CONDUCTING PATHWAYS IN NORTH TEMPERATE DECIDUOUS BROADLEAVED TREES - Brill
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
IAWA Journal, Vol. 29 (3), 2008: 247–263
CONDUCTING PATHWAYS IN NORTH TEMPERATE DECIDUOUS
BROADLEAVED TREES
Toshihiro Umebayashi1, Yasuhiro Utsumi 2,*, Shinya Koga 2, Susumu Inoue 2,
Seizo Fujikawa 3, Keita Arakawa 3, Junji Matsumura 4 and Kazuyuki Oda 4
SUMMARY
The interspecific variation of dye ascent in the stems of 44 broadleaved
deciduous species growing in Japan was studied using freeze-dried
samples after dye injection. The dye ascending pattern differed both
within and between ring-porous and diffuse-porous species. In large
earlywood vessels of all ring-porous species, the dye ascended only in
the outermost annual ring, and the inner annual rings had lost their water
transport function. The dye ascending pattern within the inner annual
rings in the ring-porous species was categorized into three types: i) the
dye ascended both in the many latewood vessels throughout the latewood
and small earlywood vessels; ii) the dye ascended in the many vessels
throughout the latewood; and iii) the dye ascended mainly in the late
latewood vessels. In diffuse-porous species, the dye ascending pattern
within the annual rings also was categorized into three types: i) the dye
ascended throughout the annual rings; ii) the dye ascended mainly in
the earlywood vessels; and iii) the dye ascended mainly in the latewood
vessels. Xylem water distribution was also examined by cryo-SEM in
three ring-porous and three diffuse-porous species that had different dye
ascending patterns. The water distribution pattern within annual rings
was correlated with the dye ascending pattern except for one diffuse-
porous species (Salix gracilistyla). In this case, water was distributed in
the whole region of the annual rings although dye was mainly distributed
in the earlywood. These results showed that the functional area of water
transport within annual rings differed among ring-porous species and
diffuse-porous species.
Key words: Dye, cavitation, diffuse-porous, ring-porous, deciduous trees.
1) Graduate School of Bioresource and Bioenvironmental Sciences, Kyushu University, Hakozaki
812-8581, Japan.
2) Shiiba Research Forest, Graduate School of Agriculture, Kyushu University, Shiiba 883-0402,
Japan.
3) Laboratory of Wood Biology, Graduate School of Agriculture, Hokkaido University, Sapporo
060-8589, Japan.
4) Laboratory of Wood Science, Faculty of Agriculture, Kyushu University, Hakozaki 812-8581,
Japan.
*) Corresponding author [E-mail: utsumi@forest.kyushu-u.ac.jp].
Associate Editor: John Sperry
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access248 IAWA Journal, Vol. 29 (3), 2008
INTRODUCTION
Transpiring woody plants move the water from the soil to the leaves through the conduits
(vessels and tracheids) of their stems, and the anatomical characteristics of the stem
water-conducting pathway can have a profound impact on the efficiency of hydraulic
conductivity (Tyree & Zimmermann 2002). In stems of broadleaved trees growing in
northern temperate zones, ring-porous species have considerably larger diameter vessels
in the earlywood, and the earlywood vessels in the outermost annual ring have a major
role in water transport (Chaney & Kozlowski 1977; Ellmore & Ewers 1986; Granier
et al. 1994). On the other hand, diffuse-porous species have similar diameter vessels
scattered throughout the whole region of an annual ring, and the vessels in the outer
several annual rings have a role in water transport (Baker & James 1933; Kozlowski &
Winget 1963; Anfodillo et al. 1993). These results indicate that the functional region for
water transport in the stem xylem is obviously different between ring-porous trees and
diffuse-porous trees. However, in broadleaved trees, there are many ring-porous and
diffuse-porous species and their xylem structures vary widely (Wheeler et al. 1989; Carl-
quist 2001). The interspecific variation of the water transport pathways in ring-porous
and diffuse-porous species is not fully understood.
To evaluate the xylem sap flow of the stems of transpiring woody plants, various
methods have been used. Thermal methods such as the heat pulse method (Swanson
1994) and the thermal dissipation method (Granier 1987; Lu et al. 2004) have revealed
the radial profile of sap flow in the trunk (Čermák et al. 1992; James et al. 2002). Tracer
injection methods such as dye injection (Baker & James 1933; Koslowski et al. 1965;
Sakamoto & Sano 2000; Kuroda 2005), radioactive isotope injection (Postlethwait &
Rogers 1958; Thomas 1967), and deuterium injection (Meinzer et al. 2003, 2006) have
been used to describe the sap pathway. To visualize water distribution in the xylem,
nuclear magnetic resonance imaging (Holbrook et al. 2001; Clearwater & Clark 2003;
Utsuzawa et al. 2005) and cryo-scanning electron microscopy (cryo-SEM, Ohtani &
Fujikawa 1990; Utsumi et al. 1996, 2003) have also been used.
In the various methods mentioned above, the dye injection method is relatively easy to
conduct and has been widely used to visualize the water-conducting pathway at the tissue
level in trees (Rudinsky & Vité 1959; Harris 1961; Essiamah & Eschrich 1986; Barnett
et al. 1993; De Faÿ et al. 2000), woody shrubs (Hargrave et al. 1994) and herbaceous
plants (Tang & Boyer 2002). However, the difficulty in clarifying the functional region
of water transport at the cellular level has also been pointed out (Čermák et al. 2002).
To prevent the diffusion or flow of dye-carrying water after sampling of the stem, Sano
et al. (2005) proposed the dye injection method, which enabled the visualization of the
water-conducting xylem tissue of living trees at the cellular level using freeze drying
and dye insoluble solution when samples were embedded, and optimal conditions for
visualizing water-conducting pathways by this method was determined (Umebayashi
et al. 2007).
In this study, we examined the patterns of dye ascent of 44 deciduous broadleaved
species with ring-porous and diffuse-porous wood structure, using the improved dye
injection method. We especially focused on the interspecific variation of the water
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 249
transport region between ring-porous and diffuse-porous species and within ring-
porous and diffuse-porous species at the cellular level. In addition, we examined the
correspondence between tissue regions that remained sap-filled throughout the winter
and tissue regions taking up dye.
MATERIALS AND METHODS
Plant materials
Forty-four deciduous broadleaved species (eight ring-porous and 36 diffuse-porous
species) growing in the Shiiba Research Forest of Kyushu University (32° 22' N, 131°
09' E; elevation 880–1150 m) in Japan were used in this study (Table 1). According to
the temperature and precipitation data from 2003 to 2005 in the Shiiba Research Forest
(elevation 600 m), the yearly mean air temperature was 12.7 °C. The daily maximum
temperature for 2003, 2004, and 2005 was 33.0, 32.5, and 33.4 °C, respectively, and
the minimum temperature for the three years was -9.1, -8.5, and -8.3 °C, respectively.
