INVOLVEMENT OF THE HISTONE ACETYLTRANSFERASE ATHAC1 IN THE REGULATION OF FLOWERING TIME VIA REPRESSION OF FLOWERING LOCUS C IN ARABIDOPSIS1 W OA
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Involvement of the Histone Acetyltransferase AtHAC1 in
the Regulation of Flowering Time via Repression of
FLOWERING LOCUS C in Arabidopsis1[W][OA]
WeiWei Deng2, ChunYan Liu2, YanXi Pei, Xian Deng, LiFang Niu, and XiaoFeng Cao*
State Key Laboratory of Plant Genomics and Center for Plant Gene Research, Institute of Genetics and
Developmental Biology, Chinese Academy of Sciences, Beijing, China 100101 (W.D., C.L., Y.P., X.D., L.N.,
X.C.); Graduate School, Chinese Academy of Sciences, Beijing, China 100039 (W.D., L.N.); and College of
Life Science and Technology, Shanxi University, Taiyuan, China 030006 (Y.P.)
Histone acetylation is an important posttranslational modification correlated with gene activation. In Arabidopsis (Arabidopsis
thaliana), the histone acetyltransferase AtHAC1 is homologous to animal p300/CREB (cAMP-responsive element-binding
protein)-binding proteins, which are the main histone acetyltransferases participating in many physiological processes, in-
cluding proliferation, differentiation, and apoptosis. The functions of p300/CREB-binding proteins in animals are well char-
acterized, whereas little is known about the roles of AtHAC1 in developmental control in Arabidopsis. Lesions in AtHAC1
caused pleiotropic developmental defects, including delayed flowering, a shortened primary root, and partially reduced fer-
tility. Analysis of the molecular basis of late flowering in hac1 mutants showed that the hac1 plants respond normally to day
length, gibberellic acid treatment, and vernalization. Furthermore, the expression level of the flowering repressor FLOWERING
LOCUS C (FLC) is increased in hac1 mutants, indicating that the late-flowering phenotype of hac1 mutants is mediated by FLC.
Since histone acetylation is usually associated with the activation of gene expression, histone modifications of FLC chromatin
are not affected by mutations in HAC1 and expression levels of all known autonomous pathway genes are unchanged in hac1
plants, we propose that HAC1 affects flowering time by epigenetic modification of factors upstream of FLC.
Posttranslational modifications of histone N termini p53, HIV-1 Tat protein, and Stat3 (Sterner and Berger,
play fundamental roles in maintaining chromatin 2000; Liu et al., 2004). Therefore, p300/CBP are not
structure and function. One such modification, histone only HATs, but also factor acetyltransferases, and both
acetylation, has been implicated in many biological types of activity are physiologically important for p300/
processes. In humans, histone acetylation is catalyzed CBP functions (Sterner and Berger, 2000). In addition,
by distinct histone acetyltransferase (HAT) families because of their transcriptional coactivator functions,
including the p300/CREB (cAMP-responsive element- p300/CBP are proposed to contribute to transcriptional
binding protein)-binding protein (CBP) family, the activation by directly interacting with DNA-binding
TAFII-250 family, the MYST (MOZ, Ybf2/Sas3, Sas2, transcription factors and/or being part of coactivation
and Tip60) family, and the GNAT (GCN5-related complexes (Dai et al., 1996; Goldman et al., 1997; Seo
N-terminal acetyltransferase) family. The p300/CBP et al., 2001). In general, the histone acetylation status
family plays a major role in transcriptional regulation of a gene correlates with its transcriptional activity
by promoting acetylation of both histones and nonhis- (Cheung et al., 2000; Strahl and Allis, 2000).
tone proteins, such as the transcription factors CREB, The p300/CBP family exists in most multicellular eu-
karyotes, including mammals, Drosophila melanogaster,
1
This work was supported by National Basic Research Program and Arabidopsis (Arabidopsis thaliana; Sterner and
of China (grant no. 2005CB522400 to X.C.), by National Natural Berger, 2000; Pandey et al., 2002). The essential roles
Science Foundation of China (grant no. 30571032 to C.L., and nos. for p300/CBP in early development have been dem-
30325015, 30430410, and 30621001 to X.C.), and by the Chinese onstrated in human patients, mice, and Drosophila.
Academy of Sciences (grant no. CXTD–S2005–2). For example, mutations within the human CBP cause
2
These authors contributed equally to the paper. the Rubinstein-Taybi syndrome, which is character-
* Corresponding author; e-mail xfcao@genetics.ac.cn; fax 86–10– ized by severe facial abnormalities, broad thumbs,
64873428. broad big toes, mental retardation, and abnormal reti-
The author responsible for distribution of materials integral to the nal development (Roelfsema et al., 2005). In mice, both
findings presented in this article in accordance with the policy
described in the Instructions for Authors (www.plantphysiol.org) is:
p300 and CBP null mice exhibit embryonic lethality
XiaoFeng Cao (xfcao@genetics.ac.cn). between 8.5 and 11.5 d of gestation (Yao et al., 1998;
[W]
The online version of this article contains Web-only data. Roth et al., 2001). In addition to the essential roles in
[OA]
Open Access articles can be viewed online without a sub- normal development, mutation of one allele of either
scription. CBP or p300 affects embryogenesis and pattern forma-
www.plantphysiol.org/cgi/doi/10.1104/pp.106.095521 tion, indicating a requirement for proper gene dosage
1660 Plant Physiology, April 2007, Vol. 143, pp. 1660–1668, www.plantphysiol.org Ó 2007 American Society of Plant Biologists
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Histone Acetyltransferase AtHAC1 Regulates Flowering Time
et al., 2002; Yuan and Giordano, 2002), the functions
of AtHACs in gene regulation and developmental
control in Arabidopsis remain unknown.
In this study, we isolated Arabidopsis mutants with
T-DNA insertions in the HAC1 gene and investigated
their effects on plant development. We showed here
that HAC1 played an important role in vegetative and
reproductive development, and lesions in HAC1
caused a late-flowering phenotype in Arabidopsis.
hac1 mutants responded normally to day length, gib-
berellin (GA), and vernalization treatments, and dis-
played an increased level of transcripts of FLOWERING
LOCUS C (FLC) and two additional MADS-box genes,
MADS AFFECTING FLOWERING 4 (MAF4) and MAF5.
