The Evolutionary History of Wild, Domesticated, and Feral Brassica oleracea (Brassicaceae)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Evolutionary History of Wild, Domesticated, and Feral
Brassica oleracea (Brassicaceae)
Makenzie E. Mabry ,*,1 Sarah D. Turner-Hissong,2 Evan Y. Gallagher,1 Alex C. McAlvay,3 Hong An,1
Patrick P. Edger,4 Jonathan D. Moore,5 David A.C. Pink,6 Graham R. Teakle,7 Chris J. Stevens,8,9
Guy Barker,7 Joanne Labate,10 Dorian Q. Fuller,9,11,12 Robin G. Allaby,7 Timothy Beissinger,13
Jared E. Decker,14 Michael A. Gore,15 and J. Chris Pires*,1
1
Division of Biological Sciences and Bond Life Sciences Center, University of Missouri, Columbia, MO, USA
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
2
Department of Evolution and Ecology, University of California Davis, Davis, CA, USA
3
Institute of Economic Botany, The New York Botanical Garden, Bronx, NY, USA
4
Department of Horticulture, Michigan State University, East Lansing, MI, USA
5
Systems Biology Centre, University of Warwick, Coventry, United Kingdom
6
Agriculture and Environment Department, Harper Adams University, Newport, United Kingdom
7
School of Life Science, University of Warwick, Coventry, United Kingdom
8
School of Archaeology and Museology, Peking University, Beijing, China
9
Institute of Archaeology, University College London, London, United Kingdom
10
USDA, ARS Plant Genetic Resources Unit, Cornell AgriTech, Geneva, NY, USA
11
School of Cultural Heritage, Northwest University, Xi’an, Shaanxi, China
12
Department of Archaeology, Max Planck Institute for the Science of Human History, Jena, Germany
13
Division of Plant Breeding Methodology, Department of Crop Sciences, University of Goettingen, Goettingen, Germany
14
Division of Animal Sciences, University of Missouri, Columbia, MO, USA
15
Plant Breeding and Genetics Section, School of Integrative Plant Science, Cornell University, Ithaca, NY, USA
*Corresponding authors: E-mails: mmabry44@gmail.com; piresjc@missouri.edu.
Associate editor: Michael Purugganan
Abstract
Understanding the evolutionary history of crops, including identifying wild relatives, helps to provide insight for con-
servation and crop breeding efforts. Cultivated Brassica oleracea has intrigued researchers for centuries due to its wide
diversity in forms, which include cabbage, broccoli, cauliflower, kale, kohlrabi, and Brussels sprouts. Yet, the evolutionary
Article
history of this species remains understudied. With such different vegetables produced from a single species, B. oleracea is
a model organism for understanding the power of artificial selection. Persistent challenges in the study of B. oleracea
include conflicting hypotheses regarding domestication and the identity of the closest living wild relative. Using newly
generated RNA-seq data for a diversity panel of 224 accessions, which represents 14 different B. oleracea crop types and
nine potential wild progenitor species, we integrate phylogenetic and population genetic techniques with ecological
niche modeling, archaeological, and literary evidence to examine relationships among cultivars and wild relatives to
clarify the origin of this horticulturally important species. Our analyses point to the Aegean endemic B. cretica as the
closest living relative of cultivated B. oleracea, supporting an origin of cultivation in the Eastern Mediterranean region.
Additionally, we identify several feral lineages, suggesting that cultivated plants of this species can revert to a wild-like
state with relative ease. By expanding our understanding of the evolutionary history in B. oleracea, these results con-
tribute to a growing body of knowledge on crop domestication that will facilitate continued breeding efforts including
adaptation to changing environmental conditions.
Key words: cabbage, domestication, crop wild relatives, Mediterranean, origin, ecological niche.
Introduction A key tenet of evolutionary and plant biology is under-
standing how plants respond and adapt to changes in envi-
“Greek legend has it that the cabbage sprung from ronmental conditions, which can be better understood by
where Zeus’ sweat hit the ground.” leveraging genotypic diversity and investigating the connec-
–N.D. Mitchell (1976) tions between genotype and phenotype. Crop wild relatives
ß The Author(s) 2021. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://
creativecommons.org/licenses/by-nc/4.0/), which permits non-commercial re-use, distribution, and reproduction in any medium,
provided the original work is properly cited. For commercial re-use, please contact journals.permissions@oup.com Open Access
Mol. Biol. Evol. doi:10.1093/molbev/msab183 Advance Access publication June 22, 2021 1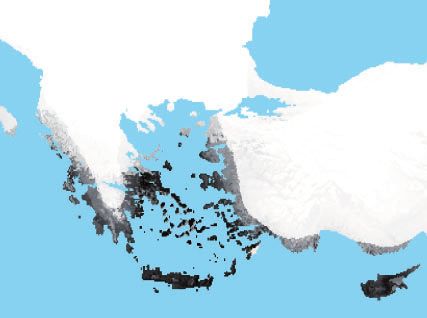
Mabry et al. . doi:10.1093/molbev/msab183 MBE
(CWRs) provide pools of allelic diversity that at one time were laconica; Snogerup et al. 1990) or only two (subsp. cretica
shared through a common ancestor with cultivated relatives. and nivea; Gustafsson et al. 1976). The taxonomic confusion
Although Vavilov recognized the potential of CWRs in the is perhaps best highlighted by Bailey (1930), who stated that
early 1900s (Vavilov 1926), advances in genomics and genome “Some of these plants appear to be more confused in litera-
editing techniques have enabled scientists to better realize ture than in nature.” The progenitor species of B. oleracea is
the potential of CWRs as a source of diversity and novel traits further obscured by the presence of weedy, cabbage-like
for the improvement of cultivated populations (Prohens et al. plants along the coastline of western Europe (England,
2017; Li et al. 2018; Fernie and Yan 2019; Khoury et al. 2020; France, and Spain), which have also been referred to as B.
Turner-Hissong et al. 2020). Yet these scientific advancements sylvestris (Mitchell 1976) or B. oleracea var. sylvestris (Gladis
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
are hindered in that we still have not identified the CWRs of and Hammer 2001). The role of these weedy populations in
many important crop species. Although cabbage may not the domestication of B. oleracea is unclear, with some studies
have exactly formed from Zeus’ sweat, its evolutionary his- suggesting these coastal wild populations represent the pro-
tory, including identifying the closest living wild relative and genitor species (Snogerup et al. 1990; Song et al. 1990), and
origin of domestication, is still left unclear due to taxonomic others identifying these wild forms as plants that escaped
confusion and the lack of genetic and archaeological cultivation (Mitchell 1976; Mitchell and Richards 1979).
evidence. Given the uncertainty surrounding wild relatives and
The horticultural crop Brassica oleracea L. has played an weedy populations, researchers have proposed numerous hy-
important role in global food systems for centuries, providing potheses for the progenitor species of B. oleracea (table 1).
