Lawrence Berkeley National Laboratory - Recent Work - eScholarship
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Lawrence Berkeley National Laboratory
Recent Work
Title
Enzymes and Models
Permalink
https://escholarship.org/uc/item/5v03f8h5
Authors
Yachandra, VK
Yano, J
Publication Date
2021-08-17
Peer reviewed
eScholarship.org Powered by the California Digital Library
University of California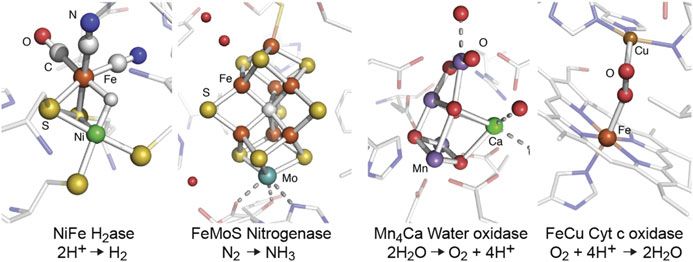
Chapter 28 Enzymes and Models Junko Yano and Vittal Yachandra 28.1 Introduction Nature uses remarkably varied systems and mechanisms to perform chemical reactions with efficiency, speed, and complexity. At the active site of many enzymes are metal centers, responsible for the rearrangement of electrons and atoms in order to carry out electron transfer and catalytic reactions. Some of the representative metal clusters, especially those that contain heterometallic clusters, that are related to important reactions in nature, are shown in Fig. 28.1 [1–4]. Many of these consist of transition metals, such as V, Mn, Fe, Co, Ni, Cu, Zn, Mo, W and others. Using the flexibility of their electronic structures that can accommodate different redox states, they catalyze multi-electron reactions in aqueous solution, under conditions of ambient temperature and pressure. Geometric and electronic structures of metal catalytic centers have been studied with X-ray absorption spectroscopy (XAS) in numerous metalloenzymes by taking advantage of the element-specificity of X-ray spectroscopy. X-ray absorption near edge spectroscopy (XANES) has been commonly used as a probe of the metal oxidation state, while extended X-ray absorption fine structure (EXAFS) has been applied for identifying the geometric structure of the metal complexes, and the method does not require single crystals. This makes the EXAFS method a powerful tool for extracting structural information of reaction intermediates, that cannot be crystallized. While solution NMR study is another way to get structural information of non-crystalline biological samples, it is difficult to apply the method to large metalloenzymes (>100 kDa) [5] or to study paramagnetic metal centers of metalloenzymes. J. Yano (*) • V. Yachandra Molecular Biophysics and Integrated Bioimaging Division, Lawrence Berkeley National Laboratory, Berkeley, CA 94720, USA e-mail: JYano@lbl.gov © Springer International Publishing Switzerland 2017 451 Y. Iwasawa et al. (eds.), XAFS Techniques for Catalysts, Nanomaterials, and Surfaces, DOI 10.1007/978-3-319-43866-5_28
452 J. Yano and V. Yachandra Fig. 28.1 Some examples of metalloenzymes that contain heterometallic clusters, such as NiFe hydrogenase, FeMo nitrogenase, Mn4Ca water oxidase, and FeCu cytc oxidase, that have been studied using X-ray spectroscopic techniques In addition to XAS, more advanced X-ray-based techniques such as RIXS (resonant inelastic X-ray scattering), XES (X-ray emission spectroscopy), and X-ray Raman spectroscopy [6], have been introduced to study biological systems, owing to the availability of bright synchrotron X-ray sources. XAS and XES data collection of biological samples have become somewhat routine. However, getting useful data often remains challenging due to several difficulties that are unique to biological systems. For example, X-ray-induced changes during data collection have been a concern. Reducing the background signal that arise from light atoms is another issue. Furthermore, interpreting X-ray spectroscopic data of biological systems often requires a set of adequate model complexes, that provide the basis for evaluating the influence of the coordination environment of the absorber elements. Study of structurally well-characterized model complexes also provides a benchmark for understanding the EXAFS from metal systems of unknown structures. In this chapter, the X-ray spectroscopy methods that have been used for studying biological samples are described, using several examples. In addition to the syn- chrotron X-ray-based methods, recent introduction of XFELs (X-ray free electron lasers) have opened a new way of collecting X-ray spectroscopy of biological systems by capturing reaction intermediates under physiological conditions, and this chapter touches on some of the applications that have been demonstrated recently. 28.2 X-Ray Spectroscopy of Biological Systems X-ray crystallography and X-ray spectroscopy of metalloenzymes provide comple- mentary information: crystallography probes the structural changes of the cofactor and the overall protein, while XAS provides detailed information about changes in

28 Enzymes and Models 453 the electronic structure of the metal of interest and its local structural information. When combined, these complementary measurements will provide a more detailed understanding of important enzymatic reactions. In general, the advantage of EXAFS over X-ray crystallography is that the local structural information around the metal of interest can be obtained even from disordered samples such as powders and solution, and usually the metal–metal and metal–ligand distance information is much better than that can be obtained from crystallography. However, ordered samples like oriented membranes and single crystals often increase the information obtained from X-ray spectroscopy. The advantages and the limitations of XAS for the structural studies of biological systems are summarized in detail below. 1. In a metalloenzyme with multiple metal centers like cytochrome oxidase (Cu and Fe), photosystem II (Mn and Ca), or nitrogenase (Fe/V and Mo), it is possible to study the structural environment of each metal atom independently, because of the element specificity of XAS. 2. The metal of interest is never “silent” with respect to X-ray spectra, and one can always probe the metal site structure by X-ray spectroscopy. It could be “silent” with respect to EPR (electron paramagnetic resonance spectroscopy), optical, or other spectroscopic methods. However, this could become a disadvantage, if there are several metal centers going through different chemical states. 