Evolutionarily conserved and divergent regulatory sequences in the fish rod opsin promoter
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399
www.elsevier.com/locate/cbpb
Evolutionarily conserved and divergent regulatory sequences
in the fish rod opsin promoter
Shoji Kawamura*, Kumiko Takeshita, Taro Tsujimura, Satoshi Kasagi, Yoshifumi Matsumoto
Department of Integrated Biosciences, Graduate School of Frontier Sciences, The University of Tokyo, Kashiwa, Chiba 277-8652, Japan
Received 3 December 2004; received in revised form 13 March 2005; accepted 15 March 2005
Available online 17 June 2005
Abstract
Fish have multiple types and subtypes of opsin genes that are expressed in a highly regulated manner in retinal photoreceptor cells. In the
rod opsin proximal promoter region (RPPR) of zebrafish (Danio rerio), the BAT 1 regulatory region contains highly conserved OTX
(GATTA) and OTX-like (TATTA) sequences that can be recognized by the mammalian cone – rod homeobox (CRX) protein. However,
binding of zebrafish crx to the OTX sequence has remained elusive. In contrast to the BAT 1 region, the Ret 1 region, located approximately
20 bp upstream of the BAT 1 region in mammals, is not conserved in zebrafish. In the Ret 1 region, even the core OTX-like sequence
(AATTA sequence in mammals) is destructed. We show in this study that a region between Ret 1 and BAT 1 (denoted IRB, Inter-Ret 1-BAT
1) is highly conserved among fish species. Using electrophoretic mobility shift assay (EMSA), we show that zebrafish crx binds to the
conserved OTX sequence and that the fish-specific IRB region specifically binds elements present in both retinal and brain nuclear extracts of
zebrafish. These results imply that the regulatory mechanisms of opsin gene expression consist not only of evolutionarily conserved but also
of divergent machinery among different animal taxa.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Zebrafish; Medaka; Rod opsin; CRX; OTX; BAT 1; Ret 1; EMSA
1. Introduction known for rod opsins. Studies of mammalian rod opsin
genes have identified a number of cis-acting regulatory
Vertebrate retinal photoreceptor cells can be classified as elements, such as Ret 1 (PCE-1), BAT 1 (OTX), NRE, and
rods or cones, with the latter being further classified into Ret 4, found within an approximate 300-bp upstream region
multiple types. Typically, one photoreceptor cell produces from the transcription initiation site of the gene [rod opsin
only one type of opsin (protein moiety of visual pigments), proximal promoter region (RPPR)] (Yu and Barnstable,
achieving its specialized scotopic or photopic visual 1994; Chen and Zack, 1996; DesJardin and Hauswirth,
function. On the basis of evolutionary relatedness, the 1996; Kumar et al., 1996). Studies also have identified
vertebrate visual opsins are classified into one rod opsin (or photoreceptor- or retina-specific trans-acting transcription
rhodopsin) type (RH1) and four cone opsin types (M/LWS, factors involved in rod opsin expression: CRX (Chen et al.,
RH2, SWS1, and SWS2) having distinct peak absorption 1997; Furukawa et al., 1997b), NRL (Swaroop et al., 1992;
spectra (Yokoyama, 2000). The regulatory mechanisms of Rehemtulla et al., 1996; Mitton et al., 2000), QRX (Wang et
the cell-type specific expression of the opsin genes are best al., 2004), and NR2E3 (Cheng et al., 2004). These factors
interact with each other synergistically.
Zebrafish have all five types of the visual opsin genes,
with multiple subtypes: SWS1, SWS2, RH2-1, RH2-2, RH2-
* Corresponding author. Tel.: +81 4 7136 5422; fax: +81 4 7136 3692. 3, RH2-4, LWS-1, LWS-2, and RH1 (Chinen et al., 2003).
E-mail address: kawamura@k.u-tokyo.ac.jp (S. Kawamura). These opsin genes are expressed in a highly regulated
1096-4959/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.cbpc.2005.03.008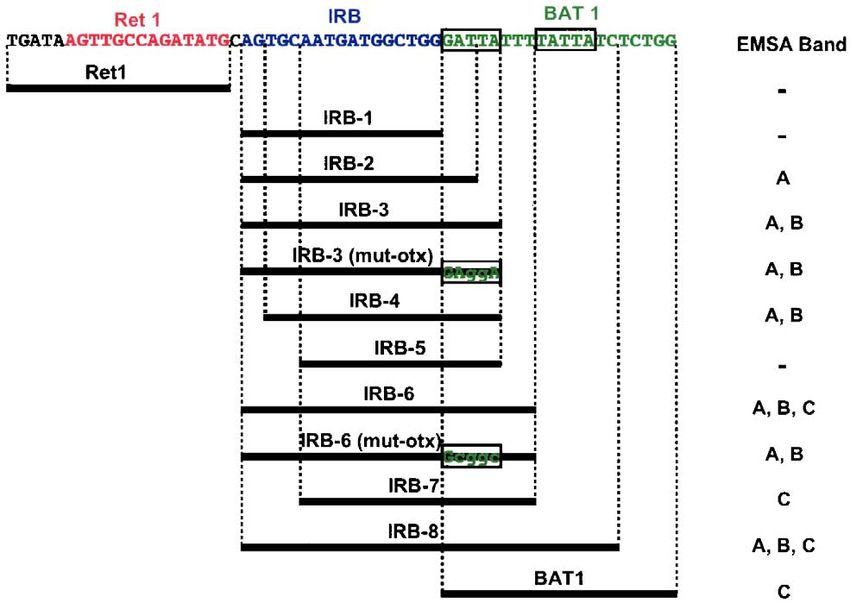
392 S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399
manner in retinal photoreceptor cells (Raymond et al., 1993, Nuclear extracts from carp retina have been shown to bind
1995; Robinson et al., 1993, 1995; Schmitt et al., 1999; oligonucleotides containing the OTX sequence in an electro-
Vihtelic et al., 1999; Takechi and Kawamura, 2005). Tight phoretic mobility shift assay (EMSA), in which use of
gene regulation and the feasibility of performing devel- antibody against a mammalian CRX peptide sequence
opmental genetics have made zebrafish an excellent model resulted in faint super-shifted bands (Ma et al., 2001).