Therefore, HAC1 is critical for the normal regulation of
flowering time in Arabidopsis.
RESULTS
Isolation and Molecular Characterization of
hac1 Mutants
To investigate the roles of the CBP homologs in
Arabidopsis (AtHAC1, AtHAC2, AtHAC4, AtHAC5,
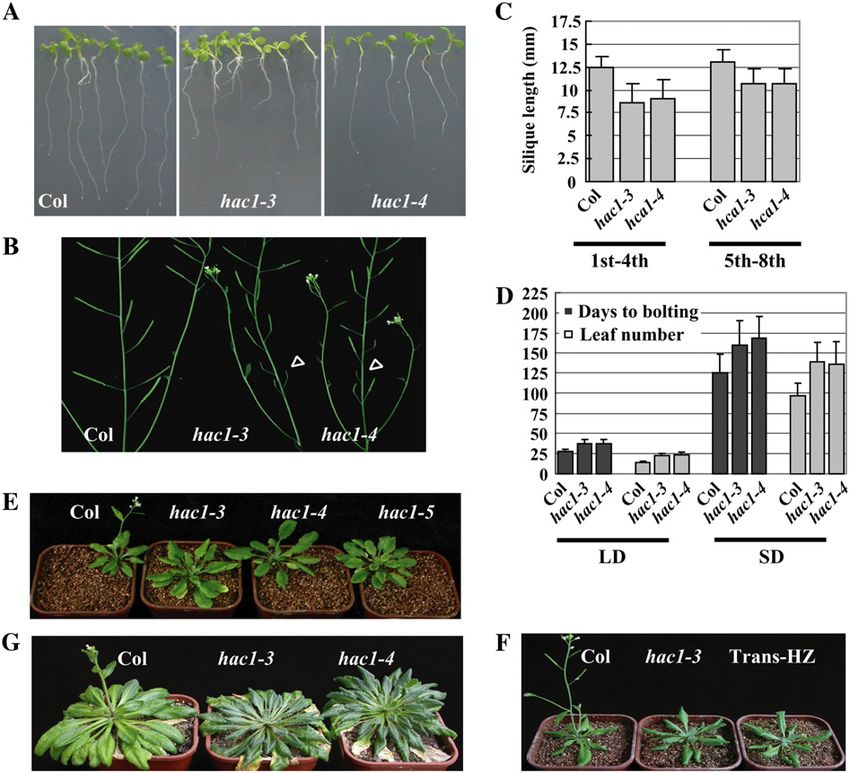
Figure 1. HAC1 gene structure, T-DNA insertion alleles, and mRNA and AtHAC12), T-DNA insertion mutants from the
expression. A, Structure of the HAC1 gene. The HAC1 gene consists of SALK or GABI-Kat collection were screened for ho-
17 exons. Exons were presented as black boxes, introns as black lines, mozygotes. No obvious developmental defects were
and 3# untranslated regions as gray boxes. The locations of hac1-3, observed from single mutants of hac2, hac4, hac5, or
hac1-4, and hac1-5 alleles were shown as triangles either above (from hac12 under normal growth conditions. However, three
SALK collection) or below (from GABI) the gene. B, Domains of HAC1. different T-DNA insertions in HAC1 (named hac1-3
Domains of TAZ, ZZ, and HAT were marked as predicted by Pandey for SALK_080380, hac1-4 for SALK_082118, and hac1-5
(Pandey et al., 2002). The T-DNA insertion sites on the genomic for GABI-314B12) showed a late-flowering phenotype.
sequences of HAC1 were marked on the corresponding positions of the
Therefore, we focused our investigations on the func-
translated protein with their allele number. aa, Amino acid. C and D,
HAC1 gene expression. HAC1 mRNA accumulation in seedlings was
tions of HAC1 gene.
sequentially analyzed by northern blot using the full-length CDS (C) The HAC1 gene contains 17 exons and encodes a
and the C terminus of HAC1 (D) as shown in Fig. 1B. The membrane 1,691 amino acid polypeptide with distinct domains
was stripped to remove the probe before blotting with another probe. including two ZZ-type and two TAZ-type zinc finger
The same membrane was also probed with an FLC probe and an Actin domains (Fig. 1, A and B). These domains have been
probe (see Fig. 4A). shown in other organisms to mediate protein-protein
interactions with transcription factors (Ponting et al.,
1996). Like CBP in animals, HAC1 also has a catalytic
(Yao et al., 1998). In Drosophila, there is only one domain (HAT domain) important for acetyltransferase
p300/CBP gene and the reduction of this activity alters activity. Both the HAT domain and zinc finger do-
gene expression such that multiple developmental mains are located in the C terminus (Pandey et al.,
processes are affected, leading to pleiotropic pheno- 2002). The location of the T-DNA insertions is up-
types (Lilja et al., 2003). stream of these conserved domains, in either the fifth
In Arabidopsis, there are five p300/CBP HAT ho- exon (hac1-3) or the sixth exon (hac1-4 and hac1-5). The
mologs, named AtHAC1 (or PCAT2), AtHAC2 (or location of each insertion was confirmed by PCR and
PCAT1), AtHAC4 (or PCAT3), AtHAC5 (or PCAT4), DNA sequencing of the junction regions. Since both
and AtHAC12 (Bordoli et al., 2001; Pandey et al., 2002; hac1-4 and hac1-5 contain the T-DNA at a similar posi-
Yuan and Giordano, 2002). All the AtHACs contain tion and show identical phenotypes, we used hac1-3
ZZ-type and TAZ-type zinc finger domains and a Cys- and hac1-4 for the rest of the analysis.
rich HAT domain at the C termini. Both ZZ-type and RNA-blot analysis was performed to confirm the
TAZ-type zinc finger domains have been implicated in efficient disruption of HAC1 mRNA in hac1 plants.
protein-protein interactions with transcription factors When full-length coding sequence (CDS) of HAC1 was
(Ponting et al., 1996) and the HAT domain of AtHAC1 used as a probe, we did not detect the full-length
confers HAT activity in vitro (Bordoli et al., 2001). transcript of HAC1 in hac1 mutants. However, a short
Although the structural conservation and diversifica- transcript was observed in both hac1-3 and hac1-4
tion of AtHACs have been characterized (Pandey plants, indicating that neither hac1-3 nor hac1-4 was a
Plant Physiol. Vol. 143, 2007 1661
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Deng et al.