3. X-ray spectroscopy is not limited by the state of the sample, because it is sensitive only to the local metal site structure. Therefore, one can either trap intermediates in the enzymatic cycle or modify the site by the addition of inhibitors or substrate, or generate other chemical modifications or site-directed mutations. Such samples can be made as frozen solutions, avoiding the problems of trying to obtain single crystals. It is also important to realize the intrinsic limitations of EXAFS for studying structures. A frequent problem is the inability to distinguish between scattering atoms with little difference in atomic number (C, N, O or S, Cl, or Mn, Fe). Distances are usually the most reliably determined structural parameters from EXAFS. But the range of data that can be collected, often-times due to practical reasons like the presence of the K-edge of another metal, or data collection time for dilute biological samples, limits the resolution of distance determinations to between 0.1 and 0.2 Å. Also it is difficult to determine whether a Fourier peak should be fit to one distance with a relatively large disorder parameter or to two distances, each having a small disorder parameter. Careful statistical analysis, taking into consideration the degrees of freedom in the fits, should precede any such analysis. The resolution in the distance Δr can be estimated from the relation that ΔrΔk ~ 1 (see Sect. 4.2). Determination of coordination numbers or number of backscatterers is fraught with difficulties. The Debye-Waller factor is strongly correlated with the coordination number and one must have recourse to other information to narrow the range that is possible from curve fitting analysis alone.
454 J. Yano and V. Yachandra
It is therefore very useful to compare the spectra from the complex in the
metalloenzyme to some known model complexes and then use Debye-Waller
parameters obtained from the model complexes in the fits.
28.3 Radiation-Induced Changes During Data Collection
Damage to biological samples by X-rays is cause for serious concern for X-ray
based experiments, in particular for the study of biological samples. However, with
the right precautions one can successfully perform these experiments under the
threshold of radiation damage. At synchrotron sources, most of the damage is
produced by free radicals and hydrated electrons that are produced by X-rays in
biological samples. The diffusion of the free radicals and hydrated electrons can be
minimized by the use of low temperatures. The use of a liquid He flow cryostat or
liquid He cryostream, where the samples are at atmospheric pressure in a He gas
atmosphere, has greatly reduced the risk of sample damage by X-rays. XAS
experiments require a lower X-ray dose than X-ray crystallography, and radiation
damage can be precisely monitored and controlled, thus allowing for data collection
from an intact form of metal catalytic centers.
An example of the X-ray-induced changes is shown in Fig. 28.2. Solutions of
Photosystem II (PSII) that consists of the MnCaO5 catalytic center were exposed to
various X-ray doses at 100 K, leading to reduction of 5, 10 or 25 %, of the Mn in the
sample from the native Mn(III,IV) to Mn(II) [7]. The corresponding XANES
spectra are shown in Fig. 28.2a. A clear shift in the K-edge toward lower energies,
due to the reduction to Mn(II) can be seen when comparing with the intact state
a b
6
PSII S1 state
Normalized Intensity
5% X-ray reduced
1.0 10% X-ray reduced
Iχ(R)I (Å-4)
4 25% X-ray reduced
PSII S1 state
0.5
5% X-ray reduced
2
10% X-ray reduced
25% X-ray reduced
0.0
0
6540 6560 6580 0 2 4 6 8
X-ray energy (eV) Apparent distance (Å)
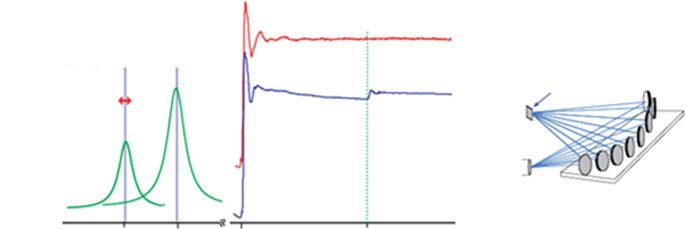
Fig. 28.2 (a) The Mn XANES spectrum of the oxygen-evolving complex from PS II that has been
photo-reduced by X-rays to different levels between 5 and 25 %, compared to a spectrum from an
intact PS II sample. (b) The corresponding Mn EXAFS Fourier transforms from the same PS II
samples, showing the disruption of the Mn4Ca oxo-bridged complex following X-ray radiation
induced reduction. This disruption manifests itself as a decrease in the intensity of FT peaks
assigned to Mn-Mn interactions at ~2.7 Å and/or Mn–Mn/Mn–Ca interactions at 3.2 Å [7]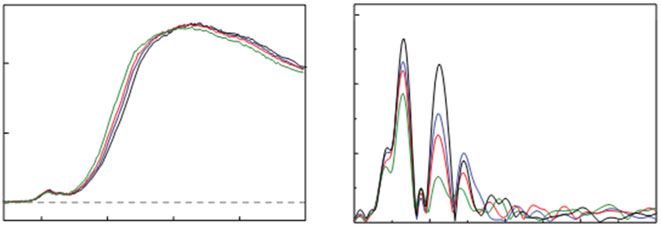
28 Enzymes and Models 455
spectrum from the native Mn(III,IV) form. In the EXAFS spectra (Fig. 28.2b), the
intensities of all three peaks are decreased upon Mn reduction, with the most
pronounced decrease in the second peak intensity, showing that upon Mn reduction
Mn-Mn distances change and they are elongated. The reduction of the metal center
by X-rays does not necessarily go through the catalytic reduction pathway, as it is
likely caused by the random attack of hydroxyl radicals that are created by the
interaction of X-rays and water molecules. Similar observations have been made
in many Fe-containing proteins such as putidaredoxin [8], Cytochrome C peroxi-
dase [9], and Fe/Fe and Fe/Mn ribonucleotide reductase [10], where the states
generated by X-ray photoreduction are significantly different from the native
catalytic state of the metal center. These studies show that radiation-induced
changes are a general problem when studying metalloenzymes with X-rays. Such
radiation-induced effects could potentially be turned into a tool to obtain new
insights into the structural and electronic structural nature of the metal cluster.
