Detection of spatial localization of Hst-1/Fgf-4 gene expression in brain and testis from adult mice
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Oncogene (2000) 19, 3805 ± 3810
ã 2000 Macmillan Publishers Ltd All rights reserved 0950 ± 9232/00 $15.00
www.nature.com/onc
Detection of spatial localization of Hst-1/Fgf-4 gene expression in brain
and testis from adult mice
Hanako Yamamoto1, Takahiro Ochiya1, Yasushi Takahama2, Yasuo Ishii3, Noriko Osumi3,
Hiromi Sakamoto1 and Masaaki Terada*,1
1
National Cancer Center Research Institute, 1-1, Tsukiji 5-chome, Chuo-ku, Tokyo 104-0045, Japan; 2First Department of Surgery,
Nara Medical University, Nara 634-8522, Japan; 3Division of Biochemistry and Cell Biology, National Institute of Neuroscience,
National Center of Neurology and Psychiatry, 4-1-1 Ogawahigashi, Kodaira, Tokyo 187-8502, Japan
HST-1, a member of the ®broblast growth factor (FGF) Drucker et al., 1993). Murine Hst-1/Fgf-4 gene works as
family (FGF-4), has been shown to be a signaling an inducer of embryonic limb (Niswander et al., 1993,
molecule whose expression is essential for embryonic 1994; Ochiya et al., 1995) and skeletal muscle develop-
development. However, HST-1/FGF-4 expression has ment (Hannon et al., 1996). As to its biological activities,
not been detected or reported in adult tissues so far we have previously shown that it is a factor with a potent
analysed. To investigate whether there is a possible role angiogenic activity (Yoshida et al., 1994) as well as a
of HST-1/FGF-4 in adult stage, we have carried out a potent inducer of platelet production from megakar-
highly sensitive RT ± PCR analysis of Hst-1/Fgf-4 gene yocytes (Sakamoto et al., 1994; Konishi et al., 1995,
expression in adult mice tissues. Results show Hst-1/Fgf- 1996). Most of the other FGF family genes have their
4 gene expression in the nervous system, intestines, and expression not only in the embryonic stage but also in
testis of normal adult mice. In situ hybridization several adult tissues; however, it has been noted that the
technique was used to localize Hst-1/Fgf-4 gene HST-1/FGF-4 gene is normally dormant in adult tissues,
expression in the cerebellum and testis from 10-week- and the physiological functions of its production of adult
old mice. Cell type-speci®c gene expression was detected: bodies were still unknown. Recently, it was reported that
Purkinje cells in the cerebellum and Sertoli cells in testis. HST-1/FGF-4 gene expression in embryonal carcinoma
These ®ndings suggest that the Hst-1/Fgf-4 gene also cells required a synergistic interaction between Oct-3 and
plays an important role in adult tissues, and may oer Sox 2 on the HST-1/FGF-4 enhancer (Yuan et al., 1995).
insights into the biological signi®cance of HST-1/FGF-4 The transcriptional factor Oct-3 mRNA was detected in
in cerebellar and testicular functions. Oncogene (2000) ovarian granulosa cells, fallopian tube, myometrium,
19, 3805 ± 3810. cervix, breast, liver, adrenal gland, pituitary, hypothala-
mus, brain cortex, prostate, and in testis in the
Keywords: Hst-1/Fgf-4; brain; testis; in situ; RT ± PCR; cynomolgus monkey (Heikinheimo et al., 1995). Sox-2
adult is a member of a large family of genes that encode
transcription factors. Previous studies have shown that
this gene is predominantly expressed in the central
Introduction nervous system during embryonic development, crystal-
line lens, peripheral nervous system, digestive tract, and
The HST-1/FGF-4 gene, originally identi®ed as a the epithelium of chick lungs (Collignon et al., 1996).
