Fish Eggr and Larvae of the Hauraki Gulf, New Zealand - Fisheries Research Bulletin No. 23 - by - Fisheries Research Division I. ew Zealand ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Fisheries Research Bulletin No. 23
Fish Eggr and Larvae of the
Hauraki Gulf, New Zealand
by
J. Crossland
Fisheries Research Division
I.[ew Zealand Ministry of Agriculrure and Fisheries
Fish Eggr and Larvae of the
Hauraki Gulf, I.{ew Zealand
$r'- ¡- -j
Fisheries Research Bulletin No. 23
Fish Eggr and Larvae of rhe
Hauraki Gulf, New Zealand
by
J. Crosslan{
Fisheries Research Division,
Ministry of Agriculture and Fisheries,
Wellington, New Zealand
Fisheries Research Division
It{ew Zea\and Ministry of Agriculture and Fisheries
19Br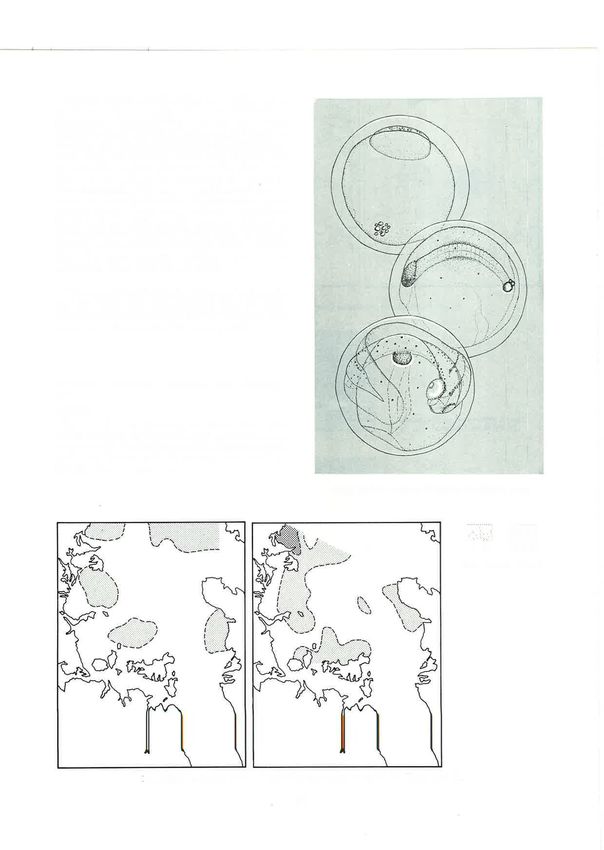
Published by the New Zealand Ministry of
Agriculture and Fisheries
Wellington
l98r
ISSN 0l l0-1749
R¿c¿iu¿d 28 February 1978; reuisíon receiu¿d 4 June 1980FOREWORI)
Jnvrs CRossI-AND has already used the distribution and abundance of the eggs
and larvae of snapper to assess the standing stocks of this most valuable species
in New Zealand's most important cornmercial fishery. llowever, despite the
rapid expansion of fishing effort over the past few years, our knowledge of the
available resources, even in the Hauraki Gulf, has remained meagre.
The data in this bulletin not only add to our undeistanding of the other
species in the Hauraki Gulf, but are an important prerequisite to our
appreciation oÍ its significance as a spawning or nursery ground {or fisheries
elsewhere. At the same time, this type of information will be of value as a
background for integrated management of the total resources of the area in the
future.
G. DUNcAN Weucn,
Director, Fisheries Research DivisionCONTENTS
Pog,
INTRODUCTION il
MATERIALAND METHODS 1l
Sampling and Sorting 1l
Identific¿tion of Eggs and Lawae ll
Descriptions of Eggs and Laruae t2
Daø Presentation l2
RESULTS l3
Family Engraulidae 13
Family Clupeidae t4
Family Muraenidae t7
Family Congridae 20
Family Moridae. .. 20
Family Exocoetidae oq
Family Trachichthyidae 22
Family Zeidae ... 23
Family Syngnathidae 23
Family Triglidae 23
Family Acanthoclinidae 2+
Famiiy Carangidae 26
Family Sparidae.. . 31
Family Mugilidae 34
Family Labridae 34
Family Uranoscopidae 35
Families Blenniidae, Clinidae, and Tripterygiidae 5t)