Nevertheless, radiation-induced changes remain one of the most challenging
aspects in biological X-ray spectroscopy. New approaches to overcome such effects
and collect radiation-damage-free data have become possible, using XFELs which
is described in Sect. 5.
28.4 Application of Various X-Ray Spectroscopy
Techniques to Study Metalloenzymes
28.4.1 Polarized X-Ray Absorption Spectroscopy
As described above, the advantage of XAS is that the local structural information
around the element of interest can be obtained from disordered samples such as
powders and solutions. However, ordered samples like membranes and single
crystals often increases the information obtained from XAS. For oriented single
crystals or ordered membranes the interatomic vector orientations can be deduced
from dichroism measurements.
Membrane proteins can be oriented on a substrate such that the lipid membrane
planes are roughly parallel to the substrate surface. This imparts a one-dimensional
order to these samples; while the z-axis for each membrane (collinear with the
membrane normal) is roughly parallel to the substrate normal, the x- and y-axes
remain disordered. Exploiting the plane-polarized nature of synchrotron radiation,
spectra can be collected at different angles between the substrate normal and the
X-ray E vector (Fig. 28.3a). The dichroism, which is the dependence of the intensity
of the absorber–backscatterer pair present in the oriented samples as a function of
the polarization of the X-rays, is reflected in, and can be extracted from, the
resulting X-ray absorption spectra [11–13]. The EXAFS of the oriented PS II
samples exhibits distinct dichroism, from which one can deduce the relative456 J. Yano and V. Yachandra
a b liq. He c
XRD detector cryostream
averaged
membrane normal Mn(V)≡O
Normalized Intensity
1.0
membrane
fragments
gonio O
XAS -meter
membrane
plane detector O N
≡
Mn(V) C
0.0
6530 6580
Incident
X-ray E vector X-ray Energy (eV)
X-rays
Fig. 28.3 (a) Experimental setup for single crystal X-ray absorption spectroscopy at the BL9-3 in
SSRL. (b) Schematic picture of the polarized XAS data collection from 1D oriented membrane
fragments. (c) Single crystal XAS of a Mn(V) inorganic complex, showing the pronounced
dichroism of the XANES spectrum [11]
orientations of several interatomic vector directions relative to the membrane
normal and derive a topological representation of the metal sites in the OEC [14].
Further refinement can be performed for samples with three-dimensional order.
Figure 28.3b shows the experimental setup for collecting single crystal XAS data at
the Stanford Synchrotron Radiation Lightsource, BL 9-3. It consists of a kappa
goniometer, a 30-element Ge detector for collecting XAS data, and a 2D detector
placed behind the sample for in situ collection of diffraction data to determine the
crystal orientation. The crystals are cooled using a liquid He cryostream to mini-
mize the radiation-induced changes of the sample. Single-crystal X-ray
spectroscopy has been performed on model complexes [15] and metalloproteins
[16–19]. These studies have been able to significantly expand the XAS information
available for these systems over what is gleaned from studies of isotropic samples.
An example of polarized XANES spectra from a Mn(V) model complex is
shown in Fig. 28.3c [11]. In this study, the orientationally resolved pre-edge
peaks were correlated to DFT calculations, and the results were used for assigning
the 1s to 3d transition components. The method was also applied to PSII in the dark
state prior to the determination of high-resolution X-ray crystal structure, and the
study improved the resolution and orientation of the metal–metal distances in
EXAFS [17].
28.4.2 Range-Extended EXAFS
In the typical EXAFS measurement of biological samples, spectra are taken up
to ~ 500 eV above the K-absorption edge, that provides a k-range of ~12.0 /Å.
However, longer the k-range, the EXAFS distance resolution improves and it
becomes important for some systems to resolve distance heterogeneity with similar28 Enzymes and Models 457
metal–ligand and metal–metal interactions. The distance resolution in a typical
EXAFS experiment is given by
ΔR ¼ π=2kmax ð28:1Þ
where kmax is the maximum energy of the photoelectron of the absorber element.