transforming gene from human gastric cancer, encodes These results led us to postulate that there was a
®broblast growth factor-4, a 206-amino acid protein, and possibility of Hst-1/Fgf-4 gene expression in adult
is located on chromosome 11q13 (Yoshida et al., 1991). tissues. Here, we performed a highly sensitive RT ±
The gene was subsequently isolated and characterized in PCR analysis to elucidate the expression of the Hst-1/
several other solid malignant tumors and it was found Fgf-4 gene in adult mice tissues. By using our RT ± PCR
that this gene is often ampli®ed in several human tumors analysis, Hst-1/Fgf-4 gene expression was predominantly
(Tsuda et al., 1989; Theillet et al., 1989) and mainly detected in the nervous system, intestines, and testis of
expressed in human testicular tumors (Yoshida et al., normal adult mice, and was weakly recognized in other
1988). The FGF family has been shown to be mitogenic tissues. In situ hybridization laid open cell type-speci®c
toward a broad spectrum of mesodermal, ectodermal, Hst-1/Fgf-4 gene expression: Purkinje cells in the
and endodermal cells, inducing cell proliferation and cerebellum and Sertoli cells in the testis. These results
dierentiation in vitro (Yoshida et al., 1991). The suggest that Hst-1/Fgf-4 may also play some speci®c
expression of mouse Hst-1/Fgf-4 transcripts is restricted roles in adult tissue as well as in embryogenesis.
in primitive streak (E7.5 ± 8.5) paraxial presomitic
mesoderm in the trunk (E7.5 ± 11.5); primitive neuroec-
toderm (E8.0 ± 8.5); pharyngeal pouch endoderm (E8.5 ±
Results
9.5); bronchial arch ectoderm (E8.5 ± 9.5); limb apical
ectoderm (E10.5 ± 12.5), and skeletal myoblast groups
Hst-1/Fgf-4 gene expression in adult mice
(E9.5 ± 13.5) (Niswander et al., 1992; Suzuki et al., 1992;
We performed a highly sensitive RT ± PCR analysis to
elucidate the expression of the Hst-1/Fgf-4 gene in
*Correspondence: M Terada
adult mice tissues. We tried 16 combinations of primer
Received 22 May 2000; revised 19 June 2000; accepted 19 June sets, and several RT ± PCR conditions. The result was
2000 that we were able to obtain the most appropriate RT ±Hst-1/Fgf-4 expression in the adult mice
H Yamamoto et al
3806
PCR system for detection of mouse Fgf-4 expression at decreased thereafter. The Hst-1/Fgf-4 gene was also
present. The RT ± PCR experiments, including the clearly expressed in embryonal (E18.5), neonatal and
extraction of RNA, were repeated at least three times adult murine testis (Figure 2b). The expression in the
to con®rm the reproducibility of the results presented. testis was relatively high during embryonal develop-
Sequencing analysis of the RT ± PCR products revealed ment and decreased with sexual maturation. These
that the ampli®ed sequence was identical to murine results suggest that Hst-1/Fgf-4 gene expression in the
Hst-1/Fgf-4 gene. Following the semi-quantitative PCR brain and testis is switched on during the embryonic
ampli®cation, the PCR products were separated by stage and detected in these organs after birth, although
agarose gel electrophoresis, transferred onto a nylon its expression in other tissues such as muscle and limb
®lter, and hybridized under high stringency conditions is limited to the embryonic stage.
with a32P labeled mouse Fgf-4 fragment cDNA as a
probe. Under optimum conditions, our RT ± PCR/
Localization of Hst-1/Fgf-4 gene expression by in situ
Southern hybridization analysis revealed a sensitivity of
hybridization
10 copies/mg total RNA.
