A Comparison of Leaf and Petal Senescence in Wallflower Reveals Common and Distinct Patterns of Gene Expression and Physiology1 W
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

A Comparison of Leaf and Petal Senescence in Wallflower
Reveals Common and Distinct Patterns of Gene
Expression and Physiology1[W]
Anna Marie Price2, Danilo F. Aros Orellana, Faezah Mohd Salleh, Ryan Stevens, Rosemary Acock,
Vicky Buchanan-Wollaston, Anthony D. Stead, and Hilary J. Rogers*
Cardiff School of Biosciences, Cardiff CF10 3TL, United Kingdom (A.M.P., D.F.A.O., F.M.S., R.S., R.A., H.J.R.);
Warwick HRI, University of Warwick, Wellesbourne, Warwick, Warwickshire CV35 9EF, United Kingdom
(V.B.-W.); and School of Biological Sciences, Royal Holloway, University of London Egham, Surrey TW20 0EX,
United Kingdom (A.D.S.)
Petals and leaves share common evolutionary origins but perform very different functions. However, few studies have
compared leaf and petal senescence within the same species. Wallflower (Erysimum linifolium), an ornamental species closely
related to Arabidopsis (Arabidopsis thaliana), provide a good species in which to study these processes. Physiological
parameters were used to define stages of development and senescence in leaves and petals and to align these stages in the two
organs. Treatment with silver thiosulfate confirmed that petal senescence in wallflower is ethylene dependent, and treatment
with exogenous cytokinin and 6-methyl purine, an inhibitor of cytokinin oxidase, suggests a role for cytokinins in this process.
Subtractive libraries were created, enriched for wallflower genes whose expression is up-regulated during leaf or petal
senescence, and used to create a microarray, together with 91 senescence-related Arabidopsis probes. Several microarray
hybridization classes were observed demonstrating similarities and differences in gene expression profiles of these two organs.
Putative functions were ascribed to 170 sequenced DNA fragments from the libraries. Notable similarities between leaf and
petal senescence include a large proportion of remobilization-related genes, such as the cysteine protease gene SENESCENCE-
ASSOCIATED GENE12 that was up-regulated in both tissues with age. Interesting differences included the up-regulation of
chitinase and glutathione S-transferase genes in senescing petals while their expression remained constant or fell with age in
leaves. Semiquantitative reverse transcription-polymerase chain reaction of selected genes from the suppression subtractive
hybridization libraries revealed more complex patterns of expression compared with the array data.
Both leaves and flowers have a finite life span, and both these tissues during senescence. These include
since it is thought that all floral organs, including genes encoding proteases, nucleases, and enzymes
petals, evolved from leaves (Friedman et al., 2004), we involved in lipid and carbohydrate metabolism
might expect commonality in their senescence mech- (Buchanan-Wollaston, 1997; Wagstaff et al., 2002). In
anisms. Both in leaves and petals, a key feature of both organs, remobilization requires a carefully or-
senescence is remobilization of resources; in both or- chestrated dismantling of the cellular machinery to
gans, this has been demonstrated experimentally using avoid cell death until remobilization is complete. In
isotope labeling (Nichols and Ho, 1975; Mae et al., 1985; leaves, senescence-associated genes (SAGs) have been
Bieleski, 1995) or pigment transport (Erdelská and classified into two expression types: those exclusively
Ovečka, 2004). This is reflected in some of the major expressed during senescence (class I) and those whose
classes of genes whose expression is up-regulated in expression increases during senescence from a basal
level (class II; Gan and Amasino, 1997). However,
1
This work was supported by grants from the Biotechnology and within these classes, there are diverse expression
Biological Sciences Research Council (to A.M.P.), the Chilean Gov- patterns (Smart, 1994; Buchanan-Wollaston, 1997),
ernment, Ministry of Agriculture (to D.F.A.O.), and the Malaysian indicating different regulatory pathways. Levels of
Government (to F.M.S.). reactive oxygen species (ROS) rise in both petals and
2
Present address: Centre for Molecular Oncology, Institute of leaves during senescence (Borochov and Woodson,
Cancer, Barts, and the London School of Medicine and Dentistry, 1989; Merzlyak and Hendry, 1994), maybe as a result of
John Vane Science Centre, Charterhouse Square, London EC1M macromolecule degradation. This is accompanied by
6BQ, UK. up-regulation of genes involved in protection against
* Corresponding author; e-mail rogershj@cf.ac.uk.
ROS, such as catalase in leaves (Buchanan-Wollaston and
The author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy Ainsworth, 1997; Zimmermann et al., 2006) and su-
described in the Instructions for Authors (www.plantphysiol.org) is: peroxide dismutase in petals (Panavas and Rubinstein,
Hilary J. Rogers (rogershj@cf.ac.uk). 1998).
[W]
The online version of this article contains Web-only data. The roles of petals and leaves are very different, as
www.plantphysiol.org/cgi/doi/10.1104/pp.108.120402 are their development and the signaling mechanisms
1898 Plant Physiology, August 2008, Vol. 147, pp. 1898–1912, www.plantphysiol.org Ó 2008 American Society of Plant Biologists
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.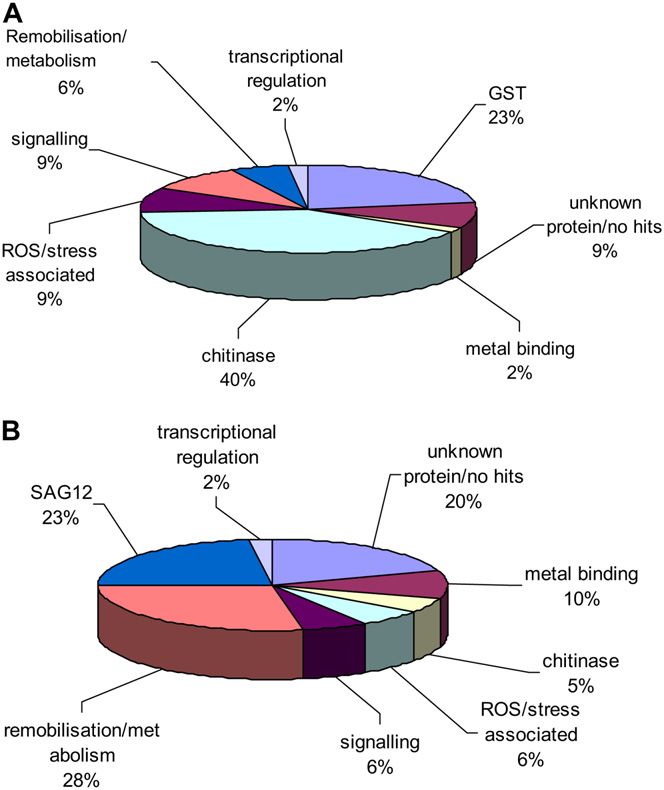
A Comparison of Leaf and Petal Senescence in Wallflower
that trigger their senescence. An early step in petal are poorly understood. Transcriptional regulation of
development is the conversion of chloroplasts to chro- senescence in both leaves and petals is also complex
moplasts (Thomson and Whatley, 1980), and this has and as yet not fully understood. Transcription factors
been compared with the transformation of chloro- that are up-regulated during leaf senescence, such as
plasts into gerontoplasts that occurs during leaf se- WRKY53 (Hinderhofer and Zentgraf, 2001) and many
nescence (Thomas et al., 2003), implying similarities others, have been identified (Buchanan Wollaston
between developing (nonsenescent) petals and senes- et al., 2005), but as yet their interactions have not
cent leaves. This would suggest that senescence- been fully elucidated. Similarly, transcription factors
associated events in petals may occur at an earlier up-regulated during petal senescence have been iden-
stage compared with leaves and that cellular degra- tified in several genera (Alstroemeria [Breeze et al.,
dation accompanied by the expression of some genes 2004] and Iris [van Doorn et al., 2003]) but not fully
that are highly up-regulated in senescent petals is characterized.
