Discovery of a dune-building hybrid beachgrass Ammophila
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Discovery of a dune-building hybrid beachgrass (Ammophila
arenaria 9 A. breviligulata) in the U.S. Pacific Northwest
REBECCA S. MOSTOW , FELIPE BARRETO , REUBEN BIEL , ELI MEYER, AND SALLY D. HACKER
Department of Integrative Biology, Oregon State University, Corvallis, Oregon 97331 USA
Citation: Mostow, R. S., F. Barreto, R. Biel, E. Meyer, and S. D. Hacker. 2021. Discovery of a dune-building hybrid
beachgrass (Ammophila arenaria 9 A. breviligulata) in the U.S. Pacific Northwest. Ecosphere 12(4):e03501. 10.1002/ecs2.
3501
Abstract. The production of novel hybrid zones is an ecologically important consequence of globally
increasing rates of species introductions and invasions. Interspecific hybridization can facilitate gene flow
between parent species or produce novel taxa that may alter invasion dynamics or ecosystem services. The
coastal sand dunes of the U.S. Pacific Northwest coast are densely populated by two non-native, con-
generic, dune-building beachgrasses (Ammophila arenaria and A. breviligulata). Here, we present morpho-
logical, cytological, and genetic evidence that the two beachgrass species have hybridized in this globally
unique range overlap. The A. arenaria 9 A. breviligulata hybrid has been found at 12 coastal sites in Wash-
ington and Oregon. It is a first-generation hybrid between the beachgrass species as evidenced by genome
size comparisons and single nucleotide polymorphism genotyping. It is intermediate between the parent
grasses in many morphological characters but exceeds both parents in shoot height, a trait associated with
dune-building potential. Understanding the ecological and population genetic consequences of this novel
hybridization event is of the utmost importance in a system where any change in dominant beachgrass
species can have large effects on both biodiversity management and coastal protection.
Key words: Ammophila; beachgrass; ecosystem engineer; hybrid zone; hybridization; invasive grass; U.S. Pacific
Northwest.
Received 16 December 2020; accepted 6 January 2021; final version received 24 February 2021. Corresponding Editor:
Debra P. C. Peters.
Copyright: © 2021 The Authors. This is an open access article under the terms of the Creative Commons Attribution
License, which permits use, distribution and reproduction in any medium, provided the original work is properly cited.
E-mail: mostowr@oregonstate.edu
INTRODUCTION speciation and adaptation (Anderson and Steb-
bins 1954, Wissemann 2007). Hybridization may
The rate of human-mediated species introduc- increase the invasiveness of plants by producing
tions is increasing (Seebens et al. 2017), posing evolutionarily novel phenotypes or phenotype
threats to biodiversity, human health, and global combinations, increasing genetic variation (in the
economies (Simberloff et al. 2013) and affecting parent species through backcrossing, or in the
biogeography at a global scale (Capinha et al. hybrid itself), facilitating genetic load dumping
2015). Introductions can produce unique species (i.e., masking deleterious alleles accumulated in
co-occurrence patterns, and if they generate small or isolated populations), and fixing hetero-
range overlaps between related species, novel sis (i.e., mechanisms that stabilize hybriditidy,
interspecific hybridization events are possible for example, allopolyploidy or clonal spread; Ell-
(Chunco 2014). Hybridization is common in both strand and Schierenbeck 2000). Transgressive
the evolutionary history and modern ecology of segregation, or the tendency to express extreme
plants (Mallet 2005) and can serve as a driver of phenotypes compared with the parent species, is
v www.esajournals.org 1 April 2021 v Volume 12(4) v Article e03501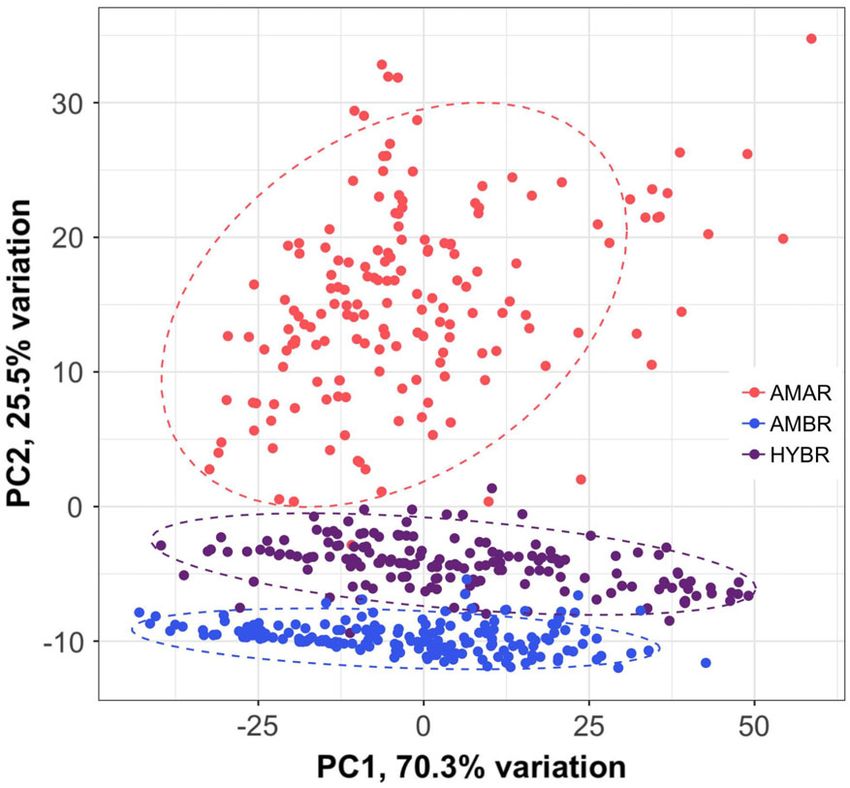
MOSTOW ET AL.