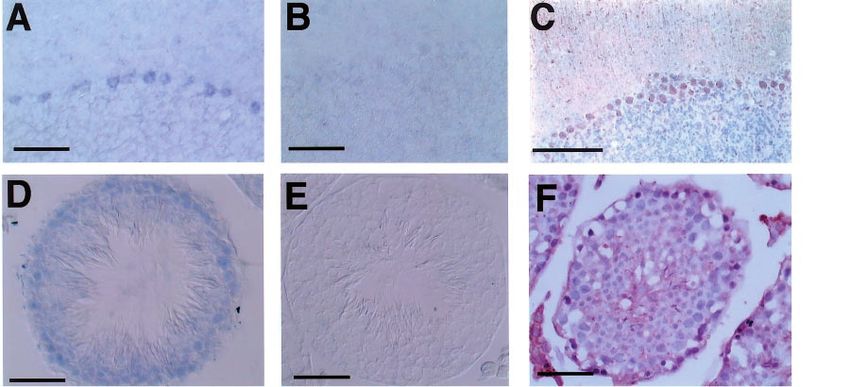
As shown in Figure 1, we detected expression of the To identify speci®cally which cells express the Hst-1/
Hst-1/Fgf-4 gene from 10-week-old mice in the Fgf-4 gene in the adult brain and testis, a 609 bp Hst-
cerebrum (26101 copies/reaction), cerebellum (26101 1/Fgf-4 antisense RNA probe was synthesized. To
copies/reaction), medulla oblongata (26102 copies/ know the accuracy of the Hst-1/Fgf-4 antisense RNA
reaction), spinal cord (26102 copies/reaction), ischiatic probe, fresh-frozen sections of embryos at E9.5 were
nerve (26102 copies/reaction), jejunum (26101 copies/ used for in situ hybridization analysis. The results show
reaction), ileum (36102 copies/reaction), vermiform that the antisense probe recognized the presence of
appendage (10 copies/reaction), colon (10 copies/ Hst-1/Fgf-4 mRNA in the apical ectodermal ridge of
reaction), and testis (26101 copies/reaction) by limbs and myotomes just as it did in the previous
ethidium staining of RT ± PCR products. These results reports (Suzuki et al., 1992; Drucker et al., 1993). In
indicate that the nervous system, digestive tract system, contrast, the sense RNA probe did not produce any
and germ cells do indeed have Hst-1/Fgf-4 gene signals. These results suggest that our in situ techniques
expression. Furthermore, in the same animals, we speci®cally capture the Hst-1/Fgf-4 signals in mice. We
could detect expression in the kidney, spleen, bone next performed in situ hybridization analysis of Hst-1/
marrow, lung, eyeball, and tongue by Southern blot Fgf-4 gene expression using the cerebellum and testis
analysis of RT ± PCR products, although their expres- from 10-week-old mice. In the sections of adult
sion level was very low (less than 10 copies/reaction). cerebellum, a DIG-labeled antisense RNA probe was
observed in the Purkinje cell layers (Figure 3a).
Immunohistochemical staining of a-calbindin which is
Detection of Hst-1/Fgf-4 gene expression in the
a speci®c marker for Purkinje cells (Figure 3c) was
developing brain and testis
used for the identi®cation of those cells. In all the
Reports show that Hst-1/Fgf-4 mRNA has not been Purkinje cells, strong signals were observed in the
detected in the brain at E8.0 ± E14.5 (Niswander et al., perikaryon. On the contrary, the dendrites of the
1992) and in postnatal (Ozawa et al., 1996) mice. To Purkinje cells were not stained. Weak labeling was also
determine the possible expression of the Hst-1/Fgf-4 found in the internal granule layer (Figure 3a). In
gene at a low amount in murine embryos, neonates and contrast, the sense RNA probe did not produce any
adults, a highly sensitive RT ± PCR method was signal (Figure 3b). In situ hybridization analysis
performed. The results show detection of Hst-1/Fgf-4 showed that Hst-1/Fgf-4 RNA was also clearly
mRNA in the cerebellum from embryos at days 14.5, detected in the spinal cord and subventricular zone of
16.5 and 18.5, at postnatal day 0, 6 and 10, and on the adult mouse brain (data not shown), although
weeks 5, 10 and 38 (Figure 2a). The expression level further study is required to determine speci®cally which
was predominantly high in embryos at E14.5 and cells express the Hst-1/Fgf-4 gene.