a source of leaf and root vegetables, fodder, and forage Hypotheses range from a single domestication with a single
(Shyam et al. 2012). When first introduced to the species, progenitor species (Song et al. 1990; Allender et al. 2007) to
Darwin drew many parallels between his theory of natural multiple domestications arising from multiple progenitor spe-
selection and the cultivation practices that led to the varied cies (de Candolle 1855; Neutrofal 1927; Lizgunova 1959; Helm
forms of this plant (Darwin 1868). Although many people 1963; Snogerup 1980; Heaney et al. 1987; Song et al. 1988;
may recognize that various dog breeds are all part of the Swarup and Brahmi 2005). Findings that point to a single
same species, they are often surprised to learn that the do- origin of domestication have proposed different wild species
mesticated forms of B. oleracea, broccoli (var. italica), Brussels as the progenitor (Snogerup et al. 1990; Song et al. 1990;
sprouts (var. gemmifera), cabbage (var. capitata), cauliflower Hodgkin 1995; Maggioni et al. 2018). For instance, Neutrofal
(var. botrytis), kale (var. acephala), and kohlrabi (var. gongyl- (1927) suggested that B. montana was the progenitor of cab-
odes) are all one species as well. The global market for B. bages and that B. rupestris was the progenitor of kohlrabi,
oleracea crops was around 70.1 million metric tons, in terms whereas Schulz (1936) identified B. cretica as the progenitor of
of production for 2019 (The Food and Agriculture only cauliflower and broccoli. Helm (1963) proposed a triple
Organization; www.fao.org). Although just six major crop origin in which a single progenitor species gave rise to cauli-
types comprise the majority of the U.S. market (Agricultural flower, broccoli, and sprouting broccoli, whereas kale and
Marketing Service, Market News Reports; www.ams.usda. Brussels sprouts were derived from another unknown wild
gov), outside of these six major cultivars there exists at least species, and that all other crop forms were derived from a
12 additional cultivated crop types (supplementary table S1, third unknown wild species. Snogerup (1980) proposed that
Supplementary Material online). These include lesser known cabbages were derived from wild B. oleracea, kales were de-
varieties such as Chinese white kale or Cantonese gai-lan rived from both B. rupestris and B. incana, and that Chinese
(Mandarin Jiè lan ; var. alboglabra), a leafy vegetable white kale was derived specifically from B. cretica subsp. nivea.
with florets, romanesco (var. botrytis) with unique fractal pat- Due to the lack of consensus on the progenitor species, the
terned curds, and walking stick kale (var. longata), which center of origin for B. oleracea has also remained obscure. One
grows 6–12 feet (1.8–3.7 m) in height. hypothesis is that domesticated B. oleracea originated in
Compared with other crops, surprisingly little is known England from weedy B. oleracea populations, with early cul-
about the progenitor species and origin of domesticated B. tivated forms brought to the Mediterranean, where selection
oleracea. Primary challenges in identifying the progenitor spe- for many of the early crop types occurred (Hodgkin 1995).
cies include the number of wild species that share a single Other studies point specifically to Sicily, which boasts a large
cytodeme and are interfertile with B. oleracea (2n¼18 chro- diversity of wild relatives, as the center of domestication
mosomes; similar genomic organization; referred to as the “C (Schiemann 1932; Lizgunova 1959). This conforms with the
genome”), the corresponding confusion surrounding taxo- observations of Vavilov (1951) that plants tended to be do-
nomic relationships, and conflicting evidence regarding the mesticated in a finite number of global centers of diversity,
center of origin. Wild relatives that share the C genome with which includes the Mediterranean. Most recently, linguistic
domesticated B. oleracea include Brassica bourgeaui, Brassica and literary evidence provided support for domestication in
cretica, Brassica hilarionis, Brassica incana, Brassica insularis, the Eastern Mediterranean, where there is a rich history of
Brassica macrocarpa, Brassica montana, Brassica rupestris, expressions related to the usage and cultivation of B. oleracea
and Brassica villosa. Throughout the literature, many of these crop types in early Greek and Latin literature (Maggioni et al.
species have been referred to by alternative names or have 2010, 2018).
multiple subspecies. For example, B. cretica is described as Using newly generated RNA-seq data for a diversity panel
having either three subspecies (subsp. aegea, cretica, and of 224 accessions that includes 14 cultivar types and nine wild
2
Evolutionary History of B. oleracea . doi:10.1093/molbev/msab183 MBE
Table 1. Wild Species Which Have Been Proposed as Progenitor with a Fisher strand (FS) value greater than 30 and quality
Species for Brassica oleracea Crop Types. depth (QD) less than 2.0. This recovered 942,357 variants in
Cultivar Wild Relative Author total, with 879,865 variants on chromosomes 1–9 and 62,492
Broccoli B. oleracea Linnaeus
variants on remaining scaffolds. Chromosomal SNPs were
B. oleracea Hedrick (1919) a then filtered to exclude sites with greater than 60% missing
B. oleracea Giles (1941) b data, sites with mean per-sample depth values less than 5, and
B. montana Hegi (1919) indels, resulting in a total of 103,525 SNPs. After a final filtering
B. oleracea (from Italy) Giles (1941) step for linkage disequilibrium (LD), a conservative final data
B. cretica Gates (1953)
B. oleracea and B. alboglabra Song et al. (1990)
set of 36,750 SNPs was generated. For all samples, no mapping
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
Brussels sprouts B. oleracea Linnaeus bias was detected when comparing the percentage of
B. oleracea (western Europe) Gates (1953) uniquely mapped reads across cultivar groups, species, and
B. oleracea (western Europe) Snogerup (1980) sequencing lane (supplementary fig. S1, Supplementary
B. oleracea and B. alboglabra Song et al. (1990) Material online).
Cabbage B. oleracea Linnaeus
B. oleracea de Candolle (1824)
B. oleracea Hedrick (1919) a Phylogeny and Population Clustering Distinguish Wild
B. oleracea Bailey (1930) and Feral Populations
B. montana Hegi (1919) Sampling of B. oleracea cultivars included eight types of kales,
B. oleracea (western Europe) Gates (1953) five types of cabbages, Brussels sprouts, broccoli, cauliflower,
B. oleracea (western Europe) Snogerup (1980)
B. oleracea and B. alboglabra Song et al. (1990)
Romanesco (var. botrytis), and kohlrabi (supplementary table
Cauliflower B. oleracea Linnaeus S2, Supplementary Material online). Together, these culti-
B. oleracea de Candolle (1824) vated types accounted for 188 of the 224 total samples.
B. oleracea Bailey (1930) The remaining 36 samples included previously identified
B. montana Hegi (1919) wild relatives: putatively wild B. oleracea, B. cretica, B. incana,
B. cretica Schulz (1936)
B. oleracea (from Cypus) Giles (1941)
B. montana, B. hilarionis, B. insularis, B. macrocarpa, B. rupest-
B. cretica Gates (1953) ris, and B. villosa. The phylogenetic reconstruction of all 224
B. oleracea and B. alboglabra Song et al. (1990) samples using SNPhylo (Lee et al. 2014) recovered several
B. cretica Tutin et al. (1964) well-supported clades with greater than 70% bootstrap sup-
Kale B. oleracea Linnaeus port, although overall support was generally poor (less than
B. oleracea Hedrick (1919) a
B. oleracea Bailey (1930)
70% bootstrap support), especially along the backbone.