common in plant hybrids and may contribute to California coastline; Cooper 1958, 1967). In the
rapid adaptation and range expansion of hybrid early 1900s, U.S. Pacific coast dunes were sys-
lineages (Rieseberg et al. 2001). For example, tematically planted with the non-native Euro-
hybridization within the genus Spartina (cord- pean beachgrass Ammophila arenaria to stabilize a
grass) has produced a variety of highly invasive shifting sand environment (Seabloom and
taxa including hybrid polyploids (e.g., S. anglica Wiedemann 1994). By the 1950s, A. arenaria had
in France, Salmon et al. 2005), sterile first-genera- spread from Mexico to Canada building tall, con-
tion (F1) hybrids (e.g., S. densiflora 9 maritima on tinuous coastal foredunes (Wiedemann and Pick-
the Iberian Peninsula in Spain, Castillo et al. art 1996). Meanwhile, in the 1930s, another non-
2010), and multiple backcrossed hybrids (e.g., native beachgrass, American beachgrass Ammo-
S. alterniflora 9 foliosa in San Francisco, Califor- phila breviligulata was planted in dunes near the
nia, Ayres et al. 2008b). These hybrids have stabi- Columbia River and, by the 1980s, had spread
lized sediment and expanded salt marsh and dominated northward along the Washington
habitats, to the detriment of native mudflat com- coast (Seabloom and Wiedemann 1994). The
munities (Hacker and Dethier 2006, Strong and spread of these beachgrasses has had a positive
Ayres 2009). Thus, when hybridization occurs in impact on coastal development; they stabilize the
plants that are highly invasive and have strong naturally shifting sand environment, build tall
ecosystem engineering properties, the genetic stable dunes, and increase coastal protection
and ecological consequences can be significant (Hacker et al. 2012, Seabloom et al. 2013, Rug-
and unpredictable. giero et al. 2018). However, they also negatively
In this paper, we provide the first documented impact biodiversity management by outcompet-
evidence of a novel hybrid formed between two ing native plants and reducing habitat value for
non-native, dune-building Ammophila beachgrass native shorebirds, particularly the threatened
species, both of which dominate the U.S. Pacific Western snowy plover (Zarnetske et al. 2010, Biel
Northwest coast. Coastal dunes are dynamic and et al. 2017).
harsh ecosystems at the land-sea interface char- The two congeneric beachgrasses differ in their
acterized by strong winds and waves, salt spray, dune building and competitive abilities. A. bre-
and sand burial. These conditions reduce the viligulata is a better competitor (Zarnetske et al.
likelihood of plant invasions (Maun 2009) but 2013) but A. arenaria creates taller dunes, even
regular disturbance events (e.g., overwash and after accounting for differences in sand supply
blowouts) can produce openings and opportuni- (Hacker et al. 2012, Zarnetske et al. 2012, Biel
ties for appropriately adapted plants to establish et al. 2019). The variation in competitive ability
(Hesp 2002, David et al. 2015, Charbonneau et al. and dune shape is ultimately the result of differ-
2020). Coastal ecosystems are especially vulnera- ences in grass density, morphology, growth form,
ble to the geomorphic changes caused by ecosys- and their effects on sand capture (Hacker et al.
tem engineering invaders (Fei et al. 2014). In 2012, Zarnetske et al. 2012, 2013). A. arenaria
addition, given the shifting and unstable nature exhibits a vertical growth form with high stem
of coastal dunes, non-native dune plants have density that results in deeper sand deposition for
been used over the last century for stabilization a given area, and thus taller and narrower dunes
purposes. One of the most often used is Ammo- (Hacker et al. 2012, Biel et al. 2019). In compar-
phila spp., a grass that has been intentionally ison, A. breviligulata exhibits more lateral growth
introduced to sandy coastlines all over the world with a lower stem density, resulting in better
where it specializes in sand capture and stabi- space occupation, but less sand accretion over a
lization, resulting in tall and wide foredunes and larger area, creating shorter and wider dunes.
increased coastal protection (Wiedemann and The intentional introduction and subsequent
Pickart 1996, Hertling and Lubke 1999, Hilton spread of the two beachgrass species over the last
et al. 2006, Hacker et al. 2012). century has resulted in overlapping distributions
One such intentional introduction of Ammo- along the U.S. Pacific Northwest coast, a globally
phila occurred on the U.S. Pacific coast, where unique occurrence pattern.
dune habitat is widespread (i.e., 45% of the Ore- In 2012, a putative hybrid was discovered in
gon and Washington coastline and 25% of the the geographic overlap between the two
v www.esajournals.org 2 April 2021 v Volume 12(4) v Article e03501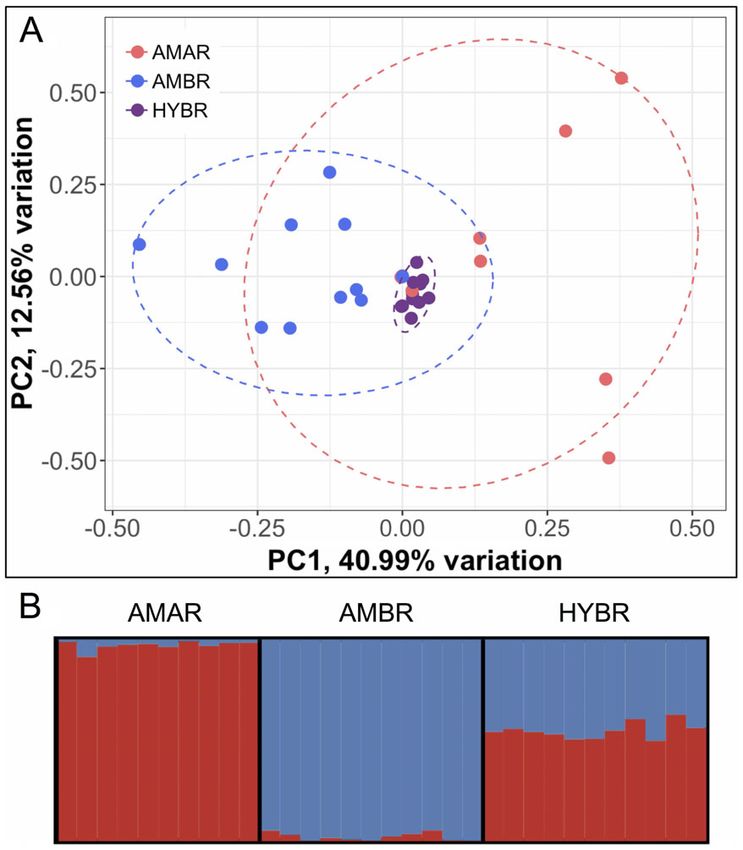
MOSTOW ET AL.
