematic representation of the MTs of each taxonomic group, with a mono-, bi-,
or multimodular organization with distinct domains (color coded) containing seven, nine, 11, or 12 cysteines, is showed at the right (see fig. 7 and
text for additional details).
Table 1. Cysteine Motifs in 11/12-Cys Domains from Different Animal Taxa.
Taxon 11/12-Cys Cysteine Motifs Metal Ions
Domains
Ascidians/thaliaceansFull-length MT [CxC]x5[CxC]x2Cx2Cx2[CxC]x4[CxC]x3–6CC 4 (this work)
Appendicularians 12C domain Cx3Cx2Cx2Cx3Cx2[CxC]x2Cx4[CxC]xCC 4 (Calatayud, Garcia-Risco, Capdevila, et al. 2021)
Vertebrates a-domaina CCxCCx3Cx2Cx3[CxC]x4–12Cx1–3CC 4 (Stillman et al. 1987)
Cephalochordates 11C domain [CxC]x5[CxC]x3–4[CxC]x3[CxC]x3[CxC]x2C nd
Mollusks a-domain [CxC]x4–6[CxC]x3Cx4–5[CxC]x3[CxC]x3[CxC]x2C 4 (Digilio et al. 2009)
Echinoderms a-domain CxCCx5[CxC]x4CCx4CCx4CC 4 (Tomas et al. 2013)
Insects MTnB/C/D Cx2Cx3[CxC]x4Cx3[CxC]x3[CxC]xC]x7CC 4 (Egli et al. 2006)
NOTE.—nd, not determined.
a
Notice that although vertebrate, echinoderm, and mollusk MTs have a-domains, the origin and evolutionary relationship of these a-domains remains uncertain.
and seven full-length sequences), whose number of repeats CroMT1 from C. robusta (phlebobranchia order), and
ranged from two to nine (fig. 2). Noteworthy, all the species HroMT1 and HroMT2 from Halocynthia roretzi (stolidobran-
with multimodular MTs belonged to the styelidae family in the chia order). These MTs were chosen because CroMT1 was the
stolidobranchia order (fig. 1), suggesting that multimodular only MT in C. robusta, whereas HroMT1 and HroMT2 repre-
MTs arose by a lineage-specific evolutionary event. sented duplicate forms in H. roretzi. These MTs were also
selected because of their K/N ratios, which in the CroMT1
Metal-Binding Features of Mono-Modular Ascidian was 3:5, opposite of that in HroMT1 and HroMT2 sequences,
MTs 6:3 and 8:2, respectively. HroMT2 showed indeed one of the
In order to analyze the metal-binding capacity and preference highest K/N ratios among tunicate MTs (supplementary table
of the ascidian MTs, we selected three MTs of two species S1, Supplementary Material online). The K/N ratio seems im-
belonging to distinct orders within the Ascidiacea class: portant for metal preference of gastropod MTs (Perez-Rafael
4437Calatayud et al. . doi:10.1093/molbev/msab184 MBE
FIG. 2. Schematic representation of multimodular Stelydae MTs. Internal duplications of a 12-Cys domain (grey box) generated multimodular MTs
with variable number of repeats (R), from 2 to 9. The number of MTs with different number of repeats (R1–R9) is indicated (DgrMT7 ¼ 2 repeats;
EtiMT8part 4 repeats; ScaMT6part and SsoMT8part 6 repeats; PmiMT10part, BleMT1, and BleMT2 7 repeats; PauMT10part and PspMT7
8 repeats; BscMT1, PmamiMT3, and PpoMT7 ¼ 9 repeats) (see supplementary fig. S2, Supplementary Material online, for further details).
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
et al. 2014; Pedrini-Martha et al. 2020), and the study of the Metal-Binding Features of Multimodular Ascidian
metal selectivity of the CroMT1, HroMT1, and HroMT2 MTs
would allow us to experimentally evaluate the K/N rule in In order to determine the metal-binding features of multi-
the ascidian system. modular MTs, we selected BscMT1 (ID: ATSW01006707.1;
We characterized the formation of metal–MT com- chromosome 6) of Botryllus schlosseri (stolidobranchia order)
plexes of CroMT1, HroMT1, and HroMT2 proteins heter- because this MT was not only identified in an accurately
ologously expressed in Escherichia coli cultures assembled genome (Voskoboynik et al. 2013), but also be-
supplemented with Cu, Cd, or Zn salts by inductively cou- cause it was supported by three EST sequences (supplemen-
pled plasma atomic emission spectroscopy (ICP-AES) and tary table S1, Supplementary Material online). BscMT1 was a
electrospray ionization mass spectrometry (ESI-MS). ICP- multimodular MT of 365 amino acids with 105 cysteines
AES was used for protein quantification and metal-to- (29%) and a K/N ratio of 1:1 (31K vs. 31N) (supplementary
protein stoichiometry determination through the mea- fig. S2 and table S1, Supplementary Material online). BscMT1
surement of element composition of the samples (S, Zn, was one of the longest MT described so far, and a detailed
Cd, and Cu) (Bongers et al. 1988), and ESI-MS allowed us to analysis of its sequence revealed that it was made of nine
determine the molecular mass of the species formed, that tandem repeats (R1–R9, the last one only partially complete)
is, the speciation of the samples (Capdevila et al. 2012). of a 12-Cys domain, each one resembling a prototypical as-
Single well-folded Cd4-MT species were recovered from cidian form (supplementary fig. S3, Supplementary Material
CroMT1, HroMT1, and HroMT2 Cd-productions (fig. online).