system in which to study the differentiation of the retina Zebrafish and carp belong to the superorder, Ostario-
(Malicki, 2000). Among the multiple visual opsin genes of physi, while the pufferfish belongs to Acanthopterygii; the
zebrafish, promoter analysis is most advanced in the rod two superorders were separated 115 –200 million years ago
opsin gene, RH1 (Kennedy et al., 2001; Hamaoka et al., (Furutani-Seiki and Wittbrodt, 2004). To better understand
2002). The 1.1-kb upstream region of zebrafish RH1 has the regulatory mechanisms of fish visual opsin expression,
been shown to be sufficient to drive reporter gene which possibly consists of both conserved and divergent
expression in rod photoreceptor cells in an identical mechanisms from mammals (Su et al., 2000; Zhang et al.,
spatio– temporal manner as endogenous RH1 (Hamaoka et 2003), we sequenced the RPPR of medaka, an acanthopter-
al., 2002). In the RPPR, the BAT 1 and NRE sequences, ygiian fish, to investigate sequence conservation and
originally identified in mammals, are conserved in zebra- divergence among fish. We show here that the HD of
fish, while the Ret 1 and Ret 4 sequences are not conserved zebrafish crx directly binds to OTX sequence by EMSA. We
(Kennedy et al., 2001). Studies of pufferfish and carp rod also show that a conserved, fish-specific sequence, located
opsin genes also revealed a different degree of conservation between the Ret 1 and BAT 1 regions (designated IRB) in
between the BAT 1/NRE and Ret 1/Ret 4 regions (Su et al., the RPPR, is specifically bound by retinal and brain nuclear
2000; Zhang et al., 2003). The BAT 1 sequence contains the extracts of zebrafish by EMSA.
OTX motif, GATTA, to which CRX binds directly in
mammals (Chen et al., 1997; Furukawa et al., 1997b;
Kimura et al., 2000), while NRE is the binding site for NRL 2. Materials and methods
in mammals (Kumar et al., 1996; Rehemtulla et al., 1996).
Although the zebrafish NRL has not been identified, its 2.1. Cloning of medaka rod opsin gene
CRX gene (crx) has been isolated and analyzed for its role
in retinogenesis (Liu et al., 2001; Shen and Raymond, Genomic DNA was extracted from a single medaka
2004). specimen (Oryzias latipes; HNI strain). A genomic library
CRX is a member of the otd/otx family of the paired-like with a total of 7.5 105 recombinant plaques was con-
homeobox proteins (Chen et al., 1997; Furukawa et al., structed using BamHI-digested EMBL3 E-phage vector and
1997b). Mammalian CRX is expressed predominantly in Sau3A I partially digested genomic DNA (12 –20 kb). For
retinal photoreceptor cells and pinealocytes, regulates probe preparation, total RNA was extracted from medaka eye
expression of many photoreceptor-specific genes [i.e., tissue and the full coding region of the rod opsin cDNA was
interphotoreceptor retinoid-binding protein (IRBP), arrestin, amplified by RT-PCR using oligonucleotide primers
and opsins] and pineal-specific genes [i.e., pineal night- designed from its published nucleotide sequence (Hisatomi
specific ATPase (PINA)], and plays a significant role in et al., 1997). The DNA probe was labeled with [a-32P] dCTP
differentiation and maintenance of photoreceptor cells (Chen using the random primer method. Plaque hybridization was
et al., 1997; Freund et al., 1997; Furukawa et al., 1997b, carried out at 55 -C in a solution consisting of 6 SSC,
1999; Li et al., 1998; Bibb et al., 2001). However, 5 Denhardt’s solution, 0.5% sodium dodecyl sulfate (SDS),
phylogenetic analyses indicated that mammalian CRX is a and 5 Ag/mL E. coli DNA. The hybridized membranes were
divergent and uniquely evolved member of the Otx5/Crx washed four times in 1 SSC containing 0.1% SDS at 55 -C
orthology class of the vertebrate otd/otx family (Plouhinec et (20 min each), which allows an approximate mismatch of
al., 2003). Unlike mammalian CRX, zebrafish crx does not 30% (Sambrook and Russel, 2001).
regulate expression of circadian genes or photoreceptor- Four overlapping clones (EMD31, EMD35, EMD65,
specific genes including opsins in the pineal gland, and is and EMD133) containing the rod opsin gene were isolated
expressed not only in adult rod and cone cells, but also in (Fig. 1A). Restriction fragments that hybridized to the
proliferating retinal progenitor cells, suggesting possible screening probe were subcloned into pBluescript II (SK-)
involvement in early optic primordium patterning and in plasmid vectors. DNA sequencing was carried out for both
promoting differentiation of retinal progenitor cells (Liu et strands using an Applied Biosystems automatic sequencer
al., 2001; Gamse et al., 2002; Shen and Raymond, 2004). (model 3100) using the Big Dye Terminator v3.1 Cycle
Zebrafish crx was found to have 50% amino acid identity Sequencing Kit.