Figure 1 Expression pro®les of Hst-1/Fgf-4 transcripts in tissues of adult mice. RT ± PCR products ampli®ed with speci®c primers
were electrophoresed on 3.5% agarose gels, stained with ethidium bromide, and then photographed (a). The same RNA samples
were subjected to RT ± PCR analysis of b-actin transcripts to verify the integrity (b). DNAs from (a) were denatured and transferred
onto a nylon membrane, and then hybridized with a a32-labeled mouse Hst-1/Fgf-4 cDNA fragment as a probe (c). For semi-
quantitative analysis, cDNA plasmid of murine Hst-1/Fgf-4 sequences at variant amounts were used and 26102 molecules/reaction
were represented (lane P). Lanes 1, cerebrum; 2, cerebellum; 3, medulla oblongata; 4, spinal cord; 5, ischiatic nerve; 6, muscle; 7,
bone; 8, bone marrow; 9, blood cells; 10, heart; 11, lung; 12, eye ball; 13, tongue; 14, digit; 15, esophagus; 16, stomach; 17, jejunum;
18, ileum; 19, vermiform appendage; 20, colon; 21, liver; 22, pancreas; 23, kidney; 24, spleen; 25, testis; 26, prostate; 27, ovary; 28,
uterus; N, no RNA samples
OncogeneHst-1/Fgf-4 expression in the adult mice
H Yamamoto et al
3807
cultured cells showed that purity of Purkinje cells and
Sertoli cells from 10-week-old mice was 590% and
598%, respectively (data not shown). RT ± PCR
analysis on mRNAs from these cells was then
performed. The results show that Hst-1/Fgf-4 mRNA
was detected in the cultured Purkinje cells (Figure 4a)
and Sertoli cells (Figure 4b). Ampli®cation of IP3R1,
which is speci®cally expressed in the Purkinje cells, in
the same mRNA sample from the Purkinje culture as a
control, produced a strong signal (data not shown).
Each RNA sample ampli®ed a b-actin gene in parallel
as internal controls. As with the in situ hybridization
analysis, it was clearly demonstrated that Hst-1/Fgf-4
transcripts present in puri®ed cell fractions of both
Purkinje and Sertoli cells.
Discussion
In this current paper, we demonstrated that expression of
the Hst-1/Fgf-4 gene is retained in mice of neonates and
adult stages. The mRNA localization of Hst-1/Fgf-4 was
predominantly detected in the nervous system, intestines,
and testis of normal adult mice at 10-week-old, and was
weakly recognized in other tissues such as the kidney,
spleen, bone marrow, lung, eyeball, and tongue.
Although data are not shown, examination of adult
human cDNA libraries showed equivalent tissue-dis-
tribution of human HST-1/FGF-4 gene expression.
Further studies on murine cerebellum and testis by in
situ hybridization and cell puri®cation analysis showed
that the signal of Hst-1/Fgf-4 lies in Purkinje cells in the
cerebellum and in Sertoli cells in the testis of adult mice.
The molecular basis of brain development and
Figure 2 Hst-1/Fgf-4 transcripts in the developing brain and central nervous system (CNS) disorders remains poorly
testis. expression of Hst-1/Fgf-4 mRNAs in ICR mouse
cerebellums (a) at E14.5, E16.5, P0, P6, P10 and adult stages at
understood. Recently, it has been reported that Sonic
5-, 10- and 38-week old and testes (b) E18.5, P0, P6, P10 and hedgehog (SHH) signaling controls the development of
adult stages at 5-, 10- and 38-week old determined by RT ± PCR the cerebellum at multiple levels (Dahmane et al.,
analysis. RT ± PCR products were separated in 3.0% agarose gel, 1999). SHH is produced by Purkinje neurons, is
stained with ethidium bromide and photographed. DNAs were required for the proliferation of granule neuron
denatured and transferred onto a nylon membrane, and then
hybridized with a a32P-labeled mouse Hst-1/Fgf-4 cDNA precursors and induces the dierentiation of Bergmann
fragment as a probe. Production of the control b-actin products glia. Blocking the SHH function in vivo results in
was observed throughout the lanes with mRNAs, N indicates no de®cient granule neuron and Bergmann glia dierentia-
RNA samples; P indicates mRNA from L361-M3E cells as a tion as well as in anomalous Purkinje neuron
template
development. Furthermore, hedgehog genes have been
implicated in inductive signaling during vertebrate limb
formation (Niswander et al., 1994). FGF-4, as a
On sections of adult testis, a signal was found secondary signal, is secreted in an apical ectodermal
localized to Sertoli cell fractions (Figure 3d) which ridge in response to SHH (Yang and Niswander, 1995).