3A–C), and a major Zn4-MT species were found in their The metal–BscMT1 complexes purified from the heterol-
recombinant Zn productions (supplementary fig. S4A, C, ogous expression in E. coli cultures supplemented with Cu,
and E, Supplementary Material online), which contrasted Cd, or Zn salts were characterized by the IPC-AES and ESI-MS.
with the mixtures of homometallic Cu-MT species purified The concentrations of metal–BscMT1 productions, despite
from Cu-supplemented E. coli cultures (supplementary fig. the several attempts, were always low, probably due to the
S4B, D, and F, Supplementary Material online). It is also high molecular weight of BscMT1 and its structural complex-
noteworthy the resemblance of the CD spectra of the Cd– ity made of repeated domains. Only the samples recovered
MT complexes of all ascidian MTs, denoting structurally from the Zn(II)-enriched culture media rendered informative
equivalent metal clusters for these proteins (fig. 4). The data through these techniques. The IPC-AES and ESI-MS anal-
exemplary exciton coupling centered at 250 nm exhib- yses showed that this preparation contained Zn36–MT com-
ited in all the CD envelopes confirms high robustness and plexes (fig. 3D). The binding of 36 metal ions agreed with its
compactness of the Cd4S(Cys)12 clusters and, therefore, multimodular organization made of nine repeated domains,
high Cd specificity. Altogether, these results showed that each one similar to a prototypical M4-MT: 4 metal ions9
Zn4- or Cd4–MT complexes were energetically the most domains ¼ 36 metal ions. To further verify the functional
favorable forms of these proteins, suggesting that their correspondence between one of the 12-Cys tandem repeats
function would be related to binding divalent metal ions, of BscMT1 and the mono-modular ascidian MTs, we analyzed
either as Zn- or Cd-thioneins [8]. Genuine Zn-thioneins, the ability of the R4 domain (R4-BscMT1) to form metal
however, render heterometallic Cu, Zn–MT complexes complexes when it was expressed alone. The election of the
when synthesized under Cu surplus, meaning that the as- R4 domain was due to the fact that it was highly similar to the
cidian MTs deviated toward a more specific Cd-thionein prototypical ascidian MTs, including some additional amino
behavior (Palacios, Atrian, et al. 2011). Regarding the K/N acids after the carboxyl-terminal CC doublet (supplementary
rule used to predict the metal-binding preference in gas- fig. S3, Supplementary Material online). We heterologously
tropod MTs, our results demonstrated that this rule did expressed the R4 domain in E. coli grown in media supple-
not apply to tunicate MTs, since the three of them dis- mented with Cu, Cd, or Zn salts. The ICP-AES and ESI-MS
played contrasted K/N ratios and their metal preferences analyses of the recovered samples confirmed the ability of this
were exactly the same. fragment to autonomously form metal complexes by itself,
4438Enigmatic Evolution of Chordate Metallothioneins . doi:10.1093/molbev/msab184 MBE
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
FIG. 3. Deconvoluted ESI-MS spectra of the Cd(II)-productions of ascidian CroMT1 (A), HroMT1 (B), HroMT2 (C), and þ14 charge state ESI-MS
spectra of the Zn(II)-production of BscMT1 (D), as well as the deconvoluted ESI-MS spectra of the Cd(II)-production of its fragment R4 BscMT1 (E).
Notice that BscMT1 is capable to bind up to 36 divalent ions, which nicely match with a multimodular organization made of nine repeated
domains (12C)9, each one binding four divalent ions alike the mono-modular (12C) MTs (A–C) or the R4 (12C) domain (E).
and revealed that the biochemical features of R4 domain were
highly similar to the mono-modular ascidian forms (compare
fig. 3A–C and E). Thus, this domain yielded major M4–R4
complexes with Zn(II) and Cd(II), being unique species only
in the latter, whereas rendered a homometallic mixture of
species in the presence of copper, as the ascidian MTs dis-
cussed above (supplementary fig. S4G and H, Supplementary
Material online). Based on this R4 fragment as a representa-
tion of the whole protein, the multidomain BscMT1 would be
a Cd-thionein capable to bind up to 36 divalent ions, and its
stoichiometry—that is, M36-BscMT1—would nicely match
FIG. 4. Circular dichroism spectra of the Cd(II)-productions of with what would be expected from the repetition of nine
CroMT1, HroMT1, HroMT2, BscMT1, and R4 BscMT1, all exhibiting domains, that is, M4-R1 to M4-R9. These results supported a
very similar CD signals. The exciton coupling centered at 250 nm direct relationship between the binding capacity and the
confirms high robustness and compactness of the clusters formed. number of the repeated domains in multimodular MTs.
4439Calatayud et al. . doi:10.1093/molbev/msab184 MBE
MTs in Thaliaceans demonstrated that the four SthMTs were functionally very
We analyzed the thaliacean MTs by surveying the databases similar among them (compare panels A–D in fig. 5), as well as
of the five available species. We identified 14 MTs from four to the mono-modular ascidian MT (compare fig. 3 and fig. 5).
thaliacean species, three from Salpida (Iasis cylindrica, Salpa The fact that thaliacean (SthMTs) and ascidian (CroMT1,
fusiformis, and Salpa thompsoni), and one from Pyrosomata HroMT1, HroMT2, and BscMT1) MTs exhibited a similar
(Pyrosomella verticillata) orders. Sequence comparison Cd-thionein character strongly supported that the prototyp-
showed that thaliacean MTs were 36–41 amino acids long, ical ascidian/thaliacean MT (and most likely the ancestral
with 12 cysteines (33–29%) organized in a single functional tunicate MT, see below) was a mono-modular protein with
domain with the C-motifs distributed as a 12C domain capable to bind four metal ions with preference
[CxC]x5[CxC]x2Cx2Cx2[CxC]x4[CxC]x3–4CC (noteworthy, we for Cd.
did not find multimodular MTs in thaliacean databases) (ta-
ble 1). This motif arrangement was identical to that of the MTs in Appendicularians
prototypical ascidian MTs, denoting a common origin of that We investigated the MTs in the Appendicularia (a.k.a.
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
thaliacean and ascidian MTs, and supporting the recently Larvacea) class. In a previous work, we had characterized
suggested phylogenetic relationship of these classes (Delsuc the OdiMT1 and OdiMT2 genes (formerly OdMT1 and
et al. 2018; Kocot et al. 2018) (fig. 1). OdMT2) of O. dioica (Calatayud et al. 2018). We showed
Importantly, MT multiplicity was found in the four thalia- that both genes encoded MTs very different from all MTs
cean species although as in ascidians, it was challenging to described so far because their cysteines motifs were arranged
determine whether the multiple sequences represented dif- in a novel pattern forming a “repeat unit” (RU) made of two
ferent genes or allelic variants. In S. thompsoni, for instance, C7 subunits (C7a and C7b) plus a carboxyl-terminal tail with
we identified six MT sequences, five full-length (SthMT1, five cysteines (C5) (Calatayud et al. 2018) (supplementary fig.