B. montana Hegi (1919) Chinese white kale, broccoli, cauliflower, romanesco, kohlrabi,
B. montana Netroufal (1927) curly kale, Brussels sprouts, B. rupestris, B. macrocarpa, and B.
B. oleracea (western Europe) Gates (1953) insularis were all recovered as monophyletic. Aside from red
B. cretica, B. incana, B. rupestris Snogerup (1980) cabbages, cabbages were also monophyletic, but with only
B. incana and B. insularis Hosaka et al. (1990)
B. oleracea and B. alboglabra Song et al. (1990)
55% bootstrap support. Seven cultivars (collards, tronchuda
Kohlrabi B. oleracea Linnaeus kale, savoy cabbage, perpetual kale, red cabbage, and marrow
B. rupestris Netroufal (1927) cabbage) were found throughout the tree as polyphyletic
Unknown Mediterranean species Gates (1953) assemblages. Several wild samples were recovered within
B. oleracea and B. alboglabra Song et al. (1990) the cultivar clade, including two samples of B. cretica (196,
NOTE.—Specific location is included in parentheses if indicated by the author. 199), one sample of B. montana (222), and all samples of
Brassica oleracea sometimes referred to as B. oleracea var. sylvestris.
a
putatively wild B. oleracea (175, 176, 177; sample names in
Edited observations by Sturtevant in the late 19th century.
b
Referring to Prof Buckman’s experiment.
bold text; fig. 1). We also recovered a group in the cultivar
clade consisting of five samples of three wild species, B. incana
(205, 208, 209), B. villosa (233), and B. cretica (195), labeled
relatives, representing the largest and most diverse collection
“WildC-2” (for wild samples with the C genome). Many of
of this species and its wild relatives to date, we integrate
these “wild” samples also share most or all of their ancestry
phylogenomics, population genomics, ecological niche
with cultivars. At K ¼ 2, in our fastSTRUCTURE analyses (Raj
modeling, archaeological, and literary evidence to clarify the
et al. 2014), samples clustered as either cultivars or wild (fig. 1).
taxonomy, identify the closest living wild relative, and provide We find that two samples of B. incana (204, 207; likely both
insight on the origin of domestication for B. oleracea. from Crimea), which are sister to all cultivated samples, share
Results 100% of their ancestry with cultivated types, as do two sam-
ples of B. cretica (196, 199), one sample of B. montana (222),
Sequencing Depth and SNP Identification and all three samples of putatively wild B. oleracea (175, 176,
RNA sequencing of 224 samples resulted in an average of 177). Together with the placement in the phylogeny, these
88,598,754 reads per sample, with a range of 59,543,560– analyses indicate that these are not truly wild samples, but
151,814,032 reads. The minimum per-sample sequencing represent feral types, defining feral here as either exoferal (a
depth recovered was 9X, with a maximum depth of 12X. domesticated population derived from admixture with either
After mapping reads to the B. oleracea TO1000 genome a divergent population, a wild conspecific, another domesti-
(Parkin et al. 2014), SNPs were filtered to exclude those cated species, or another wild species) or endoferal (a
3Mabry et al. . doi:10.1093/molbev/msab183 MBE
098_capitata
K=2 K=3 ++ K=4 K=5+
a bage
095_capitata
094_capitata
092_capitata
093_capitata
162_capitata
cab
161_capitata
163_capitata
159_capitata
164_capitata
176_oleracea
102_sabauda
179_viridis
savoy
184_viridis
108_ramosa
100_sabauda
101_sabauda
097_capitata
047_medullosa
147_alboglabra
132_alboglabra
144_capitata
collards
190_viridis
192_viridis
194_viridis
187_viridis
185_viridis
183_viridis
182_viridis
193_viridis
178_viridis
096_capitata
134_alboglabra
189_viridis
160_capitata
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
099_sabauda
091_capitata
Brussels spprouts
022_gemmifera
020_gemmifera
016_gemmifera
018_gemmifera
011_gemmifera
021_gemmifera
015_gemmifera
017_gemmifera
013_gemmifera
010_gemmifera
014_gemmifera
009_gemmifera
abbage
024_gemmifera
023_gemmifera
019_gemmifera
008_gemmifera
012_gemmifera
marrow cab
196_cretica
113_ramosa
043_medullosa
t al kalle
036_medullosa
038_medullosa
222_montana
175_oleracea
116_ramosa
115_ramosa
110_ramosa
r etu
046_medullosa
106_ramosa
045_medullosa
041_medullosa
perp
104_ramosa
114_ramosa
040_medullosa
050_sabellica
currly kale
055_sabellica
048_sabellica
049_sabellica
053_sabellica
051_sabellica
058_sabellica
052_sabellica
057_sabellica
056_sabellica
059_sabellica
054_sabellica
177_oleracea
191_viridis
186_viridis
181_viridis
063_italica
112_ramosa
174_palmifolia
078_gongylodes
080_gongylodes
087_gongylodes
a i
kohlrab
079_gongylodes
083_gongylodes
082_gongylodes
088_gongylodes
081_gongylodes
077_gongylodes
086_gongylodes
084_gongylodes
089_gongylodes
090_gongylodes
085_gongylodes
f ower
124_botrytis
118_botrytis
122_botrytis
167_botrytis
121_botrytis
152_botrytis
caulifl
128_botrytis
126_botrytis
125_botrytis
157_botrytis
119_botrytis
117_botrytis
123_botrytis
120_botrytis
060_italica
127_botrytis
065_italica
075_italica
170_italica
076_italica
168_italica
070_italica broccoli
069_italica
062_italica
067_italica
073_italica
071_italica
072_italica
066_italica
romanesco
064_italica
074_italica
169_italica
061_italica
153_botrytis
154_botrytis
156_botrytis
155_botrytis
Chinese white kale
068_italica
148_alboglabra
131_alboglabra
142_alboglabra
151_alboglabra
140_alboglabra
135_alboglabra
136_alboglabra
143_alboglabra
149_alboglabra
138_alboglabra
133_alboglabra
141_alboglabra
146_alboglabra
130_alboglabra
137_alboglabra
145_alboglabra
129_alboglabra
150_alboglabra
tronchuda kale
199_cretica
031_costata
105_ramosa
025_costata
028_costata
172_longata
034_costata
026_costata
030_costata
029_costata
033_costata
139_alboglabra
171_costata
027_costata
165_costata
035_medullosa
032_costata
180_viridis
039_medullosa
037_medullosa
WildC-2
103_ramosa
044_medullosa
209_incana
208_incana
233_villosa
205_incana
lacinato kale
195_cretica
111_ramosa
107_ramosa
004_palmifolia
173_longata
006_palmifolia
003_palmifolia
005_palmifolia
002_palmifolia
001_palmifolia
007_palmifolia
042_medullosa
109_ramosa
207_incana
204_incana
203_hilarionis
198_cretica
215_insularis
213_insularis
212_insularis
216_insularis
214_insularis
221_macrocarpa
219_macrocarpa
218_macrocarpa
220_macrocarpa
224_montana
232_rupestris
227_rupestris
228_rupestris
226_rupestris
231_rupestris
229_rupestris
237_villosa
236_villosa
234_villosa
230_rupestris
238_villosa
FIG. 1. Demographics and population structure for 224 samples of cultivated Brassica oleracea (n ¼ 188) and wild C genome species (n ¼ 36). (Left)
Individual sample phylogeny with putatively wild samples labeled in bold and black dots indicating bootstrap values less than 70%. (Middle)
Ancestry proportions for K ¼ 2 to K ¼ 5 as inferred from fastSTRUCTURE; K ¼ 3 maximizes marginal likelihood (þþ) and K ¼ 5 best explains
structure in the data (þ). (Right) Monophyletic clades indicated by a solid line, largest cluster of paraphyletic groups indicated by dashed lined.
Illustrations of corresponding crop types by Andi Kur.