Getting larger kmax is not always possible, in particular, for systems which
contain adjacent elements in the periodic table due to the presence of the rising
edge of the next element. In principle, solid-state detectors can be used for dis-
criminating fluorescence signals from different elements, by electronically
windowing the Kα fluorescence from the absorber atom and collecting XAS data
as an excitation spectrum. However, the energy resolution of the detector at the
transition metal absorption edges is about 150–200 eV (fwhm), and this makes it
difficult to discriminate signals from adjacent transition metals. For the Mn K-edge
EXAFS studies of PS II, for example, the absorption edge of the obligatory Fe in PS
II (2-3Fe/PS II) limits the EXAFS energy range (Fig. 28.4). The use of a high-
resolution crystal monochromator provides a method to selectively separate the
fluorescence signals from adjacent metals, resulting in the collection of data to
higher photoelectron energies and leading to increased distance resolution and more
precise determination in the numbers of metal–metal vectors.
28.4.3 X-Ray Emission Spectroscopy
A complementary technique to XAS is XES. The photons that are emitted after the
creation of a core hole in the 1s shell form the K emission spectrum. Among such
emission processes, Kα lines originate from 2p to 1s transitions and Kβ lines from
the 3p to 1s transitions (Kβ1,3/Kβ0 ) and valence level to 1s transitions (Kβ2,5/Kβ00 )
Mn K-edge Mn EXAFS
Mn and Fe Fluorescence
Crystal Monochromator Data
Kα natural Mn Fe
linewidth Kα Kα
Fe K-edge X-ray beam
linewidth /w
crystal analyzer Ge detector data
Sample
Ge detector
w/ 150-200eV Ge 330
energy resolution crystals
Ge Detector
5900 6400 6550 7120 (eV)
Fig. 28.4 (left) X-ray fluorescence of Mn and Fe. The multi-crystal monochromator with 1 eV
resolution is tuned to the Mn Kα1 peak. (middle) Traditional XAFS spectrum (blue) and using a
crystal monochromator (red) (note the absence of Fe contribution). (right) The schematic for the
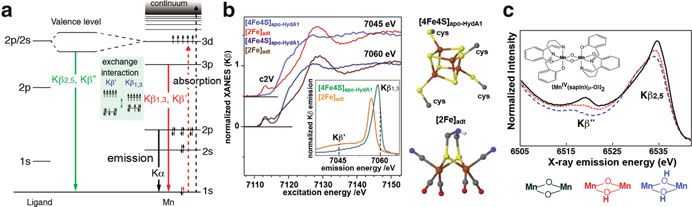
crystal monochromator used in a backscattering configuration [14]458 J. Yano and V. Yachandra Fig. 28.5 (a) Energy level diagram showing the excitation and emission processes for the Kα, Kβ1,3/Kβ0 and the Kβ2,5/Kβ00 transitions. (b) Kβ-detected XAS of apo-HydA1 protein binding only the [4Fe4S] unit of the H-cluster and of a synthetic [2Fe] model complex. The inset shows the Kβ XES, and two detection energy for the XAS (7045 and 7060 eV) are shown as dotted lines in the inset. The figure was adapted from Ref. [20]. (c) The valence to core Kβ2,5/Kβ00 emission spectra of a series of dinuclear MnIV complexes, where the bridging oxygen was protonated sequentially [21] (Fig. 28.5a). These spectra change depending on the chemical environment of the metals of interest. In particular, Kβ spectra have been used in biological systems for probing the oxidation states and identifying the ligand environment. Kβ1,3/Kβ0 spectra arise predominantly from the exchange interaction between the metal 3p and the net electron spin in the metal valence shell, i.e., the effective number of unpaired metal 3d electrons. Therefore, the spectrum is sensitive to the spin state of the metals, which indirectly reflects the oxidation state and covalency of the metal site, and has often been used for determining the oxidation states of metal catalytic sites [6]. Kβ1,3 arises from the constructive spin interactions, while Kβ0 from destructive spin interactions. The sensitivity of the Kβ1,3/Kβ0 spectra to the number of unpaired 3d electrons has the potential for being used for probing different metal sites in multinuclear clusters. As mentioned in Section II, there is no “silent” state in XAS. This can be a limiting factor for studying the system that contains several of the same element. For uniquely probing one particular reaction site, the use of different K-emission signals could become useful. One example is the [FeFe]-hydrogenases that contain 6 Fe in the metal catalytic center (H-cluster). Haumann et al. studied the H-cluster with site-selective XAS method, by using the Kβ1,3 and Kβ0 signals to distinguish between the low and high spin Fe sites in the two subcomplexes (Fig. 28.5b) [20]. By using a high-resolution spectrometer, in one case the Kβ-detected XAS was collected at the Kβ0 energy and in the other case at the Kβ1,3 energy; the former one represents the 2Fe site, while the latter one represents the 4Fe4S site. Thus the response of the two sites in different chemical states can be separately analyzed. The higher energy emission peaks above Kβ1,3 originate from valence-to-core transitions just below the Fermi level and can be separated into Kβ00 and the Kβ2,5 emission (Fig. 28.5a). Kβ2,5 emission is predominantly from ligand 2p (metal 4p) to metal 1s, and the Kβ00 emission is assigned to a ligand 2s to metal 1s; both are referred to as cross-over transitions [6]. Therefore, only direct ligands to the metal of interest are probed with Kβ2,5/Kβ00 emission, i.e., other C, N, and O atoms in the