Family Eleotridae 38
Family Gempylidae 38
Family Scombridae 38
Family Bothidae +0
Family Pleuronectidae 4l
Family Balistidae 49
Family Diodontidae 5+
DISCUSSION 56
Sampling Period 56
Effect of Temperature on Spawning.. . 56
Effect of Water Movements on Distribution of Larvae . .. 57
SUMMARY 58
REFERENCES 59
ACKNOWLEDGMENTS ... 59FIGIJRES
Page
l. The Hauraki Gulf, showing places mentioned in the text l0
2. Survey stations and study area t2
3. Distribution of anchovy eggs by cruises, 197 4-7 5 and I 975-76 t.t- I 5
4. Numbers of ancÌíovy eggs within the study area, lg74-75 and 1975-76 l5
5. Distribution of anchorry laruae by cruises, I 974-75 and I 975-76 .. . l6
6 Distribution of pilchard eggs,4-8 November 1974 t7
7. Distribution of pilchard larvae by cruises, 1974-75 and 1975-76 ... l8
8. Numbers of pilchard larvae taken by cruises, 197 4-7 5 and I 975-76 t9
9. Cumulative distribution of sprat eggs, 1974-75 and 1975-76 l9
10. Moray eel eggs l9
II. Cumulative distribution of moray eel eggs, 197 4-7 5 20
12. Morayeellarva ... 20
I 3. Distribution of ahuru eggs by cruises, 197 +-7 5 and I 975-76 21
14. Numbers of ahuru eggs taken by cruises, 1974-75 and 1975-76 22
15. Ahu¡u larvae .. 22
16. Numbers of ahuru larvae taken by cruises, 1974-75 and 1975-76
I 7. Distribution of ahuru lawae by cruises, 1974-75 and 1975-76 23-24
18. Garfish larva 2+
19. Roughylarva 2+
20. Cumulative distribution of red gurnard eggs,197+-75 and 1975-76
21. Numbers of red gurnard eggs within the study area,1974-75 and 1975-76 2s
22. Cumulative distribution of red gurnard law¿e, 197 4-7 5 and I 975-76 25
23. Rockfish larva 26
24. Cumulative distribution of rockfish larvae, 1974-75 and 1975-76... 26
25. Jack mackerel Trachuns noaae4landiae eggs 27
26. Distributionof jackmackerelT.noue¿elandiaøeggsbycruises, 1974-75and 1975-76 ... 28
27. Numbers of jack mackerel T. noaaezelandia¿ eggs within the study area, 1974-75 and 1975-76 29
28. Jack mackerel T. noaa¿zelandia¿lawae 29
29. Distributionof jackmackerelT.nooaez¿lundiaelaruaebycruises, 1974-75 and 1975-76 30
30. Numbers of jack mackerel T. noaaezelandia¿lawae taken by cruises , I97 4-7 5 and I 975-76 3t
3 I Eggs of jack mackerel Trachutas decliuis 3l
32. Cumulative distribution of jack mackerel T. declitis eggs,1974-75 and 1975-76 J1
33. Snapper lawae 32
34. Distribution of snapper larvae by cruises, 197 4-7 5 and 197 5-7 6 .. JJ
35. Numbers of snapper laruae taken by cruises, Ì97,[-75 and 1975-76 5+
36. Yellow-eyed mullet egg 34
5f,
37. Cumulative distribution o{ yellow-eyed mullet eggs, 1974-75 anð 1975-76
larva 35
38. Yellow-eyed mullet .. .Page
39. Spotty lawa 35
,10. Spotted stargazer eggs
36
41. Cumulative distribution of spotted stargazer eggs, 197,1-75 and 1975-76 36
42. Spotted stargazer larvae JI
43. Cumulative distribution of spotted stargazer larvae, 197 4-7 5 and I 975-76 ...
.14. Graham's gudgeon larvae
45. Cumulative distribution of Graham's gudgeon laruae,.1974-75 and 1975-76 39
46. Blue mackerel egg 39
47. Numbe¡s of blue mackerel eggs taken by cruises, 1974-75 39
48. Distribution of blue macke¡el eggs by cruises, I97,t-75 and t97 5-76 +0
49. Blue mackerel laruae 4l
50. Numbers of blue mackerel larvae taken by cruises, 1974-75 and 1975-76 +1
51. Distribution of blue macke rel laruae by cruises, 1974-75 and 1975-76 42
52. Crested flounder eggs +3
53. Cumulative distriburion of crested flounder eggs, 1974-75 +3
54. Crested flounder larvae ++
55. Distribution of crested flounder laruae by cruises, lg7+-75 and 1975-76 +5
56. Distribution of pleuronectid eggs by cruises, 197+-75 and 1975-76 +G-47
57. Numbers of pleuronectid eggs taken by cruises, lg7+-75 and 19?5-76 +8
58 Flounder lawa +8
59. Cumulative distribution ol flounder laruae, 1974-75 znd 197:_76... 48
60. Numbers of flounder larvae raken by cruises, 1974-75 and 1975-76 49
6l . Speckled sole larvae 49
62. Cumulative distribution of speckled soìe larvae, lg74-75 and lg75-76 50
63. Numbers of speckled sole laruae taken by cruises, 1974-75 and 1975-76 50
64. Leather jacket eggs 50
65. Numbers o{ leather jacket eggs within the study area, lg74-75 and 1975-76 50
66 Distribution of leather jacket eggs by cruises, 1974-75 and 1975-76 5 r-52
67. Leather jacket larvae 53
68. Cumulative distribution of leatherjacket lawae,1974-75 and 1975-76 )+
69. Unidentified egg I 54
70. Unidentified egg 2 55
71. Unidentified egg 3 55
72. Numbers of eggs of all species taken by cruises, 1974-75 and 197i:-16 57
73. Monthly cumulative wind speed from obseruations at Tiritiri Matangi Island 58
TABLES
Page
I. I(nds of eggs and larvae collected in the Hauraki Gulf . .. .
2. Spawning season and remperarures for the principal species in the Hauraki Gulf 56
3. Comparative eggproduction for 1974-75and 1975-76 ... 57
Inset I175"E 3ó"S
Little
Greot Borrier I
Omoho Boy
Colvílle
chonn-l
.#--th*"wou I
ärr( Cope Colville
/ 30
Tiririri
^\Motongi
I
Rokino ,0 " .',
Firth
of
Thomes
Fig. l:The Hauraki Gull, showing places mentioned in the text'INTRODUCTION
The Hauraki Gulf, New Zealand's richest fishing There are few published accounts of ichthyoplank-
ground, is a large, relatively shallow embayment with ton surveys from New Zealand. In the Hauraki Gulf
many islands and harbours (Fig. l). It is an Cassie (t956a) studied the distribution of snapper
important spawning ground for many fish species, eggs, and Colman (1973) studied the spawning of two
including snapper (Chrysophrys auratus), which domi- species of flounder. Baker (1972) surveyed pilchard
nates the commercial landings. During the spring- eggs and larvae in Tasman Bay, Robertson (ì980)
summer periods (October to January) oÍ 1974-75 and recorded the distribution of the eggs of 17 species off
1975-76,2-weekly plankton surveys were carried out the Otago coast, and Colman (1979) studied the
with the aim of estimating the stock size of snapper spawning of the sprat around the South Island.
based on egg counts (Crossland 1980). These surveys Additional information on the eggs and larvae of
were specifically designed for sampling snapper eggs; species mentioned in this bulletin is given in Elder
however, many types of fish eggs and larvae were (1966), Robertson and Raj (1971), Baker (1973),
collected, identified, and counted. These data are Robertson (1973), Castle and RÒbertson (1974), and
presented in this bulletin. James (1976). Robertson's (1975) key describes the
planktonic eggs of 56 species of New Zealand fishes.
MATERIAL AND METHODS
SAMPLING AND SORTING and larvae and sorted them according to type. When
the sorters were sufficiently experienced to identify the
Plankton samples were taken by oblique tows of a
at the stations shown in Fig. 2. different kinds accurately, the common ones were
cylinder-cone net
counted in situ in the samples. Regular checks were
Details of the net were: mouth area0.25 m2; cylinder
made on the accuracy of identifications and sample
56 cm diameter by 150 cm long; cone slope length
counts. Large samples were divided with a Folsom
185 cm; gauze 0.425-mm aperture with a porosity of
splitter until a manageable fraction was obtained. In
47"/.. Ttie total filtering area was 2.225 m2 and the
samples where eggs were abundant but larvae were
filtering'area/mouth area ratio was 8.9, made up of
not, eggs were subsampled, and the whole sample was
5.35 in the cylinder a¡rd 3.55 in the cone. The volume
sorted for larvae. Details of the dates of sampling,
of water sampled (range 50-300 m3) was recorded numbers of stations, ancl sea temperature data and
with a digital flow meter, type 2030 by General
further information on the sampling technique can be
Oceanics, Inc., which was mounted at a distance of a
found in Crossland (1980).
quarter the diameter from the rim of the net. Tows
were made at a boat speed of 2 to 2.5 knots, with the
net usually descending to within 2 to 4 m of the IDENTIFICATION OF EGGS AND LARVAE
bottom. Robertson's (1975) key was useful in identifying
Samples were preserved in 5y" neutralised many of the eggs. Other eggs and larvae were
formalin. During sorting all fish eggs and larvae were identified by building up a series of developmental
counted. Initially plankton sorters removed all eggs stages from egg to larva to recognisable juvenile.
ll.40 ll. 42.
33. 34. 35. 36. 37. 38. 39.
28. 26. 25.
aa
21. zlA!
i'frãJ iot ''.
ì5. 14. 13.
Fig 2: Suruey stations and study area. Left: 1974-75. Right: 1975-76.
Larvae of some species were identified by their DATA PRESENTATION
similarity to those of related species which had been This study was made on a more intensive scale
described from other parts of the world. Electrophor- (that is, at shorter time intervals and with stations
etic tissue analysis aided in identifying snapper larvae closer together) than is common with ichthyoplankton
(Smith and Crossland 1977) and larvae of the spotted surveys. The complete data, which are held in
stargazer. The distribution and abundance of species Fisheries Research Division library, give the counts
occurring locally as adults were also considered. recorded for each type of egg and larva for every
sample taken; these were calculated as numbers per
DESCRIPTIONS OF EGGS AND LARVAE 100 m3 (eggs) and numbers per 500 m3 (larvae).
This bu-letin describes and illustrates eggs and !'or some of the more common egg types the
Iarvae taken or refers to published accounts.
densities were integrated over the study area to give
The descriptions of larvae are intended to highlight estimates of the total numbers present at one time on
features which will allow recognition of the individual that cruise. This was done by multiplying each
species; they are not meant to be detailed, stage-by- individual station volume (station area by mean
stage descriptions of larval growth and morphology. depth) by its egg density and then summing over the
Most species can readily be distinguished. Exceptions whole study area. The calculations were adjusted for
are the two species of. Rhombosole¿ and the two species the slight variation in area between the 2 years. The
of. Trachurus, all of which may be present in the method of integration is explained fully in Crossland
sampling area. (1980) . The embryonic period was not known for any
A1l descriptions and measurements apFly to of the eggs; so it was not possible to calculate total egg
material which was preserved in 5"/" neutralised production.
formalin. Measurements were made under a stereo Distribution maps were plotted for the more
microscope at a magnification of 62 x. The length common tyPes of eggs and larvae as numbers below
recorded {or larvae was total length. The figures o{ l0 m2. Cruises not shown in the map series recorded
eggs and larvae were also made by use of a stereo fewer eggs and larvae than the minimum contour level
microscope and a drawing tube. for that species.
t2RESULTS
Both fish eggs and larvae were very abundant in the immediately recognisable, as the only other species
Hauraki Gulf. In 1974-75 the 330 samples contained known from New Zealand to have an elliptical egg,
583 000 eggs and 55 000 larvae. In 1975-76, 589 000 the carapid Echiodon sp., cloes not occur in the Hauraki
eggs and 79 000 larvae were collected in 426 samples. Gulf.