with human CRX, and 85% identity within the homeodo-
main (HD) region (Liu et al., 2001). Although zebrafish crx 2.2. Southern hybridization
can transactivate the bovine rod opsin promoter by interact-
ing with bovine NRL, its activity is weak (Liu et al., 2001) Approximately 2 Ag per lane of medaka (HNI strain)
and its OTX-binding capability has not been directly tested. genomic DNA was digested with restriction enzymes,S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399 393
A B B G E H S
(kb)
23.1
9.4
SBS B E G G H EG E H G HS E BH S
6.6
5’ 3’
4.4
λMD65
λ
λMD133
1 kb
2.3
2.0
Fig. 1. Genomic structure of the medaka rod opsin gene. (A) Restriction map of the rod opsin gene. The isolated phage clones (EMD65 and EMD133) are
indicated. The coding region is indicated by a solid box with orientation of transcription given. (B) Southern hybridization of the rod opsin cDNA to medaka
genomic DNA. Lambda HindIII size standards are indicated in kb. B: BamHI; E: EcoRI; G: BglII; H: HindIII; S: SacI. The GenBank accession number of the
isolated medaka rod opsin gene is AB180742.
electrophoresed on a 0.5% agarose gel, and transferred to a 2.4. Preparation of nuclear extract
positively charged nylon membrane (Biodyne B, Pall) using
the VacuGene vacuum blotting system (Pharmacia). A 361- Retina, brain, and decapitated body of adult zebrafish
bp region, from the initiation codon of the medaka rod opsin were homogenized in phosphate buffer saline (PBS) and
cDNA (corresponding to the exon 1 coding region of all centrifuged. The resulting pellet was resuspended in 10
terrestrial vertebrate rod opsin genes), was labeled as mM HEPES (pH 7.8), 10 mM KCl, 0.1 mM EDTA, (pH
described above and used as a probe for genomic Southern 8.0), 0.1% NP-40, 1 mM dithiothreitol (DTT), 0.5 mM
hybridization. Hybridization and washing were carried out PMSF, 2 Ag/mL aprotinin, and 2 Ag/mL leupeptin, and
as for genomic library screening using a washing temper- centrifuged. The pellet was resuspended in 50 mM HEPES
ature of 65 -C that allows an approximate mismatch of 20% (pH 7.8), 420 mM KCl, 0.1 mM EDTA (pH 8.0), 5 mM
(Sambrook and Russel, 2001). MgCl2, 20% glycerol, 1 mM DTT, 0.5 mM PMSF, 2 Ag/
mL aprotinin, and 2 Ag/mL leupeptin, gently mixed at 4
2.3. Production and purification of CRX HD -C for 30 min, and centrifuged. The supernatant (¨0.5 Ag/
AL) was analyzed on a 10% SDS-PAGE and was used as
A DNA fragment encoding the zebrafish crx HD and the nuclear extract.
six N-terminal and six C-terminal flanking amino acid
residues from the HD (residues 32 –103) and a DNA 2.5. EMSA
fragment encoding the human CRX HD and six N-
terminal and two C-terminal flanking amino acid residues For EMSA probe construction, complementary oligo-
from the HD (residues 33 – 100), were amplified by PCR nucleotides were annealed, labeled on their 5’-ends with
from their respective full-length cDNAs (Chen et al., [c-32P] ATP and T4 polynucleotide kinase (TOYOBO),
1997; Liu et al., 2001). Primer pairs for the zebrafish and and purified using the QIAquick Nucleotide Removal Kit
human cDNAs were the following: 5V-gccgtcggatccccagc- (QIAGEN). For the binding reaction, 1 – 100 ng of
cactccgaggaag-3V/5V-acgggggaattcctgaccgctggtctgctg-3Vand 5V- purified GST-CRX (HD) fusion protein or 2.5 – 5 Ag of
gctgtgggatccccaagcgcccccaggaag-3V/ 5V- ctgctggaattcctg- nuclear extract was used. The binding reaction, containing
ctgtcgctgctgcct-3V, respectively, [BamH I (ggatcc) or EcoR 15 mM Tris (pH 7.5), 60 mM KCl, 0.5 mM DTT, 7.5%
I (gaattc) sites are underlined and six extra nucleotides glycerol, 5 Ag/AL BSA, 0.1 Ag/AL poly (dI-dC), and 100
were attached to facilitate the cloning procedure]. Ampli- fmol 32P-labeled probe, was carried out in a volume of 20
fied DNA was cloned into the glutathione S-transferase AL. In the competition assay, 1 – 100 pmol (10 –1000 fold
(GST) vector, pGEX-4T-2 (Amersham Pharmacia). The molar excess) of the non-radiolabeled DNA probe was
resulting GST fusion construct was sequenced to confirm added to the standard mixture. After incubation at room
the reading frame and to exclude PCR-induced mutations. temperature for 1 h, samples were loaded onto 5%
The construct was expressed in E. coli strain BL21 and polyacrylamide gels in 0.5 Tris borate-EDTA (TBE)
purified using Glutathione-Sepharose 4B (Amersham buffer and electrophoresed at 10 V/cm for 2 h at room
Pharmacia). Purified protein was analyzed using 10% temperature. The gels were then dried and images
SDS-PAGE and detected using anti-GST antibody (Amer- obtained using X-ray film or the BAS-5000 imaging
sham Pharmacia). analyzer (Fuji Film).394 S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399
3. Results and discussion within the coding region, typical of teleost fish rod opsin
genes (Fitzgibbon et al., 1995). Southern hybridization
3.1. Genomic organization of the medaka rod opsin gene using the rod opsin cDNA probe allowed detection of a
single major band in all five restriction digests tested, and
Fig. 1A shows the genomic organization of the isolated their sizes were concordant with the restriction maps of
medaka rod opsin gene. Its nucleotide sequence shows six their genomic clones (Fig. 1B). While a minor band
synonymous and one non-synonymous substitutions in the appeared in the EcoR I digest, its nature remains to be
coding region from that of the published medaka rod opsin elucidated. However, no other rod opsin sequence was
cDNA sequence (Hisatomi et al., 1997). The non-synon- found in the medaka genome database (http://dolphin.lab.