null allele (Fig. 1C). Because the main functional do- those of the wild-type plants when measured after 6 d
mains of HAC1 were located in the C terminus, we of growth (Fig. 2A). The hac1 mutants also had re-
performed RNA-blot analysis using a probe specific to duced fertility in the first few siliques (Fig. 2, B and C).
the C-terminal portion of the HAC1 gene (Fig. 1B) and In the partial sterile flowers, the pollen produced by
no mRNA containing these conserved domains could hac1 mutants didn’t reach the stigmas due to the short
be detected (Fig. 1D). Thus, our mutant alleles are stamens resulting in the reduction of pollination and
unlikely to produce any functional protein. fertility (data not shown). The first 1 to 4 siliques of
A backcross between the hac1 homozygote and a hac1 were about 30% shorter than those of wild-type
wild-type line resulted in wild-type phenotype in F1 plants. Siliques 5 to 8 were longer than the first 1 to 4,
plants. The self-pollinated F1 progeny displayed an but still shorter than those of wild type (Fig. 2C). Si-
approximately 3:1 ratio of wild-type and hac1 pheno- liques formed after these were similar to those of wild-
type, indicating that hac1 is a recessive, single-gene type plants.
mutation. In addition, different alleles of hac1 (hac1-3, The most notable phenotype of hac1 plants is late
hac1-4, and hac1-5) do not complement each other. flowering (Fig. 2, D–G). In Arabidopsis, flowering time
For example, the hac1-3/hac1-4 trans-heterozygous F1 is regulated by four major pathways, namely the pho-
plants from a cross between homozygous hac1-3 and toperiod, GA, vernalization, and autonomous path-
hac1-4 showed the same phenotype as hac1-3 and hac1-4 ways (Simpson et al., 1999; Reeves and Coupland,
plants (Fig. 2, E and F), and no segregation was ob- 2000; Mouradov et al., 2002; Komeda, 2004; Amasino,
served in the F2 progeny (data not shown). These data 2005). To investigate the role of HAC1 in the regulation
indicate that the hac1 mutant is recessive and the of the flowering time, we first tested whether the
observed phenotypes of hac1 mutants are indeed caused photoperiod pathway was affected in hac1 mutants. By
by disrupting HAC1 activity. comparing the flowering time of the wild-type acces-
sion Columbia (Col-0) with that of hac1 plants, we
The hac1 Mutants Display Pleiotropic Developmental found that hac1 plants flowered later than wild type
Defects in Arabidopsis under both long day (LD) and short day (SD) when the
days to bolting (from germination to floral bud for-
In addition to the late-flowering phenotype, we mation) and the total leaf numbers at flowering were
examined other developmental defects in hac1 plants. measured (Fig. 2, D, E, and G). We observed only eight
The primary roots of hac1 mutants were shorter than hac1-3 and six hac1-4 plants flowering by 200 d under
Figure 2. Phenotypes of hac1 mutants.
A, The primary roots of wild-type and
hac1 plants grown on Murashige and
Skoog medium for 6 d. B, The siliques
from the primary stem of wild-type and
hac1 plants. The triangles indicate the
shortened siliques in hac1 plants. C, Sta-
tistical analysis of the first to fourth and
fifth to eighth siliques length as indicated.
Bars represent the SD of the silique length.
For each line, 20 plants were scored. D,
Flowering time of wild-type and hac1
mutants under LD and SD. The flowering
time was measured by days to bolting
(black boxes) and by total leaf numbers
(gray boxes) at flowering. Bars represent
the SD. For each line, at least 50 plants
were scored except for eight hac1-3 and
six hac1-4 under SD, respectively. E and F,
Wild-type (Col) and hac1 plants grown
under LD. Trans-HZ indicates F1 hac1-3/
hac1-4 trans-heterozygous plants from a
cross between hac1-3 and hac1-4. G,
Wild-type Col and hac1 mutants grown
under SD.
1662 Plant Physiol. Vol. 143, 2007
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Histone Acetyltransferase AtHAC1 Regulates Flowering Time
SD and the rest of the hac1 plants had not yet flowered. flowered almost at the same time (Fig. 3B). Therefore,
The experiment was terminated at this point. Since the hac1 lesions do not affect the vernalization pathway.
hac1 plants responded normally to day length, the late-
flowering phenotype of hac1 plants is not mediated by
HAC1 Controls Flowering Time through the
the photoperiod pathway.
Autonomous Pathway
The Late-Flowering Phenotype of hac1 Plants Is Rescued The autonomous and vernalization pathways con-
by GA and Vernalization Treatments verge on FLC, a MADS-box transcription factor, which
is a central regulator of floral transition in Arabidopsis
The growth regulator GA promotes flowering of (Michaels and Amasino, 1999; Sheldon et al., 1999). To
Arabidopsis. Exogenous application of GA accelerates examine whether the expression of FLC was increased
flowering in wild-type Arabidopsis, particularly un- in hac1 mutants, RNA gel-blot analysis was performed
der SD (Wilson et al., 1992; Blazquez et al., 1998). To using the FLC full-length CDS as a probe. In both
evaluate the effect of GA on the flowering time of hac1 hac1-3 and hac1-4 plants, FLC mRNA expression levels
plants, we sprayed the plants with a 100 mM GA solu- are increased (Fig. 4A). In Arabidopsis, there are five
tion once a week under SD until they flowered. Like MADS-box genes highly related to FLC, named MAF1
the wild-type controls, hac1 mutants showed acceler- to MAF5, respectively (Alvarez-Buylla et al., 2000;
ation of flowering by GA treatment (Fig. 3A). Ratcliffe et al., 2001, 2003; Scortecci et al., 2001, 2003).