The maximum number of eggs taken in one haul was
89 000, and that of larvae was 6000. For both years
combined, only 2 samples contained no eggs, and 2l The distribution of anchovy eggs for the 2 years
contained no larvae. The kinds of cggs and larvae (Fig. 3) showed a spawrring centre within the study
recognised are shown in Table l. Details of the area. Anchovy eggs were common and occurred on all
distribution, seasonality, and abundance for eggs and cruises in both years. During October spawning was
larvae of each species are given below. The species light and took place in the southern part of the gulf
have been placed in phylogenetic sequence according near Waiheke Island and the northern part of the
to Nelson (1976). Firth of Thames. The main burst of spawning
observed was during November in 1974, and
December in 1975. The centre of spawning moved
The relationship between spawning and tempera- north to the central gulf-Kawau Island area as the
ture was also considered. For demersal species (for season progressed, a similar pattern to that observed
example, flounder or gurnard) the sea bottom for snapper (Crossland 1980). Spawning was still
temperature was used; for pelagic species (for taking place at the end o{ the sampling period in each
example, anchovy or pilchard) the sea surface year.
temperature; and for species occurring throughout the
water column (for example, jack mackerel or ahuru)
the mean of bottom and surface temperatures. The relationship betw,een temperature and spawn-
ing (Fig. 4) showed that spawning began when rhe sea
surface temperature was about l6"c and was most
intense at l9o-20"c. The number of eggs produced in
Family Engraulidae the 1975-76 study period was about three times that
of the previous year (Fig. 4). These figures have been
Anchory Engraulis australis (White, I790) integrated over the stucly area, but not over time,
Eggs. The anchor,y egg is described and {igured in because the embryonic period of the anchovy is not
Robertson (1975). Its elliptical shape makes it known. Crude egg counts do not consider the effect of
TABLE l: Kinds of eggs and larvae collected in the Hauraki Gulf, October to January in 197Ç75 and 1975-76
Species Egg. Lawae Species Egg. Lawae
Anchorry (Engrailis australi) I + Snapper (Chrysoþhrys auratus) + +
Pilchard (Sardinops neopilchardus) + + Yellow-eyed mullet (Aldrichetta forster) + +
Sprat (Sprattus ant;þodum) + -l Grey mullet (Mugil cephalus) - +
Moray eel Family Muraenidae + + Spotty (Pseudolabrus celidotus) - +
Sand eel Gtathophis sp. + Spotted stargazer (Gcnyagnu monoþterygius) + +
Alauru (Auchmoccros
Garfi
Roug
þanctatus)
ihr)
chthyidae
l +
+
+
Blennies: Family Blenniidae, Family Clinidae,
Family Tripterygiidae
Graham's gudgeon (Grahamíihthys radiatus)
+
+
John as) + Bar¡acouta (Tþrsítes atun) +
Seaho¡se (Hippocampus abdominalis) + Blue mackerel (Scomber australasicus) + +
Long-snouted pipefish (Stigmatoþhora longiros tns) + Crested flounde¡ (Lophonectes gallus) + +
Flounder Rhombosolea þIebcía &. R. leporina + +
Red gurnard (Chelídonichthys kunu)
Rockfish Acanthoclinus quadridacllus I +
+ Speckled sole (Peltorhamþhus latw) +
+
+
Rockfish A. trilin¿atus + Leather jac*.et (Nooodon coruexirostris) +
Jack mackerel Trachurus noaa¿z¿landiae + + Porcupine Tts}:, Allomycterus jaatlifents +
Jack nackerel T. decliais + Three unidentified species +
+ Present.
t Absent.
IJ4-8 Nov
îÅffi
1974
î,tffi
ó-'l0 Jon 1975
5000- >t0 000
r0 000
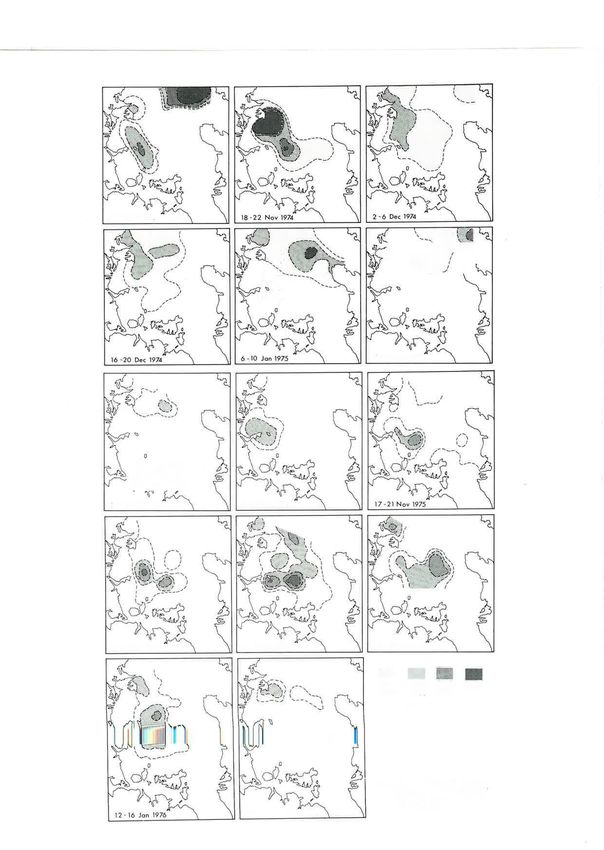
No. below l0 m2 Fig. 3 (above and right): Distribution of anchovy eggs by cruises, 1974-75 and 1975-76.
temperature on development. The egg production larvae were in the central and outer gulf, further north
shown in Fig. 4 has been adjusted on the basis that the than the eggs. Large numbers of larvae occurred on
development period is halved for a l0oç rise in the November cruises in 1974, when the numbers of
temperature. This effect was found experimentally for eggs were low. This probably resulted from bursts of
snapper eggs {rom the Hauraki Gulf (Cassie 1956b). spawning which took place between sampling cruiscs
and were thus not sampled, and it showed that even
Larvae. The anchovy larva is described and figured sampling at 2-weekly intervals may be insufficient to
in Baker (t972). It may readily be distinguished from measure the egg production of the mobile, shoaling
larvae of the other lttrew Zealand clupeoids pilchard ancholy.
(Sardinoþs neoþilchardus) and sprat (Spratns antipodum)
by its underslung jaw and the relative positions of its
anal and dorsal fins, which overlap. The latter feature
is particularly useful for identification, even in small
larvae. Family Clupeidae
Pilchard Sardinoþs neoþilchardus (Steindachner, IB79)
The distribution of anchovy larvae (Fig.'5) differed Eggs. The development of the pilchard egg is
from that of the eggs. The main concentrations of described and Iigured in Baker (1972). These eggs
I4I Z-21 Nov 1975 I -5 Dec 1975 15 -]9 Dec ]975
î,tffi
29 -31 Dec 1975
*ffi
12-16 )on197ó
îtffi
27 -3Q Jon 1976
rvere present from the beginning of sampling in each 24
year (October) until mid November in 1974 and mid
I)ecember in 1975. They were seldom abundant, and 22
on only one cruise (,1.-B November 1974) was there a
large spawning within the study area (Fig. 6). 20^
(J
Larvae. The pilchard larva is described and figured l8 c¡
f
in Baker (1972). Small pilchard larvae are difficult to
distinguish from larvae of the sprat Sþrattus antiþodum.
lóboo-
E
Larvae longer than about 25 mm can be identified by o
the differences noted by Baker. Pilchard larvae have a Ic o
,( o
melanophore below the eye and one at the base of the
l
pelvic fin, and the last ray in the anal fin is elongated. o)
o)
CJ o
These features are absent in sprat larvae. During this o 203
study all clupeid larvae large enough to be identified c)
were pilchard; smaller larvae of the same type were, -o
E
therefore, assumed to be pilchard also. Some error zf
may be introduced by this, but since pilchard is very
common in the Hauraki Gulf and sprat uncommon,
the error will be small. 1529 122ó1024721 4
Nov Dec Jon Feb
Fig.4: Number of ancholy eggs within the study area at the
The distribution of larvae in both years (Fig. 7) time of sampling for each cruise, 197 Ç-7 5 and 1975-76. Mean
showed that the main spawning areas of the pilchard sea surface temperatu¡es for each cruise are also shown.
t52- ó Dec 1974
0 r-.
i) â
A\/^L
l__r">
d
L,E-,.1-J
\uu4 0
cv"nä;t
^
l(\
//\ r^\^ .lV
(
6 -9 Oct 1975 lZ-20 Nov ì975 l-4 Dec 1975
â.
h-l / ¿¡
r !:u _-ì
./ | -- / t
\
\Wl'
s----:-- \-?'