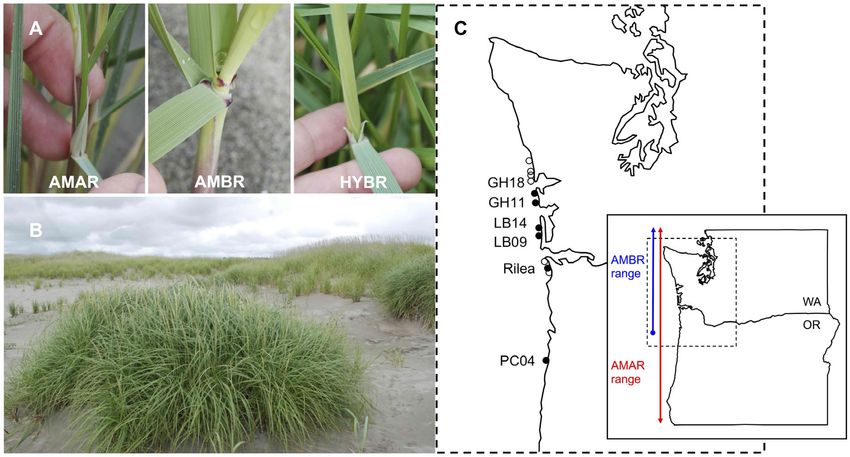
Fig. 1. (A) Photographs of the distinctive ligules of the Ammophila taxa. Ammophila arenaria (AMAR) has a long,
pointed ligule, Ammophila breviligulata (AMBR) has a short, flat-topped ligule, and the Ammophila hybrid (HYBR) has
a ligule of intermediate length with a flat top. (B) A patch of Ammophila hybrid (foreground) at the toe of an A. bre-
viligulata-dominated dune. (C) Map of all known Ammophila hybrid stands. Labeled, closed circles are sites discovered
between 2013 and 2018, from which hybrid samples were collected. Open circles are sites discovered between 2018
and 2019, after data analysis for this study had begun. Inset shows the range overlap between AMAR and AMBR in
Washington and Oregon. See Appendix S1: Table S1 for site and transect abbreviations and locations.
Ammophila species (Hacker et al. 2012, Zarnetske parents. We aim to (1) verify the hybrid origin
et al. 2013; Fig. 1). Individual stands of this mor- using cytology (genome size estimation with
phologically unusual grass have since been flow cytometry) and single nucleotide polymor-
found at 12 sites between Pacific City, Oregon, phism (SNP) genotyping and (2) compare mor-
and Ocean Grove, Washington (hybrids from six phological characters which may aid in hybrid
sites were sampled for this study, Fig. 1C). The identification and affect sand capture and dune
putative hybrid individuals were initially discov- building. If the putative hybrid exceeds its par-
ered because they were morphologically similar ents in traits associated with dune-building
to A. breviligulata but had ligules (the thin tissue potential (i.e., stem density, plant height, or bio-
at the base of the leaf) intermediate in length mass), which is possible via heterosis, then its
between the two Ammophila species (Fig. 1A). spread could affect dune morphology and have
Ligule length is a distinguishing character for substantial ecosystem-scale consequences. More-
these two beachgrass species (A. arenaria over, hybridization between the Ammophila
~25 mm and A. breviligulata ~2 mm in length; beachgrasses could produce a highly invasive
Hitchcock and Chase 1951). At the time of dis- taxon or increase the invasive potential of either
covery, other morphological differences observed parent species, leading to a change in invasion
included intermediate leaf width, which sug- dynamics or dominant beachgrass.
gested that these plants could be hybrids.
Here, we present genetic and cytological evi- METHODS
dence that confirms the hybrid origin of the
newly discovered beachgrass and a morphologi- Between 2013 and 2018, we collected whole
cal comparison between the hybrid and its plants, inflorescences, and leaf tissue samples of
v www.esajournals.org 3 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
A. arenaria, A. breviligulata, and/or the putative served as an internal standard with known
hybrid from as many as eight sites spanning a diploid genome size (2C = 8.75 pg; Greilhuber
190 km stretch of Oregon and Washington coast- et al. 2007). Samples were mixed with 400 lL of
line (Fig. 1C; see Appendix S1: Tables S1, S2 for nuclei extraction buffer (CyStain ultraviolet Pre-
collection information). We air-dried the individ- cise P Nuclei Extraction Buffer; Sysmex Partec,
ual plants (shoots attached to a single rhizome; Go €rlitz, Germany), chopped with a razor blade,
N = 523) and inflorescences (a single shoot with and then filtered through a 50-µm nylon mesh
flowers; N = 249) prior to conducting morpho- filter. The nuclei were stained by adding 1.6 mL
logical measurements. Leaf tissue samples were 40 ,6-diamidino-2-phenylindole (DAPI) to the tis-
either stored on ice and processed within 24 h sue mixture. Samples were analyzed on a Partec
for flow cytometry (N = 24) or desiccated and PA II flow cytometer (Munster, Germany) and
stored in silica gel for DNA extraction (N = 32). run until at least 3000 nuclei were counted. Gen-
To measure the morphology and growth form ome size was calculated as 2C = genome size of
of the plants, we measured individual plant standard 9 (mean fluorescence value of sam-
weight including aboveground biomass (all bio- ple mean fluorescence value of standard). Dif-
mass 1 cm above the primary node) and below- ferences between the genome sizes of each taxon
ground biomass (all biomass, including roots were assessed using one-factor ANOVA and
and rhizomes, below 1 cm above the primary Tukey’s HSD tests.
node) ligule length, number of vertical and hori- Finally, we conducted SNP genotyping to mea-
zontal shoots, shoot length, leaf number, leaf sure the potential genetic contribution of the two
length, leaf width, total rhizome length, number Ammophila species to the hybrid. Leaf tissue from
of rhizome nodes, and rhizome internode length 32 total plants at five sites was used (see
(see Hacker et al. 2019 for methods). For each Appendix S1: Table S2 for sample information).
inflorescence, we measured culm length (from DNA was extracted from 15 mg of leaf tissue
the node to tip of the inflorescence), panicle bio- using the E.Z.N.A. Plant DNA Kit (Omega Bio-
mass, panicle length (from the base of the inflo- Tek, Norcross, Georgia, USA) following the man-
rescence to the tip), panicle width (measured at ufacturer’s instructions. DNA quality was
the widest point), floret length, callus hair (hairs assessed with 1% agarose gel electrophoresis,
at the base of the floret) length, and rachilla and DNA quantity was measured using an Accu-
(small stem baring the spikelet) length. We ana- Blue NextGen dsDNA Quantitation Kit (Biotium,
lyzed the resulting data using R version 3.6.1 (R Fremont, California, USA) in the SpectraMax M3
Core Team 2019). Outliers were assessed for all plate reader (Molecular Devices, San Jose, Cali-
traits using a threshold of 10 times the mean fornia, USA).