MT2, MT3, MT5, and MT6), and one partial (SthMT4), which S6A, Supplementary Material online). We concluded that
coded for proteins 48% to 95% identical at amino acid level OdiMT1 was made of a single RU, whereas OdiMT2 was a
(supplementary table S2, Supplementary Material online). multimodular protein made of six direct tandem RU
The highest identities were between SthMT1 and SthMT5 (Calatayud et al. 2018). We wondered whether these uncon-
(88%) and SthMT2 and SthMT6 (95%) sequences, and there- ventional MTs were specific of O. dioica—gene and protein
fore, these pairs were the most plausible cases of allelic var- sequences are known to have considerably diverged during O.
iants. To investigate this possibility, we analyzed the dioica evolution (reviewed in Ferrandez-Roldan et al.
conservation of 50 and 30 flanking regions, as well as of intron [2019])—or they were widespread in the appendicularian
sequences in PCR-amplified genomic regions from two S. lineage. We took advantage of recent sequence projects of
thompsoni specimens. Nucleotide identity (excluding gaps) several appendicularian species (Naville et al. 2019) and iden-
in SthMT1–SthMT5 and SthMT2–SthMT6 alignments was tified seven new MT sequences in two additional Oikopleura
91–100%, 87–82% and 83–86% at 50 regions, introns and 30 species: O. albicans (three sequences) and O. vanhoeffeni (four
regions, respectively (supplementary table S3, Supplementary sequences) (supplementary table S1, Supplementary Material
Material online). These results would be compatible with that online). We did not identify any putative MT in databases of
SthMT5 and SthMT6 were allelic variants of SthMT1 and other oikopleurids such as O. longicauda, Bathochordaeus sp.,
SthMT2, respectively, although the possibility they may be and Mesochordaeus erythrocephalus, nor in the fritillarid spe-
recently duplicated genes cannot be fully discarded. cies Fritillaria borealis, although any conclusion about the loss
of the MTs in these lineages must await deeper sequence
Metal-Binding Features of Thaliacean MTs projects.
We analyzed the metal-binding capacity and metal prefer- Regarding Oikopleura MTs, although most of the sequen-
ence of the thaliacean MTs, and compared them with those ces were partial, they were clearly similar to O. dioica MTs.
of ascidian MTs in order to investigate whether the differ- Sequence analyses showed that these new MTs were multi-
ences in lifestyles—planktonic versus benthic—might have modular proteins made of repeats of a conserved 12-Cys
been key for MT evolution. We characterized the metal– domain with C-motifs distributed as
MT complexes rendered for the four MTs from S. thompsoni, CX3Cx2Cx2Cx3Cx2[CxC]x2Cx4[CxC]xCC (table 1 and supple-
SthMT1 to SthMT4, heterologously expressed in Zn-, Cd-, or mentary fig. S6B, Supplementary Material online). This 12-Cys
Cu-supplemented E. coli cultures. The clear Cd-thionein char- domain corresponded to the previously described C7bþC5
acter of S. thompsoni MTs was reflected, once again, by the subunits (Calatayud et al. 2018). OdiMT1 consist, therefore, of
ICP-AES and ESI-MS analyses of the recovered samples. two 12-Cys domains, one full-length (12C ¼ C7b þ C5), and
SthMTs yielded single Cd4-MT species under Cd(II) surplus one 12-Cys domain that has been trimmed (t-12C, formerly
(fig. 5), as well as major M4–MT complexes with Zn(II) (sup- known as C7a) and lacks the five last cysteines (C5) (supple-
plementary fig. S5A–D, Supplementary Material online). A mentary fig. S6A, Supplementary Material online and fig. 7).
mixture of species (for SthMT1 and SthMT3) or not detection Interestingly, the trimmed domain when expressed alone did
of the metallated species (for SthMT2 and StMT4) were not efficiently bind divalent metals by itself, suggesting that it
obtained in the presence of copper (supplementary fig. S5E has lost its capacity to independently work (Calatayud,
and F, Supplementary Material online), evidencing that these Garcia-Risco, Capdevila, et al. 2021). OdiMT2 would be a
proteins struggle to build metal clusters with Cu. Our results multimodular MT made of twelve 12-Cys domains, whose
4440Enigmatic Evolution of Chordate Metallothioneins . doi:10.1093/molbev/msab184 MBE
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
FIG. 5. Deconvoluted ESI-MS of the Cd(II)-production of S. thompsoni SthMT1 (A), SthMT2 (B), SthMT3 (C), and to SthMT4 (D). Thaliacean MTs
are mono-modular proteins made of a single 12C domain capable to bind four divalent ions, identically to ascidian MTs.
odd domains would have been also trimmed as t-12C. The from genomic sequence in B. belcheri species (BbeMT2L and
identification of this conserved 12-Cys domain in all S).
Oikopleura MTs suggested that this domain appeared in Sequence comparisons revealed that cephalochordate
the ancestor of the Oikopleura genus, and that it was dupli- MT2s were multimodular proteins made of two (MT2S) or
cated generating multimodular MTs independently in the three (MT2L) 9-Cys domains (CCx2Cx2Cx3Cx[CxC]xCC)
diverse appendicularian species (supplementary fig. S6B, depending on an alternative splicing on exon 2 (supplemen-
Supplementary Material online). tary figs. S7 and S8B, Supplementary Material online). In con-
trast, MT1s were bimodular proteins with two different
domains: a 11-Cys, N-terminal domain with the C-motifs dis-
MTs in Other Chordates tributed as [CxC]x5[CxC]x3–4[CxC]x3[CxC]x3[CxC]x2C (table
To provide a broad evolutionary perspective to our analyses 1 and supplementary fig. S8A, Supplementary Material on-
we compared the tunicate MTs with those from the other line) linked by 9–11 residues to a domain similar to the 9-Cys
chordate subphyla: cephalochordates and vertebrates. domain of MT2 (we named this domain 9-CysLike because an
In cephalochordates, MTs had been characterized in two Ile substitutes the fourth Cys: CCx2Cx2Ix3Cx[CxC]xCC), fol-
amphioxus species, Branchiostoma floridae and Branchiostoma lowed by a carboxyl-terminal tail of 16–17 amino acids devoid
lanceolatum (Guirola et al. 2012). We extended the surveys of of Cys residues. Interestingly, MT2 9-Cys and the MT1 9-
MTs to two additional amphioxus species, Branchiostoma bel- CysLike domains showed a clear sequence similarity (supple-
cheri (BbeMT1 and BbeMT2) and Branchiostoma japonicum mentary fig. S8C, Supplementary Material online), which sug-
(BjaMT1 and BjaMT2), and to Asymmetron lucayanum gested that cephalochordate MT2s might have arisen from
(AluMT1 and AluMT2), a cephalochordate distantly related the duplication of an ancestral MT1 that lost the 11-Cys
to the other Branchiostoma species (Yue et al. 2014). We domain but amplified its 9-Cys domain.