The average yearly mean precipitation for the three years was 2918 mm.
Introduction of the dye solution and collection of samples
Dye injection experiments were performed from 11:00 to 14:00 on sunny days from
mid-July to August in 2003, 2004, and 2005. A watertight collar was set at a stem height
of 1.0 m and filled with distilled water. The southern half of the stem was cut with a
fine hand saw under water and the water was then replaced with an aqueous 0.2% acid
fuchsin solution that had been filtered through a 0.22 μm filter (GV, Millipore, Billerica,
USA) before the experiment. To keep water on the cut surface and avoid the adhesion
of debris, the stem was wrapped with a fine cotton fabric during the replacement. After
perfusion for 30 min, sample discs of 2 cm thickness were taken at 0, 10, 30, and 50 cm
above the dye injection height, and at 50 cm intervals up to 1.0 m to 10 cm below the
maximum height of the stained xylem. All discs were frozen by liquid nitrogen (LN2)
immediately, transferred to the laboratory, and freeze-dried at -50 °C (Umebayashi
et al. 2007). Two experiments were carried out for each species.
Observation of the dye distribution in the xylem
The surface of the sample discs was trimmed with a razor blade and the dye distri-
bution in the cross section of stem xylem was observed macroscopically. Following
this, the stained portion of the disc was split into small blocks, the surfaces of the
cross section were planed with a sliding microtome, and the dye distribution at the
tissue level was observed with a stereomicroscope (SMZ800, Nikon, Tokyo, Japan).
Subsequently, the small blocks containing at least one annual ring were trimmed and
embedded with an epoxy resin (Sano et al. 2005). Finally, thin sections of 4–7 μm were
cut from the embedded blocks with a semi-ultramicrotome (RM2045, Leica, Wetzlar,
Germany) and the dye distribution in each xylem cell was observed by light microscopy
(Optiphoto-2, Nikon, Tokyo, Japan). Images were obtained using a digital microscope
camera system (DP70, Olympus, Tokyo, Japan) attached to the stereomicroscope and
to the light microscope.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access250 IAWA Journal, Vol. 29 (3), 2008
Table 1. Tree height, diameter at breast height, the maximum dye ascent height and the stained
annual rings. Number of stained annual ring was counted from the outermost annual ring to the
pith. Asterisks show the species that have coloured heartwood.
Max. Stained annual rings
dye (total rings in sapwood)
Tree ascent ––––––––––––––––––––––––––––––––––
Species height DBH height 10 cm above Maximum
(m) (cm) (m) dye injection dye height
height
Ring-porous species
Acanthopanax sciadophylloides* 5.2 4.3 3.3 1–13 (17) 1–2 (4)
4.8 5.0 3.1 1–11 (15) 1–2 (6)
Aralia elata* 2.8 3.5 2.1 1–3 (4) 1 (1)
2.7 4.9 1.8 1– 4 (4) 1 (1)
Elaeagnus umbellata 5.0 3.0 1.9 1– 6 (7) 1–2 (4)
6.4 5.3 0.9 1–21 (26) 1 (25)
Kalopanax pictus* 5.8 5.8 4.1 1–15 (15) 1 (1)
4.9 4.7 3.6 1–24 (46) 1 (4)
Morus australis* 7.5 5.4 2.3 1–5 (12) 1–2 (11)
7.4 5.5 1.5 1– 4 (12) 1–2 (10)
Quercus crispula* 9.2 7.0 6.3 1–25 (26) 1 (6)
8.6 5.1 6.1 1–22 (22) 1–2 (6)
Rhus javanica var. roxburghii* 5.1 6.4 2.1 1– 4 (4) 1–2 (2)
7.8 6.4 6.2 1–3 (3) 1 (1)
Rhus trichocarpa* 5.5 4.2 2.8 1–11 (11) 1 (12)
3.9 4.2 1.9 1–18 (20) 1 (14)
Diffuse-porous species
Acer carpinifolium 5.5 6.0 0.9 1–25 (25) 1–14 (19)
6.1 5.6 1.6 1–15 (21) 1–5 (16)
Acer mono var. marmoratum 5.9 7.2 1.6 1–28 (29) 1–10 (27)
7.0 7.2 2.3 1–31 (31) 1– 4 (21)
Acer palmatum 5.3 4.5 1.1 1–29 (30) 1–3 (10)
3.5 3.3 1.8 1–22 (23) 1– 4 (21)
Acer sieboldianum 9.4 9.7 1.6 1–52 (54) 1–9 (43)
6.8 9.0 1.4 1– 49 (52) 1–5 (46)
Betula grossa 9.5 8.6 2.1 1–23 (23) 4–9 (19)
8.0 6.8 2.9 1–18 (18) 1–2, 6–7 (11)
Carpinus laxiflora 6.6 4.9 2.8 1–8 (10) 2–3 (4)
5.4 5.1 1.1 1–9 (10) 1– 4 (10)
Carpinus tschonoskii 6.9 5.0 1.6 1–10 (10) 1–3 (6)
4.6 5.0 1.6 1–10 (10) 1–2 (4)
Cercidiphyllum japonicum 6.2 3.5 1.1 1–6 (9) 1 (8)
5.6 6.3 0.8 1–19 (22) 3–5, 8–9, 14, 16 (21)
Clethra barbinervis 4.5 3.9 1.1 1–15 (17) 1–13 (15)
7.1 5.1 1.2 1–17 (19) 1–2, 9–10 (16)
Cornus macrophylla 4.4 3.7 2.7 1–8 (11) 3– 4 (4)
5.2 7.4 2.3 1–9 (11) 1 (5)
Deutzia crenata 4.2 2.5 1.4 1–7 (7) 1–2 (3)
4.3 2.9 1.6 1–6 (9) 1–3 (4)
Euonymus alatus* 5.1 6.4 1.8 1–19 (45) 1– 4 (38)
5.0 4.0 1.6 1–19 (44) 1–11 (39)
Euptelea polyandra 7.3 4.0 1.3 1–11 (11) 1–3 (9)
7.4 4.1 0.9 1–15 (15) 2– 4 (14)
––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––––
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 251
(Table 1 continued)
Max. Stained annual rings
dye (total rings in sapwood)
Tree ascent –––––––––––––––––––––––––––––––––
Species height DBH height 10 cm above Maximum
(m) (cm) (m) dye injection dye height
height
Fagus japonica 5.6 5.6 1.9 1–11 (16) 1 (11)
6.4 5.2 2.1 1–12 (17) 2–3, 5–6 (12)
Hydrangea paniculata* 4.6 7.0 0.5 1–8 (11) 1– 4 (11)