Cook’s distance (Cook 1977). Outliers were Genotyping libraries followed the 2b Restric-
detected and subsequently removed from two tion Site-Associated DNA (2b-RAD) protocol
traits (panicle length and plant height). We used from Wang et al. (2012) with minor modifications
one-factor ANOVAs or Kruskal–Wallis tests (in- (Appendix S2). Libraries were pooled in equimo-
cluding Tukey’s honestly significant difference lar amounts and multiplexed in a single lane as
[HSD] post hoc test, when appropriate), in addi- 100-bp single reads on an Illumina HiSeq 3000 at
tion to principal component analysis (PCA), to Oregon State University’s Center for Genome
determine whether the two Ammophila species Research and Biocomputing (Corvallis, Oregon,
and the putative hybrid differed in their mor- USA). After quality-filtering the reads, de novo
phology and growth form. loci were generated by clustering across samples
To test for hybridization, we measured the and genotypes called for each individual. These
genome size of the plants using flow cytometry. steps were performed using methods described
Leaf tissue from three individuals of each of the in Wang et al. (2012) with publicly available
two Ammophila species and the putative hybrid scripts (https://github.com/Eli-Meyer/2brad_utili
was analyzed (see Appendix S1: Table S1 for col- ties). For the full details of SNP genotyping
lection information). Leaf tissue (~0.3 cm2) was methods, see Appendix S2.
placed in a plastic petri dish with the same area From the resulting dataset of 1870 SNPs, we
of leaf tissue from Pisum sativum Ctirad, which identified markers that would be informative for
v www.esajournals.org 4 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
verifying the parentage of the putative hybrids. The genome sizes of A. arenaria, A. breviligu-
We used the R package SNPRelate (Zheng et al. lata, and the putative hybrid all differed from
2012) to calculate the minor allele frequency for one another (mean standard deviation:
both Ammophila species at each locus and then 6.39 pg 0.05, 7.26 pg 0.07, and 6.80 pg
identified nearly fixed differences (major allele 0.05, respectively). Compared with its parents,
frequency >80%) between the two species. This the putative hybrid genome is intermediate in
filtering revealed 47 SNPs that were diagnostic size (Appendix S3: Fig. S1). Moreover, there was
between the two Ammophila species. We then no overlap in estimated genome size among the
used the program STRUCTURE (Pritchard et al. three taxa. Finally, the 95% confidence interval of
2000) to test for evidence of hybridization by the hybrid genome size (6.76–6.84 pg) encom-
clustering the putative hybrids based on their passes the expected midparent genome size
genotypes at these 47 loci. The population origin (6.83 pg), the predicted genome size for an F1
of each sample belonging to the two Ammophila diploid hybrid. Using the known chromosome
species was pre-specified and STRUCTURE was counts and genome sizes of the parent species,
used to predict the population of origin of the we can estimate the 2N chromosome count for
hybrid samples. The number of groups was set the hybrid (Ayres et al. 2008a). Both Ammophila
to 2, lambda was fixed at 1, and the migration parent species have 2N = 28 chromosomes (Bow-
prior was set to 0.05. Fifteen iterations were run den 1960, Zonneveld et al. 2005). The estimated
with 50,000 burn-in steps (sufficient for key average mass per chromosome for each species is
parameters to reach equilibrium) and 50,000 run 0.228 pg for A. arenaria (for a total of 6.39 pg
steps. The STRUCTURE output from all runs across 28 chromosomes per nucleus) and
was collated using CLUMPP (Jakobsson and 0.259 pg for A. breviligulata (7.26 pg per
Rosenberg 2007) and visualized using DIS- nucleus). A first-generation hybrid cross with a
TRUCT (Rosenberg 2003). The same SNP dataset genome comprised of 1N from A. arenaria and
was also examined via PCA on their covariance 1N from A. breviligulata would have an
matrix using SNPrelate (Zheng et al. 2012). estimated genome size of 6.82 pg ((14 chromo-
somes 9 0.228 pg per chromosome) + (14
RESULTS chromosomes 9 0.259 pg per chromosome)).
These calculations imply that the hybrid, found
Several notable morphological differences here to have a genome size of 6.80 pg, is a first-
were identified between the two beachgrass spe- generation cross between A. arenaria and A. bre-
cies and the putative hybrid (Fig. 2; viligulata and is not an allopolyploid.
Appendix S3: Table S1). The putative hybrid was To verify the origin of the putative hybrid, we
intermediate between the two Ammophila species calculated the hybrid allele frequency at genome
in ligule length (Fig. 2A) and leaf width sites diagnostic for both Ammophila species. The
(Fig. 2B), characters important for field identifi- expected allele frequency for an F1 hybrid at
cation. The putative hybrid exceeded both SNPs that are fixed between parent species is 0.5
Ammophila species in shoot height (Fig. 2C) and (Fitzpatrick 2012). For all hybrid samples, the
was intermediate in shoot number (Fig. 2D). average minor allele frequency at these diagnos-
Additionally, there were differences in inflores- tic SNPs was between 0.44 and 0.49 (mean =
cence morphology: the panicles of the putative 0.45). The PCA on the parental diagnostic SNP
hybrid were heavier than either Ammophila spe- dataset groups hybrid individuals between the
cies (Fig. 2F) although they were of intermediate two Ammophila species, which are clearly differ-
length (Fig. 2E). A PCA grouped samples by entiated from each other (Fig. 4A). As with the
taxon with the putative hybrid clustering morphological data (Fig. 3), the A. arenaria clus-
between the two Ammophila species (Fig. 3). Dif- ter has the widest spread, indicating that this
ferences in morphology within each taxon (PC1) group contains the highest level of SNP diversity.