already evident while the petals are in the early stages Global transcriptomic and EST analyses have probed
of development (Wagstaff et al., 2002, 2003). The senescence independently in leaves in Arabidopsis
primary function of petals is to attract pollinators, so (Gepstein et al., 2003; Buchanan-Wollaston et al., 2005)
they are frequently highly pigmented and scented and and petals (in Alstroemeria [Breeze et al., 2004], Iris [van
a sink rather than a source of photosynthates. Floral Doorn et al., 2003], and Rosa [Channeliere et al., 2002]);
life span is closely linked to pollination in some however, to date there is a lack of comparisons of leaf
species, which triggers rapid floral deterioration and petal senescence transcriptomes in the same spe-
(Stead and van Doorn, 1994). However, even in the cies. Wallflower (Erysimum linifolium) is a useful orna-
absence of pollination, floral life-span is finite. Al- mental species in which to compare leaf and petal
though a few environmental factors such as tempera- senescence. It is closely related taxonomically to Arab-
ture and drought can affect floral longevity, senescence idopsis (Stevens, 2001) but has larger pigmented flow-
is irreversible in the majority of species and there is ers whose development and senescence are easily
tight species-specific control over the maximum dura- staged. Thus, in the study presented here, the objec-
tion of a flower (Primack, 1985). In contrast, leaves are tives were (1) to use microarray analysis of subtractive
sources of photosynthate for most of their life span, libraries from wallflower leaves and petals to compare
and their longevity is strongly influenced by nutrient the global gene expression changes occurring during
status, light, and other environmental factors. Fertili- senescence in these two tissues and relate these to
zation does accelerate leaf senescence in some species changes in the physiology of the two organs during
(Hayati et al., 1995) but not in others, such as Arabi- senescence and (2) to take advantage of the close
dopsis (Arabidopsis thaliana; Hensel et al., 1993). How- taxonomic relationship between wallflower and Arab-
ever, as in petals, expression of some genes associated idopsis to compare and contrast expression patterns
with leaf senescence is also detected before visible between the two species in the two tissues and reveal
signs of deterioration (Buchanan-Wollaston, 1997), in- species-specific or tissue-specific differences in the
dicating that in both petals and leaves senescence senescence program.
processes may be initiated early.
Two classes of plant growth regulators, ethylene and
cytokinins, are definitely involved in both petal and
leaf senescence in some species. The sensitivity of petal RESULTS
senescence to endogenously produced, or exoge- Physiology of Leaf and Petal Senescence in Wallflower
nously applied, ethylene is species specific, and spe-
cies can be broadly divided into those in which petal One flower on the wallflower raceme opened each
senescence is ethylene sensitive and those in which it day, taking 7 d to complete its development from bud
is not (Rogers, 2006). In carnation (Dianthus caryophyl- opening to full abscission of the calyx, corolla, and
lus), an ethylene-sensitive species, ethylene produc- androecium (Fig. 1). Thus, eight stages of development
tion and ethylene biosynthetic genes are both were assigned based on number of days after opening.
up-regulated in petals late during the vase life of the Stage 0 was defined as the lowest unopened bud;
flower (ten Have and Woltering 1997). In leaves, as in additional early bud stages were designated stages
flowers, ethylene sensitivity is related to the age of the 21 and 22. No difference in morphology or in rate of
organ; however, in general, the role of ethylene in leaf development was noted for the flowers at different
senescence is less central (Grbić and Bleecker, 1995). times of year. Stage 4 was the stage at which the first
Up-regulation of cytokinins delays both leaf (Gan and signs of visible petal deterioration became evident.
Amasino, 1995) and petal (Chang et al., 2003) senes- Leaves could be characterized within one whorl and
cence, and it has been suggested that a fall in cytoki- were assigned to seven developmental groups based
nins may trigger an increase in ethylene sensitivity on relative size and chlorophyll content (Fig. 2). At
during petal senescence in unpollinated ethylene- stage 5, leaves showed the first signs of yellowing,
sensitive species (van Doorn and Woltering, 2008). indicating senescence, and this corresponded with a
Other plant growth regulators are probably also in- 20% reduction in chlorophyll levels. Dry weight-fresh
volved, but the signaling pathways and their cross talk weight ratio and total protein content were also deter-
Plant Physiol. Vol. 147, 2008 1899
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Price et al.
Figure 1. Stages of wallflower flower development. Stage 22 and stage 21, Two sequential buds below the lowest unopened
bud on the raceme. Sepals completely cover petals. Stage 0, Lowest unopened bud on raceme. Petals are dark purple in color,
tightly curled within sepals. Stage 1, Flower fully opened. Petals are pale purple, with sepals folded back midway along their
length. Stigma is yellow and fuzzy in appearance, four of six anthers are visible, all undehisced, positioned close to the stigma
with the tips curled over the stigma. Stage 2, As stage 1, but petals are darker in color. All six anthers are visible, two newly
emerged anthers are dehisced and curled back from the stigma. Stage 3, The flower is not as tightly held together as previously.
Petals are wilting slightly and darker again in color. Fuzz on stigma is not as fine as previously. All six anthers are dehisced and
curled back from the stigma. Stage 4, The flower is loosely held together. Petals are limp and curled over at the tips. Flower
appearance has deteriorated. Stage 5, As stage 4, but more extreme. Petals are wilted, stigma is discolored with dark purple areas.
Stage 6, Sepals, petals, and stamens are beginning to abscise. Remaining petals look withered and dry. Stage 7, All sepals, petals,
and stamens are abscised; only the stigma remains. Bar 5 10 mm.
mined for each developmental stage of petals and cence (Serek et al., 1995). In wallflower, detached
leaves (Figs. 3 and 4). There was a sharp reduction in flowers harvested at stage 1 and held in water se-
dry weight-fresh weight ratio between petal stages 0 nesced over the same period as attached flowers, with
and 1, coinciding with flower opening, followed by a full abscission on day 7. However, when pulsed for 1 h
rise starting from stage 4 as petals lost turgor. Protein with STS on the day of harvest, abscission was delayed
loss started after stage 3, coincident with the first signs by 2 d. STS-pulsed flowers also senesced more slowly,
of petal deterioration. In leaves, the dry weight-fresh taking 4 d to progress from stage 3 to stage 5, instead of
weight ratio started to rise after stage 5, while protein 2 d when held in water. Given that in ethylene-
loss started after stage 4, again preceding the start of sensitive species, such as carnation, cytokinins are
visual signs of senescence. also implicated in petal senescence (Taverner et al.,
Due to the close taxonomic relationship between 2000), the role of cytokinins in wallflower was tested.
wallflower and Arabidopsis, it seemed likely that Treatment with either 0.1 or 1.0 mM kinetin or with
ethylene would be an important regulator of petal 0.1 mM 6-methyl purine (an inhibitor of cytokinin
senescence in this species too. In ethylene-sensitive oxidase) delayed senescence and abscission of flow-
species, treatment with a pulse of an ethylene inhibitor ers harvested at stage 1 by 2 d (Supplemental Figs. S1
such as silver thiosulfate (STS) delays flower senes- and S2).