5.5 6.5 1.8 1–8 (12) 1–5 (12)
Hydrangea sikokiana 2.8 2.0 0.3 1–6 (9) 1–2 (7)
3.5 2.6 0.4 1–14 (16) 1–13 (16)
Ilex serrata 4.1 3.0 0.8 1–34 (40) 1–8 (35)
3.8 2.2 1.1 1–23 (28) 1– 4 (19)
Lindera erythrocarpa 7.0 5.6 1.8 1–17 (19) 1–5 (15)
6.3 9.7 1.9 1–20 (26) 1–14 (21)
Lindera praecox 5.5 3.8 1.1 1–14 (14) 1– 4 (12)
4.0 2.6 1.1 1–15 (18) 1 (15)
Lindera triloba 6.9 6.0 2.1 1–23 (27) 2–5 (20)
5.2 2.9 1.5 1–14 (15) 1 (12)
Lyonia ovalifolia var. elliptica 7.3 8.5 1.7 1–32 (33) 2 (27)
7.7 8.0 1.2 1–29 (31) 1– 4 (27)
Magnolia obovata* 5.4 5.0 1.4 1–13 (14) 1–2 (13)
5.8 4.8 1.2 1–12 (14) 1–3 (12)
Meliosma myriantha 6.5 6.9 0.9 1–14 (16) 1–3 (14)
6.0 7.5 1.1 1–22 (23) 1–5 (23)
Pourthiaea villosa var. laevis 7.0 7.2 1.6 1–18 (31) 1–12 (23)
5.2 4.5 1.6 1–12 (35) 1–2 (29)
Prunus jamasakura* 7.2 8.0 1.4 1–13 (26) 1 (21)
8.2 8.3 1.4 1–17 (25) 1–2, 5–6 (23)
Pterocarya rhoifolia 9.0 6.5 2.1 1–7 (8) 1–2 (7)
8.1 8.3 1.6 1–7 (7) 1–2, 4 (6)
Pterostyrax corymbosa 6.2 8.5 2.1 1–20 (21) 1–2 (17)
5.1 5.0 1.3 1–19 (19) 1–5 (15)
Rhododendron reticulatum 3.7 5.0 0.8 1–37 (37) 9–11 (28)
3.8 7.0 1.5 1–37 (37) 1–3 (33)
Salix gracilistyla* 5.2 6.2 2.4 1–13 (20) 1–2 (7)
4.0 3.3 1.8 1–10 (16) 1–2 (6)
Sapium japonicum 6.5 5.4 1.6 1–18 (23) 1–12 (16)
6.2 4.2 1.2 1–9 (16) 3–5 (12)
Sorbus gracilis 4.9 1.8 1.8 1–26 (29) 2–4 (19)
5.8 2.0 0.6 1–18 (23) 1–15 (23)
Stewartia monadelpha 6.1 6.4 2.8 1–17 (20) 1–3 (17)
5.8 5.8 3.6 1–20 (28) 1–2 (14)
Styrax japonica 5.5 5.5 2.7 1–22 (22) 1–3 (10)
5.8 6.2 1.4 1–18 (22) 1–3 (17)
Styrax shiraiana 3.8 2.9 1.6 1–7 (9) 1–2 (5)
5.8 3.7 1.5 1–8 (9) 1–2 (5)
Viburnum plicatum 5.3 4.0 1.8 1–23 (23) 1–12 (12)
5.3 3.9 0.8 1–27 (27) 1– 6 (19)
Weigela japonica* 6.5 6.5 3.9 1–15 (15) 1 (6)
7.1 7.1 3.8 1–11 (15) 1–2 (6)
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access252 IAWA Journal, Vol. 29 (3), 2008
Cryo-SEM
In three ring-porous species (Quercus crispula Blume, Rhus javanica var. roxburghii
(DC.) Rehd. & Wils., and Acanthopanax sciadophylloides Franch. & Sav.) and three
diffuse-porous species (Salix gracilistyla Miq., Fagus japonica Maxim., and Acer
palmatum Thunb.), two sample stems were collected in February 2006 for cryo-SEM
observation. A watertight collar with a plastic funnel was fitted to the stem at a height
of 100 cm and filled with LN2 to freeze the stem before sunrise (Utsumi et al. 2003).
Three-centimeter-thick discs were cut out with a hand saw from the frozen portion
of the stem and placed in LN2 (Utsumi & Sano 2007). The sample stem blocks were
trimmed and planed on a sliding microtome (Sano et al. 1995) in a cold room at -23 °C.
The frozen samples were slightly etched, coated with platinum and carbon in the cryo-
SEM system (JSM840-a equipped with CRU-7000, JEOL, Tokyo, Japan; Fujikawa
et al. 1988), and observed at 5 kV to assess the xylem water distribution of the outer
three or more annual rings (Utsumi et al. 1996; Utsumi & Sano 2007).
Definition of vessel arrangement in an annual ring
In ring-porous species, we described the earlywood as the initial zone of an annual
ring that contained tangentially one or several rows of larger diameter vessels in cross
section (Fig. 1a, E), and described the latewood as the other part of an annual ring. In
the earlywood of ring-porous species, two species (Acanthopanax sciadophylloides and
Kalopanax pictus (Thunb.) Nakai) have some clusters of small vessels (Itoh 1998).
In this case, we differentiated the small earlywood vessels (Fig. 1b, arrows) from the
large earlywood vessels (Fig. 1b, V). In the latewood of ring-porous species, we deter-
mined the early latewood to be the early half region of latewood (Fig. 1a, EL), and the
late latewood to be the late half region (Fig. 1a, LL). In diffuse-porous species, we de-
scribed the earlywood as the early half region of an annual ring (Fig. 1c, E) and the
latewood as the other zone (Fig. 1c, L).
RESULTS
Dye distribution in ring-porous species
At 10 cm above the dye injection height, the outer 3–25 annual rings from the cam-
bium in all ring-porous species were stained (Table 1). The samples of seven species
had colored heartwood (Table 1, asterisks), and the heartwood and innermost region
of the sapwood of these samples had no dye distribution (Fig. 2a). Quercus crispula
had the largest number of stained annual rings, and Rhus javanica var. roxburghii and
Aralia elata Miq. had the lowest number of stained rings. The stained annual ring
number in all ring-porous species decreased with an increase in stem height (Fig. 2a). At
the maximum dye ascent height, the outermost annual ring was stained in all species
(Fig. 2b), and some vessels in the late latewood (see Fig. 1) of the second annual ring
were also stained in five species (Fig. 2b, arrow; Table 1, Acanthopanax sciadophyl-
loides, Elaeagnus umbellata Thunb., Morus australis Poir, Q. crispula and R. javanica
var. roxburghii).