identified two MTs, MT1 and MT2, in all the cephalochordate In vertebrates, MTs had been classified in four different MT
species. B. lanceolatum MT2 showed two alternatively spliced types: MT1, MT2, MT3, and MT4. Vertebrate MTs had been
forms, MT2-L (for the long form, with 67–70 amino acids) and characterized in many species with significant differences of
MT2-S (for the short form, with 46–47 amino acids) (supple- multiplicity in a species-dependent manner (Seren et al. 2014).
mentary fig. S7, Supplementary Material online). Splice var- Phylogenetic and synteny analyses had suggested the existence
iants were also identified in B. floridae, B. japonicum, and A. of mammalian MT1s and MT2s, amniote MT4s, tetrapod
lucayanum cDNA databases (i.e., BflMT2L and S, BjaMT2L and MT3s, and fish MTs (fig. 1) (Seren et al. 2014). The appearance
S, and AluMT2L and S), and alternative splicing was predicted of these four types would not be related with the two rounds
4441Calatayud et al. . doi:10.1093/molbev/msab184 MBE
of whole-genome duplication events that occurred during 2019) appear a recurrent evolutionary solution in MTs of
early vertebrate evolution since they are not localized in paral- phylogenetically distant animal clades, from vertebrates and
ogous chromosomes but are located in tandem on a same tunicates to mollusks and insects (table 1). 3D structural
chromosome (Seren et al. 2014). At protein level, all vertebrate constraints to efficiently coordinate four metal ions through
MTs (even an MT we identified in the early divergent group of the formation of metal–thiolate bonds might have led to the
hagfish Eptatretus burgeri), showed a conserved bimodular convergent evolution of 11/12-Cys domains in MTs of differ-
organization made of a 9-Cys N-terminal domain (the so- ent animal phyla (Stillman et al. 1987; Egli et al. 2006; Digilio et
called b-domain: [CxC]x5[CxC]x3[CxC]x2[CxC]x2Cx3) linked al. 2009; Tomas et al. 2013; Calatayud, Garcia-Risco, Capdevila,
to a 11-Cys domain (the so-called a-domain: et al. 2021; and this work).
CCxCCx3Cx2Cx3[CxC]x6CxCC) at the carboxyl-terminal end
(supplementary table S1 and fig. S9, Supplementary Material Evolution of the Metal-Binding Preference
online). The arrangement of the C-motifs in the a and b Ascidian/thaliacean MTs and appendicularian MTs also share
domains did not show any obvious resemblance between the Cd-thionein character. The evolution of Cd-MTs seems,
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
them that might suggest of a common ancestry of these indeed, a common adaptive event in marine animals, includ-
domains, though vertebrate b-domain appeared somewhat ing mollusk (Calatayud, Garcia-Risco, Pedrini-Martha, et al.
similar to cephalochordate and tunicate 11/12-Cys domains. 2021), crustacean (Narula et al. 1995; Valls et al. 2001;
Munoz et al. 2002), echinoderm (Riek et al. 1999), and chor-
date species (Guirola et al. 2012; Calatayud et al. 2018). The
Discussion preference of tunicate MTs for Cd further support the idea
Origin and Convergent Evolution of Tunicate MTs that ancestral MTs arose and evolved probably driven by the
Ascidian and thaliacean MTs share the same arrangement of Cd(II) concentrations in the seawater because Cd is highly
their C-motifs (table 1) and are clearly similar in their amino toxic by interfering in Zn-dependent cellular processes
acid sequence, also in the noncysteine residues: ascidian MTs (Dallinger et al. 2020; Calatayud, Garcia-Risco, Pedrini-
are at least 40% identical in their noncysteine residues to Martha, et al. 2021). In addition to sporadic episodes of
any of the thaliacean MTs, and 60% identical when consid- increases in Cd emissions throughout geological eras (from
ering the full sequence. In contrast, their similarity with Paleozoic to Mesozoic and Cenozoic; Dallinger et al. [2020]
appendicularian MTs is negligible at the arrangement of the and references therein), Cd is frequently found with Zn in ore
C-motifs (table 1) and in the composition of the noncysteine deposits of the earth crust. The natural Cd(II) concentrations
residues. For instance, OdiMT1 only is, at most, 13% identical are, therefore, higher where seawater comes into contact with
to ascidian (e.g., CroMT1 and HroMT1) or thaliacean (e.g., the continental earth crust (0.8 nmol/kg) than in surface
SthMT1) MTs in noncysteine residues (notice that 25–30% pelagic waters (0.002 nmol/kg) (Bruland 1980). Cd(II) con-
sequence identity is generally taken as the minimal threshold centrations are also higher in eutrophic coastal regions of the
for presumption of homology; Rost 1999). Therefore, al- littoral and its close neritic zone (0.16 nmol/kg) (Bruland
though it cannot be ruled out that appendicularian MTs 1980), which are usual habitats for tunicates. It is noteworthy
derive from ascidian and thaliacean MTs that have diverged that all tunicate MTs analyzed in this work show a similar
too much for orthology to be recognized, their sequence and sequence and metal preference, either they are from benthic
the C-motifs are so divergent that they seem to have had ascidians or from planktonic thaliaceans, or from solitary (C.
independent evolutionary origins. This possibility agrees with intestinalis and H. roretzi) or colonial species (B. schlosseri).
the idea that de novo emergence of MTs might be more This similarity suggests that the different lifestyles of tunicates
habitual than previously thought because the only require- might have not been a significant factor in the evolution of
ment for a peptide to function as a metal ion chelator would metal-binding preference.