ymous substitution corresponds to an amino acid difference nig.ac.jp/medaka/) suggesting that this band may corre-
at residue 10, Tyr and Asn of our isolated gene and the spond to an ortholog of exo-rhodopsin that is closely related
published sequence, respectively. It was noted that this Tyr to teleost rod opsin in the RH1 gene family (Mano et al.,
residue is completely conserved among other vertebrate 1999; Philp et al., 2000). These results strongly suggest that
RH1 and RH2 opsins studied to date. There was no intron the medaka rod opsin gene is a single copy gene and that
Fig. 2. Alignment of the rod opsin proximal promoter region among vertebrates. Positions of Ret 1, IRB, BAT 1, NRE, Ret 4, the TATA box, and the ATG
initiation codon are indicated. The reported transcription initiation sites are underlined. The conserved OTX sequence (GATTA) in the BAT 1 region and the
OTX-like sequence (AATTA) in the Ret 1 region are boxed. Nucleotides conserved among all the fish genes (zebrafish, two carp paralogs, pufferfish and
medaka) are highlighted in green whereas those among all others (X. laevis, chicken, bovine, mouse and human) are blue. Nucleotides conserved among nine
or all ten vertebrate sequences are indicated in red. Genbank accession numbers for the sequences are AF331797 (zebrafish), AJ012013 (carp type 1),
AJ012014 (carp type 2), U23808 (Xenopus), M98497 (chicken), M55171 (mouse), and U16824 (human). The pufferfish sequence was retrieved from the
Takifugu rubripes genome contig scaffold 830 (http://www.fugu-sg.org) and the bovine sequence was obtained from Zack et al. (1991).S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399 395
Table 1 1993), is known to be the binding site for the RX/RAX
Sequences of the OTX and OTX-mut probes for recombinant CRX HD transcription factor (Kimura et al., 2000), a key regulator of
proteins used in EMSA
eye development (Furukawa et al., 1997a; Mathers et al.,
OTX 5VACCAACTGGATTAAACTCAGC 3V
1997). Even this core sequence was found to be disrupted in
3VTGGTTGACCTAATTTGAGTCG 5V
OTX-mut 5VACCAACTGGAGATCTCTCAGC the fish genes. It was noted that the region between the Ret 1
3VTGGTTGACCTCTAGAGAGTCG 5V and BAT 1 sequences was highly conserved among fish but
The OTX sequence is indicated in boldface. Mutations are underlined. not among the mammals, chicken, and Xenopus. We named
this region the IRB (Inter-Ret 1-BAT 1) region. We have not
identified any database sequences with significant similarity
the genomic region isolated in our study contains an to IRB.
unrearranged copy of it.
3.3. Binding of zebrafish crx HD to the OTX sequence
3.2. Sequence alignment of RPPR among vertebrates
We next evaluated the binding activity of the zebrafish
Including the medaka sequence, vertebrate RPPRs crx HD to the conserved OTX sequence. The OTX probe
[zebrafish (Kennedy et al., 2001), two paralogous rod opsin sequence (Table 1) used for EMSA, containing GATTA,
genes of carp (Su et al., 2000), pufferfish (Zhang et al., was designed based on the probe sequences tested for
2003), Xenopus laevis (Mani et al., 2001), chicken human CRX by Kimura et al. (2000). The zebrafish crx
(Sheshberadaran and Takahashi, 1994), bovine (Zack et HD interacted with the probe as strongly as human CRX
al., 1991), mouse (al-Ubaidi et al., 1990), and human HD (Fig. 3A). When we used a mutated probe (OTX-mut,
(Bennett et al., 1995)] were aligned (Fig. 2). One OTX Furukawa et al., 1997b) (Table 1), its binding to zebrafish
sequence (GATTA) in the BAT 1 region was found crx HD was much weaker (Fig. 3B). Binding of zebrafish
completely conserved throughout all vertebrate sequences. crx HD to the OTX probe was inhibited by addition of the
Another OTX sequence in the BAT 1 region was less non-radioactive OTX probe in a dose-dependent manner,
conserved (TATTA in fish and CGTTA in mouse). The 5V but binding was not inhibited by the addition of the non-
NRE region was also highly conserved. In contrast, the Ret radioactive OTX-mut probe (Fig. 3C), as shown previously
1 region, which was highly conserved among mammals, in a similar experiment using mouse CRX (Furukawa et
chicken, and Xenopus, was not evident in the fish genes. al., 1997b). These results indicate that zebrafish crx has an
The core consensus sequence of the Ret 1 region, an OTX- equivalent binding specificity to mammalian CRX for
like AATTA sequence (Morabito et al., 1991; Kikuchi et al., OTX.
A B C
zCRX Competitor
hCRX Probe OTX OTX-mut
OTX OTX-mut
zCRX
1 2 3 4 5 6 1 2 3 4 5 6
1 2 3 4 5 6
Fig. 3. EMSA using the OTX and OTX-mut probes with recombinant CRX HD proteins. (A) The OTX probe was mixed with human CRX HD (hCRX) (lanes
1 – 3) or zebrafish crx HD (zCRX) (lanes 4 – 6). Amounts of protein used were 1 ng (lanes 1 and 4), 10 ng (lanes 2 and 5), and 100 ng (lanes 3 and 6). (B) The
zCRX was mixed with the OTX probe (lanes 1 – 3) or the OTX-mut probe (lanes 4 – 6). The amount of protein used was as in (A). (C) Competition assay using
the non-radiolabeled OTX and OTX-mut for the OTX/zCRX reaction. For 100 fmol of radiolabeled OTX probe and 100 ng of zCRX, 1 pmol (lanes 1 and 4),
10 pmol (lanes 2 and 5), or 100 pmol (lanes 3 and 6) of cold competitor were added.396 S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399
Fig. 4. Schematic of Ret 1, BAT 1, and a series of IRB oligonucleotide probes, covering the Ret 1 to BAT 1 regions, of the zebrafish RPPR. The OTX (GATTA)
and OTX-like (TATTA) sequences in the BAT 1 region is boxed. Mutated OTX sequences in the oligonucleotide probes are also boxed. Double-stranded
oligonucleotides were used for EMSA. The EMSA banding patterns for the nuclear extract from the zebrafish retina are indicated on the right.