4Evolutionary History of B. oleracea . doi:10.1093/molbev/msab183 MBE
population of domesticated plants that has escaped from Domestication Is Also Reflected in the Transcriptome
cultivation without the aid of introgression/hybridization Using expression profiles (transcript abundances) of 51,438
with wild conspecifics; Gering et al. 2019). Our newly identi- genes for our original 224 samples, we tested if cultivars and
fied WildC-2 shows mixed wild and cultivar ancestry, which wild samples would still cluster separately based on the tran-
was also observed for one sample of tronchuda kale (30). The scriptome. Overall, results and clustering patterns were sim-
marginal likelihood was maximized at K ¼ 3, in which a clus- ilar to analyses using SNPs, with the axes of PC1 and PC2
ter comprised of broccoli, cauliflower, and Chinese white kale separating most wild species from cultivars (fig. 2C; supple-
separated from other cultivated types. At K ¼ 4, Chinese mentary fig. S3B and C, Supplementary Material online). We
white kale was distinct from broccoli and cauliflower. The again found the same samples of B. incana (204, 207), B.
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
structure in the data was best explained by K ¼ 5, in which cretica (196, 199), B. montana (222), and B. oleracea (175,
the clade comprised of B. insularis and B. macrocarpa was 176, 177) clustering with the cultivars, but in expression anal-
separated and had shared ancestry with Brassica cretica (198), yses WildC-2 clustered with the other wild samples, rather
than separately as in our SNP based PCA. Hierarchical clus-
B. hilarionis, B. montana (224), and one sample of tronchuda
tering of the expression profiles recovered similar patterns
kale (30). Additional K values showed similar patterns (sup-
with two major groups: wild and cultivated, again with
plementary fig. S2, Supplementary Material online).
WildC-2 clustering with the other wild samples (supplemen-
Principal component analysis (PCA) also separated culti-
tary fig. S5, Supplementary Material online). Although most
vars from most wild samples (supplementary fig. S3D–F,
cultivar groups were not recovered as unique clusters, there
Supplementary Material online). The PC1 axis distinguishes were a few exceptions. Brussels sprouts, Chinese white kale,
wild species from cultivars and the PC2 axis separates WildC- and curly kale all formed distinct clades, which corresponds to
2 from all other wild species (triangles with black outlines). what we know about their growth habit. Since RNA was
Although one sample of B. cretica (198) clusters closest to collected at the 7th leaf-stage, before substantial morpholog-
cultivated types, samples of B. incana, which were not in ical differentiation occurs between cultivars, it is not too sur-
WildC-2, along with one sample of B. montana (222), two prising that they do not cluster distinctively by cultivar. Curly
samples of B. cretica (196, 199), and all three samples of B. kale is almost immediately visually distinguishable from other
oleracea (175, 176, 177) cluster with the cultivars, corroborat- cultivars in that the first true leaves have margins which are
ing the phylogenetic analyses. To further investigate the clus- already undulate and/or frilled, in contrast to the more lan-
tering patterns of B. cretica to cultivars, we included four ceolate (i.e., long, widest in the middle, with tapered tips)
additional wild-collected samples of two B. cretica subspecies leaves observed in most cultivars. Brussels sprouts are also
(A and B¼subsp. nivea, C and D¼subsp. cretica; fig. 2A and B; easily identifiable at this early growing stage as they have
supplementary fig. S3A, Supplementary Material online; la- short, oblong to nearly circular leaves. Although Chinese
beled SRA in figure legend; Kioukis et al. 2020). Adding these white kale leaves look more similar to the lanceolate shape
samples supports the results of other studies that B. cretica, as of other cultivars, they grow more rapidly and plants in this
a species, is very diverse. Although sample C does not group group are annual instead of biennial, which may explain why
with other B. cretica samples using the PC1 axis, the PC2, PC3, these accessions cluster separately from other cultivars.
and PC4 axes show much tighter clustering among the four To identify modules of genes that might be driving the
wild-collected samples and one of our samples of B. cretica observed clustering patterns, we used weighted correlation
(198), indicating that our B. cretica (198) sample is an infor- network analysis (WGCNA; Langfelder and Horvath 2008).
mative representative of wild-collected B. cretica (fig. 2A and We found that 48 modules, ranging in size from 34 to
B; supplementary fig. S3A, Supplementary Material online). 35,981 genes, provided the best fit for the data (supplemen-
For crop samples, estimates of inbreeding coefficients from tary table S3, Supplementary Material online). To assess what
PCAngsd (Meisner and Albrechtsen 2018) roughly matched types of biological processes were overrepresented in these
modules, we used syntenic Arabidopsis thaliana genes and
expectations for the frequency of heterozygotes under
performed a GO analysis through PANTHER v. 16.0 (Mi et al.
Hardy–Weinberg equilibrium, whereas inbreeding coeffi-
2021). Overlap of B. oleracea with A. thaliana genes ranged
cients for wild species suggest excess homozygosity (supple-
from 17% to 98.3%, perhaps indicating that some modules are
mentary fig. S4, Supplementary Material online), possibly more conserved, whereas others are unique to B. oleracea.
reflecting cultivation practices for germplasm management Modules which were more conserved between the two spe-
and the relative isolation of wild populations (i.e., small effec- cies included genes related to herbivory defense compound
tive population size), respectively. Feral samples, those which production (secondary metabolite biosynthetic process, phe-
were identified as wild taxa, but were found more closely nylpropanoid biosynthetic and metabolic processes), wound
related to cultivars than to wild taxa in our phylogeny and formation (suberin biosynthetic processes), and wax forma-
clustered with cultivated samples in our PCA (B. cretica—196, tion (wax biosynthetic and metabolic processes), the latter of
199; B. incana—204, 207; B. montana—222; and wild B. oler- which may be correlated to the characteristic glaucous leaves
acea—175, 176, 177), show patterns of heterozygosity that are of cultivated B. oleracea (supplementary table S4,
similar to crop samples, as do the four samples of B. cretica Supplementary Material online). Within the top five con-
from Kioukis et al. (2020). Our WildC-2 exhibited patterns of served modules, the transcript abundance (TPM) was signif-
excess homozygosity more similar to other wild taxa. icantly different among the different groups (P value2e-16
5Mabry et al. . doi:10.1093/molbev/msab183 MBE
A Genetic Variation Between Wild and Cultivars : PC1 vs PC2
C
Cultivars
198 D B. cretica
1.3 % of genetic variance explained
0.1 B B. hilarionis
A
Principal Component 2
B. incana
B. insularis
199
B. macrocarpa
0.0 207 B. montana
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
196 B. oleracea
204
B. rupestris
222
B. villosa
Cultivar
209
195 WildC-2
−0.1
Wild
233
208 205
SRA
−0.1 0.0 0.1 0.2
Principal Component 1
18.3 % of genetic variance explained
B Genetic Variation Between Wild and Cultivars : PC2 vs PC3
Cultivars
B. cretica
1.13 % of genetic variance explained
0.1
B. hilarionis
Principal Component 3
205 233 B. incana
208 195 209 B. insularis
222 196
0.0
B. macrocarpa
204 207 199
A B. montana
D C
B 198 B. oleracea
−0.1
B. rupestris
B. villosa
Cultivar
Wild WildC-2
SRA
−0.1 0.0 0.1
Principal Component 2
1.3 % of genetic variance explained
C Expression Variation Between Wild and Cultivars : PC1 vs PC2
80
Cultivars
B. cretica
7 % of genetic variance explained
B. hilarionis
Principal Component 2
40
B. incana
208 B. insularis
204
207
198 233 B. macrocarpa
195
0
199 205
222 B. montana
196 B. oleracea
209 B. rupestris
−40
B. villosa
Cultivar
Wild WildC-2
−50 0 50 100
Principal Component 1
12 % of genetic variance explained
FIG. 2. Principal component analysis (PCA) of SNPs and expression profiles. (A) Genetic variation PCA of PC1 versus PC2, (B) Genetic variation PC2
versus PC3, and (C) Expression profile PCA for PC1 versus PC2 of wild and cultivar samples. Triangles, wild samples; circles, cultivars; triangles with
black outlines, WildC-2 samples with species identification indicated by color. Wild-collected B. cretica samples from Kioukis et al. (2020) indicated
by asterisks, labeled as SRA.