Vernalization is a process by which flowering is pro- To test whether MAFs were also affected by lesions in
moted by prolonged exposure to the cold of a typical HAC1, we employed reverse transcription (RT)-PCR to
winter. Mutants in the vernalization pathway no lon- examine mRNA levels. Like FLC, the expression levels
ger respond to prolonged cold to promote flowering of MAF4 and MAF5 were increased in hac1 plants,
(Gendall et al., 2001; Amasino, 2004; Bastow et al., while MAF1-3 expression levels were unchanged when
2004; Sung and Amasino, 2004b; Amasino, 2005; Kim grown under LD (Fig. 4B, left section). We quantified
et al., 2005; Sung and Amasino, 2005). To determine this effect using real-time PCR and found a 2- to 4-fold
whether vernalization could reverse the late-flowering increase in mRNA levels of MAF4 and MAF5 in hac1
phenotype of hac1, the hac1 mutants and wild-type mutants (Fig. 4B, right section).
plants were grown at 4°C for 6 or 10 weeks before Since hac1 mutants are late flowering under both LD
transferring to 23°C under LD. Similar to the wild-type and SD, GA and vernalization treatments reverse the
plants, the hac1 mutants flowered much earlier after late-flowering phenotype, and hac1 mutants have an
vernalization treatment (Fig. 3B). After vernalization increased level of FLC mRNA, HAC1 behaves as a
for 10 weeks, hac1 mutants and wild-type plants member of the autonomous pathway.
Figure 3. Flowering time of wild-type and hac1
plants grown under different conditions. A, Effects
of GA on flowering time. Bars represent the SD.
For each wild-type (white box), hac1-3 (gray box),
and hac1-4 (black box) line, 50 plants were
scored except for the GA untreated control. In
our assay, only seven hac1-3 and six hac1-4
plants sprayed with ethanol flowered at 200 d
under SD and the experiment was terminated at
this point. B, Vernalization effects. Wild-type and
hac1 plants were grown under LD without ver-
nalization (untreated) or after vernalization for
6 (V42d) or 10 weeks (V70d). Flowering time was
measured by days to bolting (left section, black
box) and by leaf numbers (right section) at flower-
ing. Gray boxes represent cauline leaf number
and white boxes represent rosette leaf number.
Bars indicate the SD. At least 50 plants for each
genotype were scored.
Plant Physiol. Vol. 143, 2007 1663
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Deng et al.
Figure 4. FLC and autonomous pathway
gene expression in wild-type and hac1
plants. A, The levels of FLC mRNA in
wild-type and hac1 plants were deter-
mined by northern blot using FLC full-
length CDS probe (top section). Actin
gene was used as a constitutive expres-
sion control (bottom section). B and C,
The expression levels of MAF1-5 (B) and
autonomous pathways regulators of
flowering time (C) in wild-type and
hac1 plants were analyzed by RT-PCR
and quantitated by real-time PCR. UBQ
gene was used as a control for constitu-
tive expression. a and b represent differ-
ent amplification cycles in RT-PCR. Bars
represent the SD. 22DCT corresponds to
the ratio of each gene expression ver-
sus UBQ.
The components of the autonomous pathway, in- the late-flowering phenotype of hac1 was mainly FLC
cluding FCA (Macknight et al., 1997; Page et al., 1999; dependent.
Quesada et al., 2003), FPA (Schomburg et al., 2001), Epigenetic modifications have been shown to play
FLK (Lim et al., 2004; Mockler et al., 2004), FVE (Ausin an essential role in regulating FLC expression. Two
et al., 2004; Kim et al., 2004), FLD (Sanda and Amasino, components of the autonomous pathway, FLD and
1996; Yang and Chou, 1999; He et al., 2003), LD (Lee FVE, negatively regulate FLC by participating in the
et al., 1994; Aukerman et al., 1999), FY (Simpson et al., deacetylation of chromatin (He et al., 2003; Ausin et al.,
2003; Henderson et al., 2005), and RELATIVE OF 2004; Kim et al., 2004). The vernalization pathway also
EARLY FLOWERING 6 (REF6; Noh et al., 2004), pro- regulates FLC expression by chromatin acetylation and
mote flowering by repressing FLC. If HAC1 was es- methylation (Bastow et al., 2004; Finnegan et al., 2004;
sential for the expression of one of these repressors of He et al., 2004; Sung and Amasino, 2004a, 2004b;
FLC, hac1 mutants would be predicted to reduce the Amasino, 2005; He and Amasino, 2005). Furthermore,
expression of the repressor leading to an increase in REF6, which encodes a nuclear protein with jumonji
FLC transcription. We tested this hypothesis using RT- and zinc finger domains, represses FLC transcription
PCR and real-time PCR to compare the mRNA levels through histone deacetylation (Noh et al., 2004). Thus,
of FVE, FLD, FCA, FPA, LD, FY, FLK, and REF6 histone acetylation is an essential modification of FLC
between wild-type and hac1 plants. The mRNA levels chromatin.
of all known autonomous genes were not significantly Although it is clear that histone acetylation/deacet-
affected in hac1 plants (Fig. 4C). ylation regulates FLC expression, the identity of the
HAT responsible for this process is unknown. To in-
vestigate whether HAC1 acetylates histones at FLC,
The Late-Flowering Phenotype of hac1 Is Mainly we performed chromatin immunoprecipitation assays
FLC Dependent using 12-d-old seedlings of hac1 mutant and wild-type
plants. In addition to histone acetylation, di- and
To determine whether the late-flowering phenotype trimethylation of H3K4, which are hallmarks for active
of hac1 plants was solely due to the increase of FLC chromatin, were also analyzed (Bernstein et al., 2002;
expression, hac1 mutants were crossed with an flc null Alvarez-Venegas and Avramova, 2005). However, our
allele, flc-3 (Michaels and Amasino, 1999). If the effect chromatin immunoprecipitation experiments did not
of hac1 on flowering time was fully due to the up- show significant changes of histone H3 acetylation
regulation of FLC expression, the late-flowering phe- or H3K4 methylation in regions A, B, or C at the FLC
notype of hac1 mutants would be entirely suppressed locus, suggesting that increased FLC expression in hac1
by flc null mutant. Under LD, the flowering time of mutants was not associated with changes in these
both hac1-3/flc-3 and hac1-4/flc-3 double mutants was epigenetic modifications at the assayed regions (Sup-
earlier than that of hac1 plants (Fig. 5), indicating that plemental Fig. S1).
1664 Plant Physiol. Vol. 143, 2007
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Histone Acetyltransferase AtHAC1 Regulates Flowering Time
tants could be due to other floral repressors because
the flowering time of hac1/flc-3 double-mutant plants
was still slightly later than that of flc-3 under LD (Fig.