At 10 cm above the dye injection height, many large earlywood vessels in the outer-
most annual ring were stained, but the vessels in the inner annual rings were not stained
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 253
Figure 1. Definition of earlywood and latewood in ring-porous and diffuse-porous species in
this study. – a: Cross section of Kalopanax pictus. E, earlywood; EL, early latewood; LL, late
latewood. – b: Earlywood of K. pictus. V, large earlywood vessels; arrows, small earlywood ves-
sels. – c: Cross section of Lindera erythrocarpa. E, earlywood; L, latewood. — Scale bars: a, c =
200 μm; b = 100 μm.
in all species (Fig. 2c). In A. sciadophylloides and Kalopanax pictus, many small
earlywood vessels in all stained annual rings had dye in their lumina (Fig. 2d & e, ar-
rows). Many vessels throughout the entire latewood in the outermost annual ring were
stained in all species. In the inner rings, many or some vessels in the whole latewood
were stained in Q. crispula, E. umbellata, A. sciadophylloides, and K. pictus (Fig. 2d),
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access254 IAWA Journal, Vol. 29 (3), 2008
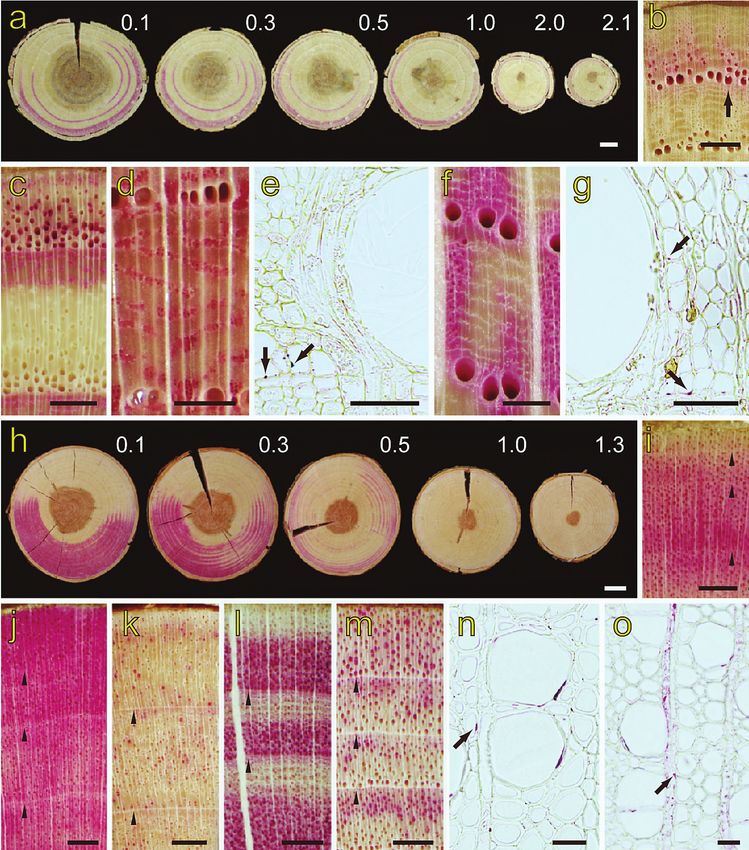
Figure 2. Micrographs of the stained xylem, cross section. – a: Discs from 10 cm above injection
to the maximum dye height in Rhus javanica var. roxburghii. The numbers give the distance (m)
from the dye injection height. – b: Outer two annual rings at the 150 cm height level in Quercus
crispula. Arrow, stained late latewood. – c: Outer two annual rings at the 10 cm height level in
R. javanica var. roxburghii. – d: Outer two annual rings at the 10 cm height level in Kalopanax
pictus. – e: Earlywood of the second annual ring at the 10 cm height level in K. pictus. Arrows,
dye in the small earlywood vessel lumina. – f: Outer two annual rings at the 10 cm height level
in Q. crispula. – g: Earlywood of the second annual ring at the 10 cm height level in Q. crispula.
Arrows, dye in the cells surrounding the vessel lumina. – h: Discs from 0.1 m to the maximum
dye height in Prunus jamasakura. The numbers give the distance (m) from the dye injection. –
i: Outer four annual rings at the 150 cm height level in Weigela japonica. Arrowheads, annual
ring boundary. – j–m: Outer three or four annual rings at the 10 cm height level in W. japonica
(j), Betula grossa (k), Fagus japonica (l), and Salix gracilistyla (m). Arrowheads, annual ring
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 255
and in the other species, only some late latewood vessels were stained (Fig. 2c). The
dye distribution pattern between each annual ring was similar in all species with the
exception of that for the outermost annual ring. Thus, we categorized the ring-porous
species into three types based on the dye distribution pattern within the annual ring
(Table 2). The first type is where small earlywood vessels and many latewood vessels of
the whole ring were stained (Fig. 2d & e; Table 2, r1). The second type is where many
vessels throughout the latewood were stained (Fig. 2f; Table 2, r2). The third type is
where mainly late latewood vessels were stained (Fig. 2c; Table 2, r3).
In all stained xylem at all heights, the tissues adjacent to stained vessels (e.g.,
vasicentric tracheids and wood fibers) contained dye in their lumina in four species
(Fig. 2g, arrows; Table 2, Q. crispula, Aralia elata, M. australis and R. javanica var.
roxburghii). Ray parenchyma cells were not stained in any species.
Dye distribution in diffuse-porous species
At 10 cm above the dye injection height, the outer 6–52 annual rings from the cam-
bium in each diffuse-porous species were stained (Table 1). Samples of six species
had colored heartwood (Table 1, asterisks), and in these samples, the heartwood and
innermost region of the sapwood had no dye (Fig. 2h). In this study, Acer sieboldianum
Miq. had the largest number of stained annual rings, and three species (Cercidiphyllum
japonicum Sieb. & Zucc., Hydrangea sikokiana Maxim., and Deutzia crenata Sieb. &
Zucc.) had the lowest number of stained annual rings. The stained annual ring number
in all diffuse-porous species decreased with an increase in stem height (Fig. 2h). At the
maximum dye ascent height, the outer several annual rings were stained uniformly in
most samples. Only one sample from six species was discontinuously stained across
annual rings (Table 1, Betula grossa Sieb. & Zucc., Cercidiphylllum japonicum, Clethra
barbinervis Sieb. & Zucc., Fagus japonica, Prunus jamasakura Sieb. ex Koidz. and
Pterocarya rhoifolia Sieb. & Zucc.).