3.4. Binding of retinal nuclear extract to the IRB region these band shifts, EMSA was performed using the series of
probes and purified zebrafish crx HD. EMSA results
Conservation of the IRB region only in fish species revealed that the probes yielding band C in the presence
implies that this region may serve as a novel cis-acting of the nuclear extract always bound the purified crx HD but
regulatory sequence, specific for fish rod opsin expression. the other probes did not (Fig. 5B). In addition, when IRB-3
For a genomic region to be a cis-regulatory element, it must and IRB-6 were mutated in the OTX region [probes IRB-3
be recognized by a trans-acting transcription factor. Our (mut-OTX) and IRB-6 (mut-OTX) in Fig. 4] and mixed
preliminary DNase I footprint experiment using zebrafish with the retinal nuclear extract, bands A and B were not
retinal nuclear extract showed protection of this region (data affected but band C disappeared. These results strongly
not shown). To initially characterize possible IRB function, suggest that the nuclear factor resulting in band C is crx and
we tested the binding activity of the IRB region to the that the factors resulting in bands A and B do not require a
nuclear extract from zebrafish retina using EMSA.
A series of EMSA probes, covering the Ret 1 to BAT 1
regions of zebrafish, was prepared (Fig. 4). Using the Ret 1 A B
probe, no band shift appeared, as predicted from the lack of IRB-3 IRB-2 IRB-6 Probe
core OTX-like sequence (AATTA) (Kimura et al., 2000)
IRB-1
IRB-3
BAT1
IRB-2
IRB-6
IRB-8
Ret1
NE
conserved among mammals, chicken, and Xenopus (see Fig. - + ++ - + ++ - + ++
2). When using only the IRB region (probe IRB-1), again no
band shift appeared. However, when the probe was
extended 3 bp toward the BAT 1 region (probe IRB-2), a B
single band shift appeared (band A in Fig. 5A). When the C CRX
A
probe included the entire OTX sequence (probe IRB-3), an
additional band was detected (band B in Fig. 5A). Extension
of an additional 3 bp (probe IRB-6) resulted in yet another
band (band C in Fig. 5A). The intensities of these band
shifts were dependent on the amount of nuclear extract
mixed with the probes (Fig. 5A).
The zebrafish crx was expected to bind to the probes Fig. 5. EMSA using the IRB probes for the zebrafish retinal nuclear extract
containing the OTX sequence, as demonstrated in Fig. 3. To (A) and for the recombinant zebrafish crx HD (B). (A) The three band shifts
examine whether crx may correspond to one or more of (A, B and C) are shown. Amount of nuclear extract: +, 2.5 Ag; ++, 5 Ag.S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399 397
specific nucleotide sequence in the OTX region and hence, IRB-2 IRB-3 BAT1 Probe
are distinct from crx.
Retina
Retina
Retina
Brain
Brain
Brain
Body
Body
Body
To identify nucleotide sequences necessary for resulting Nuclear Extract
in bands A and B, we first tested two probes, IRB-4 and
IRB-5, which lack 2 bp and 5 bp from the IRB-3 at the
Ret 1 side, respectively (Fig. 4). The abovementioned 2 bp
are not conserved between zebrafish/carp and pufferfish B
(Takifugu rubripes)/medaka (Fig. 2) and, as expected, C
deletion of the 2 bp did not affect the band shift pattern. In A
contrast, IRB-5, which lacks the additional 3 bp conserved
in fish (Fig. 2), yielded no band shift. When the same 5 bp
was deleted from IRB-6 (probe IRB-7 in Fig. 4), only
bands A and B disappeared while band C was unaffected.
We then introduced point mutations in the IRB-3 and IRB-
2 probes (Fig. 6). When a conserved 3 bp was mutated
[IRB-3 (mut-1)], only band B disappeared, implying that
putative nuclear factors resulting in bands A and B are
distinct and that the latter factor requires a specific
nucleotide sequence while the former does not. While Fig. 7. EMSA for nuclear extracts from the retina, brain, and decapitated
mutations to the next 5-bp region [IRB-3 (mut-2) and IRB- body of zebrafish using the IRB-2, IRB-3, and BAT 1 probes.
3 (mut-3)] affected both bands A and B, other mutations
affected only one of the two, implying that factors
resulting in bands A and B have distinct but overlapping these putative factors appeared to compete for binding to
recognition sequences in the IRB region. Furthermore, the IRB region because the IRB-2 probe, yielding only
band A when intact, resulted in band B when mutated
EMSA Band [IRB-2 (mut-2) and IRB-2 (mut-3)].
IRB-3 AGTGCAATGATGGCTGGGATTA A, B We then asked whether the putative nuclear factors
IRB-3(mut-1) --GTA----------------- A were specific to the retina or not since vertebrate visual
IRB-3(mut-2) -----CC--------------- - opsins are known to be expressed in the pineal gland and
IRB-3(mut-3) -------GTC------------ -
IRB-3(mut-4) ----------GTT--------- B other brain regions (Kawamura and Yokoyama, 1997;
IRB-3(mut-5) -------------AGT------ A Okano et al., 2000; Wada et al., 2000; Forsell et al., 2001;
IRB-3(mut-6) ----------------TTC--- A Masuda et al., 2003). Since the zebrafish pineal organ was
too small for us to collect a sufficient amount of nuclear
IRB-2 AGTGCAATGATGGCTGGGAT A
IRB-2(mut-1) ----------G--------- -
extract for EMSA, whole brain was used for the assay.