!
I
-
fiQ s
- - -
'
/-t
l-/
!'>L
\J'Þ
^\/
il rsJ
^ r,4lJ,4 ov"Ëfd^L J çz-¿r,t¿)
t--'l\^L
*j. "'"q¡"J¿
!
auy4
/\.c> ! /\¡^..- ü As>
ló - 19 Dec 1975
rLiii ll|illiill
50-200 200-500 500- >looo
1000
No. below ìO m'
Fig. 5: Distribution of anchory larvae by cruises, 197'l-75 and 1975-76.
27 -3O Jon1976took place outside the study area; however, the sea
surface temperatures at the outer stations were in the
range 16.lo-17.8"c at the peak occurrence of larvae.
Sprat Sþrattus antipodum (Hector, 1872)
Eggs. The sprat egg is described and figured in
Baker (1973). These eggs were taken in only small
numbers and occurred sporadically in time and space.
Their cumulative distributions for the 2 years are
shown in Fig. 9. The largest patch of eggs was found
in late October 1975, off Kauri Point, Waiheke
Island.
Larvae. Sprat larvae were not recognised in any
sample, but they may have been confused with small
(less than 25 mm) pilchard larvae.
Adult sprat are occasionally taken by bottom trawl
4- 8 Nov 1924 in the Hauraki Gulf if a cod-end liner is fitted, but
they do not appear to be abundant. Baker (1973)
recorded sprat spawning in the Marlborough Sounds
from July to January. Robertson (I980), sampling off
the Otago coast, found that spawning lasted from
May to November and peaked in June-August. In the
500- 2000- 5000- >10 000 Hauraki Gulf the sampling period was outside the
2000 5000 r0 000 main spawning season found by these workers.
However, it is unlikely that major spawning took
No. below lO m2 place in the winters oÍ 1974 and 1975, because sprar
larvae or juveniles would have been recognised in the
Fig. 6: Distribution of pilchard eggs, 4-B Novembe¡ 1974. samples.
were north of the study area, probably in the Jellicoe
and Cradock Channels. The numbers of larvae taken
in each cruise are shown in Fig. 8. A similar pattern
was apparent in both years, but the peak lagged by 2
Family Muraenidae
weeks in 1975-76, when temperatures were lower.
Larval numbers reached a maximum in the first week Morayeel (unidentified)
of November in 1974, and in the third week in 1975. Eel eggs and larvae of a previously undescribed
Peak spawning would therefore have taken place type were occasionally taken in the outer stations of
earlier, possibly in late October or early November. the study area. Because of their close similarity to the
eggs and larvae of moray eels from European waters
(see Sparta 1939), they were identified as one of the
These results can be compared with those of Baker species of morays found in New Zealand. The most
(1972), who studied a population in Tasman Bay and commonly observed species locally is the yellow
found the breeding season to be lrTovember to moray (Llcodontis þrasinus (Richardson, 1847)).
February, with a peak in December-January; that is,
some 6 weeks to 2 months later than in the Hauraki
Gulf . The difference in latitude between Tasman Bay Eggs. These eggs hadi a diameter of 3.9-4.5 mm; a
(41" S) and the Hauraki Gulf (36' 30' S) is the wide perivitelline space (especially in early stages);
probable reason for this. Baker found that most and a finely segmented yolk with 6 to 40 oil droplets,
pilchard eggs were spawned when the sea surface 0.07-0.15 mm in diameler (Fig. l0). Eggs were first
temperature was between 16.0' and l6.4oc It was observed in mid December and were still present at
difficult to make a direct comparison with tempera- the end of the sampling period in late January. The
ture data from this study, because most spawning cumulative egg distribution for 1974-75 is shown in
T7â\\, -*_ _/--\\ sÈ{il!,
õJ ._--_-r 'r._____
*ffi
0
21 -25 Oct 197Á. l8 - 22 Nov 1974
al (_----
.-'-/'-l i.
13'4
l')Ä> ,y
16 - 20 Dec 1974
2\
ql
\\\\ i
lr
..' z-: ,/
?
',/
\.r'
0
)\ a
/\l¡-J-J
t)v> f-'>\.l r,'-'t?)
\J">
^L
l,â.2_. \
*!yrÌ " t¡ê,.i
-1N ](\ xuy4¡
o
//\ r\^ \l /lÄ> ,v
(
ó -9 Oct ì975 20 - 23 Oct 1975 3-7Nov1975
't''l q"rn ,;
0
îÅs
/-\ ^L/ d .rJ
\JVÞ 1,¡¿7e,-,.1 o
\uy4
\l\ )
2\ ñ
/À¡\Þ"v \) I
lZ- 20 Novì ló - 19 Dec 1975
,iffiffiffi
50-200 200-500500- >1000
1000
No below lO m'
Fig. 7: Distribution of pilchard larvae by cruises,1974-75 and 1975-76.Fig. ll. Spawning appeared to be located mainly
outside the study area, probably over the rocky
bottom around Great and Little Barrier: Islands, a
habitat favoured by morays. Fewer eggs were taken in
1975-76 because the outer part of the Hauraki Gulf
o was not sampled.
x
0)
o Larvae. Five larvae were taken in 1974-75 and two
o in 1975-76; they were found close to the eggs. They
o were immediately recognisable by their long, slender
shape and fearsome array of íorward-pointing teeth
¡
o
E
1975-76 (Fig. I2).
l
z
lrs29 1226 1024721 4 Fig.8: Numbers of pilchard la¡vae taken by cruises, I974-75
Ocl Nov Dec Jon Feb and 1975-76
5-20 20-50 50 - 100
No. below l0 m2
Fig. 9: Cumulative distribution
of sprat eggs. Left: 1974-75
Right: 1975-76.
,{
Fig. l0: Moray eel eggs
t9Family Moridae
Ahuru (Hutton, 1873)
Auchenoceros punctatus
Eggs. The ahuru egg is described and figured in
Robertson (1975). The distribution of ahuru eggs for
the 2 years is shown in Fig. 13. The centre of
spawning was in the northern part of the Firth of
Thames. Figure 14 shows seasonal changes in egg
numbers for the 2 years and the mean sea
temperatures in the spawning area. Few eggs were
observed in 1974, but they were common during
October and November 1975, when sea temperatures
were lower than in the previous year. These findings
are consistent with those of Robertson (1980), who
found that in Otago waters ahuru were winter-spring
spawners, preferring cool, Iow-salinity, in-shore
waters.
Larvae. The. main distinguishing features of the
larvae are the large numbers of rays in the dorsal and
anal fins, and in older larvae the single hair-like ray in
front of the dorsal fin (Fig. 15). Small larvae have no
particularly distinctive features and were recognised
after building up a developmental series.
..'..''.'''..'..''.''.':
Ahuru larvae were common in the study area and
...........Jir¡¡¡; were more abundant than the eggs, particularly in the
r -5
5-20 first year.In 1974-75,6800 were recorded, and in
1975-76, 9700. They were most numerous during
No. below lO m2 October and Nlovember (Fig. l6), with a marked peak
Fig. ll: Cumulative dist¡ibution of moray eel eggs, Ig?4-75
in November 1975, 2 weeks after the peak in egg
numbers. The relatively high numbers of larvae and
low numbers of eggs in 197+-75 indicated that
spawning either took place mainly outside the study
area (to the south) or before sampling began.
Family Congridae
Sand eel Gnathophis incognitus Castle, 1963 or G. The distribution of larvae (Fig. l7) shows distinct
habenatus (Richardson, I848) and puzzling differences from that of the eggs. In both
Eggs. The egg of Gnathoþhis sp. is described and years few larvae were found in the immediate vicinity
{igured in Castle and Robertson (1974) . Five eggs of of spawning. In 1974-75 larvae were commonest in
this type were taken in one sample on 22 January the eastern gulf, off the Coromandel Peninsula. In
1975; two were at the early blastodisc stage and three 1975-76 they were found both there and more widely
had well-developed embryos. spread over the central gulf.
Fig. 12: Moray eel Larva (9.6 mnr).