Figure 2. Stages of wallflower leaf development. Stage 1, Very young leaves, less than 50% expanded. Stage 2, Very young
leaves, 50% to 75% expanded. Stage 3, Young leaves, 75% to 100% expanded. Stage 4, Mature green leaves. Stage 5, Older
mature leaves, green with signs of yellowing on the tip. Stage 6, Old leaves, up to 50% of leaf area yellow. Stage 7, Very old
leaves, mostly or all yellow. Below each image is the total chlorophyll for that leaf stage expressed as a percentage of maximum.
1900 Plant Physiol. Vol. 147, 2008
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.A Comparison of Leaf and Petal Senescence in Wallflower
Six of the possible nine classes of expression (i.e. up-
regulated in both old petals and old leaves compared
with the young tissue, up-regulated in petals but
unchanging in leaves, up-regulated in petals but
down-regulated in leaves, etc.) were represented in
the microarrays (Table I). Of the 427 probes (excluding
the replicates described above), expression of 305
probes was up-regulated reproducibly in old petals
compared with young petals. Of these, the expression
of 232 probes was up-regulated in both old organs,
while the expression of 61 probes was up-regulated in
old petals but remained stable in leaves, and the
expression of a further 12 probes was up-regulated
in petals with age but was down-regulated in old
leaves. As expected from the enrichment of the genes
by suppression subtractive hybridization (SSH), the
majority of probes on the array indicated up-regulated
expression with senescence in the tissue from which
they were derived, confirming that the subtraction of
the SSH libraries was effective. Of 164 probes from the
petal cDNA library, whose expression could be reli-
ably determined in both tissues, the expression of 98%
Figure 3. Fresh weight (FW), dry weight (DW), and ratio of dry weight
to fresh weight during petal (A) and leaf (B) development and senes-
was up-regulated with age in petals. For 263 probes
cence. Dry weight was determined by drying 20 to 100 petals or leaves derived from the leaf cDNA library, 52% showed up-
at 60°C for 5 d. Error bars represent 6 SE (n 5 3). regulated expression with age in leaves, although
larger numbers of leaf-derived probes on the array
Construction of Wallflower Petal and Leaf Subtracted were stable in expression with leaf senescence (47%;
Libraries and Screening by Microarray Analysis Supplemental Table S1).
Based on the physiological characterization of leaf
Sequence Analysis of Wallflower Genes from the
and petal senescence, subtracted libraries were con-
SSH Libraries
structed for use in transcriptomic analysis to identify
genes whose expression is up-regulated during the Following microarray analysis, fragments represent-
senescence of these two organs. For this purpose, ing selected probes on the array were chosen for
petals from stages 22, 21, and 0 (early to mature
buds) were combined to represent young petals, and
petals from stages 3, 4, and 5 (early to late visible signs
of petal wilting) were combined to represent old
petals. Leaf stage 3 (75%–100% expansion, 80% chlo-
rophyll) was used to represent young leaves that had
not yet reached their full photosynthetic capability,
and stages 5 and 6 (early to later stages of leaf yellow-
ing, in which chlorophyll levels had fallen to 81% and
44% of maximum, respectively) were combined to
represent old leaves. A total of 1,018 and 614 clones for
leaves and petals, respectively, were obtained from the
subtraction. PCR-amplified inserts from all 1,632
clones from the subtracted libraries were used to
generate a cDNA microarray, and 431 probes showed
a consistent expression pattern with both pairs of
labeled RNA when analyzed using GeneSpring soft-
ware. The results from the microarray analysis are
summarized in Supplemental Table S1. Two fragments
representing known genes WLS63 and WPC11A were
spotted in three replicate dilutions (36 data points) and
showed very similar changes in expression with low
variability between replicates (for WLS63, leaves down,
1.1 6 0.2 [values are mean fold 6 SE], petals up, 3.8 6
0.4; for WPC11A, leaves up, 10 6 1.0, petals up, 136 6 Figure 4. Total protein content of petals (A) and leaves (B). Error bars
27), indicating the reproducibility of the array results. represent 6 SE (n 5 15).
Plant Physiol. Vol. 147, 2008 1901
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Price et al.
Table I. Expression classes from microarray analysis sequences (40%) were related to chitinases. The ma-
jority of these chitinase-related sequences (20) were
Petal Petal Petal
Unchanged Down-Regulated Up-Regulated
most closely related to an Arabidopsis class IV chiti-
nase (At2g43570; contigs WC1 and WC2), while one
Leaf unchanged 103 0 61 was more closely related to an Arabidopsis family 18
Leaf down-regulated 1 0 12
glycosyl hydrolase (At4g19810; contig WC3); both Arab-
Leaf up-regulated 18 0 232
idopsis genes are putatively involved in cell wall metab-
olism.
sequencing to represent the different classes of gene A further 23% of the sequences from this array
expression presented in Table I. In addition, a random expression class (up-regulated in old petals, either
selection of clones from the SSH libraries were also unchanged or down-regulated in senescent leaves)
sequenced. Once poor and short sequences had been showed homology to GSTs. All of the 10 sequenced
removed, 210 ESTs were obtained (GenBank accession probes that were up-regulated in senescent petals but
numbers are listed in Supplemental Table S1) and 127 down-regulated in senescent leaves showed closest
of the sequences clustered into 27 contigs (WC1– homology to the f class of GSTs (Wagner et al., 2002),
WC27), with the largest contig containing 28 se- and nine were assigned to one contig (WC10). All the
quences (Table II). The remaining 83 sequences were WC10 sequences were closest to AtGSTF3, while the
singletons (i.e. represented only once). Thus, the re- singleton sequence was closer to AtGSTF7. However,
dundancy (number of sequences clustered divided by as all of the clones were partial, it is difficult to assign
the total number of sequences; Breeze et al., 2004) of the sequences unambiguously to an Arabidopsis ho-
the EST collection was 60% (which means that the molog, as a key diagnostic triplet of amino acids at
chance of finding the same sequence again in any new positions 66 to 68 relative to AtGSTF2 (Wagner et al.,
clones sequenced is 60%). However, there may be 2002) was not included in the wallflower clones and, in
further redundancy due to nonoverlapping fragments addition, AtGSTF3 and AtGSTF2 were 95% identical.