28 Enzymes and Models 459 protein media do not contribute to the spectra. In particular, the energy of the Kβ00 transition depends on the difference between the metal 1s and ligand 2s binding energies, which reflects the environment of the ligand owing to orbital hybridization. Detecting the chemical states of ligands is important for understanding the reaction mechanism of numerous metalloenzymes. In particular, protonation/ deprotonation states of ligands moderate reaction chemistry in metal centers in biologically important processes. For example, copper oxo/hydroxo/peroxo com- plexes play vital roles in respiration, such as in hemocyanin [22], in biological metabolic pathways such as catechol oxidase activity or activation of aliphatic C-H bonds by dopamine β-monooxygenase [23], peptidyl-glycine α-amidating enzyme, and particulate methane monooxygenase [24]. It is also known that FeFe clusters in methane monooxygenase catalyze the hydroxylation of methane to methanol [25]. Understanding these events requires techniques that are sensitive enough to differentiate species that differ only by a single proton. Fig. 28.5c shows how the Kβ2,5 and Kβ00 spectra change depending on the protonation state of the bridging oxygen ligands of the metal cluster [21]. Several techniques have the potential to detect such a single protonation event, that includes vibrational spectroscopy and ligand sensitive EPR techniques such as ENDOR (electron nuclear double reso- nance), ESEEM (electron spin echo envelope modulation), and HYSCORE (hyper- fine sub-level correlation). The advantage of X-ray-based methods over EPR techniques is their element specificity and that they are unrestricted by the spin states of the compounds. The Kβ2,5 and Kβ00 XES was also applied for the study of nitrogenase to identify the central atom of the FeMoco cluster [26], using the element sensitivity of the Kβ00 peak position. A combination of the detailed data analysis and the theoretical calculations revealed that the central atom is a carbon. 28.4.4 Resonant Inelastic X-Ray Scattering Spectroscopy The pre-edge region of metal K-edge XAS, that corresponds to the 1s to 3d transition, is a probe of metal oxidation state and local geometry of metal catalytic centers. However, the K pre-edge spectra are usually weak compared to the K-main edge, as they mostly arise from quadrupole transition matrix elements. In addition, it suffers from a strong background from the dipole-allowed transition of the main edge, particularly for early transition metals. Transition metal L-edge XAS are in some ways a superior chemical probe to K-edge spectra as (1) 2p ! 3d transitions are allowed under dipole selection rules, while only s ! p transitions are allowed at the K-edge, (2) greater sensitivity to the occupancy of the 3d orbitals of metal provides a better indication of the oxidation states and symmetry of the complex involved. It is therefore anticipated that examination of the L-edge region will provide oxidation state information of the metal centers with significantly better resolution than that from K-edges. However, the disadvantage is that the energy of
460 J. Yano and V. Yachandra
these transitions requires the use of soft X-rays for direct excitation, leading to
drastically increased radiation damage compared to high energy hard X-rays
(100 times higher) and only surface absorption for bulk samples. These issues
make soft X-ray (metal L-edge) spectroscopy less applicable to biological systems.
The above-described problems can be overcome in 1s2p RIXS (resonant inelas-
tic X-ray scattering spectroscopy. In this method, hard X-rays tuned to the energy of
the K pre-edge are used to excite a 1s electron into an unoccupied valence orbital
(1s to 3d transition), and the emission due to decay of a 3p or 2p electron into the 1s
shell is measured as a function of the excitation energy. In experiments, the incident
energy is varied across the metal K-edge (1s to 3d) using hard X-rays, and the
crystal analyzer is scanned over the Kα emission energy (2p to 1s). The energy
difference between excitation and emission corresponds to the energy difference
between 2p or 3p and 3d orbitals and, therefore, one can get L-edge-like XAS
spectrum while taking advantage of using hard X-rays (high-excitation energy and
therefore the high X-ray penetration depth and larger attenuation length) to probe
these transitions indirectly. In addition, the RIXS 2D plot also makes the back-
ground separation of the pre-edge structure from the main K-edge feature easier.
As an example, the Fe site of the cytochrome c (cyt c) and its model compound
was studied by Fe 1s2d RIXS [27]. The RIXS study was used to quantify the highly
covalent nature of the porphyrin environment in the heme cofactor, through the
analysis of the L-edge XAS-like data from RIXS, and showed the increased
covalency for the Fe-S(Met) relative to Fe-N(His) axial ligand and a higher degree
of covalency for the ferric states relative to the ferrous states. Namely, it suggests
that the bond strengths for the Fe(II) and Fe(III) states are a critical factor for the
enzymatic function.
28.5 Application of XFELs for the Study
of Metalloenzymes Using X-Ray Spectroscopy
X-ray based techniques, such as X-ray spectroscopy, X-ray crystallography, as well
as X-ray scattering methods, have contributed significantly to the structural and
functional studies of metalloenzymes in the last decades, and have become indis-
pensable methods. However, to minimize or eliminate radiation-induced changes
during data collection has remained challenging. As described in Section II, the
data collection of biological samples is usually carried out at cryogenic temperature
to minimize the X-ray induced effect. However, there is an increasing effort to
understand biological phenomena as the enzymes function, through studies under
physiological/functional conditions. Additionally, the need to probe rare transi-
tions, which often are superior probes of chemical states but require very high
photon fluxes, , within a reasonable time, is another issue.