204-8 Nov 1974
ó - l0 Oct 1975 20 -24 Oct 1975 3 -7 lNov 1975
50-200 200-500 500- > r000
1000
No. below lO m'
,l975 -5 Dec 1975
l7-21 Nov 1
Fig. 13: Distribution of ahuru eggs by cruises, 197'[-75 and 1975-76
A possible expianation for the disjunctive egg and ' and the smallest larvae taken were about 4 mm long.
larval distributions is that the larvae were carried by Flowever, ahuru juveniles, some up to 45 mm long,
water movements from the spawning area and were were sometimes tallen and were the largest
not sampled until they reached a size large enough to ichthyoplankton caught during this survey. Adults
be retaiñed by the plankton net. The newly hatched seldom exceed 120 mm and are susPected to live off
larva from the small ahuru egg (0.525-O.600 mm the bottom. Graham (1956) found that ahuru fed
diameter) is unlikely to exceed 1.5 mm total length principally on a surface-living copepod.
2tFamily Exocoetidae
Garfish Hyþorhamþhus zår Phillips , Ig32
The early development of the garfish is described
and figured in Graham (1956). This species has
p
adhesive, demersal eggs.
I róã
:
r; 9
Larvae. Two larvae (measuring 10.6 and I 1.0 mm) þo
x o_
were taken in 1975, and one (meâsuring ll.3 mm) in E
1976. In both years they were taken in the northern $r 20P
o o
central gulf in late January. The heaviiy pigmenred o2 c,
garfish larva is distinctive (Fig. IB), with a prominent
18.
o o
! o
double row of rouncl pigment spots along its back. 5r ló=
The lower jaw projects beyond the upper. z
0 14
I 1529122610247214
Family Trachichthyidae Oct Nov Dec Jon Feb
Roughy (unidentified) Fig. 14: Numbers of ahuru eggs taken by cruises, 1974-75 and
1975-76. Mean sea temperatures in the spawning area are
Larvae. During December and January, larvae also shown.
(Fig. I9) identified to this farnily were taken
occasionally at the outer stations of the study area.
Fig. 15: Ahuru (Auchnoceros þunctatus) larvae (5.6, 12.8 mm).
22single juvenile ll.9 mmL lÒng was taken in the outer
gulf on 16 January 1976.
Family Syngnathidae
'o Small juveniles of the seahorse (Hippocampus
x abdominalis Lesson, l8l8) and the long-snouted
(¡)
o pipefish (Stigmatoþhora longirostris llutton, I 8 72) were
5 197s-76 occasionally recorded.
_9
o 4
(¡)
-o
E Family Triglidae
l
z Red gurnard Chelidonichthys kumu (Lesson and
Garnot, l826)
Eggs. The red gurnard egg is described and figured
in Mito (1963) and Robertson (1975). Red gurnard
eggs were taken on every cruise during both years, but
12 721 were never abundant. The cumulative egg distribu-
l15 26 1
tion was remarkably similar in both years (Fig. 20)
Oct Nov Dec Jú Feb
and showed the main spawning centre to be at the
Fig. 16: Numbers of ahuru larvae taken by cruises, 1974-75 and northern edge of the study area or still further to the
1975-76.
north. The seasonal variation in egg numbers for
197+-75 indicated a spring spawning peak in
They may have been larvae of the slender roughy October, but in 1975-76 peak egg numbers occurred
(Optiaus elongatus, (Günther, 1859)), the most common in December (Fig. 2t). However, in both years the sea
species of roughy observed by divers along rocky bottom temperatures in the spawning area were
coastlines in northern New Zealand (Doak 1972). similar at peak spawning (15.6'c in 1974-75 and
Most larvae were small (3.5-4.5 mm long). The 15.7'c in 1975-76). Ekler (1976) considered that the
Iargest was 8.3 mm and the skin on the dorsal surface time of peak spawning was influenced by temPera-
of its head and body was covered with tiny spines. ture, through its effect on the maturation rate of the
gonads.
Family Zeidae
John Dory Zeas japonicas (Richardson, IB45) Larvae. Early stages of the red gurnard larva are
The eggs of this species have'been described figured in Mito (1963) . The cumulative distrit¡utions
(Robertson 1975), but were not taken in this study. A of larvae (Fig. 22) showed a pattern of dispersal
2t-25
*ffi
æ 1971 l8 -22 Nov 1974
Fig. 17: Distribution of ahuru lawae by cruises, 1974-75 (see key on following page).
23{t z-ì /
li
\
I
)r-s.-- -----**
\ z
)\ lL
al\/^/L ¡r^^.-.t"!
vìqt\rJ
z---r\ / cr,-a r:J
>
^--"-" o
Y^""qnJ¿
/J\ * t'\
//\ r^r^ V
\t\ -J I
l/ ì.\ Y
"w
6-9 Oct1975 20 - 23 Oct 1975 3 -Z Nov 1975
20-s0 s0-200 200-500
No. below l0 m2
l7 -20 Nov 1975 1 -4 Dec 1975
Fig l7 (continued): Distribution of ahuru larvae by cruises, 1975-76.
southwards from the spawning area; this was more
marked ín 1974-75, perhaps because spawning began
earlier that year (see Fig. 2l). These findings concur
with those of Elder (1976), who postulated that larvae
drifted south a{ter â northward, contranatant
migration of adult red gurnard from the inner
Hauraki Gulf to a spawning ground in the Cradock
Fig. 18: GarÍish (Hyþorhamþhus ih) larua (1t.3 mm). and Colville Channels.
Family Acanthoclinidae
Rockfish Acanthoclinus quadridactylus (Bloch and
Schneider, l80l) and A. trilineatus (Griffin, 1933)
Jillett (1968) described the early life history of A.
quadridactylus, a small littoral fish which lays demersal
eggs. The adults of both A. quadridactylus and A.
trilineatus are described and figured in Griffin (1933).
Fig. 19: Roughy (Family Trachichthyidae) larva (5.3 mm).
2+r-5 5-20 >20
below l0 m2
î,1 ffi
Fig' 20: cumulative disrriburion or red gurnard eggs Lefr: rg74-75.
Righr: 1975-76
l8
o)
o)
o
ló U
o o
f
o
t4
E o
l o-
z r50 E
1975-76 o
Ê
100 r8 o
o
tó o
rn
Fig. 2.1: of red gurnard eggs taken by cruises, lg74-75
^Numbers
and 1975-76. Mean sea bottom temperaturés in the Ápawning
area are also shown.
rt5 1226t024721 4
14
Oct Nov Dec Jon Feb
r-5 5 -r0 r0-20 >20
No. below lO m2
\ '\ sï?/!-/-{-\,
Fig.22: Cumulative distribution of red gurnard larvae. Left: 1,974-75. Right: 1975-76
25
Inset 3Larvae. Small larvae oÍ A. quadridacQlus up to Family Carangidae
5.5 mm long are figured in Jillett (1968) . Lawae of the There are two species oÍ Trachurus (jack mackerel)
two species were not recorded separately during this found in New Zealand waters (Stephenson and
study. The illustrated specimen (Fig. 23) is A' Robertson 1977). Trachurus nouaezelandiae is abundant
trilineatus, which has more dorsal and anal fin rays in in-shore waters in northern New Zealand. Trachurus
than A. quadridactllus and a more slender body. The decliuis usually lives further off shore and is found
larvae are heavily pigmented with dark brown sPots' throughout New Zealand. The eggs of both species
are described in Robertson (1975), but this study and
recent findings of Robertson (pers. comm.) have
'l'he cumulative distribution for the 2 years shown these descriptions to be inaccurate.
(Fig. 2a) showed the larvae to be well dispersed' but
with the largest numbers usually recorded close to (Richardson,
land. Larvae were collected on all cruises in both Jack mackerel Trachurus noaaezelandiae
rB43)
years, except the làst cruise in 1974-75. Their
numbers declined steadily as the sea temPerature Eggs. The eggs of T. nouaezelandiae (: T. mccullochl
increased. More larvae were collected in 1975-76 than are described in Robertson (1975) as having a
in the previous year, which was warmer. These results diameter of 0.720-0.820 mm, a segmented yolk, and a
are consistent with Jillett's findings that A. single oil droplet. During this study eggs conforming
winter spawner, with peak spawning
quadridacQlus is a to this description were not taken, but an egg slightly
in September and October. (Jillett considered Sep- Iarger than this was regularly caught and was very
tember and October as winter.) abundant. The likely causes of this size discrepancy
Fig. 23: Rockfish Acanlhoclinus trilin¿alus
Iarva (10.0 mm).