The dye distribution pattern within the annual rings of inner rings was the same at
all heights for all species (Fig. 2i & j). In most species, a similar pattern of the stained
region was also observed between the outermost annual ring and inner annual rings
(Fig. 2j–l). In the outermost annual ring of two species (Salix gracilistyla and Clethra
barbinervis), vessels of the whole region were stained, and in the inner annual rings
the latewood was mainly stained (Fig. 2m). In the inner annual rings, the dye ascent
pattern was classified into three types. In species of the first pattern, most or some ves-
sels of the whole ring were stained (Fig. 2j & k; Table 2, d1). In species of the second
pattern, mainly earlywood vessels were stained (Fig. 2l; Table 2, d2). In species of the
third pattern, mainly latewood vessels were stained (Fig. 2m; Table 2, d3).
←
boundary. – n: Earlywood of the second annual ring at the 10 cm height level in Pterostyrax
corymbosa. Arrow, dye in a pit between vessel and surrounding tissue. – o: Earlywood of
the second annual ring at the 10 cm height level in Magnolia obovata. Arrow, the dye in a ray
parenchyma cell. — Scale bars: a, h = 1 cm; b, c, i–m = 1 mm; d, f = 500 μm; e, g = 50 μm;
n, o = 25 μm.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access256 IAWA Journal, Vol. 29 (3), 2008
Table 2. Staining pattern within annual rings. – r: ring-porous species; d: diffuse-porous species;
rl: whole latewood and small-earlywood vessels were stained; r2: whole latewood vessels were
stained; r3: mainly late latewood vessels were stained; d1: vessels of whole ring were stained;
d2: mainly earlywood vessels were stained; d3: mainly latewood vessels were stained; none:
the tissues adjacent to the vessels were not stained; portion: several tissues adjacent to vessels
were stained; whole: many tissues adjacent to vessels were stained.
Stained tissue adjacent
Stained tissue adjacent
maximum dye height
maximum dye height
Staining pattern of
Staining pattern of
to vessels
to vessels
Species Species
Ring-porous species Hydrangea paniculata – none
Hydrangea sikokiana – none
r1:
Ilex serrata – portion
Acanthopanax sciadophylloides r3 none
Lindera erythrocarpa – none
Kalopanax pictus r3 none
Lindera praecox – portion
r 2: Lindera triloba – portion
Elaeagnus umbellata r3 none Magnolia obovata – none
Quercus crispula r3 whole Meliosma myriantha – none
Pourthiaea villosa var. laevis – none
r 3:
Prunus jamasakura – portion
Aralia elata – portion
Pterocarya rhoifolia – portion
Morus australis – portion
Pterostyrax corymbosa – portion
Rhus javanica var. roxburghii – portion
Sapium japonicum – none
Rhus trichocarpa – none
Sorbus gracilis – none
Stewartia monadelpha – portion
Diffuse-porous species
Styrax japonica – none
d1: Styrax shiraiana – none
Acer carpinifolium – none Viburnum plicatum – none
Acer mono var. marmoratum – none Weigela japonica – portion
Acer palmatum – none
d2:
Acer sieboldianum – none
Euptelea polyandra – portion
Betula grossa – none
Fagus japonica – portion
Carpinus laxiflora – portion
Lyonia ovalifolia var. elliptica – none
Carpinus tschonoskii – portion
Rhododendron reticulatum – portion
Cercidiphyllum japonicum – none
Cornus macrophylla – whole d3:
Deutzia crenata – portion Clethra barbinervis – portion
Euonymus alatus – portion Salix gracilistyla – none
In 13 species of the d1 type, three species in d2 type and one species in d3 type,
dead xylem cells adjacent to stained vessels had dye in their lumina (Fig. 2n, arrow,
Table 2). Furthermore, in Lindera triloba Blume, Magnolia obovata Thunb., Sapium
japonicum Pax & K. Hoffm., Cercidiphyllum japonicum, Ilex serrata Thunb., and
Weigela japonica Thunb., the lumina of parenchyma cells adjacent to the vessels was
also stained (Fig. 2o, arrow).
Water distribution
Of the six species examined by cryo-SEM (Rhus javanica var. roxburghii, Quercus
crispula, Acanthopanax sciadophylloides, Acer palmatum, Fagus japonica, and Salix
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 257
Figure 3. Cryo-scanning electron micrographs of xylem cross section. – a: The second and third
annual rings of Acanthopanax sciadophylloides. Arrow, small earlywood vessel. – b: The second
annual ring of Quercus crispula. Arrow, water-filled latewood vessel. – c & d: The second and
third annual rings of Rhus javanica var. roxburghii. – e: The second annual ring of Acer pal-
matum. – f: The fourth and fifth annual rings of Fagus japonica. – g: The fourth annual ring of
Salix gracilistyla. – e–g: Arrowhead, annual ring boundary. — Scale bars: a–c, e–g = 100 μm;
d = 50 μm.
gracilistyla), all except for S. gracilistyla showed similar patterns of water distribution
in the outer three annual rings. In all three ring-porous species, most large earlywood
vessels did not have water in their lumina (Fig. 3a–c). In A. sciadophylloides, most
small earlywood vessels (Fig. 3a, arrow) and all latewood vessels had water in their
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access258 IAWA Journal, Vol. 29 (3), 2008
lumina. The wood fibers were rarely water-filled. In Q. crispula, some latewood vessels
had water in their lumina (Fig. 3b, arrow), and many cells adjacent to the vessels also
had water in their lumina. In R. javanica var. roxburghii, no early latewood vessels had
water (Fig. 3c), and some late latewood vessel clusters had water in their lumina (Fig.
3d). The wood fibers were rarely water-filled, except for the tissues adjacent to late
latewood vessels. In A. palmatum and F. japonica, most vessels throughout an annual
ring had water in their lumina (Fig. 3e & f). In S. gracilistyla, most outermost annual
ring vessels were filled with water. In the second and inner annual ring, only latewood
vessels had water in their lumina (Fig. 3g). In the three diffuse-porous species, many
wood fibers had water in their lumina (Fig. 3e–g).
DISCUSSION
The stem water-conducting pathway in 44 deciduous broadleaved species varied among
ring-porous and diffuse-porous species as well as between these two groups (Table 2).