The recent advent of XFELs [28, 29] brought the possibility of probing labile
biological systems with high dose X-rays, by outrunning the radiation damage28 Enzymes and Models 461 processes and extracting the information before manifestation of the X-ray-induced changes to the sample using fs X-ray pulses. XFELs produce high intensity X-ray pulses in the fs time regime. In practice the number of photons in a single
462 J. Yano and V. Yachandra
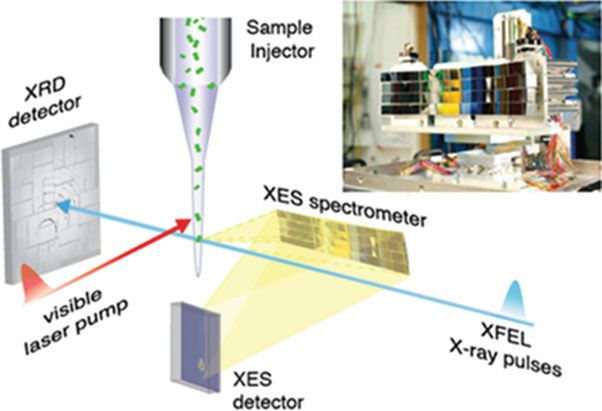
Fig. 28.6 Setup for simultaneous XES/XRD at the LCLS XFEL. The XES spectrometer is
mounted at 90 to the fs X-ray pulses and the XRD is recorded in forward scattering. The sample
is introduced into the interaction region using a jet as shown here, but other methods can also be
used. The energy dispersive XES spectrometer is shown at top right. Illumination of the samples is
achieved by visible laser pumps and are used for time resolved optical pump X-ray probe
measurements [36]
reaction paths in metalloenzymes and the important information of how to control
electron/proton flow and product/substrate transport, spatially and temporally,
during catalysis using the protein framework.
References
1. Ogata H, Nishikawa K, Lubitz W (2015) Hydrogens detected by subatomic resolution protein
crystallography in a [NiFe] hydrogenase. Nature 520:571
2. Kaiser JT, Hu Y, Wiig JA, Rees DC, Ribbe MW (2011) Structure of precursor-bound NifEN: a
nitrogenase femo cofactor maturase/insertase. Science 331:91
3. Suga M, Akita F, Hirata K, Ueno G, Murakami H, Nakajima Y, Shimizu T, Yamashita K,
Yamamoto M, Ago H, Shen J-R (2015) Native structure of photosystem II at 1.95 Å resolution
viewed by femtosecond X-ray pulses. Nature 517:99
4. Tsukihara T, Aoyama H, Yamashita E, Tomizaki T, Yamaguchi H, Shinzawa-Itoh K,
Nakashima R, Yaono R, Yoshikawa S (1995) Structures of metal sites of oxidized bovine
heart cytochrome c oxidase at 2.8 Å. Science 269:1069
5. Tzakos AG, Grace CRR, Lukavsky PJ, Riek R (2006) NMR techniques for very large proteins
and RNAs in solution. Annu Rev Biophys Biomol Struct 35:319
6. Glatzel P, Bergmann U (2005) High resolution 1s core hole X-ray spectroscopy in 3d transition
metal complexes - electronic and structural information. Coord Chem Rev 249:6528 Enzymes and Models 463
7. Yano J, Kern J, Irrgang K-D, Latimer MJ, Bergmann U, Glatzel P, Pushkar Y, Biesiadka J, Loll
B, Sauer K, Messinger J, Zouni A, Yachandra VK (2005) X-ray damage to the Mn4Ca
complex in single crystals of photosystem II: a case study for metalloprotein crystallography.
Proc Natl Acad Sci U S A 102:12047
8. Corbett MC, Latimer MJ, Poulos TL, Sevrioukova IF, Hodgson KO, Hedman B (2007)
Photoreduction of the active site of the metalloprotein putidaredoxin by synchrotron radiation.
Acta Cryst 63:951
9. Meharenna YT, Doukov T, Li HY, Soltis SM, Poulos TL (2010) Crystallographic and single-
crystal spectral analysis of the peroxidase ferryl intermediate. Biochemistry 49:2984
10. Sigfridsson KGV, Chernev P, Leidel N, Popović-Bijelić A, Gräslund A, Haumann M (2013)
Rapid X-ray photoreduction of dimetal-oxygen cofactors in ribonucleotide reductase. J Biol
Chem 288:9648
11. Yano J, Robblee J, Pushkar Y, Marcus MAM, Bendix J, Workman JM, Collins TJ, Solomon
EI, George SD, Yachandra VK (2007) Polarized X-ray absorption spectroscopy of single-
crystal Mn(V) complexes relevant to the oxygen-evolving complex of photosystem II. J Am
Chem Soc 129:12989
12. George GN, Prince RC, Frey TG, Cramer SP (1989) Oriented X-ray absorption-spectroscopy
of membrane-bound metalloproteins. Physica B 158:81
13. George GN, Cramer SP, Frey TG, Prince RC (1993) X-ray-absorption spectroscopy of oriented
cytochrome-oxidase. Biochim Biophys Acta 1142:240
14. Pushkar Y, Yano J, Glatzel P, Messinger J, Lewis A, Sauer K, Bergmann U, Yachandra VK
(2007) Structure and orientation of the Mn Ca cluster in plant photosystem II membranes
studied by polarized range-extended X-ray absorption spectroscopy. J Biol Chem 282:7198
15. Pickering IJ, George GN (1995) Polarized X-ray-absorption spectroscopy of cupric chloride