be a high content of coordinating residues (i.e., cysteines) and
a relative small length that would favor its proper folding Evolution of MT Multiplicity and Multimodular MTs
(Capdevila and Atrian 2011; Calatayud, Garcia-Risco, Increasing the metal-binding capacity enhance the physiolog-
Pedrini-Martha, et al. 2021). The independent emergence of ical capabilities of the organisms to adapt to diverse condi-
MTs in distant animal groups have been, indeed, implicitly tions of metal bioavailability and other environmental stress
suggested in diverse evolutionary studies (Capdevila and situations (Jenny et al. 2016; Baumann et al. 2017). In a sim-
Atrian 2011; Blindauer 2014; Isani and Carpene 2014; Ziller plified view, such increments might be attained through two
and Fraissinet-Tachet 2018), and we have recently showed de strategies: increasing the number of MT genes (i.e., augment-
novo evolution of MT domains within the Mollusca phylum ing MT multiplicity) (Maroni et al. 1987; Adamo et al. 2012;
(Calatayud, Garcia-Risco, Pedrini-Martha, et al. 2021). Luo et al. 2020) or increasing the number of metal-binding
Intriguingly, ascidian/thaliacean MTs and appendicularian domains in a given MT (i.e., generating multimodular MTs)
MTs seem to have converged in the number of cysteines— (Jenny et al. 2016; Baumann et al. 2017). Tunicates have used
12—of their basic functional organizations: the prototypical both strategies.
12-Cys MTs of ascidians and thaliaceans, and the repeated 12-
Cys domains of appendicularian MTs. Functional domains Evolution of MT Multiplicity
with 11/12 cysteines organized in different patterns of C- Multiplicity in the MT system has been associated to neo-
motifs but converging to similar 3D architectures (Beil et al. functionalization or subfunctionalization processes, in which
4442Enigmatic Evolution of Chordate Metallothioneins . doi:10.1093/molbev/msab184 MBE
different paralogs have distinct metal-binding preferences The Enigmatic Origin of the MTs in Chordates
and/or expression patterns. For instance, in mollusks, several The extensive catalog of tunicate MTs collected in this work
MTs can be found in a given species, each one with distinct together with the analysis of cephalochordate and vertebrate
metal selectivity—that is, Cd-MT, Cd/Cu-MT, and Cu-MT— MTs has allowed us to investigate the origin and evolution of
and differential transcription patterns (Palacios, Perez-Rafael, the MTs in the chordate phylum. This has been a particularly
et al. 2014; Dvorak et al. 2018; Dallinger et al. 2020). Similarly, difficult challenge because when comparing distant taxo-
in the mammalian system, ubiquitous metal-induced forms nomic groups (e.g., at phylum, subphylum, or class level),
MT1 and MT2 have preference for Zn(II), whereas restrictedly one has to be cautious with the MT sequence alignments.
expressed MT3 and MT4 have intermediate Zn/Cu- and Cu- The general cysteine richness of the MTs distorts the com-
thionein characters (Capdevila and Atrian 2011; Vasak and parisons by forcing the alignment of Cys positions and insert-
Meloni 2011; Artells et al. 2013). MT multiplicity has been ing many gaps in the aligned sequences. This distortion results
observed in 36 tunicate species, patchily distributed among in a false impression of similarity between nonhomologous
the three tunicate classes. However, in contrast with the ob- MT sequences, and if the alignments are used for phyloge-
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
served in the MT system of other animal groups, the analyzed netic reconstructions, they can lead to wrong evolutionary
tunicate MT paralogs do not seem to have essentially di- inferences. To circumvent this drawback, we focused our
verged in their metal-binding selectivity. The two paralogs analyses on the patterns of the cysteine motifs (CxC, CC,
of the ascidian H. roretzi (HroMT1 and HroMT2), the four and CCC) in the MT domains that, as it has been proved
paralogs of the thaliacean S. thompsoni (SthMT1 to SthMT4), for other molecular markers (Rokas and Holland 2000), might
and the two paralogs of the appendicularian O. dioica have enormous potential as a tool for phylogenetic
(OdiMT1 and OdiMT2; Calatayud et al. 2018) have a con- inferences.
served Cd selectivity. These results suggest that tunicate Regarding the ancestral chordate MT, it is generally ac-
paralogs have undergone a subfunctionalization process par- cepted that due to their substantial genomic stasis
titioning the expression of the predecessor, rather than a (Ca~ nestro and Albalat 2012; Somorjai et al. 2018; Coppola
neofunctionalization event acquiring new metal preferences. et al. 2019), cephalochordate genes and genomes are more
Future detailed spatiotemporal analyses of the transcription similar to those of the chordate ancestor than those of tuni-
patterns of each paralog will provide a definitive answer to cates and vertebrates (Louis et al. 2012). It can be argued,
this question, and will allow us to understand the ecophysi- therefore, that the MT of the chordate ancestor was a bimod-
ological needs that led the evolution of multiple MTs in these ular protein with an amino-terminal 11/12-Cys domain and a
species. carboxyl-terminal 9-Cys domain (fig. 6), resembling the orga-
nization of the current cephalochordate MT1s. The similarity
of cephalochordate and vertebrate domains (fig. 7) supports
Evolution of Multimodular MTs the bimodular organization of the ancestral chordate MT. In
Evolution of multimodular MTs seems a recurrent solution addition, the fact that cephalochordate MT1s are also similar
for creating large MTs with high cysteine content and a high to the a and b1 domains of mollusks (Calatayud, Garcia-
capacity of metal binding (Niederwanger et al. 2017; Palacios Risco, Pedrini-Martha, et al. 2021) would push back in evolu-
et al. 2017; Calatayud et al. 2018; and this work). tionary time this MT organization to the origins of the bilat-
Multimodular MTs have been found in very diverse organ- erian animals.