explained the most variation (70.3%) while differ- In contrast, the hybrids cluster together closely,
ences among taxa (PC2) explained 25.5% of the indicating that there are very few SNP differ-
variation. Notably, A. arenaria had the greatest ences between individuals. Four A. arenaria sam-
within-species morphological variation (Fig. 3). ples and one A. breviligulata sample are clustered
v www.esajournals.org 5 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
Fig. 2. Comparisons (mean standard error) of grass species morphology, including (A) ligule length, (B) leaf
width, (C) shoot height, (D) number of shoots per plant, (E) panicle length, and (F) panicle weight of the two
beachgrass species and the hybrid (abbreviations in Fig. 1). Results of one-factor ANOVAs and Kruskal–Wallis
rank sum tests are given in plots. Bars that do not share letters are statistically different (Tukey’s honestly signifi-
cant difference P ≤ 0.05). P < 0.0001, P < 0.00001.
with the hybrids. Four of these five samples are sample), which results in their positioning near
missing significant data (i.e., some of the species- zero on both PCA axes. Finally, the STRUCTURE
differentiating SNPs were not sequenced for this analysis suggested that the hybrids are of
v www.esajournals.org 6 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
Fig. 3. Dimensional reduction on morphological
data using principal component analysis. Dashed lines
represent 95% confidence ellipses for each species (spe-
cies abbreviations in Fig. 1) based on a multivariate t
distribution. The first principal component is most Fig. 4. Genomic ancestry of hybrids. Both (A) princi-
strongly correlated with shoot height while the second pal component analysis and (B) STRUCTURE analysis
principal component is most strongly correlated with were performed with 47 species-diagnostic single
ligule length. nucleotide polymorphism loci. Dashed lines in A rep-
resent 95% confidence ellipses for each species (abbre-
viations in Fig. 1) based on a multivariate t
distribution.
intermediate population ancestry (Fig. 4B), with
an average ancestry of 55% A. arenaria and 45%
A. breviligulata. We show that the hybrid has some morpholog-
ical traits that are intermediate between the two
DISCUSSION Ammophila species (i.e., ligule length, leaf width,
and callus hair length; Fig. 2A, B, Appendix S3:
The morphological, cytological, and genomic Table S1) and a few traits for which it exceeds
data presented here all support the hypothesis both parents (i.e., tiller height, panicle width and
that the distinct Ammophila morphotype recently biomass, and spikelet and rachilla length;
discovered on the Pacific Northwest coast is an Fig. 2C, F, Appendix S3: Table S1). F1 hybrids
F1 hybrid between A. arenaria and A. breviligu- may experience heterosis, or hybrid vigor, which
lata. The hybrids we identify are undoubtedly a can produce exceptional traits and likely explain
unique and unintended consequence of the some of these novel phenotypic values. The taller
sequential introduction and subsequent invasion shoots and larger inflorescences of the hybrid
of two non-native beachgrass species to the Paci- suggest greater energy input into growth and
fic Northwest over the last century. Although reproduction.
both species have been introduced to a number The cytological and genomic evidence is con-
of coastlines (Hertling and Lubke 1999, Hilton cordant with our hypothesis that the putative
et al. 2006), as far as we know the Pacific North- hybrid is an F1 cross between the two Ammophila
west is the only place in the world where the two species. The genome sizes of the three taxa are
Ammophila species significantly overlap in their significantly different and non-overlapping, and
range and thus have had the opportunity to the hybrid genome size and number of chromo-
hybridize. somes are as predicted for a F1 hybrid
v www.esajournals.org 7 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
(Appendix S3: Fig. S1). Although we do not yet using the same technique, A. arenaria has a much
know if the hybrid can sexually reproduce, the longer history of planting on the U.S. Pacific
lack of allopolyploidy (chromosome duplication coast (Seabloom and Wiedemann 1994, Wiede-
following hybridization) and the large differ- mann and Pickart 1996), which perhaps explains
ences between average chromosome sizes of the the higher genetic diversity we observed in that
parent species suggest that the hybrid is likely species. Further study is needed to understand
sterile (Rieseberg and Willis 2007). Allopoly- the relationship between planting patterns in the
ploidy can lead to previously sterile hybrids Pacific Northwest, the population genetic diver-
developing sexual reproduction, which can sity of both A. arenaria and A. breviligulata in
increase spread and speciation as has been seen their invasive ranges, and the impact of these
in invasive Spartina (Ayres and Strong 2001) and patterns on hybridization success.
Salsola (Ayres et al. 2009). Further research into The direction of hybridization (i.e., which par-
the potential for sexual reproduction and moni- ent species is the pollen donor and which is the
toring for changes in the ploidy level of the seed parent) is unknown. To infer the direction
hybrid will be increasingly important as we con- of parentage, we are planning to sequence sec-
sider the possibilities of speciation, backcrossing, tions of the chloroplast genome, which is exclu-
and spread. sively maternally inherited in most grasses
The species-diagnostic SNP data clearly indi- (Harris and Ingram 1991). The majority of the
cate that the putative hybrid is an F1 cross hybrid stands are found on dunes that are domi-
between the two beachgrass species (Fig. 4). At nated by A. breviligulata with very low A. arenar-
this point, we have found no evidence of later ia abundance (Hacker et al. 2012, Biel et al. 2019).
generation hybrids or backcrossing, which We hypothesize that in these sites, the high ratio
would produce genome sizes intermediate to the of A. breviligulata to A. arenaria flowers leads to
F1s and their parent species. Genetic variation pollen swamping, likely producing hybrids with
among hybrids and their current geographic an A. arenaria seed parent and an A. breviligulata
range of 190 km of the coast suggests that there pollen parent. The only known hybrid stand that
have likely been multiple independent is on an A. arenaria-dominated dune is found in
hybridization events. However, both Ammophila Pacific City, Oregon. Comparison of patterns of
parent species have been shown to disperse long chloroplast genome inheritance between hybrids
distances during winter storm events, which from A. arenaria- and A. breviligulata-dominated
erode rhizomes and stems from beaches and sites will help us better determine the direction
deposit them in different locations (Maun 1984, of hybridization and the ecological and biologi-
Konlechner et al. 2016). This mechanism could cal controls on hybrid formation.
contribute to further spread of the hybrid with- Besides the population genetic consequences
out additional hybridization events. Ayres et al. of the hybridization, there are ecological and
(2004) documented the dramatic spread, a 100- ecosystem service implications as well. Past
fold increase in area in 30 yr, of the hybrid cord- research shows that beachgrass shoot density,
grass, S. alterniflora 9 S. foliosa, in San Francisco height, and biomass are important characteristics
Bay, California. The authors attributed this in determining species-specific differences in
spread to both high asexual growth rate and sand capture (Zarnetske et al. 2012, Biel et al.