of the same gene. Genes within contigs were given All of the GST-related probes in this expression class
codes: WLC (Wallflower Leaf Contig) and WPC (Wall- showed similar expression patterns on the microarray
flower Petal Contig), and singletons were denoted (leaf, 0.35 6 0.02 [values are mean fold 6 SE]; petal,
WLS and WPS. Contigs are hereafter referred to in the 15.3 6 1.69). The expression of two further probes on
form WC1 (Supplemental Table S1). Putative gene the microarray whose sequence showed homology to
functions were assigned based on a BLAST search, and GSTs was up-regulated in senescent petals but was
in most cases the closest match was to Arabidopsis genes, stable in senescent leaves (leaf, 1.5 6 0.23; petal, 4.3 6
as wallflower is of the same subfamily (Brassicoideae; 0.09). These sequences formed a separate contig
Stevens, 2001). In total, 193 wallflower sequences (WC21) showing closest homology to AtGSTZ1. The
could be assigned to a closely matching Arabidopsis remainder of the sequenced probes on the microarray,
gene, and 73 Arabidopsis genes were identified as the for which putative functions could be ascribed and
closest match. Analysis of gene functions revealed that whose expression was up-regulated in senescent
three contigs (WC4, WC5, and WC26, comprising petals but stable in senescent leaves, represented
altogether 26 sequences) and four further sequences metal-binding proteins (one probe), proteins associ-
that did not overlap the contigs, amounting to 14% of ated with ROS/stress (five probes) or signaling (five
the sequences, matched SAG12. Three contigs (WC1, probes), proteins involved in remobilization/metabo-
WC2, and WC16, comprising 25 sequences) matched lism (three probes), and one gene involved in mRNA
nonoverlapping regions of the same chitinase gene stability. The metal-binding protein was a putative
(At2g43570), and a fourth contig (WC3 of three sequences) copper chaperone most closely homologous to CCH/
matched most closely a different chitinase gene ATX1 that is thought to play a role in remobilization of
(At4g19810). Thus, 13% of the sequences represented copper from metalloprotein degradation (Himelblau
chitinase-like genes. A further 7% of the sequences and Amasino, 2000) and was 4-fold up-regulated in
matched glutathione S-transferases (GSTs), 7% matched petals. ROS/stress-related proteins include a PR5-like
metallothioneins, and 4% matched a lipid transfer protein (petals, up-regulated by 6.3-fold), a cytosolic
protein. thioredoxin (petals, up-regulated by 3.0-fold), SAG21
(petals, up-regulated by 5.7-fold), and a cytochrome
Representation of the Functional Categories in the P450 family protein (petals, up-regulated by 23.3-fold).
Different Expression Classes Signaling proteins include a rhodopsin-like receptor
(petals, up-regulated by 2.9-fold), a Rab acceptor
There was a striking difference in the representation (petals, up-regulated by 7.5-fold), and a Rab subfamily
of putative functional categories between the different GTPase (petals, up-regulated by 2.3-fold). Finding
gene expression classes on the microarray (Fig. 5). genes encoding proteins involved in remobilization
Sequences were obtained for 75% of the probes on the is not surprising, although genes whose role may be
microarray whose expression was up-regulated in specific to remobilization in petals and not leaves may
senescent petals and was either unchanged or down- be significant in defining the difference between re-
regulated in senescent leaves. Over one-third of these mobilization in the two organs. The three up-regulated
1902 Plant Physiol. Vol. 147, 2008
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.A Comparison of Leaf and Petal Senescence in Wallflower
Table II. Most abundant sequenced transcripts from the SSH libraries
Arabidopsis
Genome Initiative Wallflower No. of Putative Function/Closest Functional
Code for Closest Contig Clones Arabidopsis Homolog Class
Arabidopsis Match
At2g43570 WC1 28 Chitinase class IV Stress/defense
At5g45890 WC4 26 SAG12 Remobilization
At1g07600 WC24 13 Metallothionein Metal binding
At2g02930 WC10 10 ATGSTF3 Defense
At3g22600 WC6 9 Lipid transfer protein Remobilization
At2g43570 WC2 9 Chitinase class IV Stress/defense
At1g32450 WC11 5 Peptide transporter Remobilization
At3g22600 WC7 5 Lipid transfer protein Remobilization
At5g01600 WC17 4 Ferretin Metal binding
At2g23790 WC8 3 Unknown protein Unknown
At2g45570 WC14 3 Cytochrome P450 ROS/stress
At1g73260 WC15 3 Endopeptidase inhibitor Remobilization
– WC18 3 No hits –
At4g19810 WC3 2 Chitinase Stress/defense
At1g11190 WC9 2 Bifunctional nuclease Remobilization
At4g02520 WC10 2 ATGSTF2 Defense
At5g02040 WC13 2 Rab acceptor Signaling
At5g01220 WC19 2 Unknown protein Unknown
At1g05560 WC20 2 UDP glycosyl transferase ROS/stress
At2g02390 WC21 2 ATGSTZ1 Defense
At5g40690 WC22 2 Unknown protein Unknown
At2g45220 WC23 2 Pectin esterase inhibitor Remobilization
At5g45890 WC5 2 SAG12 Remobilization
– WC12 2 No hits –
At2g43570 WC16 2 Chitinase class IV Stress/defense
At1g07600 WC25 2 Metallothionein Metal binding
At5g45890 WC26 2 SAG12 Remobilization
genes identified here were a lipid transfer protein petals than in leaves. Three sequences were homolo-
(leaves, 1.59-fold; petals, 5.14-fold), a thiol protease gous to transcriptional regulators, and the expression
(leaves, 1.41-fold; petals, 2.77-fold), and a AAA-type of these genes was also more highly up-regulated in
ATPase family protein (leaves, 1.51-fold; petals, 4.95- aging petals compared with leaves: a WRKY75 tran-
fold). Only one gene involved in the regulation of gene scription factor (At5g13080) and two members of the
expression and up-regulated only in petal senescence plant-specific NAC family of transcription factors
was identified, and it showed homology to CCR4- (At2g33480 and At5g64530).
related proteins (WLS63, three replicates on the array; The expression of only a few probes (18) was up-
leaf, 0.88 6 0.15; petal, 3.8 6 0.43). CCR4-NOT proteins regulated in senescent leaves while remaining un-
in yeast are involved in the regulation of gene expres- changed in petals. Sequences were obtained for seven
sion via mRNA stability (Chen et al., 2002). of these: four were putative ferretin genes (leaf, 3.6 6
In contrast, of those probes that were up-regulated 1.7), while the rest were of unknown function (Sup-
in both senescent leaves and petals and for which plemental Table S1).
meaningful sequence was obtained, the highest pro-
portion (23%) was represented by SAG12, while chiti- Semiquantitative Reverse Transcription-PCR of
nase genes represented only 5% and no GST genes Selected Wallflower Genes
were up-regulated in both tissues (Fig. 5). The expres-
sion of all of the SAG12 probes was reliably deter- Genes were selected for semiquantitative reverse
mined from the microarray, and all were up-regulated transcription (RT)-PCR based on their putative func-
in both leaves and petals, although more strongly in tion and results from the microarray experiments, to
petals (leaves, 95 6 17; petals, 216 6 35). A lower confirm the validity of the arrays and also to determine
proportion of the sequences in this expression class, more precise timing of expression for selected genes of
compared with those that were only up-regulated in interest. SAG12 was selected as it represented a high pro-
petals with age, related to signaling and included three portion of probes whose expression was up-regulated
genes with putative functions in auxin responses, one in both old leaves and petals (Supplemental Table S1).