isms, from fungi (Palacios, Espart, et al. 2014; Iturbe-Espinoza The evolutionary scenario of a chordate MT ancestor sim-
et al. 2016) and mollusks (Calatayud, Garcia-Risco, Pedrini- ilar to the current cephalochordate MT1 implies a number of
Martha, et al. 2021; and references therein), to chordates evolutionary events independently shaping the MTs in each
(Guirola et al. 2012; Calatayud et al. 2018). Here, we have chordate subphylum: (I) In cephalochordates, an ancient tan-
identified multimodular MTs in 20 species of ascidian and dem duplication of the ancestral MT followed by the loss of
appendicularian classes (thaliaceans appear devoid of multi- the 11-Cys domain and the amplification of the 9-Cys domain
modular MTs). These multimodular MTs are made of tan- in one of the copies would have led the MT2 forms of the
dem repeats of a prototypical organization within each subphylum (fig. 6); (II) In vertebrates, the ancestral MT would
tunicate class—[CxC]x5[CxC]x2Cx2Cx2[CxC]x4[CxC]x3–6CC have readjusted its Cys motifs, leading to the current amino-
for ascidian and CX3Cx2Cx2Cx3Cx2[CxC]x2Cx4[CxC]xCC for terminal b- and the carboxyl-terminal a-domains (fig. 6).
appendicularian MTs–, implying independent evolutionary Successive lineage-specific tandem duplications would have
origins for ascidian and appendicularian multimodular MTs. led to a variable degree of the MT multiplicity in the different
In ascidians, the multimodular MTs arose during the evolu- vertebrate species (Seren et al. 2014); (III) In tunicates, the
tion of the Styelidae family in the Stolidobranchia order, after ancestral MT would have lost the 9-Cys domain in the ascid-
its split from the Pyuridae family, 277 Ma (Delsuc et al. 2018). ian/thaliacean lineage. This mono-modular 12-Cys MT was
In appendicularians, multimodular MTs arose after its diver- patchily duplicated, giving rise to a variable number of MTs in
gence from the other tunicate classes, 447 Ma (Delsuc et al. different ascidian/thaliacean species, or internally repeated in
2018). The adaptive causes that favored the evolution of large the multimodular MTs of styelidae species (fig. 6); (IV) Finally,
MT in some benthic ascidian species but not in others, or in in the fast–evolving appendicularians, the ancestral chordate
some planktonic tunicates such as appendicularians but not MT would have been lost, and a new MT with a different
in thaliaceans, are still an unsolved question. pattern of Cys motifs seems to have evolved de novo (fig. 6).
4443Calatayud et al. . doi:10.1093/molbev/msab184 MBE
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
FIG. 6. Reconstruction of MT evolution in the Chordata phyla. The most parsimonious scenario would be that the ancestral chordate MT was a
bimodular MT made of an N-terminal 11/12-Cys domain (in blue) and a C-terminal 9-Cys domain (in red), similar to the current cephalochordate
MT1. In cephalochordates, this ancestral form was tandem duplicated. One copy lost the 11/12-Cys domain and expanded the 9-Cys domain,
yielding the current bi/trimodular MT2 form. In vertebrates, the number of cysteines in each domain was readjusted to the current 9 Cys of the N-
terminal b-domains, and 11 Cys of the C-terminal a domain. Successive lineage-specific tandem duplications of this primeval vertebrate MT led to
different MT types (MT1, MT2, MT3, and MT4) with a variable degree of multiplicity in different species. In tunicates, the ancestral form lost the C-
terminal 9-Cys domain in ascidian/thaliacean lineages, leading a mono-modular. This MT was frequently duplicated generating a high degree of
MT multiplicity in many ascidian and thaliacean species. In contrast, internal duplications led to the emergence of multimodular MTs in the
stelydae lineage. Finally, in the appendicularian clade, the ancestral MT appears to have been lost, and a new MT would have emerged de novo by
the duplication of a 12-Cys domain (in green). During O. dioica evolution, the new bimodular MT lost five Cys in the N-terminal domain, yielding a
7-Cys/12-Cys arrangement (as it is found in current OdiMT1). Next, this 7-Cys/12-Cys MT was internal duplicated generating multimodular MTs
(as it is OdiMT2).
The primeval appendicularian MT would have been a bimod- independent amplifications of domains that generated multi-
ular form made of two 12-Cys domains, apparently unrelated modular MTs with high metal-binding capacities. All these
with the other 12-Cys domains of chordates. This ancestral results have led us to reconstruct an intricate evolutionary
appendicularian MT lost five Cys in the N-terminal domain in history for the chordate MTs, from a bimodular, cadmium-
the O. dioica lineage (but remained intact in other Oikopleura selective form in the last common ancestor, to a high struc-
lineages), yielding the t-12C þ 12C organization of the current tural and functional MT diversity currently found in the di-
OdiMT1. This MT was later duplicated, and internal duplica- verse species of the phylum. To discover the adaptive causes
tions in one of the copies of a repeat unit (RU) made of the t- that shaped this history and the molecular mechanisms that
12C þ 12C domains gave rise to the large multimodular made it possible will certainly be a fascinating challenge.
OdiMT2 (Calatayud et al. 2018).
In summary, our analyses have shown: 1) de novo emer- Materials and Methods
gence of MT genes and domains during chordate diversifica- Chordate MT Search in Databases
tion; 2) convergent evolution to 12-Cys domains in chordate Tunicate and cephalochordate MT sequences were retrieved
MTs, but also in other animals, probably favored by essential from public databases (NCBI database: https://www.ncbi.nlm.
constraints of the metal coordination; 3) convergent evolu- nih.gov/; and Octopus database: http://octopus.obs-vlfr.fr/
tion in the metal preference for Cd ions, probably triggered by public/botryllus/blastbotryllus.php) by BlastP and TBlastN
an ancestral requirement for cadmium-selective forms in searches using known chordate MTs as queries: tunicate
many marine animals; 4) lineage-specific events of gene dupli- CroMT1 (ACN32211), HcuMTa (AY314949), OdiMT1
cations yielding significant levels of MT multiplicity; and 5) (AYN64372), and OdiMT2 (AYN64376); cephalochordate
4444Enigmatic Evolution of Chordate Metallothioneins . doi:10.1093/molbev/msab184 MBE
FIG. 7. Comparison of the arrangement of Cys motifs in different MT domains. The arrangement of the Cys motifs in the 11-Cys amino-terminal
domain of cephalochordate MT1s are somewhat similar to that in the vertebrate 9-Cys b domains and in the ascidian/thaliacean 12-Cys MTs (blue
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
boxes), whereas the Cys motifs in the carboxyl-terminal domains of cephalochordate MT1s and MT2s (9C and 9CLike domains) are vaguely similar
to those in vertebrate 11-Cys a domains (pink boxes). In contrast, the arrangement of the Cys motifs in the appendicularian MT1 seems totally
different (green boxes).