IRB-2(mut-2) -----------T-------- B When the IRB-2 and IRB-3 probes were tested, the
IRB-2(mut-3) ------------T------- A, B EMSA banding pattern for the brain nuclear extract was
IRB-2(mut-4) -------------AG----- A identical to that of retina, with IRB-2 hybridizing to band
IRB-2(mut-5) ---------------T---- A
IRB-2(mut-6) ----------------T--- A A and IRB-3 hybridizing to bands A and B (Fig. 7).
However, when using the BAT 1 probe, band C was not
IRB-3(mut-otx)
detected in brain nuclear extract. This finding is consistent
IRB-3(mut-1)
IRB-3(mut-2)
IRB-3(mut-3)
IRB-3(mut-4)
IRB-3(mut-5)
IRB-3(mut-6)
IRB-2(mut-1)
IRB-2(mut-2)
IRB-2(mut-3)
IRB-2(mut-4)
IRB-2(mut-5)
IRB-2(mut-6)
with the notion that band C corresponds to crx expressed
specifically in the retina and pinealocytes (Liu et al.,
IRB-3
IRB-2
IRB-2
IRB-3
2001), since pinealocytes represent only a minor fraction
of entire brain cells. No band shift was detected using any
probe when nuclear extract from the decapitated body was
B used.
B
A
A
4. Conclusions
We isolated the medaka rod opsin gene from a genomic
library. Comparison of nucleotide sequences of RPPR
among fish (zebrafish, carp, pufferfish, and medaka), X.
laevis, chicken, and mammals (bovine, mouse, and
Fig. 6. Mutation sequences in the IRB-3 and IRB-2 probes. The resulting human) revealed strict conservation of the OTX sequence
EMSA bands are indicated on the right and shown in the lower panel. in the BAT 1 region throughout the vertebrates. It also398 S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399
revealed genomic regions well-conserved only in fish. The DesJardin, L.E., Hauswirth, W.W., 1996. Developmentally important DNA
IRB region, located between the Ret 1 and BAT 1 elements within the bovine opsin upstream region. Invest. Ophthalmol.
Visual Sci. 37, 154 – 165.
regions, is one such region. Our EMSA results, using the Fitzgibbon, J., Hope, A., Slobodyanyuk, S.J., Bellingham, J., Bowmaker,
OTX sequence, IRB probes, CRX HD, and nuclear J.K., Hunt, D.M., 1995. The rhodopsin-encoding gene of bony fish
extracts, revealed that zebrafish crx binds to the OTX lacks introns. Gene 164, 273 – 277.
sequence as strongly as human CRX and that the Forsell, J., Ekstrom, P., Flamarique, I.N., Holmqvist, B., 2001. Expression
of pineal ultraviolet- and green-like opsins in the pineal organ and retina
zebrafish IRB region contains sequences recognized by
of teleosts. J. Exp. Biol. 204, 2517 – 2525.
at least two nuclear factors, distinct from crx, which are Freund, C.L., Gregory-Evans, C.Y., Furukawa, T., Papaioannou, M.,
produced in zebrafish retina and brain. Further studies to Looser, J., Ploder, L., Bellingham, J., Ng, D., Herbrick, J.A., Duncan,
examine other conserved genomic regions as well as A., Scherer, S.W., Tsui, L.C., Loutradis-Anagnostou, A., Jacobson,
studies to isolate putative nuclear factors for the IRB S.G., Cepko, C.L., Bhattacharya, S.S., McInnes, R.R., 1997. Cone – rod
region are needed. It is also important to evaluate IRB dystrophy due to mutations in a novel photoreceptor-specific homeobox
gene (CRX) essential for maintenance of the photoreceptor. Cell 91,
regulatory activity and the activity of its binding factors 543 – 553.
for rod opsin expression. These studies are of crucial Furukawa, T., Kozak, C.A., Cepko, C.L., 1997a. rax, a novel paired-type
importance to our understanding of the evolution of the homeobox gene, shows expression in the anterior neural fold and
opsin transcription machinery that possibly consists of developing retina. Proc. Natl. Acad. Sci. U. S. A. 94, 3088 – 3093.
both conserved and divergent components among different Furukawa, T., Morrow, E.M., Cepko, C.L., 1997b. Crx, a novel otx-like
homeobox gene, shows photoreceptor-specific expression and regulates
animal taxa. photoreceptor differentiation. Cell 91, 531 – 541.
Furukawa, T., Morrow, E.M., Li, T., Davis, F.C., Cepko, C.L., 1999.
Retinopathy and attenuated circadian entrainment in Crx-deficient mice.
Acknowledgements Nat. Genet. 23, 466 – 470.
Furutani-Seiki, M., Wittbrodt, J., 2004. Medaka and zebrafish, an evolu-
tionary twin study. Mech. Dev. 121, 629 – 637.
We greatly appreciate Dr. P.A. Raymond for zebrafish Gamse, J.T., Shen, Y.C., Thisse, C., Thisse, B., Raymond, P.A.,
crx cDNA, Drs. A. Kimura and T. Shinohara for human Halpern, M.E., Liang, J.O., 2002. Otx5 regulates genes that show
CRX cDNA, and Dr. K. Naruse, H. Mitani and A. Shima for circadian expression in the zebrafish pineal complex. Nat. Genet. 30,
medaka. This study was supported by Grants-in-Aid for 117 – 121.
Scientific Research (B) (12440243) and for Exploratory Hamaoka, T., Takechi, M., Chinen, A., Nishiwaki, Y., Kawamura, S., 2002.
Visualization of rod photoreceptor development using GFP-transgenic
Research (13874105) from the Japan Society for the zebrafish. Genesis 34, 215 – 220.
Promotion of Science. The manuscript was proofread by Hisatomi, O., Satoh, T., Tokunaga, F., 1997. The primary structure and
BioMed Proofreading Service. distribution of killifish visual pigments. Vision Res. 37, 3089 – 3096.