in cytokinin responses, and one in ethylene synthesis. Semiquantitative RT-PCR (Fig. 6) showed that the
The expression of all of these genes with putative roles expression of SAG12 remained low in leaves until
in signaling was more highly up-regulated with age in stage 6, at which point chlorophyll levels were re-
Plant Physiol. Vol. 147, 2008 1903
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Price et al.
complex pattern than was evident from the arrays in
which pooled tissue stages were used (Fig. 6). Thus,
although WLS63 expression was low in stage 3 leaves,
selected to represent young tissue, and remained low
as leaves aged, the highest levels of expression were
early in leaf development at stage 1. In petals, expres-
sion reached a maximum before the final stages of
petal senescence, at stage 4. WC11 expression fluctu-
ated during leaf age, being already high in young
leaves and reaching a peak at stage 4, when fresh
weight, dry weight, protein levels, and chlorophyll
levels were maximal, falling thereafter. Its expression
in petals was very low in young buds (stages 22 and
21) but increased already to 60% of maximum from
stage 0, when the buds were not yet open. It reached a
maximum expression level at stage 1 (young open
flowers) and remained high until it dropped slightly in
late senescence (stage 5).
Use of Arabidopsis Gene Probes in Cross-Species
Microarray Analysis and Comparisons with
Arabidopsis Gene Chip Data
In addition to the wallflower probes, 91 Arabidopsis
probes were also printed onto the arrays. Many of
these Arabidopsis sequences were selected as genes
whose expression was already known to change with
leaf senescence in Arabidopsis. Expression patterns of
Figure 5. Putative functional classes of wallflower genes from differ- 52 of these genes in wallflower petals and leaves were
ent microarray expression classes. Comparison of putative functional
reliably detected on the arrays for both tissues. Gene
classes of genes represented in two expression categories from the
microarray analysis. A, Up-regulated in senescent petal but either
expression patterns in Arabidopsis mature green
stably expressed or down-regulated in senescent leaf. B, Up-regulated leaves (MG, analogous to wallflower stage 4 leaves)
in both senescent petal and senescent leaf. Functional classes were and two stages of leaf senescence (S1, between stage 4
derived from Gene Ontogeny annotations and from putative functions and stage 5, and S2, between stage 5 and stage 6 of wall-
based on sequence homology.
duced to 44% of maximum and protein levels to 35% of
maximum. Expression then increased significantly in
stage 6 leaves, reaching a maximum at the oldest stage
used in the RT-PCR, stage 7. At this stage, both the
protein and chlorophyll levels had decreased to less
than one-quarter of their maximum. In petals, how-
ever, although SAG12 expression was very low in buds
and young open flowers, it was already substantially
up-regulated in mature, stage 2 flowers, at which time
protein levels, fresh weight, and dry weight were at or
close to their maximum. Thereafter, SAG12 levels in
petals fell until by stage 5 they were less than 20% of
the maximum value.
Two additional genes were selected: first, the CCR4-
like protein (WLS63), and second, a gene with a
putative role in remobilization, a peptide transporter
(WC11). On the array, expression of WLS63 was up-
regulated only in petals with age, while the expression
of WC11 was up-regulated in both, although to a much
greater extent in petals. In both cases, the expression Figure 6. RT-PCR of selected genes from the SSH libraries. Semiquan-
pattern from semiquantitative RT-PCR was consistent titative RT-PCR over petal (A) and leaf (B), young (Y) and old (O) stages
with the array data, but a better resolution was as defined in the text, expressed as percentage of maximum value 6 SE
obtained from the RT-PCR due to the larger number (n $ 3) for SAG12, WLS63, and WC11. Note that data for WLS63 and
of separate tissue stages used. This revealed a more WC11 expression levels for stage 7 leaves were not determined.
1904 Plant Physiol. Vol. 147, 2008
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.A Comparison of Leaf and Petal Senescence in Wallflower
flower leaf senescence) of 10 of these genes were verified sequenced wallflower genes on the wallflower array
by northern analysis (Fig. 7), showing a range of expres- (Supplemental Table S2). Of these, six genes were
sion patterns. Data on the expression of all of the Arabi- included in the 52 Arabidopsis genes discussed above.
dopsis genes was also obtained from AtGenExpress Thus, data were available from all three combinations,
(Supplemental Table S2). Leaf and petal stages were enabling a three-way comparison of the expression of
chosen to resemble most closely the stages used for the the Arabidopsis gene and wallflower gene when
wallflower SSH and arrays. All four data sets for probed with the wallflower transcripts and the Arabi-
young/old leaves and petals were obtained from the dopsis gene expression pattern on the Arabidopsis
Weigel laboratory experiments (Schmid et al., 2005). Affymetrix arrays (the At codes for these genes are
Senescing leaves (from 6-week-old Arabidopsis plants), shown in boldface in Supplemental Table S2). For
corresponding to stage 5 to 6 wallflower leaves, were three genes (SAG12, up-regulated in both petals and
compared with leaf 8 (from 4-week-old Arabidopsis leaves; ferretin, up-regulated only in leaf; SAG21, up-
plants), which is not fully expanded and thus resem- regulated only in petals) there was exact correspon-
bles wallflower leaf stage 3. For petals, petal stage 15 dence; for another two genes (a cytochrome P450 and a
(Smyth et al., 1990), which equates to stage 3 wall- copper homeostasis factor gene) the expression pat-
flower petals, was compared with stage 12 petals tern was in broad agreement, although with the wall-
(unopened bud, nondehisced), comparable to stage 0 flower probe the leaf signal was below the threshold
in wallflower. Comparing the northern expression for an up-regulated response; finally, a xylosidase gene
data with the Weigel laboratory array data indicated (At5g49360) was up-regulated only in petals on the
that the senescing leaf material used in the arrays Affymetrix array while it was stable in leaves. This
included leaves at the same stage as the S2 of the result, however, contrasts with the northern analysis for
northern blots, since in Arabidopsis, SAG12 expres- this gene (Fig. 7), which showed a clear up-regulation
sion was only detected late in senescence. Expression of expression in the later stages of leaf senescence. The
patterns of nine of the 10 genes for which northern data from the Arabidopsis gene on the wallflower
data are presented here were detected on the Affyme- array hybridized to wallflower transcripts, and for the
trix Weigel arrays, and eight of them showed increased wallflower homolog WLS27, were in better agreement
gene expression in senescent leaves by both methods. with the northern data, showing up-regulation of
In the case of LSC141 (At5g49360), Affymetrix array expression with age in both tissues.