dihydrate. Inorg Chem 34:3142
16. Flank AM, Weininger M, Mortenson LE, Cramer SP (1986) Single-crystal EXAFS of nitro-
genase. J Am Chem Soc 108:1049
17. Yano J, Kern J, Sauer K, Latimer M, Pushkar Y, Biesiadka J, Loll B, Saenger W, Messinger J,
Zouni A, Yachandra VK (2006) Where water is oxidized to dioxygen: structure of the
photosynthetic Mn Ca cluster. Science 314:821
18. George GN, Hilton J, Temple C, Prince RC, Rajagopalan KV (1999) Structure of the
molybdenum site of dimethyl sulfoxide reductase. J Am Chem Soc 121:1256
19. Scott RA, Hahn JE, Doniach S, Freeman HC, Hodgson KO (1982) Polarized X-ray absorption-
spectra of oriented plastocyanin single-crystals - investigation of methionine copper coordi-
nation. J Am Chem Soc 104:5364
20. Chernev P, Lambertz C, Brunje A, Leidel N, Sigfridsson KG, Kositzki R, Hsieh CH, Yao S,
Schiwon R, Driess M, Limberg C, Happe T, Haumann M (2014) Hydride binding to the active
site of [FeFe]-hydrogenase. Inorg Chem 53:12164
21. Lassalle-Kaiser B, Boron TT 3rd, Krewald V, Kern J, Beckwith MA, Delgado-Jaime MU,
Schroeder H, Alonso-Mori R, Nordlund D, Weng TC, Sokaras D, Neese F, Bergmann U,
Yachandra VK, DeBeer S, Pecoraro VL, Yano J (2013) Experimental and computational X‐ray
emission spectroscopy as a direct probe of protonation states in Oxo-bridged Mn dimers
relevant to redox-active metalloproteins. Inorg Chem 52:12915
22. Magnus KA, Tonthat H, Carpenter JE (1994) Recent structural work on the oxygen-transport
protein hemocyanin. Chem Rev 94:727
23. Klinman J (2006) The copper-enzyme family of dopamine β -monooxygenase and
peptidylglycine α-hydroxylating monooxygenase: resolving the chemical pathway for sub-
strate hydroxylation. J Biol Chem 281:3013
24. Himes RA, Karlin KD (2009) Copper-dioxygen complex mediated C-H bond oxygenation:
relevance for particulate methane monooxygenase (pMMO). Curr Opin Chem Biol 13:119
25. Kryatov SV, Taktak S, Korendovych IV, Rybak-Akimova EV, Kaizer J, Torelli S, Shan XP,
Mandal S, MacMurdo VL, Payeras AMI, Que L (2005) Dioxygen binding to complexes with464 J. Yano and V. Yachandra
Fe (μ-OH) cores: steric control of activation barriers and O -adduct formation. Inorg Chem
44:85
26. Lancaster KM, Roemelt M, Ettenhuber P, Hu Y, Ribbe MW, Neese F, Bergmann U, DeBeer S
(2011) X-ray emission spectroscopy evidences a central carbon in the nitrogenase iron-
molybdenum cofactor. Science 334:974
27. Kroll T, Hadt RG, Wilson SA, Lundberg M, Yan JJ, Weng TC, Sokaras D, Alonso-Mori R,
Casa D, Upton MH, Hedman B, Hodgson KO, Solomon EI (2014) Resonant inelastic X-ray
scattering on ferrous and ferric bis-imidazole porphyrin and cytochrome c: nature and role of
the axial Methionine-Fe bond. J Am Chem Soc 136:18087
28. Emma P, Akre R, Arthur J, Bionta R, Bostedt C, Bozek J, Brachmann A, Bucksbaum P, Coffee
R, Decker FJ, Ding Y, Dowell D, Edstrom S, Fisher A, Frisch J, Gilevich S, Hastings J, Hays
G, Hering P, Huang Z, Iverson R, Loos H, Messerschmidt M, Miahnahri A, Moeller S, Nuhn
HD, Pile G, Ratner D, Rzepiela J, Schultz D, Smith T, Stefan P, Tompkins H, Turner J, Welch
J, White W, Wu J, Yocky G, Galayda J (2010) First lasing and operation of an Å-wavelength
free-electron laser. Nat Photonics 4:641
29. Ishikawa T, Aoyagi H, Asaka T, Asano Y, Azumi N, Bizen T, Ego H, Fukami K, Fukui T,
Furukawa Y, Goto S, Hanaki H, Hara T, Hasegawa T, Hatsui T, Higashiya A, Hirono T,
Hosoda N, Ishii M, Inagaki T, Inubushi Y, Itoga T, Joti Y, Kago M, Kameshima T, Kimura H,
Kirihara Y, Kiyomichi A, Kobayashi T, Kondo C, Kudo T, Maesaka H, Marechal XM, Masuda
T, Matsubara S, Matsumoto T, Matsushita T, Matsui S, Nagasono M, Nariyama N, Ohashi H,
Ohata T, Ohshima T, Ono S, Otake Y, Saji C, Sakurai T, Sato T, Sawada K, Seike T, Shirasawa
K, Sugimoto T, Suzuki S, Takahashi S, Takebe H, Takeshita K, Tamasaku K, Tanaka H,
Tanaka R, Tanaka T, Togashi T, Togawa K, Tokuhisa A, Tomizawa H, Tono K, Wu SK,
Yabashi M, Yamaga M, Yamashita A, Yanagida K, Zhang C, Shintake T, Kitamura H,
Kumagai N (2012) A compact X-ray free-electron laser emitting in the sub-Å region.