3-4
â\* -- ------._,t
-
n( 10 m2
-'.f-Ð---''.
\*-'
\-v1. \
1
\i ,'
Fig 24: Cumularive distribution of rockfish laruae LeÍv 1974-75. Right: 1975-76.
26are that Robertson's measurements were of unfertil- The egg distribution (Fig. 26) showed the Hauraki
ised eggs which were probably not fully mature, and Gulf to be an important spawning area Íor T.
that a slight increase in size normally occurs at nouae4landiae. Most sparvning occurred in the western
fertilisation. side of the gulf, a few rniles off shore, and in depths
greater than 30 m. Spawning extended from Rakino
The eggs shown in Fig. 25 had a diameter of 0.78- Island in the south to Cape Rodney in the north, with
0.88 mm; a narrow perivitelline space; a segmented the densest patches of eggs east of Tiritiri Matangi
yolk (segmentation irregular and not always distinct, and Kawau Islands, the areas in which snapper eggs
in the early stages more noticeable just below the were also most abundant (Crossland 1980). In 1974-
blastodisc) (see Fig. 25 top); and a single, yellowish 75 there was another important spawning area off the
oil droplet, 0.204.27 rnrn in diameter, situated south-west shore of Great Barrier Island; this part of
anteriorly in late stage eggs. The embryo was stout the gulf was not sampled in t975-76.
bodied with numerous prominent melanophores
along the dorsal surface.
The seasonal changes in egg production are shown
These eggs were considered to be those of T. in Fig. 27. TÌr'e figures have been adjusted for
nouae4landiae for the following reasons:
temperature effect (as described for anchovy eggs)
and the numbers in the .t974-75 season are for a;trarea
They had the typical carangid egg features of
1.
equivalent to that sampled in 1975-76. The observed
segmented yolk; single, anterior oil droplet; and
egg production was similar in both years. peak
relatively undeveloped embryo at hatching.
spawning occurred in nrid November in 1g74, when
2. An embryological series was built up and the late the mean sea temperature was 17.3"c, and in mid
December in 1975 at a mean sea temperature of l7.9oc.
stage embryos were indistinguishable from yolk-sac
larvae of T. nouaelelandiae.
3; Measurements of 100 unfertilised eggs from a ripe Larvae. The larvae of. T. nouae¿elandiae (Fig. 28) are
specimen of T. gave the following
nouaezelandiae similar to those of. T. symmetreczs (Ahlstrom and Ball
results:.diameter 0.775-0.865 mm (mean 0.825 mm); 1954) from the north-eastern Pacific. They show the
oil droplet 0.191-0.250 mm diameter (mean distinctive carangid feature of three rows of
0.226 mm) (Robertson pers. comm.). melanophores on each side, one each along the dorsal
and ventral margins of the body and one along the
4. The eggs are not those of trevally (Caranx georgianus lateral line. They can readily be distinguished lrom
(Cuvier, 1833)), another carangid known to occur in the larvae of trevally because the latter are more
the Hauraki Gulf. Although of a similar size, the dlkly pigmented and have a deeper body (James
trevally egg has an unpigmented embryo (James
r s76).
r e76).
The distribution of larvae (Fig. 29) was similar for
both years. Larvae were found close to the eggs for the
corresponding period, but were displaced a few miles
to the north-east, further into the central gulf. Larval
numbers (FiS. 30) showed a distinct peak in
November of each year. In 1974 this coincided with
the peak in egg numbers, but in 1975 it occurred a
month before the egg peak. This could be the result of
a large spawning inside the study area between
sampling periods, or it could have resulted from a
spawning to the north of the study area, with the
larvae carried south lty currents (compare with
pilchard and gurnard).
Jack mackerel Trachurus decliais (Jenyns, 1841)
Eggs. Robertson's (1975) description gives the egg
diameter as l.l-1.3 mm. This was based on a poorly
preserved sample and is inaccurate. A more recent
Fig. 25: Jack macke¡el Trachurus nouac¿clandiae eggs. sample of 100 artificially fertilised eggs had a diameter
274-8 Nov 1974
20-23 Jo¡1975
gK4;^L
l\ r^.,^ tl
20-24 Oc¡ 1975 3 -Z Nov 1975
I -5 Dec l9Z5 l5 -19 Dec 1975 29 - 31 De< 1975
500- 2000- 5000- >t0000
2000 5000 10000
No. below l0 m'
Fie. of jack
- 26: Distribution
mackerel T. noaaez¿Iandia¿
eggs bY cruises, 1974-75
and 1975-76.
27 -3O )on 1976range of 0.970-1.030 mm (mean 0.998 mm) and a
segmented yolk with an oil droplet of 0.238-0.302 mm
(mean 0.266 mm) diameter (Robertson pers. comm.).
Eggs conforming to this description were occasionally
l8
recorded in the Hauraki Gulf. They ranged in size
=,o
I
c)
from 0.95 to 1.02 mm diameter and had an oil droplet
of 0.23-0.29 mm diameter. The late stage embryo was
lói more slender and muc.h more lightly pigmented than
x o
o
that of T. nouae4landiae (Fig. 3l). Cumulative egg
q)
o)
o distributions for the 2 years are shown in Fig. 32.
o E
CJ
o
o
{J Larvae. The larvae oÍ T. decliuis are probably so
-o c
)E similar to those of. T. nouaelelandiae that separation of
z 'rg the two species may only be possible by biochemical
means. Stephenson and Robertson (t977) could not
distinguish juveniles of the two species if they were
shorter than 35 mm standard length. Because the eggs
oL T. decliuis were only rarely recorded, and nearly all
adult jack mackerel observed in the Hauraki Gulf are
11529 1226 1024721 4 T. nouae4landiae, all Trachurus lar.'ae were assumed to
Oct Nov Dec Jon Feb
be T. nooaelelandiae.
Fig. 27: Number of jack mackerel T. noraezelandiaø eggs within
the study area at the tirre of sampling for each cruise, 197.f-
75 and 1975-76. Mean sea tempe¡atu¡es for each cruise a¡e
also shown.
Fig. 28: Jack mackerel Trachums noaa¿-
zelandiae larvae (4.4, 7.6, 8.9 mm).
29I
U\l
\¿
^ tz'-.-¡")
f-.r\/
o u4-4t.9
:.
21 - 25 Oct 1974 18 -22 Nov 1974
93"4
//\ r^\^ V
ll;:t:iitiiilli:tlrill
ii::rliiiiii:i:l:ri:ii
50-200 200-500 500- >t000
\t t000
r'------/
No. below lO m'1
0
i,Effi Fig. 29: Distribution of jack mackerel T. noaae4landiaelarvaeby
2Q-23 Jon1975 cruises, 197,S-75 and 1975-76.
/\ L
/-\/ d
!)v* l,rÞ- -J \
-ê^41ì/'¿ .
\t\ ) I
/ìÀÞ,v
/)\.a\\
aJi I'o
x
o
o
o
o
o
E
zl
Fig. 3l: Eggs of jack mackerel Trachwas decliuis.
Larvae. The early stage larva uP to a length of
3.2 mm is described and figured in Cassie (1956b) .
The patches of yellow pigment he observed in living
11529 1226 1024721 4
specimens were not aPParent in preserved samples,
Oct Nov Dec Jon Feb
and the absence of distinctive features in small
Fig. 30: Numbers of jack mackerel T. nooaezelandiae larvae taken snapper larvae made initial identification difficult.
by cruises, 197+-75 and 1975-76. Gradually a series was built up and it showed that the
most useful identifying character, before the develop-
ment of the fin ray pattern, was the Presence of two
Family Sparidae pigment spots on the ventral su¡face of the gut (Fig.
Snapper Chrysoþhrys auratus (Forster, l80l) 33 top). These larvae '*'ere confirmed to be snapper by
Eggs. The snapper egg is described and figured in electrophoretic analysis of their enzyme patterns and
Cassiè (1956b) and Crossland (1980). The distribu- comparison with those of the adult (Smith and
tion and abundance of snapper eggs in the Hauraki Crossland 1977). Lawae large enough to be identified
Gulf have been described previously (Crossland by their fin ray count (that is, Iarvae more than
1980). They were the most abundant of all egg tyPes. l0 mm long) were rarely taken.