expression was strong in both leaf stages (means, stage Of the 61 sequenced wallflower probes that matched
8, 513; senescing, 799); however, the increase in ex- Arabidopsis genes and for which Affymetrix expres-
pression (leaf, 1.6-fold) was below a 2-fold threshold. sion data were available for senescent leaves and
For 47 Arabidopsis genes, data were available from petals from the Weigel data, 85% shared the same
both the Affymetrix Arabidopsis arrays and the wall- expression pattern with their Arabidopsis homolog in
flower arrays. Of these, 81% (38 genes) showed the at least one of the two tissues and 53% shared the same
same pattern of expression in at least one tissue in both expression pattern in both tissues. However, there
species and 38% (18 genes) showed the same pattern in were some notable differences in those genes that were
both tissues in both species. The expression of five particularly abundant in the wallflower array or that
genes, 1-aminocyclopropane-1-carboxylic acid (ACC) are of interest because of potential roles in signaling or
oxidase, catalase, blue copper-binding protein, SAG21, regulation (Supplemental Table S2). Thus, expression
and an unknown protein that is strongly induced by of the major class of chitinase genes (WC1/2/16 in
brassinolide (At2g38640), was unchanged in leaves Supplemental Table S2, which is the mean of contigs
but up-regulated in senescent petals of both species. WC1, WC2, and WC16) on the wallflower array was
Ferretin was up-regulated in senescing leaf but not strongly up-regulated with age in wallflower petals
petal tissue of both species, while six genes, histone (mean, 36-fold) but not in leaves. However, in Arabi-
H1-3, a hydrolase, a Cys protease, an RNase, SAG12, dopsis, expression of the homolog (At2g43570) on the
and xyloglucan transferase, were up-regulated in both Affymetrix arrays was strongly up-regulated in both
petals and leaves of both species. The remainder were tissues (leaves, 10.1-fold; petals, 4.8-fold). Expression
unchanged with age in both tissues of both species. of the largest group of wallflower GST sequences
Ten genes were up-regulated in both petals and leaves (WC10, mean of contig WC10 on Supplemental Table
with age in Arabidopsis but only in petals in wallflower. S2) homologous to Arabidopsis AtGSTF3 (At2g02930)
These were cytochrome P450, copper homeostasis was strongly up-regulated in senescent wallflower
factor, POP dikinase, NADPH-dependent oxidore- petals (16.5-fold) but down-regulated in senescent wall-
ductase, Gln synthetase, two Cys proteases, alcohol flower leaves. Expression of Arabidopsis AtGSTF3 on
dehydrogenases, ERD1, and an unknown protein. the Affymetrix arrays showed a similar pattern, with
Conversely, xylosidase and b-glucosidase were up- up-regulation in petals with age (2.7-fold) but no
regulated in both wallflower tissues with age but only change in leaves. However, two of the wallflower
in one of the two tissues in Arabidopsis. sequences (WC21, mean of contig WC21 in Supple-
Affymetrix data, from the Weigel data sets (Schmid mental Table S2) showed closest homology to AtGSTZ1
et al., 2005) described above, were available for 61 of (At2g02390). Expression of these wallflower probes
the 73 genes identified as the closest hits to the was up-regulated strongly in senescent wallflower
Plant Physiol. Vol. 147, 2008 1905
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Price et al.
Figure 7. Northern analysis of 10 Arabidopsis genes
represented on the microarrays. MG represents ma-
ture Arabidopsis green leaf with maximum chlo-
rophyll levels (100%); S1 and S2 are stages of
Arabidopsis leaf senescence with 98% and 60% chlo-
rophyll levels, respectively (Buchanan-Wollaston and
Ainsworth, 1997).
petals (4.3-fold) but was only very mildly up-regulated (7.5-fold) but only very weakly in old wallflower leaves
in leaves (1.5-fold). Expression of Arabidopsis AtGSTZ1 (1.6-fold). In contrast, expression of the closest Arabi-
on the Affymetrix arrays was strongly up-regulated in dopsis homolog, At5g02040, was up-regulated in old
both leaves (4.9-fold) and petals (6-fold). leaves (2.3-fold) but was stable with age in petals on
Four genes with potential roles in signaling differed the Affymetrix arrays. Expression of the second Rab-
in expression patterns between Arabidopsis and wall- related wallflower sequence (WPS55 in Supplemental
flower. Although expression of the three genes relating Table S2), a putative member of the Rab small GTPases,
to auxin signaling (WPS46, WPS103, and WPS53 in was weakly up-regulated in old wallflower petals (2.3-
Supplemental Table S2) was up-regulated with age in fold) but was stable in wallflower leaves. However,
both tissues of both species, expression of a putative the Arabidopsis homolog (At1g49300) was stable with
cytokinin oxidase (wallflower probe, WPS96; Arabi- age in both Arabidopsis petals and leaves on the
dopsis gene, At1g75450) was strongly up-regulated in Affymetrix arrays. Finally, a putative rhodopsin-like
Arabidopsis leaf (8.5-fold) but only very weakly in receptor gene also differed in expression pattern in the
petals (1.5-fold) on the Affymetrix arrays. In contrast, two species. Expression of the Arabidopsis gene
the wallflower homolog (WPS96 in Supplemental (At1g12810) was up-regulated in both tissues (leaves,
Table S2) was strongly up-regulated in both wallflower 2.8-fold; petals, 2.1-fold) on the Affymetrix arrays,
tissues (leaves, 5.8-fold; petals, 8.9-fold). Two sequences while the wallflower homolog (WPS95 in Supplemen-
relating to Rab signaling were identified from the tal Table S2) was only up-regulated in old wallflower
wallflower libraries. Expression of a wallflower Rab petals (2.9-fold) but not in old leaves.
acceptor homolog, WLC13A (Supplemental Table S2), Four transcription factors were also identified on the
was strongly up-regulated in old wallflower petals wallflower arrays. Expression of a WRKY75 homolog
1906 Plant Physiol. Vol. 147, 2008
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.A Comparison of Leaf and Petal Senescence in Wallflower
(WLS67) and two members of the No Apical Meristem the sequenced genes that were up-regulated in senes-
(NAM) family (WPS52 and WLS62) was up-regulated cent wallflower petals related to remobilization (38%
in both tissues in both species (Supplemental Table S2). overall). This is in agreement with transcriptomic
Expression of the Arabidopsis homolog of the WRKY75 studies of petals from other species in which wilting
transcription factor (At5g13080) was up-regulated occurs (Alstroemeria [Breeze et al., 2004] and Iris [van
much more strongly in old leaves compared with old Doorn et al., 2003]). The vast majority of genes related
petals (leaves, 202-fold; petals, 13.8-fold) on the Affy- to remobilization were up-regulated in both senescent
metrix arrays, whereas the expression pattern of its wall- petals and leaves, and the largest proportion of the
flower homolog, WLS67, was reversed, with much genes whose expression was up-regulated with age in
stronger up-regulation in old wallflower petals (67-fold) both tissues were putatively involved in remobiliza-
compared with old wallflower leaves (7-fold). There tion. Including SAG12, these represent just over half of
was a similar contrast in pattern for one of the NAM the genes in this category. Again, this agrees with other
family transcription factors (At5g64530/WLS62 in transcriptomic studies of leaves (Guo et al., 2004;
Supplemental Table S2). Expression of this gene was Buchanan-Wollaston et al., 2005) and petals (van
much more highly up-regulated in old petals com- Doorn et al., 2003; Breeze et al., 2004). All of the
pared with old leaves in wallflower (leaves, 2.2-fold; SAG12 targets belonged to this expression class as
petals, 22-fold), while on the Arabidopsis Affymetrix expected, and metal-binding proteins were also well
arrays the pattern was reversed (leaves, 4.3-fold; represented, again reflecting other studies discussed
petals, 2-fold). Finally, expression of a CCR4 family above. However, three genes related to remobilization
protein (WLS63 in Supplemental Table S2) was up- were specifically up-regulated in wallflower petals
regulated in both aging Arabidopsis leaves and petals and not leaves, a AAA-type ATPase family protein, a
(leaves, 2.8-fold; petals, 4.0-fold) on the Affymetrix lipid transferase, and a thiol protease. These may be
arrays, while in wallflower it was only up-regulated in interesting genes to study further.
petals with age (3.7-fold) and stable in leaves. While the process of remobilization, and many of
the genes involved, are shared between petals and
leaves in wallflower, the timing of both physiological
events and gene expression in the two organs differs.