NatPhotonics 6:540
30. Chapman HN, Fromme P, Barty A, White TA, Kirian RA, Aquila A, Hunter MS, Schulz J,
DePonte DP, Weierstall U, Doak RB, Maia FRNC, Martin AV, Schlichting I, Lomb L,
Coppola N, Shoeman RL, Epp SW, Hartmann R, Rolles D, Rudenko A, Foucar L, Kimmel
N, Weidenspointner G, Holl P, Liang MN, Barthelmess M, Caleman C, Boutet S, Bogan MJ,
Krzywinski J, Bostedt C, Bajt S, Gumprecht L, Rudek B, Erk B, Schmidt C, Homke A, Reich
C, Pietschner D, Struder L, Hauser G, Gorke H, Ullrich J, Herrmann S, Schaller G, Schopper F,
Soltau H, Kuhnel KU, Messerschmidt M, Bozek JD, Hau-Riege SP, Frank M, Hampton CY,
Sierra RG, Starodub D, Williams GJ, Hajdu J, Timneanu N, Seibert MM, Andreasson J,
Rocker A, Jonsson O, Svenda M, Stern S, Nass K, Andritschke R, Schroter CD, Krasniqi F,
Bott M, Schmidt KE, Wang XY, Grotjohann I, Holton JM, Barends TRM, Neutze R,
Marchesini S, Fromme R, Schorb S, Rupp D, Adolph M, Gorkhover T, Andersson I,
Hirsemann H, Potdevin G, Graafsma H, Nilsson B, Spence JCH (2011) Femtosecond X-ray
protein nanocrystallography. Nature 470:73
31. Alonso-Mori R, Kern J, Gildea RJ, Sokaras D, Weng TC, Lassalle-Kaiser B, Tran R, Hattne J,
Laksmono H, Hellmich J, Glockner C, Echols N, Sierra RG, Schafer DW, Sellberg J, Kenney
C, Herbst R, Pines J, Hart P, Herrmann S, Grosse-Kunstleve RW, Latimer MJ, Fry AR,
Messerschmidt MM, Miahnahri A, Seibert MM, Zwart PH, White WE, Adams PD, Bogan
MJ, Boutet S, Williams GJ, Zouni A, Messinger J, Glatzel P, Sauter NK, Yachandra VK, Yano
J, Bergmann U (2012) Energy-dispersive X-ray emission spectroscopy using an X-ray free-
electron laser in a shot-by-shot mode. Proc Natl Acad Sci U S A 109:19103
32. Zhang WK, Alonso-Mori R, Bergmann U, Bressler C, Chollet M, Galler A, Gawelda W, Hadt
RG, Hartsock RW, Kroll T, Kjaer KS, Kubicek K, Lemke HT, Liang HYW, Meyer DA,
Nielsen MM, Purser C, Robinson JS, Solomon EI, Sun Z, Sokaras D, van Driel TB, Vanko G,
Weng TC, Zhu DL, Gaffney K (2014) Tracking excited-state charge and spin dynamics in iron
coordination complexes. J Nature 509:34528 Enzymes and Models 465
33. Bertoni R, Cammarata M, Lorenc M, Matar SF, Letard JF, Lemke HT, Collet E (2015)
Ultrafast light-induced spin-state trapping photophysics investigated in Fe(phen) (NCS)
spin-crossover crystal. Acc Chem Res 48:774
34. Lemke HT, Bressler C, Chen LX, Fritz DM, Gaffney KJ, Galler A, Gawelda W, Haldrup K,
Hartsock RW, Ihee H, Kim J, Kim KH, Lee JH, Nielsen MM, Stickrath AB, Zhang WK, Zhu
DL, Cammarata M (2013) Femtosecond X-ray absorption spectroscopy at a hard X-ray free
electron laser: application to spin crossover dynamics. J Phys Chem A 117:735
35. Mitzner R, Rehanek J, Kern J, Gul S, Hattne J, Taguchi T, Alonso-Mori R, Tran R, Weniger C,
Schroder H, Quevedo W, Laksmono H, Sierra RG, Han G, Lassalle-Kaiser B, Koroidov S,
Kubicek K, Schreck S, Kunnus K, Brzhezinskaya M, Firsov A, Minitti MP, Turner JJ, Moeller
S, Sauter NK, Bogan MJ, Nordlund D, Schlotter WF, Messinger J, Borovik A, Techert S, de
Groot FM, Fohlisch A, Erko A, Bergmann U, Yachandra VK, Wernet P, Yano J (2013) L-edge
X-ray absorption spectroscopy of dilute systems relevant to metalloproteins using an X-ray
free-electron laser. J Phys Chem Lett 4:3641
36. Kern J, Alonso-Mori R, Tran R, Hattne J, Gildea RJ, Echols N, Glockner C, Hellmich J,
Laksmono H, Sierra RG, Lassalle-Kaiser B, Koroidov S, Lampe A, Han G, Gul S, Difiore D,
Milathianaki D, Fry AR, Miahnahri A, Schafer DW, Messerschmidt M, Seibert MM, Koglin
JE, Sokaras D, Weng TC, Sellberg J, Latimer MJ, Grosse-Kunstleve RW, Zwart PH, White
WE, Glatzel P, Adams PD, Bogan MJ, Williams GJ, Boutet S, Messinger J, Zouni A, Sauter
NK, Yachandra VK, Bergmann U, Yano J (2013) Simultaneous femtosecond X-ray spectros-
copy and diffraction of photosystem II at room temperature. Science 340:491You can also read

















