6Evolutionary History of B. oleracea . doi:10.1093/molbev/msab183 MBE
for modules 7, 13, 31, & 34; P value¼2.19e-11 for module 30). cabbage]. To further test this event, we used four-population
Post hoc comparisons using Tukey’s honestly significant dif- (f4) tests for treeness as implemented in TreeMix, where a
ference (HSD) revealed that transcript abundance in cultivars significant nonzero value indicates the presence of gene flow
was significantly different compared with that of wild relatives (Reich et al. 2009; Pickrell and Pritchard 2012; fig. 4B).
across conserved modules, except for B. hilarionis, which was Although the tree [[tronchuda cabbage, kohlrabi],[B. cretica
not recovered as significantly different from cultivars for any (198), B. hilarionis]] showed no significant evidence of gene
module. WildC-2 along with other identified feral samples flow (f4 ¼ 0.0008, Z ¼ 1.094), replacing tronchuda cabbage
had significantly different transcript abundance compared with Chinese white kale indicated significant gene flow
with cultivars for modules 7, 13, 31, and 34, but not for mod- from B. cretica (198) to Chinese white kale (f4¼0.0055, Z
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
ule 30. Significant differences were also found between ¼5.113). This result was further verified when adding a
WildC-2 and feral samples compared with wild relatives for second migration edge, as the migration edge only included
several modules, with no obvious patterns across modules Chinese white kale, but the direction was reversed (from
(supplementary fig. S6 and table S5, Supplementary Material Chinese white kale to B. cretica [198]). The second event,
online). from kohlrabi to a presumably feral sample of B. cretica
(199), was supported by f4 tests, with the tree [[kohlrabi, B.
Species Tree and Admixture Inference Indicate cretica (196)],[B. cretica (199), marrow cabbage]] indicating
Brassica cretica Is the Closest Living Wild Relative significant evidence of gene flow from kohlrabi to B. cretica
Given the results of population clustering using both SNPs (199) (f4 ¼ 0.012, Z ¼ 10.5). This migration event is also seen
and expression profiles, we further interrogated the species phenotypically, as B. cretica (199) has a swollen stem when
level relationships between wild relatives and cultivar groups grown to maturity. No significant evidence of gene flow was
by resolving the backbone of the phylogeny. Using the PoMo found when substituting B. cretica (199) with B. oleracea
model (Schrempf et al. 2016) as implemented in IQ-Tree (175), which is not expected to be involved in the admixture
(Nguyen et al. 2015) and only including samples representing event (f4 ¼ 0.00023, Z ¼ 2.68). Two admixture events provide
monophyletic groups as determined in the sample-level phy- evidence of potential exoferal origins for at least two samples,
logeny, we found strong support for B. cretica as the closest B. oleracea (175) and B. cretica (199). The four-population tree
living wild relative to cultivated B. oleracea (fig. 3A). Notably, of [[B. montana (222), curly kale],[B. oleracea (175), broccoli]]
for our species tree analyses, we included only one sample of suggests significant gene flow from B. montana (222) to B.
B. cretica (198). This sample was used for species reconstruc- oleracea (175) (f4 ¼ 0.315, Z ¼ 15.77), as does the tree of
tion due to its placement near other wild taxa in the sample [[tronchuda cabbage, Chinese white kale],[B. cretica (199),
level phylogeny and its clustering with wild-collected B. cretica broccoli] for gene flow from Chinese white kale to B. cretica
from Kioukis et al. (2020) in the PCA. The current distribution (199) (f4 ¼0.009, Z ¼7.98). The fifth added migration
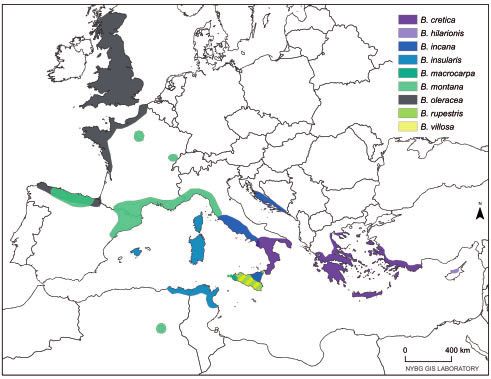
of B. cretica occurs throughout the Eastern Mediterranean, edge from B. rupestris to WildC-2 explains the shared ancestry
primarily in Greece, highlighting a potential origin of domes- recovered in the fastSTUCTURE results. The test for treeness
tication (fig. 3B). Another suggested wild relative, B. incana, is with [[curly kale, WildC-2],[B. rupestris, B. macrocarpa]] indi-
strongly supported as belonging to the cultivar clade, sister to cated significant admixture from B. rupestris to WildC-2 (f4
lacinato kale. Although our sampling is limited in regard to ¼0.006, Z¼6.50), but was nonsignificant when substitut-
the distribution of B. incana as a whole, this result supports ing WildC-2 with cauliflower (f4 ¼0.0003, Z ¼0.338). In
our other findings that B. incana is not a completely wild general, these analyses highlight that the evolutionary history
assemblage, but that at least some populations are feral. of B. oleracea is characterized by many admixture events and
Within cultivars, several expected relationships were recov- lineages of exoferal origins.
ered: collards and cabbage as sister lineages (Song et al. 1988;
Farnham 1996), with Brussels sprouts sister to both; cauli- Archaeological and Literary Evidence Point to a Late-
flower and broccoli as sister clades (Song et al. 1988; Holocene Domestication
Stansell et al. 2018), with romanesco sister to both; and To further investigate the origins of domesticated B. oleracea,
Chinese white kale as sister to all other cultivars, agreeing we surveyed archaeological, literary, and artistic evidence
with recent literature (Cheng et al. 2016; Stansell et al. 2018). (supplementary tables S6 and S7, Supplementary Material
With the overall species relationships resolved, we aimed online). The earliest reported claim of B. oleracea comes
to tease apart the evolutionary history of the wild samples from an archaeological collection from the Austrian Alps.
that clustered within the cultivar clade. Specifically, we asked This collection comprises three seeds dated to the Middle
if any of the identified feral samples were the products of Bronze Age (ca. 3550–3350 years before present or BP;
admixture using TreeMix (Pickrell and Pritchard 2012). Schmidl and Oeggl 2005). However, the lack of illustrations
Although the tree model without any migration edges and discussion of separation criteria from other Brassica spe-
explained 87.3% of the variance in the data set, sequentially cies makes us question the reliability of this species-level iden-
adding migration events to the tree resulted in five migrations tification, as seeds of Brassica species are difficult to tell apart.