In ring-porous species, some reports indicate that the water in tree stems is transported
mainly through the outermost annual ring (Greenidge 1955; Kramer & Kozlowski
1979; Kubler 1991; Kozlowski & Pallardy 1997). However, other reports indicate that
inner annual rings as well as the outermost annual ring have water transport functions
(Kozlowski & Winget 1963; Granier et al. 1994). In all ring-porous species we stud-
ied, we found that large earlywood vessels of the outermost annual ring functioned
in water transport, but there was no water transport function in the second and inner
annual rings (Fig. 2b–g & 3a–c). In general, cavitation, the rupture of water continuity
in the water transport system, is induced by water stress and freezing stress (Tyree &
Sperry 1989). It is unlikely that water stress-induced cavitation occurred in most large
earlywood vessels, because many large earlywood vessels in the outermost annual ring
of all ring-porous species in this study maintained their water transport function during
the growing season when most leaves transpired and the water potential decreased. On
the other hand, some reports indicate that the hydraulic conductivity of ring-porous
species declines rapidly after the first frost (Cochard & Tyree 1990; Sperry et al. 1994)
and that earlywood vessels in the outermost annual ring are cavitated (Utsumi et al.
1999). It is suggested that the freeze-thaw cycle causes embolism in conduits when the
diameter is greater than 43 μm at the -0.5 MPa condition (Davis et al. 1999; Pittermann
& Sperry 2003). The water in dead xylem cells freezes at several degrees below zero
(Zimmermann 1964), although cold acclimated living tissue freezes commonly between
-8 and -10 °C (Sakai & Larcher 1987). The large earlywood vessels of the examined
species were over 43 μm in diameter (Fig. 3a–c). And the minimum yearly temperature
at our study site was less than -8 °C during the experiment. Therefore, freeze-induced
cavitation would occur in the large earlywood vessels of the outermost annual ring,
which would lose their water transport function during winter.
We showed that small earlywood and latewood vessels in some ring-porous spe-
cies maintained their water transport system over several years (Fig. 2c–e). There is a
positive correlation between vulnerability to cavitation caused by freezing and conduit
diameter (Ewers 1985; Cavender-Bares 2005), and smaller conduits tend to experience
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 259
less freeze-thaw embolism than larger ones (Cochard & Tyree 1990; Feild & Brodribb
2001). Functioning periods for water transport will differ between large earlywood
vessels and small earlywood and latewood vessels. These small vessels would contrib-
ute less to water transport under normal conditions. However, ring-porous deciduous
species leaf out later than many diffuse-porous species (Wang et al. 1992) and expand
their leaves before the maturation of earlywood vessels in the outermost annual ring
(Suzuki et al. 1996). In the earliest stage of leaf expansion, small vessels of the second
and inner annual ring would have a major role in water transport.
In the diffuse-porous species studied, the water-conducting pattern within an an-
nual ring was categorized into three types. Many previous reports have only shown
the conducting annual ring number macroscopically (e.g., Kozlowski & Winget 1963;
Čermák et al. 2002), and only a few studies analyzed variation of the water-conducting
pathway within individual annual rings (Baker & James 1933; Chaney & Kozlowski
1977). Our study is the first report showing the variation of the water transport path-
way of annual rings for various diffuse-porous species. Species in which all vessels
in each annual ring conduct water (d1 type) probably avoid embolism throughout the
growing season or reverse it by a mechanism such as root pressure. It is known that the
water transport system in Betula spp. and Acer spp. generates high positive pressures
in the stems and roots (Merwin & Lyon 1909; Kramer & Kozlowski 1960; Milburn &
OʼMalley 1984). All species of Betula and Acer studied showed the d1 type. Positive
high pressure may enable the vessels in the d1 type to be functional in the whole ves-
sel region. In species that only transport water in the earlywood (d2 type), the whole
vessel region was filled with water (Fig. 3f). In this case, the water ascending region
and water distribution in xylem did not correspond. Relatively small latewood vessels
would have a water transport function, but the amount of moving water would be small
when large vessels are functioning because the vessel diameter strongly affects water
transport efficiency according to the Hagen-Poiseuille law (Tyree & Zimmermann
2002). In the d3 type, all vessels in the outermost annual ring and the latewood vessels
in the inner annual rings had water in their lumina and a transport function (Fig. 2m
& 3g). These results indicate that the earlywood vessels of the outermost annual ring
maintained the water transport function during the first growing season, but lost water
during the first dormant season. Freeze-induced cavitation would occur in the larger
earlywood vessels of the outermost annual ring in winter.
A clear relationship between the dye distribution pattern of the tissue surround-
ing the vessel lumen and the tissue cell type was not observed. In ring-porous trees,
Quercus species have vasicentric tracheids (Wheeler et al. 1989) and the surrounding
tissue of Q. crispula was stained with dye in this study. On the other hand, the species
that have no vasicentric tracheids and where the vessels are surrounded by axial paren-
chyma such as Morus australis (Itoh 1995), were also stained. Even in the same ge-
nus, the surrounding tissue of Rhus javanica var. roxburghii was stained, but that of
R. trichocarpa Miq. was not stained. In diffuse-porous species, the surrounding tis-
sue of Lindera praecox (Sieb. & Zucc.) Blume and Lindera triloba was stained but
that of Lindera erythrocarpa Makino was not stained. These congeneric species had
vasicentric parenchyma and showed no apparent structural difference under the light
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access260 IAWA Journal, Vol. 29 (3), 2008
microscope. The staining property of the cells surrounding the vessel lumina may not
be determined by the cell type but regulated by the porosity and chemical property of
the pit membranes between the vessel and the adjacent cells.
In ring-porous species, most of the water was transported in the outermost annual
ring, although many annual rings of sapwood had a water transport function. This is
because the large earlywood vessels can maintain their water transport function only
during the first growth season. On the other hand, the radial sap flow pattern in diffuse-
porous species was unclear in this study. In a sample of Cercidiphyllum japonicum,
Fagus japonica, Cornus macrophylla Wall., Lindera praecox, L. triloba and Weigela
japonica Thunb., the fastest water ascent region was in the outermost annual ring, while
in a sample of Rhododendron reticulatum D. Don ex G. Don, water ascended mainly in
the inner annual ring (Table 1). In diffuse-porous and coniferous species, the sap flow
varies radially (Čermák & Nadezhdina 1998; Jiménez et al. 2000). Some reports indicate
that the middle portion of the sapwood transports water rapidly (Čermák et al. 1992),
and in other studies, it was suggested that the outer part of the sapwood was the fastest
region (Čermák et al. 2002; Nadezhdina et al. 2002). In diffuse-porous trees, the radial
sap flow pattern might differ between species and even within a species. However, our
results only show the water conducting pathways because dye was introduced from a
notch in the stem. To clarify the actual radial distribution pattern of sap flow among
ring-porous and diffuse-porous species, further qualitative and quantitative studies such
as dye injection from the root are needed.