Kawamura, S., Yokoyama, S., 1997. Expression of visual and nonvisual
opsins in American chameleon. Vision Res. 37, 1867 – 1871.
Kennedy, B.N., Vihtelic, T.S., Checkley, L., Vaughan, K.T., Hyde, D.R.,
References 2001. Isolation of a zebrafish rod opsin promoter to generate a
transgenic zebrafish line expressing enhanced green fluorescent protein
al-Ubaidi, M.R., Pittler, S.J., Champagne, M.S., Triantafyllos, J.T., in rod photoreceptors. J. Biol. Chem. 276, 14037 – 14043.
McGinnis, J.F., Baehr, W., 1990. Mouse opsin. Gene structure and Kikuchi, T., Raju, K., Breitman, M.L., Shinohara, T., 1993. The proximal
molecular basis of multiple transcripts. J. Biol. Chem. 265, promoter of the mouse arrestin gene directs gene expression in
20563 – 20569. photoreceptor cells and contains an evolutionarily conserved retinal
Bennett, J., Sun, D., Kariko, K., 1995. Sequence analysis of the 5.34-kb 5V factor-binding site. Mol. Cell. Biol. 13, 4400 – 4408.
flanking region of the human rhodopsin-encoding gene. Gene 167, Kimura, A., Singh, D., Wawrousek, E.F., Kikuchi, M., Nakamura, M.,
317 – 320. Shinohara, T., 2000. Both PCE-1/RX and OTX/CRX interactions are
Bibb, L.C., Holt, J.K., Tarttelin, E.E., Hodges, M.D., Gregory-Evans, K., necessary for photoreceptor-specific gene expression. J. Biol. Chem.
Rutherford, A., Lucas, R.J., Sowden, J.C., Gregory-Evans, C.Y., 2001. 275, 1152 – 1160.
Temporal and spatial expression patterns of the CRX transcription Kumar, R., Chen, S., Scheurer, D., Wang, Q.L., Duh, E., Sung, C.H.,
factor and its downstream targets. Critical differences during human and Rehemtulla, A., Swaroop, A., Adler, R., Zack, D.J., 1996. The bZIP
mouse eye development. Hum. Mol. Genet. 10, 1571 – 1579. transcription factor Nrl stimulates rhodopsin promoter activity in
Chen, S., Zack, D.J., 1996. Ret 4, a positive acting rhodopsin regulatory primary retinal cell cultures. J. Biol. Chem. 271, 29612 – 29618.
element identified using a bovine retina in vitro transcription system. Li, X., Chen, S., Wang, Q., Zack, D.J., Snyder, S.H., Borjigin, J., 1998. A
J. Biol. Chem. 271, 28549 – 28557. pineal regulatory element (PIRE) mediates transactivation by the
Chen, S., Wang, Q.L., Nie, Z., Sun, H., Lennon, G., Copeland, N.G., pineal/retina-specific transcription factor CRX. Proc. Natl. Acad. Sci.
Gilbert, D.J., Jenkins, N.A., Zack, D.J., 1997. Crx, a novel otx-like U. S. A. 95, 1876 – 1881.
paired-homeodomain protein, binds to and transactivates photoreceptor Liu, Y., Shen, Y., Rest, J.S., Raymond, P.A., Zack, D.J., 2001. Isolation and
cell-specific genes. Neuron 19, 1017 – 1030. characterization of a zebrafish homologue of the cone rod homeobox
Cheng, H., Khanna, H., Oh, E.C., Hicks, D., Mitton, K.P., Swaroop, A., gene. Invest. Ophthalmol. Visual Sci. 42, 481 – 487.
2004. Photoreceptor-specific nuclear receptor NR2E3 functions as a Ma, G.C., Wang, T.M., Su, C.Y., Wang, Y.L., Chen, S., Tsai, H.J., 2001.
transcriptional activator in rod photoreceptors. Hum. Mol. Genet. 13, Retina-specific cis-elements and binding nuclear proteins of carp
1563 – 1575. rhodopsin gene. FEBS Lett. 508, 265 – 271.
Chinen, A., Hamaoka, T., Yamada, Y., Kawamura, S., 2003. Gene Malicki, J., 2000. Harnessing the power of forward genetics—analysis of
duplication and spectral diversification of cone visual pigments of neuronal diversity and patterning in the zebrafish retina. Trends
zebrafish. Genetics 163, 663 – 675. Neurosci. 23, 531 – 541.S. Kawamura et al. / Comparative Biochemistry and Physiology, Part B 141 (2005) 391 – 399 399
Mani, S.S., Batni, S., Whitaker, L., Chen, S., Engbretson, G., Knox, B.E., Robinson, J., Schmitt, E.A., Dowling, J.E., 1995. Temporal and spatial
2001. Xenopus rhodopsin promoter. Identification of immediate patterns of opsin gene expression in zebrafish (Danio rerio). Vis.
upstream sequences necessary for high level, rod-specific transcription. Neurosci. 12, 895 – 906.
J. Biol. Chem. 276, 36557 – 36565. Sambrook, J., Russel, D.W., 2001. Molecular cloning: a laboratory
Mano, H., Kojima, D., Fukada, Y., 1999. Exo-rhodopsin: a novel manual, 3rd ed. Cold Spring Harbor Laboratory Press, Cold Spring
rhodopsin expressed in the zebrafish pineal gland. Mol. Brain Res. Harbor.
73, 110 – 118. Schmitt, E.A., Hyatt, G.A., Dowling, J.E., 1999. Erratum: temporal and
Masuda, T., Iigo, M., Mizusawa, K., Aida, K., 2003. Retina-type rhodopsin spatial patterns of opsin gene expression in the zebrafish (Danio rerio):
gene expressed in the brain of a teleost, ayu (Plecoglossus altivelis). corrections with additions. Vis. Neurosci. 16, 601 – 605.