::.:. .:
':,: ':r'''':
5 -20 20-50
No. below l0 m2
Fig.32: Cumulative distribution of jack mackerel T. decliais eggs. Left: 1974-75' fught: 1975-76'
3l' i.i'l'::;ïi;j;
'- I '. ',
¡,rí"
ç'
. .'"
-¿"
/.'rr'1.1
{ù^..''i
_ a :e
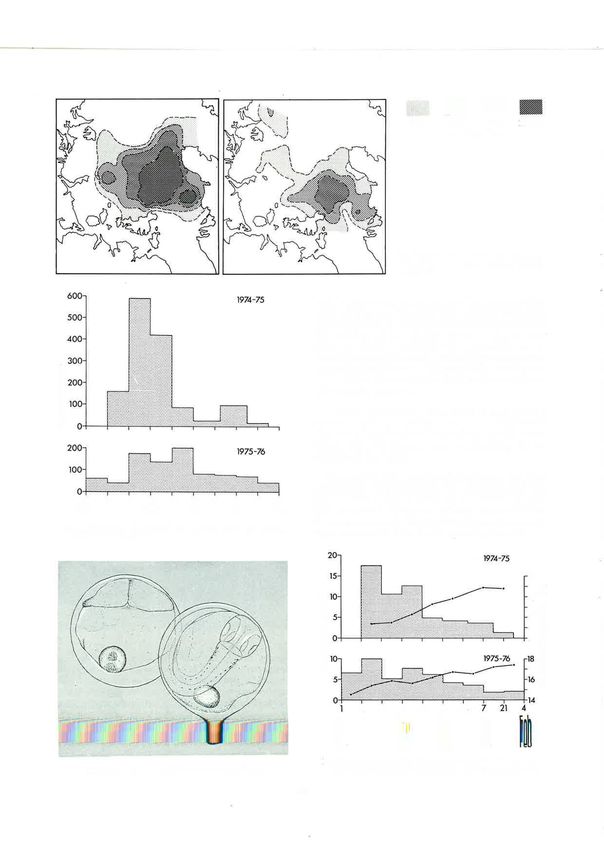
Fig. 33: Snapper (Chrysoþhrys auratus) la*ae (5.6, 9.7, 13.5 mm).
The distribution of snapper larvae was patchy in it was lower in 1975-76 than in
2 years indicated that
197+-75 (Fig. 34), aqd they were not abundant (Fig. the year before and that about the same number of
35). In I975-76 they were much more common, and eggs survived to hatch in each year.
more evenly distributed, mainly over the central gulf
(Fig. 3a). Consideration of these facts alone would How can the five times greater number of larvae in
l97l-70 be explainedl The relative scalci 0f
I
I
heavier in I975-76 th'an 1974-75, yet the opposite was snapper larvae (even in 1975-76) compared with the
true (Crossland 1980) . A study of egg mortality in the larvae of other species whose eggs were much less
32!t
t2
â\
L^l
L,'
/\ L
î54Ë
//\ -r^\- V
18 -22 Nov 1974
17- 20 Nov ì975 1-4 Dec 1975 ló - l9 Dec 1975
20-50 50-200 200-500 >500
No. below ]O m2
Fig. 34: Distribution of snapper laruae by cruises, 197.[-75 and 197Í176.o
'o
x
0)
o
!
o
ot
_o
E
l
z
11529 1226 l0 247 21 4
Oct Nov Dec Jon Feb
,\i
Fig. 35: Numbers of snapper lawae taken by cruises, t974-7 5
and 1975-76.
abundant (such as jack mackerel) indicates that the
snapper larva has a short pelagic phase. Cassie
(I956b) observed that newly hatched snapper larvae Fig. 36: Yellow-eyed mullet (Aldrichan forsten) egg.
preferred the bottom of their container.
the If
demersal habit is quickly adopted, the larvae collected
in the plankton may be mainly those displaced by small juveniles the pigmentation merges to form a
currents or tidal movements. Thus the chances of uniform, dark grey background.
sampling them will depend not only on their actual
abundance, but also on the effects of the prevailing Larvae, and juveniles up to 33 mm iong, were
weather or the state of the tide. One sample in recorded throughout the study area, but were taken
November 1975 taken off the south shore of Kawau only occasionally. The occurrence of juveniles during
Island, where tidal movements are.rnore pronounced the early part of the study period indicated that
than in open water, contained more laryae than the spawning also occurred during late winter and that
total for the whole of the previous year. This is also the yellow-eyed mullet has an extended breeding
the only locality in which Cassie (1956b) recorded season.
snapper larvae in any numbers.
Grey mullet Mugil æphaløs Linnaeus, 1758
A single juvenile I1.5 mm long was recorded in mid
Family Mugilidae January 1976. It was very similar to juvenile yellow-
Yellow-eyed mullet Aldrichetta forsteri eyed mullet, but could be distinguished by its having
(Valenciennes, 1836) only 8 anal soft rays (compared with 12 in the yellow-
eyed mullet); both species have 3 spinous rays.
Eggt. The egg of the yellow-eyed mullet is
described in Cassie (1956b). Eggs of this species were
taken in the inner gulf throughout the study period,
but never in large numbers. They ranged in size from
0.84 to 0.93 mm diameter. The number of oil droplets Family Labridae
varied from 3 to 13. Late stage eggs were never
recorded; most eggs were at the stage shown in Fig.
Spotty Pseudolabrus celidotus (Forster, l80l)
36. The cumulative egg distributions for the 2 years Eggs of this common littoral fish, which are
(Fig. 37) showed a spawning centre east of Tiritiri described in Robertson (1975), were not recorded
Matangi Island. Eggs were more abundant in 1974- during this study.
75 than in the following year.
Larvae. Larvae (Fig. 39) were recorded in small
numbers between October and the beginning of
Lônae, Lawae 0i this sPecies ate very dark' December and were three times as abundant in 1975-
I
because of rnany small melanophores over the whole
of the body and head (Fig. 3B). In older larvae and
345-20 20 -50
No. below l0 m2
Fig. 37: Cumulative distribution of yellow-eyed mullet eggs. Left: 1974-75. I{ight: 1975-76.
Fig. 38: Yellow-eyed mullet (Aldrichettø
forstcn) Iawa (10.5 mm).
Fig. 39: Spotty (Pseudolabrus celídotus)
larua (13.2 mm).
Family lJranoscopidae droplets, and in the late stages only a single,
Spotted stargazer Genyagnus monoþterygius (Bloch and posteriorly placed droplet remained (Fig. a0 bottom).
Schneider, l80l) The late stage embry¡o was spotted with numerous
melanophores; there were also some on the surface of
Eggs. The spotted stargazer egg is described in the yolk. The dorsal median fin fold was strongly
Robertson (1975) (diameter 1.800-2.000 mm; non- developed and began at the front of the head.
segmented yolk; three to nine oil droplets). Eggs with
a size range of 1.79-1.96 mm and multiple oil droplets Spotted stargazer eggs were recorded consistently
were recorded in both years (Fig. 40 top). During during Octobe¡ and lrTovember of each year, but in
sorting a developmental series of these eggs was built only small numbers; from December onwards they
up and it showed that as the egg matured the oil were rare. Their distribution (Fig. 4l ) was patchy and
droplets coalesced, so that by the middle stages (Fig. widespread, consistent with the occurrence of this
40 centre) there were only two or three larger species.
35Larvae. These were identified by building up a
developmental series (Fig. a2). The smallest larva
(top) was consistent with the most advanced embryo
(Fig. a0 bottom). The adult fish has a small barbel on
the chin, and this was observed in the largest larva
taken (Fig. 42 bottom), which also showed the
characteristic forward placement of the pelvic fins.
Although no specimen was developed far enough to
allow comparison of fin ray counts, biochemical tests
confirmed the identification. Electrophoretic tissue
analysis by*y colleague P. J. Smith on one of these
Iarvae and on adult spotted stargazer detected a
glucosephosphate isomerase (GPI) Iocus of similar
mobility in both. These loci are good species markers
because of electrophoretic variability of phenotypes
(Avise and Kitto 1973, Dando 1974).