ACKNOWLEDGEMENTS
The authors thank Hisami Nagasawa, Katsuyoshi Kubota, Sachiko Inoue, Shigeru Osaki, Yasuki Shiiba
and Toshiaki Itoh for their technical support. This work was supported in part by a Grant-in-Aid for
Scientific Research (No. 19780121) from the Ministry of Education, Science and Culture, Japan.
REFERENCES
Anfodillo, T., G.B. Sigalotti, M. Tomasi, P. Semenzato & R. Valentini. 1993. Applications of
a thermal imaging technique in the study of the ascent of sap in woody species. Plant Cell
Environ. 16: 997–1001.
Baker, H. & W.O. James. 1933. The behavior of dyes in the transpiration stream of sycamores
(Acer pseudoplatanus L.). New Phytol. 32: 245–260.
Barnett, J.R., P. Cooper & L.J. Bonner. 1993. The protective layer as an extension of the apoplast.
IAWA Bull. n.s. 14: 163–171.
Carlquist, S. 2001. Comparative wood anatomy. Ed. 2. Springer Verlag, Berlin.
Cavender-Bares, J. 2005. Impacts of freezing on long distance transport in woody plants. In:
N.M. Holbrook & M.A. Zwienieck (eds.), Vascular transport in plants: 401–424. Elsevier
Academic Press, Burlington.
Čermák, J., E. Cienciala, J. Kučera & J. Hällgren. 1992. Radial velocity profiles of water flow
in trunks of Norway spruce and oak and the response of spruce to severing. Tree Physiol.
10: 367–380.
Čermák, J., M.S. Jimenéz, A.M. González-Rodríguez & D. Morales. 2002. Laurel forests in
Tenerife, Canary Islands. II. Efficiency of the water conducting system in Laurus azorica
trees. Trees 16: 538–546.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 261
Čermák, J. & N. Nadezhdina. 1998. Sapwood as the scaling parameter: Defining according to
xylem water content or radial pattern of sap flow? Ann. Sci. For. 55: 509–521.
Chaney, W.R. & T.T. Kozlowski. 1977. Patterns of water movement in intact and excised stems
of Fraxinus americana and Acer saccharum seedlings. Ann. Bot. 41: 1093–1100.
Clearwater, M. J. & C. J. Clark. 2003. In vivo magnetic resonance imaging of xylem vessel con-
tents in woody lianas. Plant Cell Environ. 26: 1205–1214.
Cochard, H. & M.T. Tyree. 1990. Xylem dysfunction in Quercus: vessel sizes, tyloses, cavitation
and seasonal changes in embolism. Tree Physiol. 6: 393–407.
Davis, S.D., J.S. Sperry & U.G. Hacke. 1999. The relationship between xylem conduit diameter
and cavitation caused by freezing. Amer. J. Bot. 86: 1367–1372.
De Faÿ, E., V. Vacher & F. Humbert. 2000. Water-related phenomena in winter buds and twigs
of Picea abies (L.) Karst. until bud-burst: A biological, histological and NMR study. Ann.
Bot. 86: 1097–1107.
Ellmore, G.S. & F.W. Ewers. 1986. Fluid flow in the outermost xylem increment of a ring porous
tree. Ulmus americana. Amer. J. Bot. 73: 1771–1774.
Essiamah, S. & W. Eschrich. 1986. Water uptake in deciduous trees during winter and the role
of conducting tissues in spring reactivation. IAWA Bull. n.s. 7: 31–38.
Ewers, F.W. 1985. Xylem structure and water conduction in conifer trees, dicot trees, and lianas.
IAWA Bull. n.s. 6: 309–317.
Feild, T.S. & T. Brodribb. 2001. Stem water transport and freeze-thaw xylem embolism in conifers
and angiosperms in a Tasmanian treeline heath. Oecologia 27: 314–320.
Fujikawa, S., T. Suzuki, T. Ishikawa, S. Sakurai & Y. Hasegawa. 1988. Continuous observation
of frozen biological materials with cryo-scanning electron microscope and freeze-replica by
a new cryo-system. J. Electron Microsc. 37: 315–322.
Granier, A. 1987. Evaluation of transpiration in a Douglas-fir stand by sap flow measurements.
Tree Physiol. 3: 309–320.
Granier, A., T. Anfodillo, M. Sabatti, H. Cochard, E. Dreyer, M. Tomasi, R. Valentini & N. Bréda.
1994. Axial and radial water flow in the trunks of oak trees: a quantitative and qualitative
analysis. Tree Physiol. 14: 1383–1396.
Greenidge, K.N.H. 1955. Studies in the physiology of forest trees. III. The effect of drastic inter-
ruption of conducting tissues on moisture movement. Amer. J. Bot. 42: 582–587.
Hargrave, K.R., K. J. Kolb, F.W. Ewers & S.D. Davis. 1994. Conduit diameter and drought-in-
duced embolism in Salvia mellifera Greene (Labiatae). New Phytol. 126: 695–705.
Harris, J.M. 1961. Water-conduction in the stems of certain conifers. Nature 4765: 678–679.
Holbrook, N., E.T. Ahrens, M.J. Burns & M.A. Zwienieck. 2001. In vivo observation of cavitation
and embolism repair using magnetic resonance imaging. Plant Physiol. 126: 27–31.
Itoh, T. 1995. Anatomical description of Japanese hardwoods. I. Wood Res. Technical Notes
31: 81–181.
Itoh, T. 1998. Anatomical description of Japanese hardwoods. IV. Wood Res. Technical Notes
34: 30–166.
James, S.A., M. J. Clearwater, F.C. Meinzer & G. Goldstein 2002. Heat dissipation sensors of
variable length for the measurement of sap flow in trees with deep sapwood. Tree Physiol.
22: 277–283.
Jimenéz, M.S., N. Nadezhidina, J. Čermák & D. Morales. 2000. Radial variation in sap flow in
five laurel forest tree species in Tenerife, Canary Islands. Tree Physiol. 20: 1149–1156.
Kozlowski, T.T., L. Leyton & J.F. Hughes. 1965. Pathways of water movement in young coni-
fers. Nature 205: 830.
Kozlowski, T.T. & S.G. Pallardy. 1997. Physiology of woody plants. Ed. 2. Academic Press, Cali-
fornia.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free access262 IAWA Journal, Vol. 29 (3), 2008
Kozlowski, T.T. & C.H. Winget. 1963. Patterns of water movement in forest trees. Bot. Gaz.
124: 301–311.
Kramer, P. J. & T.T. Kozlowski. 1960. Physiology of trees. McGraw-Hill, New York.