5). Consistent with this hypothesis, the expression of
two additional MADS-box genes in the FLC clade,
MAF4 and MAF5, were also up-regulated in hac1 mu-
tants (Fig. 4B). However, the up-regulation of MAF4
and MAF5 was not unique in hac1 mutants because
some autonomous pathway mutants also resulted in
increased expression of MAF4 and MAF5 (Supple-
mental Fig. S2).
In addition to the late-flowering phenotypes, muta-
tions in HAC1 caused pleiotropic developmental de-
fects, including shortened primary roots and partially
reduced fertility. These pleiotropic effects observed in
hac1 may be caused by loss of acetylation at multi-
ple target genes, similar to mutations in other genes
responsible for chromatin modifications. The Arabi-
dopsis genome encodes five HACs, whereas the num-
ber of p300/CBP proteins in animals is only one to two
(Pandey et al., 2002). In Arabidopsis, HAC1, HAC4,
HAC5, and HAC12 are highly related and could be
functionally redundant, which may explain why the
mutations in humans and mice p300/CBP are much
more severe than that of the hac1 mutation in Arabi-
dopsis. This hypothesis is supported by a recent study
in which a much stronger flowering phenotype was
observed with two different hac double mutants, hac1/
hac5 and hac1/hac12 (Han et al., 2007).
In humans, the transcriptional regulation by p300/
CBP appears to be exerted through multiple mecha-
nisms. p300/CBP have been shown to acetylate both
Figure 5. FLC-dependent late flowering caused by lesions in HAC1. nucleosomal histones and certain sequence-specific
Flowering time of hac1-3/flc-3 (top section) and hac1-4/flc-3 (bottom transcription factors, leading to transcriptional activa-
section) double mutants. Wild-type (Col), flc-3, hac1/flc double-mutant
tion in most cases (Sterner and Berger, 2000; Liu et al.,
and hac1 single-mutant plants were grown under LD. LN means leaf
number. The flowering time was measured by days to bolting (black
2004). Furthermore, as transcriptional coactivators,
box), cauline leaf number (gray box), and rosette leaf number (white p300/CBP are proposed to contribute to transcriptional
box) at bolting. Bars represent the SD. For each line, at least 50 plants activity by directly interacting with transcription fac-
were scored. tors and/or being part of the coactivator complex
(Dai et al., 1996; Goldman et al., 1997; Seo et al., 2001).
A variety of transcription factors have been demon-
DISCUSSION strated to interact with the bromodomain, HAT,
KIX, and zinc finger domains of p300/CBP (Vo and
Histone acetylation is known to regulate flowering Goodman, 2001). It is possible that HAC1 is also a
time in Arabidopsis (Tian and Chen, 2001; He et al., factor acetyltransferase and that this contributes to the
2003; Ausin et al., 2004; Tian et al., 2005). However, the regulation of flowering time.
HATs involved have not been identified. Our studies In general, histone acetylation correlates with gene
provide a link between a HAT, HAC1, and flowering- activation and when HAC1 is disrupted, one would
time control in Arabidopsis. The late-flowering phe- expect that the expression of its direct target genes
notype of hac1 mutants was FLC dependent. First, hac1 would be reduced. In hac1 mutants, the expression
plants flowered later than Col under LD and SD, levels of FLC, MAF4, and MAF5 are increased (Fig. 4, A
which is a characteristic of FLC repressing flowering and B). Therefore, FLC, MAF4, and MAF5 may not be
(Fig. 2, D–G). Second, hac1 mutants had a proper ver- the direct targets of HAC1. Instead, HAC1 might
nalization response that promotes flowering by re- regulate the expression of a trans-acting repressor of
pressing the expression of FLC (Fig. 3B). Third, the FLC, MAF4, and MAF5. Since the vernalization and
late-flowering phenotype of hac1 mutants was sup- autonomous pathway genes repress the expression of
pressed by an flc null mutant (Fig. 5). Finally, the FLC, and vernalization rescues the late-flowering phe-
mRNA levels of FLC were increased in hac1 mutants notype of hac1 mutants, it is unlikely that the targets
(Fig. 4A). In addition to the increase of FLC expression, of HAC1 are in the vernalization pathway. Therefore,
a portion of the late-flowering phenotype of hac1 mu- we examined the mRNA expression of all known
Plant Physiol. Vol. 143, 2007 1665
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Deng et al.
autonomous pathway genes by RT-PCR and real-time For RNA gel-blot analysis, RNA was transferred onto N1 nylon mem-
brane and probed with [a-32P]dCTP-labeled FLC full-length CDS fragment
PCR, but did not find any significant differences in (NM_121052, from 110–700 bp), or HAC1 full-length CDS (NM_106550, from
hac1 plants (Fig. 4C). One possible explanation is that 1–5,076 bp) or C-terminal fragments from 1,873 to 5,076 bp, respectively. Blots
HAC1 acetylates the histones of an unknown compo- were also probed with an Actin probe as a loading control.
nent that represses FLC and/or MAF4/5. Alterna-
tively, HAC1 might acetylate a known or an unknown Chromatin Immunoprecipitation
component of the autonomous pathway to affect its
activity at posttranslational level. A third possibility is The chromatin immunoprecipitation experiments were performed as de-
scribed by Johnson et al. (2002) using 12-d-old seedlings. Anti-acetyl-histone
that the direct target of HAC1 (or HAC1 itself) might H3 (AcH3; Upstate Biotechnology, 06-599), anti-dimethyl-histone H3 (H3K4m2;
be involved in the recruitment of the known or un- Upstate Biotechnology, 07-030), or anti-trimethyl-histone H3 (H3K4m3; Up-
known autonomous pathway components to FLC chro- state Biotechnology, 07-473) were used to immunoprecipitate the chromatin.
matin. Further studies aimed at identifying the direct The amount of immunoprecipitated FLC chromatin was determined by semi-
quantitative PCR on three different regions of FLC locus as previously reported
targets of HAC1 in the FLC-dependent pathway would (Bastow et al., 2004). Primer sequences were listed in Supplemental Table S3.
help to distinguish between these possibilities.
Supplemental Data
The following materials are available in the online version of this article.