events explaining 92% of the variation (fig. 4A and supple- The only other find of similar antiquity is B. oleracea seeds
mentary fig. S7, Supplementary Material online). Adding a from the Late Bronze Age/Early Iron Age, identified by scan-
single migration edge resulted in an admixture event from ning electron microscopy and radiocarbon dated directly be-
B. cretica (198) to a clade of [Chinese white kaleþtronchuda tween ca. 3250–2970 BP (Kaniewski et al. 2011). These finds
7Mabry et al. . doi:10.1093/molbev/msab183 MBE
A 100
100
Brussels sprouts
cabbage
B
100 collards
marrow cabbage
100
100
curly kale
93
lacinato kale
99
B. incana
Cultivars kohlrabi
91
100
cauliflower
83 100 broccoli
100
romanesco
tronchuda cabbage
100
Chinese white kale
100
B. cretica
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
100
B. hilarionis
100
B. montana
100
B. insularis
B. macrocarpa
B. rupestris
B. villosa
C Habitat
D
Suitability Habitat
Suitability
High
High
Low
Low
FIG. 3. Species tree with current distribution and historical environmental niche modeling. (A) Species tree of wild and cultivar samples. Bootstrap
support indicated above branches. (B) Current species distribution of wild relatives. (C) Suitable habitat for B. cretica and (D) B. hilarionis during the
late-Holocene. Map of current distrubution provided by Elizabeth Gjieli, the Geographical Information Manager at the New York Botanical Garden
GIS Laboratory.
A B_villosa
B_macrocarpa
B
curly_kale
wildC-2
[[tronchuda cabbage, kohlrabi]; [B. cretica (198), B. hilarionis]]
B_cretica_199 a [[kohlrabi, Chinese white kale]; [B. cretica (198), B. hilarionis]]
B_montana_222
c. [[B. cretica(198), B. hilarionis]; [broccoli, Chinese white kale]]
lacinatio_kale
romanesco
cauliflower [[kohlrabi, B. cretica (196)]; [B. oleracea (176), marrow cabbage]]
broccoli
b [[kohlrabi, B. cretica (196)]; [B. cretica(199), marrow cabbage]]
tronchuda_cabbage
Chinese_white_kale
b. [[B. montana (222), curly kale]; [cauliflower, broccoli]]
Migration
kohlrabi
cabbage
c [[B. montana (222), curly kale]; [B. oleracea (175), broccoli]]
weight
Brussels_sprouts
0.5 a.
collards
[[tronchuda cabbage, Chinese white kale]; [cauliflower, broccoli]]
e. B_cretica_196 d [[tronchuda cabbage, Chinese white kale]; [B. cretica (199), broccoli]]
B_oleracea_176
B_oleracea_175
marrow_cabbage [[curly kale, WildC-2]; [B. rupestris, B. macrocarpa]]
0 B_oleracea_177 e [[curly kale, cauliflower]; [B. rupestris, B. macrocarpa]]
B_incana
B_cretica_198
B_insularis
f. [[B. montana, B. insularis]; [B. cretica (198), broccoli]]
B_montana f [[B. montana, B. hilarionis]; [B. cretica (198), broccoli]]
10 s.e. B_hilarionis
B_rupestris
−0.02 0.00 0.02
0.00 0.01 0.02 0.03 0.04 0.05 0.06 0.07 F4
Drift parameter
FIG. 4. Inferred admixture events. (A) Phylogeny five migrations labeled a–f. (B) Corresponding four-population tests for treeness.
are associated with destruction levels at Gibala, Tell Tweini in commonly eaten Brassica leaves in England at this time, this
western Syria on the Mediterranean coast. Although most of would appear a likely identification.
the archaeological finds are of seeds (supplementary table S6, The earliest literary references to B. oleracea date to Greek
Supplementary Material online), there is at least one docu- scholars 2500–2000 BP (supplementary table S7,
mentation of pottery residues where lipids of Brassica leaf Supplementary Material online). Hipponax’s writing refers
waxes were identified and dated to 850–750 BP (Evershed to a seven-leaf cabbage in an iambic verse (West 2011),
et al. 1992, 1994). The authors attribute this to the boiling of whereas Hippocrates On the Nature of Women, written
leaves of B. oleracea, and given the lack of evidence for other around 2410–2320 BP, refers to the use of cabbage, or
8Evolutionary History of B. oleracea . doi:10.1093/molbev/msab183 MBE
krambe, in a few recipes (Totelin 2009). As early as 2320 BP, these species being likely progenitor species of B. oleracea
there is evidence for cultivar diversity. Theophrastus refers to cultivars. In our species tree reconstructions, we find just B.
three varieties: a curly-leaved type, a smooth-leaved type, and cretica as sister to all cultivars, specifically sample 198, which
a wild type with a bitter taste, many branches, and many clusters with wild-collected B. cretica samples from Kioukis et
small round leaves (Yonge 1854). Pliny in his Natural al. (2020) in our PCA (fig. 2A and B), lending further support
History writing some 200 years later describes at least ten for B. cretica as the progenitor species. This same sample of B.
varieties in addition to those seen in the previous classical cretica (198) as well as our sample of B. hilarionis are recov-
works (The Elder and Rackham 1950). However, whereas ered as fairly homozygous, therefore they would likely be
most scholars accept that the Greek or Latin translations of good starting material for future research related to de
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
“cabbage” refer to B. oleracea, it is important to note that novo domestication via selective breeding or gene editing.
“cabbage” is not a Greek word and that the word “raphanos” Although we do recover evidence of admixture between B.
is translated as both cabbage and B. cretica in the Greek- cretica (198) and both wild and cultivated taxa, the place-
English Lexicon (Liddell and Scott 1940) and in Hort’s ment of B. cretica (198) as the closest living wild relative does
(1916) translation of Theophrastus’ Historia Plantarum. not change. However, an inferred admixture event from B.
Certainly, there are differences between the subspecies of B. cretica (198) to B. hilarionis does result in a topological change
cretica that might be reflective of the varieties described by in the placement of B. hilarionis as sister to B. montana (224;
Theophrastus and Pliny, and which may explain the diversity originally collected in Spain) (supplementary fig. S7,
we observed among B. cretica samples in our PCA results. Supplementary Material online). This novel relationship has
Further, the description by Nicander (quoted by Athenaeus; not been identified before and warrants additional study with
Yonge 1854; p. 582) indicates that wild or perhaps feral forms greater taxon sampling. The second migration event involving
of B. cretica were known in Ionia, the western coast of B. cretica (198) is from Chinese white kale. This event lends
present-day Turkey, ca. 2150–2050 BP. further evidence of admixture with wild germplasm during
the domestication process, consistent with other examples
Late-Holocene Environmental Niche Modeling demonstrating that domestication is not a single event, but a
Highlights Wild Relatives’ Ranges series of events characterized by continuous gene flow be-
Based on archaeological information, the oldest relatively re- tween wild and cultivated populations (Beebe et al. 1997;
liable occurrence for B. oleracea cultivation is dated 3250– Wang et al. 2017). Together with the phylogeographic dis-
2970 BP in Gibala NW Syria (Kaniewski et al. 2011). To predict continuity of wild B. oleracea samples and their Eastern
what would be a suitable habitat for the wild relatives during Mediterranean progenitors (fig. 3B), the more distant phylo-
the late-Holocene, we compiled occurrence records from genetic placement of B. insularis, B. macrocarpa, and B. villosa
GBIF (www.gbif.org) and (Snogerup et al. 1990), along with (fig. 3A), and strong patterns of shared ancestry between B.
environmental data, to perform environmental niche model- incana and cultivars (fig. 1), these results lead us to support
ing using MaxEnt 3.4.1 (Phillips et al. 2017). Notably, we find the hypothesis of domestication in the Eastern
that B. cretica has an expanded Eastern Mediterranean hab- Mediterranean with B. cretica as the closest living wild relative.
itat suitability (fig. 3C) that includes Cyprus. Presently, only B.