Kramer, P. J. & T.T. Kozlowski. 1979. Physiology of woody plants. Academic Press, New
York.
Kubler, H. 1991. Function of spiral grain in trees. Trees 5: 125–135.
Kuroda, K. 2005. Xylem dysfunction in Yezo spruce (Picea jezoensis) after inoculation with the
blue-stain fungus Ceratocystis polonica. Forest Pathol. 35: 346–358.
Lu, P., L. Urban & Z. Ping. 2004. Granierʼs thermal dissipation probe (TDP) method for measur-
ing sap flow in trees: Theory and practice. Acta Bot. Sin. 46: 631–646.
Meinzer, F.C., J.R. Brooks, J.C. Domeck, B.L. Gartner, J.M. Warren, D.R. Woodruff, K. Bible &
D.C. Shaw. 2006. Dynamics of water transport and storage in conifers studied with deuterium
and heat tracing techniques. Plant Cell Environ. 29: 105–114.
Meinzer, F.C., S.A. James, G. Goldstein & D. Woodruff. 2003. Whole-tree water transport
scales with sapwood capacitance in tropical forest canopy trees. Plant Cell Environ. 26:
1147–155.
Merwin, H.E. & H. Lyon. 1909. Sap pressure in the birch stem. Bot. Gaz. 48: 442–458.
Milburn, J.A. & P.E. OʼMalley. 1984. Freeze-induced sap absorption in Acer pseudoplatanus:
a possible mechanism. Can. J. Bot. 62: 2101–2106.
Nadezhdina, N., J. Čermák & R. Ceulemans. 2002. Radial patterns of sap flow in woody stems
of dominant and understory species: scaling errors associated with positioning of sensors.
Tree Physiol. 22: 907–918.
Ohtani, J. & S. Fujikawa. 1990. Cryo-SEM observation on vessel lumina of a living tree: Ulmus
davidiana var. japonica. IAWA Bull. n.s. 11: 183–194.
Pittermann, J. & J.S. Sperry. 2003. Tracheid diameter is the key trait determining the extent of
freezing-induced embolism in conifers. Tree Physiol. 23: 907–914.
Postlethwait, S.N. & B. Rogers. 1958. Tracing the path of the transpiration stream in trees by
the use of radioactive isotopes. Amer. J. Bot. 45: 753--757.
Rudinsky, J.A. & J.P. Vité. 1959. Certain ecological and phylogenetic aspects of the pattern of
water conduction in conifers. Forest Sci. 5: 259–266.
Sakai, A. & W. Larcher. 1987. Frost survival of plants : responses and adaptation to freezing stress.
Springer Verlag, Berlin.
Sakamoto, Y. & Y. Sano. 2000. Inhibition of water conductivity caused by watermark disease in
Salix sachalinensis. IAWA J. 21: 49–60.
Sano, Y., S. Fujikawa & F. Fukazawa. 1995. Detection and features of wetwood in Quercus
mongolica var. grosseserrata. ??? 9: 261–268.
Sano, Y., Y. Okamura & Y. Utsumi. 2005. Visualizing water-conduction pathways of living trees:
selection of dyes and tissue preparation methods. Tree Physiol. 25: 269–275.
Sperry, J.S., K.L. Nichols, J.E.M. Sullivan & S.E. Eastlack. 1994. Xylem embolism in ring-
porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska. Ecology
75: 1736–1752.
Suzuki, M., K. Yoda & H. Suzuki. 1996. Phenological comparison of the onset of vessel forma-
tion between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest.
IAWA J. 17: 431– 444.
Swanson, R.H. 1994. Significant historical developments in thermal methods for measuring sap
flow in trees. Agr. For. Met. 72: 113–132.
Tang, A.C. & J.S. Boyer. 2002. Growth-induced water potentials and the growth of maize leaves.
J. Exp. Bot. 53: 489–503.
Thomas, W.A. 1967. Day and calcium ascent in dogwood tree. Plant Physiol. 42: 1800–1802.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessUmebayashi et al. — Conducting pathways in temperate trees 263
Tyree, M.T. & J.S. Sperry. 1989. Vulnerability of xylem to cavitation and embolism. Annu. Rev.
Plant Physiol. Plant Mol. Biol. 40: 19–38.
Tyree, M.T. & M.H. Zimmermann. 2002. Xylem structure and the ascent of sap. Ed. 2. Springer
Verlag, Berlin.
Umebayashi, T., Y. Utsumi, S. Koga, S. Inoue, Y. Shiiba, K. Arakawa, J. Matsumura & K. Oda.
2007. Optimal conditions for visualizing water-conducting pathways in a living tree by the
dye injection method. Tree Physiol. 27: 993–999.
Utsumi, Y. & Y. Sano. 2007. Cryoplaning technique for visualizing the distribution of water in
woody tissues by cryoscanning electron microscopy. In: J. Kuo. (ed.), Electron microscopy:
Methods and protocols. Ed. 2: 497–506. Humana Press, Totawa.
Utsumi, Y., Y. Sano, R. Funada, S. Fujikawa & J. Ohtani. 1999. The progression of cavitation
in earlywood vessels of Fraxinus mandshurica var. japonica during freezing and thawing.
Plant Physiol. 121: 897–904.
Utsumi, Y., Y. Sano, R. Funada, J. Ohtani & S. Fujikawa. 2003. Seasonal and perennial changes
in the distribution of water in the sapwood of conifers in a sub-frigid zone. Plant Physiol.
131: 1826–1833.
Utsumi, Y., Y. Sano, J. Ohtani & S. Fujikawa. 1996. Seasonal changes in the distribution of water
in the outer growth rings of Fraxinus mandshurica var. japonica: A study by cryo-scanning
electron microscopy. IAWA J. 17: 113–124.
Utsuzawa, S., K. Fukuda & D. Sakaue. 2005. Use of magnetic resonance microscopy for the
nondestructive observation of xylem cavitation caused by pine wilt disease. Phytopathol.
95: 737–743.
Wang, J., N.E. Ives & M.J. Lechowicz. 1992. The relation of foliar phenology to xylem embolism
in trees. Funct. Ecol. 6: 469– 475.
Wheeler, E.A., P. Baas & P.E. Gasson. 1989. IAWA list of microscopic features for hardwood
identification. IAWA Bull. n.s. 10: 219–332.
Zimmermann, M.H. 1964. Effect of low temperature on ascent of sap in trees. Plant Physiol.
39: 568–572.
Downloaded from Brill.com09/16/2020 07:30:29PM
via free accessYou can also read

















