DISCUSSION Whereas in petals the dry weight-fresh weight ratio
Remobilization during Petal and Leaf Senescence was rising well before any visible signs of wilting, in
in Wallflower leaves the first signs of visible senescence, and the
drop from maximal chlorophyll levels, coincided with
Species can be broadly divided into those in which the start of the rise in dry weight-fresh weight ratio.
petals wilt before abscission and those in which petals The loss of both fresh weight and dry weight was
abscise at full turgor (van Doorn and Stead, 1997). comparable between petals and leaves; however, the
Generally, the longer the petals persist, the more extent of protein breakdown differed, with 65% of the
remobilization of nutrients is likely to occur. Patterns maximal level of protein remaining in petals by stage 5
of dry weight-fresh weight ratio changes during wall- compared with only 5% in stage 7 leaves. The fall in
flower petal senescence are consistent with data from leaf protein coincided with chlorophyll degradation,
other genera, such as Alstroemeria and Tulipa (Collier, reflecting the fact that the majority of remobilized
1997), Hemerocallis (Lay Yee et al., 1992), Digitalis protein from leaves is from chloroplasts (Thomas and
(Stead and Moore, 1977), and Sandersonia (Eason and Donnison, 2000). The precise timing of SAG12 expres-
Webster, 1995), in which some wilting occurs before sion also differed between the two organs when ex-
abscission. However, in wallflower, the magnitude of amined more closely by RT-PCR. The leaf data are in
change between the maximal values of open flowers agreement with data from Arabidopsis (Lohman et al.,
and heavily wilted flowers is quite low (at stage 5, 1994), with up-regulation of SAG12 late in senescence.
fresh weight and dry weight are 41% and 67%, respec- However, in petals, SAG12 is already substantially up-
tively, of the maximum) compared with Hemerocallis, regulated in mature nonsenescent flowers and falls to
in which fresh weight decreases to 2% of maximum less than 20% of maximal levels by late senescence.
and dry weight decreases to 33% of maximum. The SAG12 encodes a papain-like Cys protease located in
change, however, is greater than in Digitalis (dry senescence-associated vacuoles. It is presumed to play
weight remains at 88% of maximum) or Alstroemeria a role in proteolysis; however, sag12 knockouts are not
(dry weight remains at 80% and fresh weight at 40% of perturbed in their leaf senescence (Otegui et al., 2005).
maximum). Thus, wallflower petals appear to be more Assuming that the role of SAG12 is equivalent in
similar to Alstroemeria in their loss of fresh weight (40% petals and leaves, the different expression programs
of maximum in Alstroemeria) but closer to Tulipa in the could reflect different patterns of cellular degradation.
loss of dry weight (60% of maximum in Tulipa). This Electron microscopy of petals reveals very early cel-
indicates a flower in which there is substantial, but not lular death in much of the mesophyll while the epi-
extreme, wilting before petal abscission and in which dermal cells remain intact (Weston and Pyke, 1999;
some remobilization is probably taking place. This is Wagstaff et al., 2003). Hence, perhaps the majority of
supported by the array results: a large proportion of SAG12 activity is already complete in many petal cells
Plant Physiol. Vol. 147, 2008 1907
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Price et al.
at a relatively earlier stage of organ senescence. The nescence in wallflower may be a reduction in cytokinin
temporal difference in expression patterns in different levels via cytokinin oxidase. In carnation (Dianthus
cell types is something that array experiments often caryophyllus) petals, sensitivity to ethylene of excised
overlook, and it is only with laser dissection micros- petals was reduced by exogenous application of cyto-
copy, or other single cell-based PCR techniques, that kinin (Taverner et al., 2000), indicating cross talk be-
these differences will be elucidated. tween these two growth regulators, which is worthy of
The expression of another gene with a presumed further investigation in wallflower. The Arabidopsis
role in remobilization, WC11, encoding a putative cytokinin oxidase gene, At1g75450, was only very
peptide transporter, was also examined by RT-PCR, weakly up-regulated in petals (1.5-fold) on the Affy-
and the expression pattern of this gene also differs metrix arrays, whereas up-regulation in leaves was
between the two organs. Although WC11 expression is much more significant (8.5-fold). This could indicate
strongly up-regulated according to the array during either temporal or species-specific differences in the
both petal and leaf senescence, RT-PCR shows that the role of this enzyme in petal senescence.
pattern is more complex. It was expressed from young One aim of this study was to identify genes that
leaves through to mature leaves, with increased ex- might indicate differences in the regulation of petal
pression early in senescence. In contrast in petals, and leaf senescence. Only one wallflower sequence
there is a clear up-regulation that precedes other signs related to transcriptional regulation was identified in
of senescence, and expression remains high. Peptide the class of probes from the wallflower microarray that
transporters form a superfamily of structurally related were up-regulated in petals but not in leaves; this was
membrane proteins (Chiang et al., 2004). Different WLS63, a CCR4-related gene. In yeast, the CCR4
members of the Arabidopsis gene family show tissue- protein forms part of the CCR4-NOT complex, which
specific expression. The closest Arabidopsis gene to acts as an RNA deadenylase, and is involved in
WC11, At1g32450, is part of the PTR family, trans- nutrient and stress sensing (Collart, 2003). The role
porting dipeptides and tripeptides (Waterworth and of these genes in plants has not been fully investigated.
Bray, 2006), although it does not fall into one of the RT-PCR showed that expression patterns of WLS63 in
major subfamilies. The Arabidopsis gene is expressed wallflower are very different between leaves and
in mature tissues and is strongly up-regulated in leaf petals. WLS63 peaks in expression in petals relatively
senescence (Affymetrix data from Genevestigator). late, at stage 4, when petals are already showing
Hence, this gene may have a function both during visible signs of senescence, after the peak in SAG12
leaf development and during the remobilization oc- expression. This suggests that it may be important in
curring during leaf senescence. In petals, the role may mRNA stability late in senescence, perhaps targeting
be different, in that expression during early develop- specific transcripts for degradation. In leaves, WLS63
ment is very low and there is a far greater up-regulation transcripts are at their highest levels in young leaves
during senescence, indicating a more specific role in and fall thereafter to lower levels of expression. This
senescence-associated remobilization. could imply either that it is not involved in leaf
senescence in this species or, alternatively, that its
down-regulation stabilizes specific transcripts.