The Role of Ferality in the Domestication of Brassica
hilarionis is known to occur in Cyprus (fig. 3B), however
modeling predicts that in the late-Holocene it would also
oleracea
Multiple lines of evidence highlight the role of wild and feral
have had an expanded habitat suitability in the surrounding
populations as pools of diversity that contributed to crop
mainland coastal regions (fig. 3D). Since most of these wild
diversification during domestication (Beebe et al. 1997;
species are narrow island endemics (Snogerup et al. 1990),
Allaby 2010; Fuller et al. 2014; Wang et al. 2017). Our data
species are generally estimated to have little change from
support a similar phenomenon in the domestication of B.
current day distributions (supplementary fig. S8 and table
oleracea: it appears that introgression from wild or feral pop-
S8, Supplementary Material online).
ulations contributed to the genetic composition of particular
Discussion crops, and vice versa, which is revealed by in-depth analyses of
admixture using population structure and tree-based meth-
Multiple Lines of Evidence Support a Single Eastern ods (figs. 1 and 4; supplementary fig. S2 and S7,
Mediterranean Origin Supplementary Material online). Several samples of wild rel-
Our evidence from genome-scale, multilocus data along with atives, including B. cretica, as well as wild B. oleracea, B. incana,
archeology, literature, and environmental niche modeling B. montana, and B. villosa, are recovered as feral in all analyses.
best support a single Eastern Mediterranean domestication Although we find one sample of B. cretica (198) as the
origin for B. oleracea, corroborating the conclusions of closest living wild relative, we also identify two samples of
Maggioni et al. (2018) based on literary sources and B. cretica (196 and 199) as likely feral and fall within the
(Maggioni et al. 2010) using linguistics. When modeling phy- cultivar clade (fig. 1; see supplementary fig. S9,
logeny and population structure, two Eastern Mediterranean Supplementary Material online, for photos). Interestingly,
species, B. cretica and B. hilarionis, are found as sister species Song et al. (1988) also recovered a polyphyletic B. cretica using
to cultivars and are assigned ancestry from all cultivar pop- RFLPs. Results presented here support previous findings that
ulations for values of K from 2 to 5 (fig. 1), consistent with B. cretica was at one point at least partially domesticated.
9Mabry et al. . doi:10.1093/molbev/msab183 MBE
Snogerup et al. (1990) state that wild B. cretica was consumed attributed to the spread of crops both within and on the
as late as 1962 and, as noted in our literary results, some early periphery of the Roman Empire (Van der Veen 2011).
references to B. oleracea in the literature could be translated Additionally, several studies that sampled wild B. oleracea
as B. cretica, meaning the vast amount of described morphol- populations in the British Isles (Mitchell 1976; Mitchell and
ogy in these works, which may be the result of cultivation, Richards 1979), South West England (Raybould et al. 1999),
could now be reflected in the multiple named subspecies and Atlantic coasts of western Europe (Mittell et al. 2020), and
described genetic diversity of modern B. cretica (Snogerup et Atlantic coast of France (Maggioni et al. 2020) support that
al. 1990; Widen et al. 2002; Allender et al. 2007; Edh et al. these wild B. oleracea populations are feral populations, typ-
2007). Further, B. cretica was known to occur in Ionia (western ically with low levels of genetic diversity and some degree of
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
coast of present day Turkey) ca. 2150–2050 BP and the evi- isolation from other populations. Lanner-Herrera et al. (1996)
dence of B. cretica populations today in Lebanon, which are sampled populations across Spain, France, and Great Britain,
morphologically similar to B. cretica subsp. nivea, suggests concluding that each population evolved independently,
widespread trade of these species by the earliest whereas more recently, Mittell et al. (2020) found that geo-
Mediterranean civilizations (Dixon 2006). However, these graphically close populations were more genetically different
plants may have been introduced into these localities without than distant populations. Our results provide additional evi-
cultivation as was proposed by Snogerup et al. (1990). dence that feralization is commonplace for B. oleracea crops
Previous researchers have noted that B. cretica populations and that references to wild B. oleracea likely represent multi-
are typically found in coastal locations associated with an- ple, independent feralization events. Additional sampling of
cient seaports, occupying their preferred ecological niche on wild populations will enable opportunities to further investi-
chalk cliffs undisturbed by grazing (Mitchell 1976; Snogerup et gate the relationships among these feral populations and
al. 1990). We believe that these early forms of B. cretica may cultivated crops.
have played underappreciated roles in the domestication of B. Brassica incana, another suggested progenitor species
oleracea crops and to fully understand the evolutionary his- (Snogerup 1980), is also supported as feral for the samples
tory of B. oleracea, the demographic history and domestica- included in our analyses. Two of our five samples (204 and
tion story of B. cretica must be resolved. 207) are recovered as sister to all cultivars in our individual
Sources have hypothesized that wild populations of B. level phylogeny but are found to share 100% of their ancestry
oleracea in England are the progenitor(s) for modern cultivars with cultivars rather than other wild taxa using
(Snogerup et al. 1990; Song et al. 1990), whereas others have fastSTRUCTURE when K ¼ 2 (fig. 1). Further, these two sam-
proposed that these are escaped cultivars (Mitchell 1976; ples were resolved as sister to lacinato kale in our species tree
Mitchell and Richards 1979). Consistent with these hypothe- analysis, providing additional evidence that these samples
ses, we find that the three wild B. oleracea samples in our represent a feral lineage, possibly of lacinato kale. This result
study cluster with cultivars both phylogenetically and in PCA may lend insight into why previous studies have found B.
for both SNP data and expression profiles. Although these incana as sister to B. oleracea (Lazaro and Aguinagalde
samples are from Canada (175), Denmark (176), and 1998; Mei et al. 2010; Arias and Pires 2012) and the observa-
Germany (177), well outside the natural distribution range tion by Snogerup et al. (1990) that samples of B. incana from
for B. oleracea—notably not from England, one of the hypoth- the Crimea are more interfertile with cultivated B. oleracea
esized geographic origins—we suggest that an origin in than others. Although Snogerup et al. (1990) suggested that B.
England is unlikely given the archeological and literary data. incana was more interfertile due to historical introgression,
Although the oldest archaeobotanical record for B. oleracea we do not find evidence of this for samples 204 and 207.