BflMT1 (XP_035672826), BflMT2 (BW841405), BlaMT1 DNA Polymerase (Invitrogen, Thermo Fisher Scientific,
(BL11229), and BlaMT2 (JT872034); and vertebrate Waltham, MA) in a final 25 ll reaction. PCR conditions
HsaMT1A (NP_005937), HsaMT2A (NP_005944), HsaMT3 were 98 C 30 s (s); 35 cycles of 98 C 10 s, 50–54 C 30 s
(NP_005945), and HsaMT4 (NP_116324). In addition, RNA- (supplementary table S4, Supplementary Material online)
SRA for each tunicate species deposited in NCBI were BLAST and 72 C 3 min; and 72 C 10 min. PCR products were visu-
searched using as queries MT sequences from the nearest alized in 0.8% agarose gels, isolated with the GelElute Plasmid
phylogenetic species as well as from different species covering Miniprep Kit (Sigma-Aldrich, Merck KGaA, Darmstadt,
the main tunicate clades. Raw sequence data were retrieved Germany), and cloned with TOPO TA Cloning Kit
and assembled using SeqMan 8.0.2 (Pro Assembler) software (Invitrogen, Thermo Fisher Scientific, Waltham, MA).
from the DNASTAR Lasergene package, and manually Plasmid DNA was purified from bacteria using the
inspected in order to reconstruct new MT sequences. The GeneElute Plasmid Miniprep Kit (Sigma-Aldrich), screened
MT nature of each new identified sequence was evaluated by for insert presence by digestion with Fast Digest EcoRI
BlastX searches against metazoan NCBI nonredundant pro- (Invitrogen, Thermo Fisher Scientific, Waltham, MA), and se-
tein sequence database. The amino acid sequences and the quenced at the Scientific and Technological Centers of the
accession numbers of the retrieved MTs are provided in sup- University of Barcelona using the Big Dye Terminator v3.1
plementary table S1, Supplementary Material online. Cycle Sequencing Kit (Applied Biosystems) in an automatic
Sequence alignments were generated with Aliview program sequencer (ABIPRISM 310, Applied Biosystems). Sequences
(Larsson 2014) and reviewed manually. were analyzed with Unipro UGENE software
(Okonechnikov et al. 2012).
Characterization of S. thompsoni Genes
Salpa thompsoni specimens were kindly provided by Nora- Production and Purification of Recombinant Metal–
Charlotte Pauli from the Bettina Meyer’s group at Alfred MT Complexes
Wegener Institute for Polar and Marine Research (AWI). Synthetic cDNAs codifying the selected MTs (i.e., CroMT1,
Genomic DNA was extracted using Quick-DNA/RNA HroMT1, HroMT2, BscMT1, R4-BscMT1, SthMT1, SthMT2,
Microprep Plus Kit Protocol (Zymo Research, CA) using SthMT3, and SthMT4) were provided by Synbio
5 mg of tissue and following the manufacturer’s instructions. Technologies (Monmouth Junction, NJ), cloned in the
The DNA concentration, purity, and integrity were checked pGEX-4T-1 expression vector (GE Healthcare, Chicago, IL),
using Tecan Infinite M200 (Tecan Group Ltd, Switzerland) and transformed in protease-deficient E. coli BL21 strain for
measuring the absorbance at 260 and 280 nm. heterologous expression. For heterologous protein produc-
Based on S. thompsoni genome and transcriptome projects tion, 500 ml of Luria–Bertani (LB) medium with 100 lg/ml
(PRJNA318929 and GFCC00000000.1, respectively) and the ampicillin was inoculated with E. coli BL21 cells transformed
raw data from S. thompsoni SRA projects, several pairs of with the corresponding recombinant plasmid. After over-
primers were designed in order to PCR amplify the 50 and night growth at 37 C/250 rpm, the cultures were used to
30 flanking regions, as well as of intron sequences of the dif- inoculate 5 l of fresh LB-100 lg/ml ampicillin medium. Gene
ferent SthMTs (supplementary table S4, Supplementary expression was induced with 100 lM isopropyl-b-D-thiogalac-
Material online). topyranoside (IPTG) for 3 h. After the first 30 min of induc-
For SthMT1 we used the primer pairs FwMT1/MT5 with tion, cultures were supplemented with ZnCl2 (300 lM),
RvMT1/MT2; for SthMT5, FwMT1/MT5 with RvMT5, and CdCl2 (300 lM), or CuSO4 (500 lM) in order to generate
finally, for SthMT6 we used FwMT2/MT6 with RvMT6. metal–MT complexes. Cells were harvested by centrifugation
For each PCR reaction, 1 ng of genomic DNA was amplified for 5 min at 9,100 g (7,700 rpm), and bacterial pellets were
using selected pairs of primers and the Phusion High-Fidelity suspended in 125 ml of ice-cold phosphate-buffered saline
4445Calatayud et al. . doi:10.1093/molbev/msab184 MBE
(PBS: 1.4 M NaCl, 27 mM KCl, 101 mM Na2HPO4, 18 mM d’Analisi Quımica (SAQ) de la Universitat Autonoma de
KH2PO4, and 0.5% v/v b-mercaptoethanol). Resuspended Barcelona (ICP-AES, ESI-MS) for allocating instrument time.
cells were sonicated (Sonifier Ultrasonic Cell Disruptor) R.A. was supported by BIO2015-67358-C2-1-P, C.C. was sup-
8 min at voltage 6 with pulses of 0.6 s, and then centrifuged ported by BFU2016-80601-P and PID2019-110562GB-I00, and
for 40 min at 17,200 g (12,000 rpm) and 4 C. M.C. and O.P. were supported by BIO2015-67358-C2-2-P, the
Soluble protein extracts containing GST-MT fusion pro- Spanish Ministerio de Ciencia e Innovacion. M.C. and O.P. are
teins were incubated with glutathione sepharose beads (GE members of the “Grup de Recerca de la Generalitat de
Healthcare) for 1 h at room temperature with gentle rotation. Catalunya,” ref. 2017SGR-864, and R.A. and CC. of ref.