Zoolog. Sci. 20, 989 – 997. Shen, Y.C., Raymond, P.A., 2004. Zebrafish cone – rod (crx) homeobox
Mathers, P.H., Grinberg, A., Mahon, K.A., Jamrich, M., 1997. The Rx gene promotes retinogenesis. Dev. Biol. 269, 237 – 251.
homeobox gene is essential for vertebrate eye development. Nature 387, Sheshberadaran, H., Takahashi, J.S., 1994. Characterization of the chicken
603 – 607. rhodopsin promoter: identification of retina-specific and glass-like
Mitton, K.P., Swain, P.K., Chen, S., Xu, S., Zack, D.J., Swaroop, A., 2000. protein binding domains. Mol. Cell. Neurosci. 5, 309 – 318.
The leucine zipper of NRL interacts with the CRX homeodomain. A Su, C.Y., Lim, J., Tsai, H.J., 2000. Structural characterization and
possible mechanism of transcriptional synergy in rhodopsin regulation. transcriptional pattern of two types of carp rhodopsin gene. Comp.
J. Biol. Chem. 275, 29794 – 29799. Biochem. Physiol., B 125, 37 – 45.
Morabito, M.A., Yu, X., Barnstable, C.J., 1991. Characterization of Swaroop, A., Xu, J.Z., Pawar, H., Jackson, A., Skolnick, C., Agarwal,
developmentally regulated and retina-specific nuclear protein binding N., 1992. A conserved retina-specific gene encodes a basic
to a site in the upstream region of the rat opsin gene. J. Biol. Chem. 266, motif/leucine zipper domain. Proc. Natl. Acad. Sci. U. S. A. 89,
9667 – 9672. 266 – 270.
Okano, K., Okano, T., Yoshikawa, T., Masuda, A., Fukada, Y., Oishi, T., Takechi, M., Kawamura, S., 2005. Temporal and spatial changes in the
2000. Diversity of opsin immunoreactivities in the extraretinal tissues of expression pattern of multiple red and green subtype opsin genes during
four anuran amphibians. J. Exp. Zool. 286, 136 – 142. zebrafish development. J. Exp. Biol. 208, 1337 – 1345.
Philp, A.R., Bellingham, J., Garcia-Fernandez, J., Foster, R.G., 2000. A Vihtelic, T.S., Doro, C.J., Hyde, D.R., 1999. Cloning and character-
novel rod-like opsin isolated from the extra-retinal photoreceptors of ization of six zebrafish photoreceptor opsin cDNAs and immuno-
teleost fish. FEBS Lett. 468, 181 – 188. localization of their corresponding proteins. Vis. Neurosci. 16,
Plouhinec, J.L., Sauka-Spengler, T., Germot, A., Le Mentec, C., Cabana, T., 571 – 585.
Harrison, G., Pieau, C., Sire, J.Y., Veron, G., Mazan, S., 2003. The Wada, Y., Okano, T., Fukada, Y., 2000. Phototransduction molecules in the
mammalian Crx genes are highly divergent representatives of the Otx5 pigeon deep brain. J. Comp. Neurol. 428, 138 – 144.
gene family, a gnathostome orthology class of orthodenticle-related Wang, Q.L., Chen, S., Esumi, N., Swain, P.K., Haines, H.S., Peng, G.,
homeogenes involved in the differentiation of retinal photoreceptors and Melia, B.M., McIntosh, I., Heckenlively, J.R., Jacobson, S.G., Stone,
circadian entrainment. Mol. Biol. Evol. 20, 513 – 521. E.M., Swaroop, A., Zack, D.J., 2004. QRX, a novel homeobox gene,
Raymond, P.A., Barthel, L.K., Rounsifer, M.E., Sullivan, S.A., Knight, modulates photoreceptor gene expression. Hum. Mol. Genet. 13,
J.K., 1993. Expression of rod and cone visual pigments in goldfish and 1025 – 1040.
zebrafish: a rhodopsin-like gene is expressed in cones. Neuron 10, Yokoyama, S., 2000. Molecular evolution of vertebrate visual pigments.
1161 – 1174. Prog. Retin. Eye Res. 19, 385 – 419.
Raymond, P.A., Barthel, L.K., Curran, G.A., 1995. Developmental Yu, X., Barnstable, C.J., 1994. Characterization and regulation of the
patterning of rod and cone photoreceptors in embryonic zebrafish. protein binding to a cis-acting element, RET 1, in the rat opsin
J. Comp. Neurol. 359, 537 – 550. promoter. J. Mol. Neurosci. 5, 259 – 271.
Rehemtulla, A., Warwar, R., Kumar, R., Ji, X., Zack, D.J., Swaroop, A., Zack, D.J., Bennett, J., Wang, Y., Davenport, C., Klaunberg, B., Gearhart,
1996. The basic motif-leucine zipper transcription factor Nrl can J., Nathans, J., 1991. Unusual topography of bovine rhodopsin
positively regulate rhodopsin gene expression. Proc. Natl. Acad. Sci. promoter-lacZ fusion gene expression in transgenic mouse retinas.
U. S. A. 93, 191 – 195. Neuron 6, 187 – 199.
Robinson, J., Schmitt, E.A., Harosi, F.I., Reece, R.J., Dowling, J.E., 1993. Zhang, T., Tan, Y.H., Fu, J., Lui, D., Ning, Y., Jirik, F.R., Brenner, S.,
Zebrafish ultraviolet visual pigment: absorption spectrum, sequence, Venkatesh, B., 2003. The regulation of retina specific expression of
and localization. Proc. Natl. Acad. Sci. U. S. A. 90, 6009 – 6012. rhodopsin gene in vertebrates. Gene 313, 189 – 200.You can also read

















