MATERIALS AND METHODS
Supplemental Figure S1. Chromatin immunoprecipitation of FLC in wild-
Plant Materials and Growth Conditions type and hac1 plants.
All the Arabidopsis (Arabidopsis thaliana) lines used in this study were in Supplemental Figure S2. Expression levels of MAF4, MAF5, and FLC in
ecotype Col-0. Plants were grown at 23°C 6 1°C. The photoperiods for LD autonomous pathway mutants.
were 16 h of light followed by 8 h of darkness and 8 h of light followed by 16 h Supplemental Table S1. Primers for genotyping of hac1 mutants.
of darkness for SD. For flowering-time measurements, plants were sowed side
by side and transferred into Versatile Environmental Test Chamber (MLR- Supplemental Table S2. Primers for RT-PCR and real-time PCR analysis
350H, SANYO) at 23°C 6 1°C. Flowering time was measured by counting the of flowering-time regulatory genes.
total number of rosette and cauline leaves at flowering and the days from Supplemental Table S3. Primers for chromatin immunoprecipitation
germination to floral bud formation. For GA treatment, plants were grown on analysis at FLC locus.
soil under SD, and a GA solution of 100 mM was sprayed once a week until
flowering. Ethanol was sprayed as untreated control. For vernalization treat-
ment, the hac1 mutants and wild-type plants were grown side by side at 4°C
for 6 or 10 weeks under SD and then transferred to 23°C under LD. Non-
vernalized control seeds were grown for 3 d under vernalization conditions. ACKNOWLEDGMENTS
We thank Dr. R. Jorgensen for analysis of the HAC1 protein structure, Dr.
L. Johnson for critical reading and comments on the manuscript, and Dr. R.
Identification of T-DNA Insertion Mutants Amasino for providing flc-3 seeds. We also thank the Arabidopsis Biological
DNA was isolated from hac1-3, hac1-4, and hac1-5 mutants. T-DNA border Resources Center at The Ohio State University and Bernd Weisshaar for
primers cx0101 LBb1 or cx0102 LBa1 were used to amplify DNA from mutants providing SALK and GABI T-DNA insertion lines in the Col background.
obtained from the SALK collection and T-DNA border primer cx1475 was Received January 5, 2007; accepted January 31, 2007; published February 9,
used for GABI T-DNA insertion mutant hac1-5. The gene-specific primers 2007.
were cx0517 and cx0516 for hac1-3 and cx0515 and cx0514 for hac1-4. hac1-5
was amplified with cx0514 and cx0517. Primer sequences were listed in
Supplemental Table S1.
LITERATURE CITED
Root Growth Conditions Alvarez-Buylla ER, Liljegren SJ, Pelaz S, Gold SE, Burgeff C, Ditta GS,
Seeds of Col-0 and hac1 mutants were surface sterilized with 10% bleach Vergara-Silva F, Yanofsky MF (2000) MADS-box gene evolution beyond
for 15 min, washed with sterile water, and plated on 9 3 9-cm2 dishes flowers: expression in pollen, endosperm, guard cells, roots and tri-
containing 30 mL of agar-solidified culture medium. This medium contains chomes. Plant J 24: 457–466
13 Murashige and Skoog basal medium with vitamins (PhytoTechnology Alvarez-Venegas R, Avramova Z (2005) Methylation patterns of histone H3
Laboratories) supplemented with 3% Suc and 0.8% to 1% plant tissue culture Lys 4, Lys 9 and Lys 27 in transcriptionally active and inactive Arabidopsis
agar. The plates were kept at 4°C for 3 d and then placed vertically to allow the genes and in atx1 mutants. Nucleic Acids Res 33: 5199–5207
downward growth of roots in a growth chamber under LD at 23°C. After 6 d, Amasino R (2004) Vernalization, competence, and the epigenetic memory
measurements of primary root length were made for each sample. of winter. Plant Cell 16: 2553–2559
Amasino RM (2005) Vernalization and flowering time. Curr Opin
Biotechnol 16: 154–158
Analysis of Transcript Levels Aukerman MJ, Lee I, Weigel D, Amasino RM (1999) The Arabidopsis
flowering-time gene LUMINIDEPENDENS is expressed primarily in
Transcript levels were measured either by real-time PCR and RT-PCR or by regions of cell proliferation and encodes a nuclear protein that regulates
RNA gel blots. Total RNA extraction and RT-PCR were performed as previ- LEAFY expression. Plant J 18: 195–203
ously described by Liu et al. (2005). Real-time PCR analysis was performed Ausin I, Alonso-Blanco C, Jarillo JA, Ruiz-Garcia L, Martinez-Zapater JM
using the Chromo4 Real-Time PCR instrument (MJ) and SYBR Green I (2004) Regulation of flowering time by FVE, a retinoblastoma-associated
(Invitrogen, S-7567). Primer sequences were listed in Supplemental Table S2. protein. Nat Genet 36: 162–166
The cycle number at which the amplification plot crosses a fixed threshold Bastow R, Mylne JS, Lister C, Lippman Z, Martienssen RA, Dean C (2004)
above baseline is defined as the threshold cycle (Ct). Specific gene expression Vernalization requires epigenetic silencing of FLC by histone methyla-
was normalized to the internal control gene Ubiquitin (UBQ; Mockler et al., tion. Nature 427: 164–167
2004) given by the formula 22DCt. DCt is the Ct of the target gene subtracted Bernstein BE, Humphrey EL, Erlich RL, Schneider R, Bouman P, Liu
from the Ct of the UBQ gene. JS, Kouzarides T, Schreiber SL (2002) Methylation of histone H3
1666 Plant Physiol. Vol. 143, 2007