GST-MT fusion proteins bound to the sepharose beads were 2017SGR-1665. M.G.-R. acknowledges the UAB the PIF grant.
washed with 30 ml of cold 1xPBS bubbled with argon to
prevent oxidation. After three washes, GST-MT fusion pro-
teins were digested with thrombin (SERVA Electrophoresis Data Availability
GmbH Heidelberg, Germany, 25 U/l of culture) overnight at
Downloaded from https://academic.oup.com/mbe/article/38/10/4435/6306413 by guest on 01 November 2021
17 C, thus enabling separation of the metal–MT complexes The data underlying this article are available in NCBI (https://
from the GST that remained bound to the sepharose matrix. www.ncbi.nlm.nih.gov/) and Octopus (http://octopus.obs-
The eluted metal–MT complexes were concentrated with a vlfr.fr/public/botryllus/blastbotryllus.php) databases. The
3 kDa Centripep Low Concentrator (Amicon, Merck- amino acid sequences and the accession numbers of the
Millipore, Darmstadt, Germany), and fractionated on a MTs used in this work are available in supplementary table
Superdex-75 FPLC column (GE Healthcare) equilibrated S1, Supplementary Material online.
with 20 mM Tris–HCl, pH 7.0. The protein-containing frac-
tions, identified by their absorbance at 254 nm, were pooled References
and stored at 80 C until use. Adamo GM, Lotti M, Tamas MJ, Brocca S. 2012. Amplification of the
CUP1 gene is associated with evolution of copper tolerance in
Analysis of Metal–MT Complexes Saccharomyces cerevisiae. Microbiology 158(Pt 9):2325–2335.
Artells E, Palacios O, Capdevila M, Atrian S. 2013. Mammalian MT1 and
Protein quantification and element composition of all the MT2 metallothioneins differ in their metal binding abilities.
samples were achieved by ICP-AES measurements performed Metallomics 5(10):1397–1410.
in a Optima 4300DV apparatus (Perkin-Elmer, MA) (S, Baumann C, Beil A, Jurt S, Niederwanger M, Palacios O, Capdevila M,
182.040 nm; Zn, 213.856 nm; Cd, 228.802 nm; Cu, Atrian S, Dallinger R, Zerbe O. 2017. Structural adaptation of a pro-
tein to increased metal stress: NMR structure of a marine snail
324.803 nm) under conventional conditions following an al-
metallothionein with an additional domain. Angew Chem Int Ed
ready stablished method (Bongers et al. 1988). Molecular Engl. 56(16):4617–4622.
weights were determined by ESI-MS, in a MicroTof-Q instru- Beil A, Jurt S, Walser R, Schonhut T, Guntert P, Palacios O, Atrian S,
ment (Bruker Daltonics Gmbh, Bremen, Germany) connected Capdevila M, Dallinger R, Zerbe O. 2019. The solution structure and
to a Series 1100 HPLC pump (Agilent Technologies) con- dynamics of Cd-metallothionein from helix pomatia reveal optimi-
zation for binding Cd over Zn. Biochemistry 58(45):4570–4581.
trolled by the Compass Software. The instrument was cali-
Blindauer CA. 2014. Metallothioneins. In: Maret W, Wedd A, editors.
brated with ESI-L Low Concentration Turning Mix (Agilent Binding, transport and storage of metal ions in biological cells.
Technologies, Santa Clara, CA). Metallated forms were Cambridge: The Royal Society of Chemistry. p. 594–653.
detected under native conditions: 20 ml of sample injected Blindauer CA, Leszczyszyn OI. 2010. Metallothioneins: unparalleled di-
through a PEEK tube at 30–50 ml min1 in a 3.5–5.0 kV versity in structures and functions for metal ion homeostasis and
more. Nat Prod Rep. 27(5):720–741.
capillary-counter voltage, at 90–110 C of desolvation tem-
Bofill R, Capdevila M, Atrian S. 2009. Independent metal-binding features
perature, and with dry gas at 6 l min1. The spectra were of recombinant metallothioneins convergently draw a step grada-
recorded between a m/z range from 800 to 3,000. The liquid tion between Zn- and Cu-thioneins. Metallomics 1(3):229–234.
carrier was a 90:10 mixture of 15 mM ammonium acetate and Bongers J, Walton CD, Richardson DE, Bell JU. 1988. Micromolar protein
acetonitrile at pH 7.0. All molecular masses were calculated concentrations and metalloprotein stoichiometries obtained by in-
ductively coupled plasma atomic emission spectrometric determi-
according to the bibliography (Fabris et al. 1996).
nation of sulfur. Anal Chem. 60(24):2683–2686.
Bruland KW. 1980. Oceanographic distributions of cadmium, zinc,
Supplementary Material nickel, and copper in the North Pacific. Earth Planetary Sci Lett.
47(2):176–198.
Supplementary data are available at Molecular Biology and Calatayud S, Garcia-Risco M, Capdevila M, Ca~ nestro C, Palacios O,
Evolution online. Albalat R. Forthcoming 2021. Modular evolution and population
variability of Oikopleura dioica metallothioneins. Front Cell Dev
Acknowledgments Biol. doi: 10.3389/fcell.2021.702688.
Calatayud S, Garcia-Risco M, Pedrini-Martha V, Eernisse DJ, Dallinger R,
The authors thank Nora-Charlotte Pauli and Bettina Meyer at Palacios O, Capdevila M, Albalat R. 2021. Modularity in protein
Alfred Wegener Institute for Polar and Marine Research for evolution: modular organization and de novo domain evolution in
providing Salpa thompsoni specimens, and all tunicate labo- mollusk metallothioneins. Mol Biol Evol. 38(2):424–436.
ratories for sharing sequence results ahead of their publica- Calatayud S, Garcia-Risco M, Rojas NS, Espinosa-Sanchez L, Artime S,
Palacios O, Ca~ nestro C, Albalat R. 2018. Metallothioneins of the
tion. They thank Sebastian Artime for experimental support. urochordate Oikopleura dioica have Cys-rich tandem repeats, large
They also thank the Centres Cientıfics i Tecnologics (CCiT) de size and cadmium-binding preference. Metallomics
la Universitat de Barcelona (DNA sequencing) and the Servei 10(11):1585–1594.
4446You can also read

















