(Middle Bronze Age; ca. 3550–3350 BP) is from Austria, we However, the three other samples of B. incana (205, 208,
regard this evidence with caution as wild populations of B. 209), which belong to WildC-2, do show evidence of admix-
oleracea are not presently found in Austria and the major ture with B. rupestris, likely explaining their clustering to-
Brassica crops in this region include B. nigra (Tutin et al. 1964) gether both in the PCAs and phylogeny with B. cretica
or potentially cultivated turnip (B. rapa). Additionally, there is (195) and B. villosa (233) which also show admixture with
no compelling archaeological evidence to suggest the possible B. rupestris (see supplementary, fig. S10, Supplementary
cultivation of cabbages in Europe prior to the Late Iron Age Material online, for photos). All three B. incana were collected
(2350–2050 BP) and Roman periods (1950–1650 BP), but in Italy from two locations and therefore do not well repre-
there is evidence for knowledge of B. oleracea in Greece dur- sent the known B. incana range (fig. 3B), whereas the two
ing this time (Maggioni et al. 2018; supplementary tables S6 other samples found in this clade, B. cretica (195) and B. villosa
and S7, Supplementary Material online). Overall, there are no (233), were collected in Greece and Italy, respectively.
records for B. oleracea from before this period within data- Although all five WildC-2 samples share an introgression
bases relating to the Eastern Mediterranean (Reihl 2014), event from B. rupestris (figs. 1 and 4; supplementary fig. S7,
Europe (Kroll 2001, 2005), Britain (Tomlinson and Hall Supplementary Material online), they are from different
1996), the Czech Republic (Kreuz and Sch€afer 2002), or within germplasm collections (IPK-gatersleben and USDA National
predynastic and Pharaonic Egypt (Murray 2000), despite hav- Plant Germplasm System), ruling out the inferred migration
ing documentation for other Brassica species. Evidence for B. being the result of current cultivation practices. It is possible
oleracea in Europe does not start appearing until ca. 1850 BP, that at least three of these samples (B. incana 205, 208, 209)
when the appearance of seeds increased and can be are related to the wild kale of Crimea, which is posited as a B.
10Evolutionary History of B. oleracea . doi:10.1093/molbev/msab183 MBE
rupestris–incana hybrid that was transferred to the Crimea via 2018). A review of Chinese historical sources concluded that
trade (Dixon 2006). This suggests that there was early wide- B. oleracea may have been introduced to China 1450–1350 BP
spread cultivation of these B. rupestris–incana types (Dixon and had evolved into Chinese white kale in Southern China
2006) and provides a plausible explanation for why B. incana by the period of the Tang Dynasty (1350–1250 BP; Zhang
and B. rupestris are closely related in previous studies (Lanner 2009). Due to its position as sister to all other cultivars and as
et al. 1997; Mei et al. 2010). The other two samples in WildC-2 the only Asian B. oleracea crop type, as well as its annual
(B. cretica 195 and B. villosa 233), possibly represent misiden- growth habit, this taxon warrants additional study to under-
tifications, which is supported by their intermediate pheno- stand its own unique domestication story.
types (i.e., B. rupestris margins with varying amounts of The dispersal of B. oleracea by human translocation west-
Downloaded from https://academic.oup.com/mbe/advance-article/doi/10.1093/molbev/msab183/6304875 by guest on 11 August 2021
trichomes; supplementary fig. S10, Supplementary Material ward, ultimately to the Atlantic coast of Europe, appears to
online). have established both regional feral populations and the va-
The last feral identification is that of B. montana, for which riety of modern crop types. Archaeological evidence suggests
we find one sample as more closely related to wild taxa (224) that this process may have begun with Late Bronze Age sea-
and one more closely related to cultivars (222). The feral faring (3000–3300 years ago), when the whole Mediterranean
sample (222) is of unknown origin, but again the literature became linked in trade perhaps for the first time (Broodbank
indicates that this may not be a surprising result. Many stud- 2015), and continued to provide a corridor for introgression
ies have previously indicated a close relationship between B. and varietal diversification through the Iron Age (up to
montana and B. oleracea. For example, Panda et al. (2003) 2000 years ago). Trade links along the Atlantic seaboard
concluded that B. montana may be a subspecies of B. oleracea, from North Africa and Iberia through Britain and Ireland
whereas Lanner et al. (1997) found that B. montana and B. are clearly indicated in archaeology (Cunliffe 2004), and are
oleracea clustered together using chloroplast data. associated with the first peopling of the Canary Islands from
Furthermore, several authors have suggested that some pop- the north, where walking stick kale is endemic. Notably, many
ulations of B. montana were feral B. oleracea (Paolucci 1890; cultivars do not form monophyletic groups in our sample
Onno 1933; Snogerup et al. 1990), which may be reflected in level phylogeny, likely indicative of admixture between crop
the overlapping ranges produced by our niche modeling of types. This is supported by previous findings that broccoli is
these two species (supplementary fig. S8, Supplementary paraphyletic (Song et al. 1988; Stansell et al. 2018), as well as
Material online). Therefore, in combination with results collards (Pelc et al. 2015), and by our findings that kale types
from previous studies, our results support that at least such as tronchuda kale and perpetual kale are highly poly-
some B. montana populations are of feral origin. phyletic, suggesting that the kale morphotype has been se-
Taken together, it is clear that the current taxonomy of B. lected for multiple times independently.
oleracea and its wild relatives is confounded by gene flow In conclusion, we confirm a single Eastern Mediterranean
between wild and cultivated populations, resulting in confu- origin for B. oleracea and find B. cretica as the closest living
sion between wild and feral lineages and obscuring the true wild relative. We highlight several feral samples that are not
evolutionary history of this species. Additionally, although reflected by the current taxonomy but likely reflect important
there is much interest in crop improvement using CWRs aspects of the domestication history for B. oleracea. Moving
(Meyer et al. 2012; Khoury et al. 2020), feral lineages offer forward, it will be important to identify, collect, study, and
another, potentially more direct route to reintroducing ge-
preserve these feral samples as pools of allelic diversity, which
netic diversity into cultivated populations, as gene flow is less
may play an important role in future crop improvement, for
likely to be impeded by barriers such as reproductive isolation
example, as a source of potential pest and pathogen resis-
(Mabry et al. 2021). These feral populations may also provide
tance (Mithen et al. 1987; Mithen and Magrath 1992;
additional avenues to explore the evolutionary capacity for
Mohammed et al. 2010). In clarifying the evolutionary history
range expansion and phenotypic plasticity.
of B. oleracea and its wild relatives, we hope to enable this
Postdomestication Cultivar Relationships model system for additional studies on evolutionary phenom-
Although our knowledge of the spread and diversification of ena such as parallel selection, polyploidy, and ferality.
B. oleracea crops after domestication is confounded by both Additionally, since many of these wild species are very narrow
the difficulties of identifying seeds of individual crop types endemics and are valuable for both crop improvement and
and frequent introgression between crop types, we can infer for nature conservation, their identification and preservation
some patterns using the species phylogeny. Like other studies are urgent. We hope this study can serve as a steppingstone,
(Cheng et al. 2016; Stansell et al. 2018), we find Chinese white as the work before us has, for those who, like Darwin was, are
kale sister to all other cultivars, representing the only Asian intrigued by this group of plants and wish to further its study.
clade of crop types (fig. 3A). Although the spread of B. oler-
acea to eastern Asia is still undocumented archaeologically,
Materials and Methods
recent pollen analysis has provided evidence for cultivation of Taxon Sampling
other Brassica species, including B. rapa, in the Yangtze valley Samples from cultivars accounted for 188 of the 224 total
3250–3350 BP, likely corresponding to movement across “Silk samples with the remaining 36 samples included being pre-
Road” trade routes (Zhang 2009). However, this only provides viously identified wild relatives (supplementary table S2,
identification criteria, not archaeological evidence (Yang et al. Supplementary Material online). These include accessions
11You can also read

















