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Histone Acetyltransferase AtHAC1 Regulates Flowering Time
Lys 4 in coding regions of active genes. Proc Natl Acad Sci USA 99: in Arabidopsis by K homology domain proteins. Proc Natl Acad Sci
8695–8700 USA 101: 12759–12764
Blazquez MA, Green R, Nilsson O, Sussman MR, Weigel D (1998) Mouradov A, Cremer F, Coupland G (2002) Control of flowering time: inter-
Gibberellins promote flowering of Arabidopsis by activating the LEAFY acting pathways as a basis for diversity. Plant Cell (Suppl) 14: S111–130
promoter. Plant Cell 10: 791–800 Noh B, Lee SH, Kim HJ, Yi G, Shin EA, Lee M, Jung KJ, Doyle MR,
Bordoli L, Netsch M, Luthi U, Lutz W, Eckner R (2001) Plant orthologs Amasino RM, Noh YS (2004) Divergent roles of a pair of homologous
of p300/CBP: conservation of a core domain in metazoan p300/CBP jumonji/zinc-finger-class transcription factor proteins in the regulation
acetyltransferase-related proteins. Nucleic Acids Res 29: 589–597 of Arabidopsis flowering time. Plant Cell 16: 2601–2613
Cheung WL, Briggs SD, Allis CD (2000) Acetylation and chromosomal Page T, Macknight R, Yang CH, Dean C (1999) Genetic interactions of the
functions. Curr Opin Cell Biol 12: 326–333 Arabidopsis flowering time gene FCA, with genes regulating floral
Dai P, Akimaru H, Tanaka Y, Hou DX, Yasukawa T, Kanei-Ishii C, initiation. Plant J 17: 231–239
Takahashi T, Ishii S (1996) CBP as a transcriptional coactivator of Pandey R, Muller A, Napoli CA, Selinger DA, Pikaard CS, Richards EJ,
c-Myb. Genes Dev 10: 528–540 Bender J, Mount DW, Jorgensen RA (2002) Analysis of histone acetyl-
Finnegan EJ, Sheldon CC, Jardinaud F, Peacock WJ, Dennis ES (2004) A transferase and histone deacetylase families of Arabidopsis thaliana
cluster of Arabidopsis genes with a coordinate response to an environ- suggests functional diversification of chromatin modification among
mental stimulus. Curr Biol 14: 911–916 multicellular eukaryotes. Nucleic Acids Res 30: 5036–5055
Gendall AR, Levy YY, Wilson A, Dean C (2001) The VERNALIZATION 2 Ponting CP, Blake DJ, Davies KE, Kendrick-Jones J, Winder SJ (1996) ZZ
gene mediates the epigenetic regulation of vernalization in Arabidopsis. and TAZ: new putative zinc fingers in dystrophin and other proteins.
Cell 107: 525–535 Trends Biochem Sci 21: 11–13
Goldman PS, Tran VK, Goodman RH (1997) The multifunctional role of Quesada V, Macknight R, Dean C, Simpson GG (2003) Autoregulation of
the co-activator CBP in transcriptional regulation. Recent Prog Horm FCA pre-mRNA processing controls Arabidopsis flowering time. EMBO J 22:
Res 52: 103–120 3142–3152
Han SK, Song JD, Noh YS, Noh B (2007) Role of plant CBP/p300-like Ratcliffe OJ, Kumimoto RW, Wong BJ, Riechmann JL (2003) Analysis of
genes in the regulation of flowering time. Plant J 49: 103–114 the Arabidopsis MADS AFFECTING FLOWERING gene family: MAF2
He Y, Amasino RM (2005) Role of chromatin modification in flowering- prevents vernalization by short periods of cold. Plant Cell 15: 1159–1169
time control. Trends Plant Sci 10: 30–35 Ratcliffe OJ, Nadzan GC, Reuber TL, Riechmann JL (2001) Regulation of
He Y, Doyle MR, Amasino RM (2004) PAF1-complex-mediated histone flowering in Arabidopsis by an FLC homologue. Plant Physiol 126: 122–132
methylation of FLOWERING LOCUS C chromatin is required for the Reeves PH, Coupland G (2000) Response of plant development to envi-
vernalization-responsive, winter-annual habit in Arabidopsis. Genes ronment: control of flowering by daylength and temperature. Curr Opin
Dev 18: 2774–2784 Plant Biol 3: 37–42
He Y, Michaels SD, Amasino RM (2003) Regulation of flowering time by Roelfsema JH, White SJ, Ariyurek Y, Bartholdi D, Niedrist D, Papadia F,
histone acetylation in Arabidopsis. Science 302: 1751–1754 Bacino CA, den Dunnen JT, van Ommen GJ, Breuning MH, et al (2005)
Henderson IR, Liu F, Drea S, Simpson GG, Dean C (2005) An allelic series Genetic heterogeneity in Rubinstein-Taybi syndrome: mutations in both
reveals essential roles for FY in plant development in addition to flowering- the CBP and EP300 genes cause disease. Am J Hum Genet 76: 572–580
time control. Development 132: 3597–3607 Roth SY, Denu JM, Allis CD (2001) Histone acetyltransferases. Annu Rev
Johnson L, Cao X, Jacobsen S (2002) Interplay between two epigenetic Biochem 70: 81–120
marks: DNA methylation and histone H3 lysine 9 methylation. Curr Biol Sanda SL, Amasino RM (1996) Ecotype-specific expression of a flowering
12: 1360–1367 mutant phenotype in Arabidopsis thaliana. Plant Physiol 111: 641–644
Kim HJ, Hyun Y, Park JY, Park MJ, Park MK, Kim MD, Lee MH, Moon J, Schomburg FM, Patton DA, Meinke DW, Amasino RM (2001) FPA, a gene
Lee I, Kim J (2004) A genetic link between cold responses and flowering involved in floral induction in Arabidopsis, encodes a protein contain-
time through FVE in Arabidopsis thaliana. Nat Genet 36: 167–171 ing RNA-recognition motifs. Plant Cell 13: 1427–1436
Kim SY, He Y, Jacob Y, Noh YS, Michaels S, Amasino R (2005) Establish- Scortecci K, Michaels SD, Amasino RM (2003) Genetic interactions be-
ment of the vernalization-responsive, winter-annual habit in Arabidop- tween FLM and other flowering-time genes in Arabidopsis thaliana.