seedling germination, an example that further 2019, Hacker et al. 2019) and competition (Zar-
emphasizes the need to understand the sexual netske et al. 2013). The number of shoots per
and asexual reproductive potential of the Ammo- plant was found to be highest in A. arenaria and
phila hybrid. lowest in A. breviligulata (Fig. 2D), in agreement
The low genetic diversity observed in A. bre- with earlier studies of beachgrass functional mor-
viligulata in this study is congruent with previous phology (Hacker et al. 2012). The hybrid had an
studies of out-planting using this species. Clonal intermediate number of shoots per plant but was
propagules typically come from limited source not statistically different from the trait average
populations (Carlson et al. 1991), resulting in for A. breviligulata (Fig. 2D). Additionally, the
field populations with low genetic diversity hybrid exceeded both parents in shoot height,
(Slaymaker et al. 2015). Although propagated belowground biomass, and average rhizome
v www.esajournals.org 8 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
internode distance (Appendix S3: Table S1). This There is a history of hybrid grasses being
combination of taller plants with intermediate preferentially planted for their value as sedi-
density and a greater investment in rhizomes has ment stabilizers. In Europe, a hybrid between
the potential to produce dunes that are as wide A. arenaria and Calamagrostis epigejos is more
as those produced by A. breviligulata but signifi- vigorous than A. arenaria, growing more later-
cantly taller. Proliferation of the hybrid could ally with longer and wider leaf blades, and is
result in dunes intermediate in morphology therefore preferentially planted in dune stabi-
between those dominated by A. arenaria and lization projects (Rihan and Gray 1985). S. an-
A. breviligulata. However, both Ammophila spe- glica, a polyploid hybrid species of cordgrass,
cies have been shown to be phenotypically plas- has been intentionally planted in marshes in
tic (Gray 1985) and to have growth rates and Puget Sound, Washington, USA (Hacker et al.
competition interactions that respond strongly to 2001), and across China (Zhang et al. 2004,
their environment (Zarnetske et al. 2013). There- Chung 2006) to capture sediment and minimize
fore, studies of the morphological differences erosion. A. arenaria 9 breviligulata may have
between the parent species and their hybrid properties that make it an exceptional or valu-
under controlled environmental conditions, espe- able dune builder and could be of interest to
cially sand deposition and climate, will be critical coastal planners and land managers. However,
for predicting the effect of this novel taxon on any plans to intentionally plant the hybrid must
dune morphology and related ecosystem ser- consider its invasion potential and effects on the
vices. The two beachgrass species both nega- genetic diversity of its parent species. Addition-
tively impact native coastal shorebird and plant ally, introduction of the hybrid into the Great
populations but do so to different degrees Lakes native range of A. breviligulata could have
(Hacker et al. 2012, Biel et al. 2017). Specifically, irreversible negative consequences for popula-
A. arenaria-dominated dunes tend to have lower tions already threatened by land use change and
plant species diversity than those dominated by genetic swamping from restoration (Fant et al.
A. breviligulata (Hacker et al. 2012) and removal 2008, Slaymaker et al. 2015). Both Ammophila
of both species improves breeding habitat for the species are still intentionally planted in their
threatened Western snowy plover (Zarnetske native and invasive ranges as dune stabilizers.
et al. 2010, Biel et al. 2017). Given the intermedi- Considering the ability of these two species to
ate morphological characteristics of the hybrid, hybridize, introducing them into the same range
we hypothesize that it will also negatively affect should be carefully considered. Clearly, more
both native plant biodiversity and plover habitat research into the effects of the hybrid on impor-
value. The effects of a possible spread of the tant ecosystem functions and services is neces-
Ammophila hybrid on biodiversity conservation sary to fully understand the management
are unknown but potentially detrimental. implications of this discovery.
It is interesting to consider how the spread of Monitoring the size and expansion of this new
the hybrid beachgrass might affect dune mor- hybrid zone is of critical management impor-
phology. Expansion of the hybrid beachgrass tance along the Pacific Northwest coast, where
into areas dominated by A. arenaria (i.e., spread any change in dominant beachgrass species as a
to the south of its current range) might decrease result of the spread of this new invader could
dune height and compromise coastal protection. have substantial effects on dune geomorphology,
However, expansion of the hybrid on dunes coastal protection, and biodiversity conservation
currently dominated by A. breviligulata would (Seabloom et al. 2013, Biel et al. 2017, 2019).
likely increase dune height and increase coastal Additionally, further study into the reproductive
protection. A more nuanced understanding of interactions between the hybrid and its parents,
the likelihood of spread of the hybrid beach- including studies of flowering phenology, repro-
grass, and the consequences for dune shape and ductive compatibility, and tests for introgression,
coastal protection is of critical management is essential for determining the potential effects
importance, especially with relation to the dif- on the population genetics and invasibility in
ferent invasion contexts posed by the two par- beachgrasses, a critical ecosystem engineer in
ent species. coastal dune systems.
v www.esajournals.org 9 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
ACKNOWLEDGMENTS Capinha, C., F. Essl, H. Seebens, D. Moser, and H. M.
Pereira. 2015. The dispersal of alien species rede-
We thank Dr. R. Contreras for use of his flow fines biogeography in the Anthropocene. Science
cytometer; D. Batson, A. David, and J. Henderson for 348:1248–1251.
help with fieldwork during the initial hybrid discov- Carlson, J. R., F. F. Reckendorf, and W. Ternyik. 1991.
ery; M. Barker, M. Marie-Rose, and E. Tom, who mea- Stabilizing coastal sand dunes in the Pacific North-
sured hundreds of grass samples; and J. Borland, who west. Agriculture Handbook 687. USDA Soil Con-
completed the first morphological analysis of the servation Service, West National Technical Center,
hybrid. This work was supported by an NSF Graduate Portland, Oregon, USA.
Research Fellowship and a Washington Native Plant Castillo, J. M., D. R. Ayres, P. Leira-Doce, J. Bailey, M.
Society grant to RSM, and a NOAA Oregon Sea Grant Blum, D. R. Strong, T. Luque, and E. Figueroa.
(NA18OAR4170072) to SDH and FB. 2010. The production of hybrids with high ecologi-
cal amplitude between exotic Spartina densiflora
LITERATURE CITED and native S. maritima in the Iberian Peninsula.
Diversity and Distributions 16:547–558.