Regulation of Wallflower Petal and Leaf Senescence
Shared and Petal-Specific Gene Expression
Pulse treatment of cut flowers with STS indicated
that ethylene is involved in both petal senescence and The high prevalence of SAG12 clones (14% overall;
abscission in this species. It was a surprise, therefore, 8% of petal clones and 21% of leaf clones) is expected
not to find more genes related to ethylene biosynthesis due to the close taxonomic relationship to Arabidopsis
or responses in the petal SSH library. In fact, only one and Brassica, in which SAG12 is a highly abundant
ACC oxidase-like gene was found. This gene, how- transcript in senescent leaves (Lohman et al., 1994;
ever, was strongly up-regulated in both senescent Guo et al., 2004). Although metallothioneins were
leaves and petals, as expected. In addition, Arabidop- represented in both libraries (6% in petal and 7% in
sis ACC oxidase on the array was up-regulated 3-fold leaf), the levels were not as high as those found in
when hybridized to messages from wallflower petals. other EST studies of petal senescence, in which they
Many of the SSH library genes represented 3# untrans- were present at levels of 19% in Alstroemeria (Breeze
lated region sequences and were thus difficult to annotate; et al., 2004) and 11% in Rosa (Channeliere et al., 2002),
therefore, it seems likely that further ethylene-related indicating species-specific differences in the expres-
genes are up-regulated in both leaf and petal wallflower sion of these genes and perhaps in their role in petal
senescence but were not identified as such. senescence. Metallothioneins have been found in other
Treatment with cytokinin (kinetin) delayed both petal studies of leaf ESTs, although not at such high levels as
senescence and abscission, as did treatment with the in wallflower (e.g. rice [Oryza sativa] mature leaves, 3%
inhibitor of cytokinin oxidase, 6-methyl purine. A cyto- [Gibbings et al., 2003]; senescent Arabidopsis leaves,
kinin oxidase gene (At1g75450, WPS96), was strongly 3% [Guo et al., 2004]).
up-regulated in old petals in wallflower (9-fold). Thus, Two genes were found at unexpectedly high fre-
part of the mechanism for the regulation of petal se- quency in the array class up-regulated in senescent
1908 Plant Physiol. Vol. 147, 2008
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.A Comparison of Leaf and Petal Senescence in Wallflower
petals but not leaves: chitinases and GSTs. The very high level of agreement. Thus, although caution must
high abundance of chitinase genes in the wallflower be exercised in the interpretation of data from cross-
petal libraries (23%) was a surprise, and there was a species experiments, due to the complications of gene
clear interorgan difference, with only 2% of the genes families and inherent difficulties in precisely assigning
found in the leaf library identified as chitinase. Al- stages of development, these data strongly support the
though chitinase transcripts have been reported as use of this approach here.
senescence enhanced in other species in leaves in Due to the close taxonomic relationship between the
Brassica (Guerrero et al., 1990; Hanfrey et al., 1996) two species, floral architecture in wallflower and
and petals in Alstroemeria (Breeze et al., 2004), EST Arabidopsis is similar, and in both species leaves
studies of senescent petals or leaves have not revealed form sequentially in a spiral. However, wallflower
the high abundance found here. The major role of petals differ from Arabidopsis petals in their purple
chitinases was usually thought to be in pathogen pigmentation and much slower development and
defense, and they are classed as pathogenesis-related senescence. Differences in leaf senescence strategy
proteins. However, it is becoming clear that chitinases might also occur due to the diverse life cycles in the
may also have roles in signaling and programmed cell two species: perennial in wallflower and ephemeral
death (Kasprzewska, 2003). Arabidopsis. Genes that share expression patterns
GSTs are up-regulated in petal senescence in other between the two species thus reflect perhaps the
species, such as carnation (Meyer et al., 1991). Al- underlying evolutionary conservation, while those
though the role of most plant GSTs is unclear (Wagner with differing patterns may reflect species-specific
et al., 2002), some, including those of the Arabidopsis strategies. Over one-third (38%) of the Arabidopsis
f class, may act as glutathione peroxidises, protecting genes on the array and 53% of the wallflower genes
cells from ROS damage, while others may have roles in shared gene expression patterns in the two species,
hormone metabolism. Two wallflower targets were indicating a conservation of a significant portion of the
most closely homologous to AtGSTZ1, which is in- gene expression profile. However, a number of genes
volved in Tyr metabolism (Dixon et al., 2000). The differed in expression pattern between the two spe-
other wallflower clones were closest in amino acid cies. These include both the AtGSTZ1 gene and the
sequence to f class GSTs from Arabidopsis: AtGSTF2 Arabidopsis chitinase gene (At2g43570), which were
and AtGSTF3. AtGSTF2 is membrane associated (Zettl up-regulated with senescence in Arabidopsis leaves
et al., 1994) and ethylene responsive; both AtGSTF2 while the wallflower homologues were not. These
and AtGSTF3 have a putative ethylene-responsive differences may reflect a divergence of senescence
enhancer element in their promoter sequences similar strategies in the two species and, again, would be
to that found in the petal senescence-enhanced carna- interesting for future studies.
tion GST (Itzhaki et al., 1994) and are also up-regulated
by salicylic acid (Wagner et al., 2002). AtGSTZ1 tran-
scription is not induced by ethylene but is induced by CONCLUSION
methyl jasmonate, and both AtGSTF2 and AtGSTZ1 This study has revealed considerable differences in
are induced by the auxin analog 2,4-dichlorophenoxy- gene expression during senescence both between
acetic acid (Wagner et al., 2002). Thus, GSTs are clearly petals and leaves and between two closely related
involved in processes related to senescence, and their species. Further work to understand petal and leaf
up-regulation in wallflower petals but not in leaves senescence in these species will exploit the advantages
may reflect important differences in the regulation of of wallflower for biochemical studies and the myriad
senescence by plant growth regulators or in the fine resources for forward and reverse genetics available
control of senescence progression in these two tissues. for Arabidopsis.
Clearly, the role of GSTs in wallflower petal senescence
is also worthy of further investigation.
MATERIALS AND METHODS
Cross-Species and Cross-Tissue Comparisons of Plant Material
Expression Patterns
Leaves and petals were collected from wallflower (Erysimum linifolium
‘Bowles Mauve’) and staged (Figs. 1 and 2). Material for RNA extraction was
Overall, 57% of the Arabidopsis genes on the array
immediately frozen in liquid nitrogen and stored at 280°C until required.
gave consistent results when hybridized to the wall-
flower transcripts. This compares favorably with other
studies using species taxonomically related to Arabi- Cut Flower Treatments
dopsis (e.g. in Thlaspi arvense arrays, only 31% of Individual flowers were detached from the raceme at stage 1, and the
probes cross-hybridized to Arabidopsis [Sharma et al., pedicel was immediately submerged in water. Flowers were held at 20°C and
2007]). Likewise, for the six wallflower genes on the 16 h of light either in water or in solutions of kinetin (1.0 or 0.1 mM) or 6-methyl
purine (0.1 mM; Sigma-Aldrich). For ethylene inhibitor treatment, flowers
array, for which the closest Arabidopsis homolog was were held in STS (4 mM AgNO3:32 mM NaS2O3) for 1 h and then transferred to
also included on the array and data were available water. Each experiment consisted of 10 replicate flowers, which were mon-
from the Affymetrix experiments, there was a very itored daily to record senescence stage and day of petal abscission.
Plant Physiol. Vol. 147, 2008 1909
Downloaded on February 16, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