sis requires a putative histone H3 methyl transferase. Plant Cell 17: Plant Mol Biol 52: 915–922
3301–3310 Scortecci KC, Michaels SD, Amasino RM (2001) Identification of a MADS-
Komeda Y (2004) Genetic regulation of time to flower in Arabidopsis box gene, FLOWERING LOCUS M, that represses flowering. Plant J 26:
thaliana. Annu Rev Plant Biol 55: 521–535 229–236
Lee I, Aukerman MJ, Gore SL, Lohman KN, Michaels SD, Weaver LM, Seo SB, McNamara P, Heo S, Turner A, Lane WS, Chakravarti D (2001)
John MC, Feldmann KA, Amasino RM (1994) Isolation of LUMINIDE- Regulation of histone acetylation and transcription by INHAT, a human
PENDENS: a gene involved in the control of flowering time in Arabi- cellular complex containing the set oncoprotein. Cell 104: 119–130
dopsis. Plant Cell 6: 75–83 Sheldon CC, Burn JE, Perez PP, Metzger J, Edwards JA, Peacock WJ, Dennis
Lilja T, Qi D, Stabell M, Mannervik M (2003) The CBP coactivator ES (1999) The FLF MADS box gene: a repressor of flowering in Arabidopsis
functions both upstream and downstream of Dpp/Screw signaling in regulated by vernalization and methylation. Plant Cell 11: 445–458
the early Drosophila embryo. Dev Biol 262: 294–302 Simpson GG, Dijkwel PP, Quesada V, Henderson I, Dean C (2003) FY is
Lim MH, Kim J, Kim YS, Chung KS, Seo YH, Lee I, Hong CB, Kim HJ, an RNA 3# end-processing factor that interacts with FCA to control the
Park CM (2004) A new Arabidopsis gene, FLK, encodes an RNA binding Arabidopsis floral transition. Cell 113: 777–787
protein with K homology motifs and regulates flowering time via Simpson GG, Gendall AR, Dean C (1999) When to switch to flowering.
FLOWERING LOCUS C. Plant Cell 16: 731–740 Annu Rev Cell Dev Biol 15: 519–550
Liu B, Li P, Li X, Liu C, Cao S, Chu C, Cao X (2005) Loss of function of Sterner DE, Berger SL (2000) Acetylation of histones and transcription-
OsDCL1 affects microRNA accumulation and causes developmental related factors. Microbiol Mol Biol Rev 64: 435–459
defects in rice. Plant Physiol 139: 296–305 Strahl BD, Allis CD (2000) The language of covalent histone modifications.
Liu C, Lu J, Tan J, Li L, Huang B (2004) Human interleukin-5 expression is Nature 403: 41–45
synergistically regulated by histone acetyltransferase CBP/p300 and Sung S, Amasino RM (2004a) Vernalization and epigenetics: how plants
transcription factors C/EBP, NF-AT and AP-1. Cytokine 27: 93–100 remember winter. Curr Opin Plant Biol 7: 4–10
Macknight R, Bancroft I, Page T, Lister C, Schmidt R, Love K, Westphal L, Sung S, Amasino RM (2004b) Vernalization in Arabidopsis thaliana is
Murphy G, Sherson S, Cobbett C, et al (1997) FCA, a gene controlling mediated by the PHD finger protein VIN3. Nature 427: 159–164
flowering time in Arabidopsis, encodes a protein containing RNA- Sung S, Amasino RM (2005) Remembering winter: toward a molecular
binding domains. Cell 89: 737–745 understanding of vernalization. Annu Rev Plant Biol 56: 491–508
Michaels SD, Amasino RM (1999) FLOWERING LOCUS C encodes a novel Tian L, Chen ZJ (2001) Blocking histone deacetylation in Arabidopsis
MADS domain protein that acts as a repressor of flowering. Plant Cell induces pleiotropic effects on plant gene regulation and development.
11: 949–956 Proc Natl Acad Sci USA 98: 200–205
Mockler TC, Yu X, Shalitin D, Parikh D, Michael TP, Liou J, Huang J, Tian L, Fong MP, Wang JJ, Wei NE, Jiang H, Doerge RW, Chen ZJ (2005)
Smith Z, Alonso JM, Ecker JR, et al (2004) Regulation of flowering time Reversible histone acetylation and deacetylation mediate genome-wide,
Plant Physiol. Vol. 143, 2007 1667
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Deng et al.
promoter-dependent and locus-specific changes in gene expression time and floral initiation in Arabidopsis thaliana. Plant Cell Physiol 40:
during plant development. Genetics 169: 337–345 647–650
Vo N, Goodman RH (2001) CREB-binding protein and p300 in transcrip- Yao TP, Oh SP, Fuchs M, Zhou ND, Ch#ng LE, Newsome D, Bronson RT,
tional regulation. J Biol Chem 276: 13505–13508 Li E, Livingston DM, Eckner R (1998) Gene dosage-dependent embry-
Wilson RN, Heckman JW, Somerville CR (1992) Gibberellin is required for onic development and proliferation defects in mice lacking the tran-
flowering in Arabidopsis thaliana under short days. Plant Physiol 100: scriptional integrator p300. Cell 93: 361–372
403–408 Yuan LW, Giordano A (2002) Acetyltransferase machinery conserved in
Yang CH, Chou ML (1999) FLD interacts with CO to affect both flowering p300/CBP-family proteins. Oncogene 21: 2253–2260
1668 Plant Physiol. Vol. 143, 2007
Downloaded on May 12, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.CORRECTIONS
Due to a software error, six articles in the April 2007 issue were printed with an incorrect
DOI (digital object identifier) number at the bottom of the first page. These numbers allow
the articles to be uniquely identified online. The online versions of the articles have been
updated, and the correct numbers are listed below.
Chen et al., Vol. 143: 1954–1967, www.plantphysiol.org/cgi/doi/10.1104/pp.107.095588
Deng et al., Vol. 143: 1660–1668, www.plantphysiol.org/cgi/doi/10.1104/pp.107.095521
Kalituho et al., Vol. 143: 1861–1870, www.plantphysiol.org/cgi/doi/10.1104/pp.107.095562
Marino et al., Vol. 143: 1968–1974, www.plantphysiol.org/cgi/doi/10.1104/pp.107.097139
Park et al., Vol. 143: 1547–1560, www.plantphysiol.org/cgi/doi/10.1104/pp.107.096396
Schaarschmidt et al., Vol. 143: 1827–1840, www.plantphysiol.org/cgi/doi/10.1104/
pp.107.096446
www.plantphysiol.org/cgi/doi/10.1104/pp.104.900225
Plant Physiology, June 2007, Vol. 144, p. 1233, www.plantphysiol.org Ó 2007 American Society of Plant Biologists 1233You can also read

















