Anderson, E., and G. L. Stebbins. 1954. Hybridiza- Charbonneau, B. R., R. Nicoletta, and L. S. Wootton.
tion as an evolutionary stimulus. Evolution 8:378– 2020. A decade of expansion of the invasive plant
388. Carex kobomugi in a coastal foredune system. Bio-
Ayres, D. R., E. Grotkopp, K. Zaremba, C. M. Sloop, logical Invasions 22:2099–2112.
M. J. Blum, J. P. Bailey, C. K. Anttila, and D. R. Chunco, A. J. 2014. Hybridization in a warmer world.
Strong. 2008a. Hybridization between invasive Ecology and Evolution 4:2019–2031.
Spartina densiflora (Poaceae) and native S. foliosa in Chung, C.-H. 2006. Forty years of ecological engineer-
San Francisco Bay, California, USA. American Jour- ing with Spartina plantations in China. Ecological
nal of Botany 95:713–719. Engineering 27:49–57.
Ayres, D. R., K. Zaremba, C. M. Sloop, and D. R. Cook, R. D. 1977. Detection of influential observation
Strong. 2008b. Sexual reproduction of cordgrass in linear regression. Technometrics 19:15–18.
hybrids (Spartina foliosa 9 alterniflora) invading Cooper, W. S. 1958. Coastal sand dunes of Oregon and
tidal marshes in San Francisco Bay. Diversity and Washington. Geological Society of America, Boul-
Distributions 14:187–195. der, Colorado, USA.
Ayres, D., F. J. Ryan, E. Grotkopp, J. Bailey, and J. Cooper, W. S. 1967. Coastal dunes of California. Geolog-
Gaskin. 2009. Tumbleweed (Salsola, section Kali) ical Society of America, Boulder, Colorado, USA.
species and speciation in California. Biological David, A. S., P. L. Zarnetske, S. D. Hacker, P. Ruggiero,
Invasions 11:1175–1187. R. G. Biel, and E. W. Seabloom. 2015. Invasive con-
Ayres, D. R., D. L. Smith, K. Zaremba, S. Klohr, and D. geners differ in successional impacts across space
R. Strong. 2004. Spread of exotic cordgrasses and and time. PLOS ONE 10:e0117283.
hybrids (Spartina sp.) in the tidal marshes of San Ellstrand, N. C., and K. A. Schierenbeck. 2000.
Francisco Bay, California, USA. Biological Inva- Hybridization as a stimulus for the evolution of
sions 6:221–231. invasiveness in plants? Proceedings of the National
Ayres, D. R., and D. R. Strong. 2001. Origin and Academy of Sciences USA 97:7043–7050.
genetic diversity of Spartina anglica (Poaceae) using Fant, J. B., R. M. Holmstrom, E. Sirkin, J. R. Etterson,
nuclear DNA markers. American Journal of Botany and S. Masi. 2008. Genetic structure of threatened
88:1863–1867. native populations and propagules used for
Biel, R. G., S. D. Hacker, and P. Ruggiero. 2019. Eluci- restoration in a clonal species, American beach-
dating coastal foredune ecomorphodynamics in grass (Ammophila breviligulata Fern.). Restoration
the US Pacific Northwest via Bayesian networks. Ecology 16:594–603.
Journal of Geophysical Research: Earth Surface Fei, S., J. Phillips, and M. Shouse. 2014. Biogeomorphic
124:1919–1938. impacts of invasive species. Annual Review of
Biel, R. G., S. D. Hacker, P. Ruggiero, N. Cohn, and E. Ecology, Evolution, and Systematics 45:69–87.
W. Seabloom. 2017. Coastal protection and conser- Fitzpatrick, B. M. 2012. Estimating ancestry and
vation on sandy beaches and dunes: context-de- heterozygosity of hybrids using molecular mark-
pendent tradeoffs in ecosystem service supply. ers. BMC Evolutionary Biology 12:1–14.
Ecosphere 8:e01791. Gray, A. J. 1985. Adaptation in perennial coastal
Bowden, W. M. 1960. Chromosome numbers and taxo- plants-with particular reference to heritable varia-
nomic notes on Northern grasses: III. Twenty-five tion in Puccinellia maritima and Ammophila arenaria.
genera. Canadian Journal of Botany 38:541–557. Vegetatio 61:179–188.
v www.esajournals.org 10 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
Greilhuber, J., E. M. Temsch, and J. C. M. Loureiro. Maun, M. A. 1984. Colonizing ability of Ammophila bre-
2007. Nuclear DNA content measurement. Pages viligulata through vegetative regeneration. Journal
67–101 in J. Doleel, J. Greilhuber, and J. Suda, edi- of Ecology 72:565–574.
tors. Flow cytometry with plant cells. Wiley-VCH Maun, M. A. 2009. The biology of coastal sand dunes.
Verlag GmbH & Co. KGaA, Weinheim, Germany. Oxford University Press, Oxford, UK.
Hacker, S. D., and M. N. Dethier. 2006. Community Pritchard, J. K., M. Stephens, and P. Donnelly. 2000.
modification by a grass invader has differing Inference of population structure using multilocus
impacts for marine habitats. Oikos 113:279–286. genotype data. Genetics 155:945–959.
Hacker, S. D., D. Heimer, C. E. Hellquist, T. G. Reeder, R Core Team. 2019. R: a language and environment for
B. Reeves, T. Riordan, and M. N. Dethier. 2001. A statistical computing. R Foundation for Statistical
marine plant (Spartina anglica) invades widely Computing, Vienna, Austria.
varying habitats: potential mechanisms of invasion Rieseberg, L. H., M. A. Arther, and R. K. Wayne. 2001.
and control. Biological Invasions 3:211–217. Transgressive segregation, adaptation and specia-
Hacker, S. D., K. R. Jay, N. Cohn, E. B. Goldstein, P. A. tion. Heredity 83:363–372.
Hovenga, M. Itzkin, L. J. Moore, R. S. Mostow, E. Rieseberg, L. H., and J. H. Willis. 2007. Plant specia-
V. Mullins, and P. Ruggiero. 2019. Species-specific tion. Science 317:910–914.
functional morphology of four US Atlantic coast Rihan, J. R., and A. J. Gray. 1985. Ecology of the hybrid
dune grasses: biogeographic implications for dune marram grass 9 Calammophila baltica in Britain.
shape and coastal protection. Diversity 11:1–16. Vegetatio 61:203–208.