The cumulative distribution of larvae is shown in
Fig. 43. They were less scattered than the eggs,
commonest in the central gulf during both years, and
taken at all times during the study period.
Families Blenniidae, Clinidae, and Triptery-
giidae
Blennies
Larvae of these ubiquitous families of fishes (the
adults of which lay demersal eggs) were a common
component of the ichthyoplankton. Most were small
(less than 8 mm), and no attempt was made to
identify them to species level.
Fig. 40: Spotted sta¡gazer (Genyagnus monoþtcrygius) eggs.
iiii: t ::::: äi!ii::::t!tl
r -5
5-20
No. below l0 m2
Fig. 4l: Cumulative distribution of spotted stargazer eggs' Left: 1974-75. Right: 1975-76.
36Fig.42: Spotted stargazer (Gnlagnus monoþterygius) larvae (5.6, 7.2, 13.8,20.0 mm).
3t1-2 2- 5
No. below l0 m2
Fig. 43: Cumulative distribution of sported stargazer larvae. Left: 197+-75. Right: 1975-76.
Family Eleotridae Family Scombridae
Graham's gudgeon Grahamichthys radiatus (Quoy and Blue mackerel Scomber australasicus Cuvier and
Gaimard, IB37) Valenciennes, 1831
Larvae. Larvae of this species (another which lays The eggs and larvae of this species were identified
demersal eggs) are shown in Fig. 44.Tlne young larva from their close similarity to those of the Pacific
(top) is transparent and has a conspicuous swim (Japanese) mackerel Scomber japonicus (:
bladder which becomes less prominent as the larva Pneumatoþhorus diego) (Kramer 1960).
matures (bottom). These larvae-were common and
were taken throughout the sampling period, being Eggs. These eggs (Fig. 46) had a diameter of I.05-
most abundant from late November onwards in 1974 1.20 mm; a narrow perivitelline space; a non-
and from early December in 1975. Their distribution segmented yolk; anda single oil droplet of 0.26-
was similar in both years (Fig. 45) and extended in a 0.33 mm diameter, situated posteriorly in late stage
broad band across the northern half of the studv area. eggs. The advanced embryo is moderately pigmented
dorsally and laterally, from the head almost to the tip
Family Gempylidae of the tail. A few melanophores also occur on the
Barracouta Thyrsites atun Euphrasen, l79l surface of the yolk near the embryo.
Ten specimens of larvae and pre-juveniles were
taken in the outer stations on the first cruise in 1974. BIue mackerel eggs were taken throughout the
They ranged in length from 7.5 to 25 mm. study period in 1974-75. Their abundance varied
Fig. .14: Graham's gudgeon (Gralary-
lchtþs radiatu) larvae (4.9,
12.2 mm),
38r-.5 5-t0 10-20
No. below l0 m'?
Fig. 45: cumulative dis¡¡ibution of G¡aham's gudgeon larvae. Lelt: lg74-75. -Right: 1975-76.
greatly from cruise to cruise (Fig. 47), which is to be top of the head and derLse pigmentation on rhe sides of
expected with this highly mobile schooling {ish. The the visceral cavity. These leatures distinguish them
spatial distribution (FiS. 48) showed the main from snapper larva€:, which they s.uperficially
spawning area to be outside the study a¡ea or on irs resemble in the early stages. Details of pigmentation
northern periphery. In 1975-76 this part of the gulf are remarkably similar ro rhose of the Pacific mackerel
was not sampled and blue mackerel eggs were less described by Kramer (1960). However, larvae of S.
common, being taken in large numbers only during australasicus are more slender and have less developed
one cruise. fin rays than S. jaþonints larvae of comparable length.
The largest larva recorded was 19 mm long.
Larvae. Blue mackerel larvae (FiS. 49) are
characterised by a patch of round meianophores on Blue mackerel were among the most abuñdant
Iarvae in plankton from the outer stations; on several
occasions over 1000 were taken in one sample, and a
maximum of 2704 tvas recorded. The seasonal
variation in larval numbers (Fig. 50) showed two
marked peaks in both seasons. I was not able to
determine whether these were caused by chance
during sampling or were a significant feature of the
spawning of this species. The distribution of larvae
(Fig. 5l) was similar in both years and was centred
close to the observed spawning areas, but dispersed
further south into the central gulf.
,o
x
o)
o)
(¡)
o
o
_o
E
l l'r529
z Oct 1226 1024721 4
Nov Dec Jon Feb
Fig. 46: Blue macke¡el (Scomber australasims) egg.
Fig.47: N_umbers of blue mackerel eggs taken by cruises,
r974-7 5.
39îÅffi
l8 - 22 Nov 1974
lrrr:rl:l:i:r.:
::r ¡::!:i*i:i::.¡i:i:i
50-200 200-500 500- >1000
1000
No. below l0 m'
16 - 20 Dec 1974 20-23 Jon1975
lZ-21 Nov 197 Fig. 48: Distribution of blue mackerel eggs by cruises, 1974-75 and 1975-76
Family Bothidae (Arnoglossus scapha (BIoch and Schneider, 1B0l)) (see
Crested flounder Loþhonectes gallus Gunther, 1880 Robertson 1975 for eggs and Robertson 1973 for
larvae).
The eggs of this species have not previously been
described from New Zealand, and the larvae have
been described only in an unpublished account Eggs. These eggs (Fig. 52) had a diameter of 0.81-
(Robertson 1973). Eggs and larvae were identilied by 0.88mm; a non-segmented yolk; and a single oil
and raryar series' rh1 droolet of 0.I2-0.16 mm diameter. Crested flounder
fllllülllf l¡ ft[ IlllfIÎt'¡nut lll
closely resemble those of the related species witch eggs were taken at the outer stations on every crulse In
40Fig. 49: Blue mackerel (Seomber
australasicus) larvae (9 4, I4.4 mm).
or 1974-75 bothid larvae taken in this study were positively
identified as those oÍ L. gallus by their fin ray counts.
8-l
Loþhonectes gallushas 87 to 93 dorsal rays and 7l to 77
anal rays; A. scaþha has ltl to 126 and 87 to 99
respectively (Norman 1934). The largest lal.va
'o recorded was 14 mm long, at which size metamor-
)( phosis had not begun.
(¡)
o
o
IJnlike the eggs, Iarvae of crested flounder were at
times abundant, particularly in 1975-76, when as
o
many as 152 were recorded in a sample. The larval
(¡)
-o distribution (Fig. 55) shows dispersion south into the
z)
E
central gulf from the northern spawning centre. This
was more marked in 1975-76, when larvae were
common as far south as Tiritiri Matangi Island.
Although larvae were taken on all cruises in both
years, their numbers varied greatly, and periods of
abundance did not correspond in the 2 years. In
1974-75larvae were common on the first three cruises
11529 1226r0 24721 4
(October and November); in 1975-76 they were rare
Oct Nov Dec Jon Feb
on the first three cruises, suddenly became abundant
Fig. 50: Numbers of blue mackerel larvae taken by cruises, on the fourth cruise (mid November), and continued
1974-75 and 1975-76. to be so until the end of December.
The thin, Ieaf-life form of the crested flounder larva
1974-75, but were never common excePt at one indicates that it is a weak swimmer and is probably
station off the south-west shore of Great Barrier more truly planktonic than many other fish larvae and
Island (Fig. 53). The spawning area aPPears to be thus more affected by water movements. This, with its
north of the study area. Iong larval period, means that it can occur in samples
taken a considerable distance from the spawning site.
Larvae. The ybung larva (Fig. 54 top) is quickly
recognisable by a prominent band of black pigment
near the tail and by the bifid whisker on top of the
head. (Sometimes only a single ray was present, Family Pleuronectidae
possibly as a result of damage.) Larger larvae are Three members of this family are commonly found
transparent and every detail of the viscera and in the Hauraki Gulf: the sand flounder (Rhombosolea
structure of the gills is visible. They also have the bifid þlebeia Richardson, 1843), the yellow-belly flounder
whisker. These features also apply to the larvae of the (R. leporina Günther, lB73), and the speckled sole
more southerly distributed A. scaþha. ÉIowever, the (Peltorhamþhus latus James, 1972). The development of
+tYou can also read

















