Hacker, S. D., P. Zarnetske, E. Seabloom, P. Ruggiero, J. Rosenberg, N. A. 2003. distruct: a program for the
Mull, S. Gerrity, and C. Jones. 2012. Subtle differ- graphical display of population structure. Molecu-
ences in two non-native congeneric beach grasses lar Ecology Notes 4:137–138.
significantly affect their colonization, spread, and Ruggiero, P., S. Hacker, E. Seabloom, and P. Zarnetske.
impact. Oikos 121:138–148. 2018. The role of vegetation in determining dune
Harris, S. A., and R. Ingram. 1991. Chloroplast DNA morphology, exposure to sea-level rise, and storm-
and biosystematics: the effects of intraspecific diver- induced coastal hazards: a U.S. Pacific Northwest
sity and plastid transmission. Taxon 40:393–412. perspective. Pages 337–361 in L. J. Moore and A. B.
Hertling, U. M., and R. A. Lubke. 1999. Use of Ammo- Murray, editors. Barrier dynamics and response to
phila arenaria for dune stabilization in South Africa changing climate. Springer International Publish-
and its current distribution; perceptions and prob- ing, Cham, Switzerland.
lems. Environmental Management 4:467–482. Salmon, A., M. L. Ainouche, and J. F. Wendel. 2005.
Hesp, P. 2002. Foredunes and blowouts: initiation, geo- Genetic and epigenetic consequences of recent
morphology and dynamics. Geomorphology hybridization and polyploidy in Spartina (Poaceae).
48:245–268. Molecular Ecology 14:1163–1175.
Hilton, M., N. Harvey, A. Hart, K. James, and C. Seabloom, E. W., P. Ruggiero, S. D. Hacker, J. Mull,
Arbuckle. 2006. The impact of exotic dune grass and P. Zarnetske. 2013. Invasive grasses, climate
species on foredune development in Australia and change, and exposure to storm-wave overtopping
New Zealand: a case study of Ammophila arenaria in coastal dune ecosystems. Global Change Biology
and Thinopyrum junceiforme. Australian Geogra- 19:824–832.
pher 37:313–334. Seabloom, E. W., and A. M. Wiedemann. 1994. Distri-
Hitchcock, A. S., and A. Chase. 1951. Manual of the bution and effects of Ammophila breviligulata Fern.
grasses of the United States. Volume 2. United States (American Beachgrass) on the foredunes of the
Department of Agriculture, Washington, D.C., USA. Washington coast. Journal of Coastal Research
Jakobsson, M., and N. A. Rosenberg. 2007. CLUMPP: a 10:178–188.
cluster matching and permutation program for Seebens, H., et al. 2017. No saturation in the accumula-
dealing with label switching and multimodality in tion of alien species worldwide. Nature Communi-
analysis of population structure. Bioinformatics cations 8:1–9.
23:1801–1806. Simberloff, D., et al. 2013. Impacts of biological inva-
Konlechner, T. M., D. A. Orlovich, and M. J. Hilton. sions: what’s what and the way forward. Trends in
2016. Restrictions in the sprouting ability of an Ecology & Evolution 28:58–66.
invasive coastal plant, Ammophila arenaria, from Slaymaker, D. H., M. S. Peek, J. Wresilo, D. C. Zeltner,
fragmented rhizomes. Plant Ecology 217:521–532. and Y. F. Saleh. 2015. Genetic structure of native
Mallet, J. 2005. Hybridization as an invasion of the and restored populations of American beachgrass
genome. Trends in Ecology & Evolution 20:229– (Ammophila breviligulata Fern.) along the New Jersey
237. coast. Journal of Coastal Research 31:1334–1343.
v www.esajournals.org 11 April 2021 v Volume 12(4) v Article e03501MOSTOW ET AL.
Strong, D. R., and D. A. Ayres. 2009. Spartina introduc- Zarnetske, P. L., S. D. Hacker, E. W. Seabloom, P. Rug-
tions and consequences in salt marshes: arrive, sur- giero, J. R. Killian, T. B. Maddux, and D. Cox. 2012.
vive, thrive, and sometimes hybridize. Pages 3–22 Biophysical feedback mediates effects of invasive
in B. R. Silliman, E. Grosholz, and M. D. Bertness, grasses on coastal dune shape. Ecology 93:1439–
editors. Human impacts on salt marshes: a global 1450.
perspective. University of California Press, Berke- Zarnetske, P. L., E. W. Seabloom, and S. D. Hacker.
ley, California, USA. 2010. Non-target effects of invasive species man-
Wang, S., E. Meyer, J. K. McKay, and M. V. Matz. 2012. agement: beachgrass, birds, and bulldozers in
2b-RAD: a simple and flexible method for genome- coastal dunes. Ecosphere 1:art13.
wide genotyping. Nature Methods 9:808–810. Zhang, R. S., Y. M. Shen, L. Y. Lu, S. G. Yan, Y. H.
Wiedemann, A. M., and A. Pickart. 1996. The Ammo- Wang, J. L. Li, and Z. L. Zhang. 2004. Formation of
phila problem on the Northwest coast of North Spartina alterniflora salt marshes on the coast of
America. Landscape and Urban Planning 34:287– Jiangsu Province, China. Ecological Engineering
299. 23:95–105.
Wissemann, V. 2007. Plant evolution by means of Zheng, X., D. Levine, J. Shen, S. M. Gogarten, C. Lau-
hybridization. Systematics and Biodiversity 5:243– rie, and B. S. Weir. 2012. A high-performance com-
253. puting toolset for relatedness and principal
Zarnetske, P. L., T. C. Gouhier, S. D. Hacker, E. W. Sea- component analysis of SNP data. Bioinformatics
bloom, and V. A. Bokil. 2013. Indirect effects and 28:3326–3328.
facilitation among native and non-native species Zonneveld, B. J. M., I. J. Leitch, and M. D. Bennett.
promote invasion success along an environmental 2005. First nuclear DNA amounts in more than 300
stress gradient. Journal of Ecology 101:905–915. Angiosperms. Annals of Botany 96:229–244.
SUPPORTING INFORMATION
Additional Supporting Information may be found online at: http://onlinelibrary.wiley.com/doi/10.1002/ecs2.
3501/full
v www.esajournals.org 12 April 2021 v Volume 12(4) v Article e03501You can also read

















