were positively stained with anti-vimentin antibody Thus, our ®ndings that the Hst-1/Fgf-4 gene is
(Figure 3f). Weak labeling was also detected in adult expressed in Purkinje cells may provide a molecular
Leydig cells and pachytene spermatocytes of stage V ± model for the growth and patterning of the cerebellum
XI tubules (data not shown). The sense RNA probe by SHH through the coordination of the development
did not label any cells, and no nonspeci®c labeling was of cortical cerebellar cell types.
observed (Figure 3e). The intensity of the Hst-1/Fgf-4 Localization of the expression of other members of
signal in Sertoli cells was generally uniform in all the FGF family in the brain has been well recognized
tubules (data not shown), indicating that its expression in acidic FGF, basic FGF (FGF-2), FGF-5 and FGF-
was not associated with a speci®c stage of the 9. Acidic FGF expression is restricted to neurons (Bean
spermatogenic cycle. et al., 1991; Wilcox and Umerstall, 1991). FGF-2-like
immunoreactivity was noted not only throughout the
Hst-1/Fgf-4 transcripts in the primary cultured cells cytoplasm of Purkinje cells but also in the nuclei of the
immunopositive cells from postnatal day 14 to
To con®rm further the cell type-speci®c expression of postnatal day 28 in rat cerebellum (Matsuda et al.,
Hst-1/Fgf-4 gene, Purkinje cells and Sertoli cells were 1992, 1994). FGF-5 has been demonstrated to be
puri®ed from adult mice, respectively, and cultured in localized in neurons in the rat brain (Gomez-Pinilla et
vitro. Immunohistochemical analysis of the primary al., 1993). FGF-9 immunoreactivity was also noted in
OncogeneHst-1/Fgf-4 expression in the adult mice
H Yamamoto et al
3808
Figure 3 Expression of Hst-1/Fgf-4 in the adult mice tissues. In situ hybridization of antisense or sense Hst-1/Fgf-4 RNA probe to
sections of cerebellum (a ± b) and testis (d ± f) from 10-week old mice. (a) Hst-1/Fgf-4 antisense probe signal on section of the
cerebellum. (b) Signal from the control sense probe. (c) Immunohistochemical staining with anti-a-calbindin antibody and
counterstained with hematoxylin. (d) Hst-1/Fgf-4 antisense probe signal on section of testis. (e) Signal from the control sense probe.
(f) Immunohistochemical staining with anti-vimentin antibody and counterstained with hematoxylin. ml, molecular layer; pcl,
Purkinje cell layer; gcl, granule cell layer
1/FGF-4, were expressed in the rat brain during
postnatal development (el-Husseini et al., 1994) and
developing chicken nervous system (Heuer et al., 1990).
These reports suggest that the FGF family and their
receptors play an important role in the CNS. Our
®ndings that the HST-1/FGF-4 gene is also expressed
in adult CNS indicate the requirement of HST-1/FGF-
4 in maintenance of some of adult brain functions. In
this connection, it may be of interest to study
expression pro®les of HST-1/FGF-4 in the weaver
and reeler mutant cerebella, which show abnormal
cytoarchitecture and neural circuitry.
Activity-dependent long-term modi®cation of trans-
mission eciency at synapses is thought to be a cellular
basis of learning and memory (McGaugh, 2000). Since
the discovery of long-term potentiation in the
hippocampus, synaptic plasticity has been described
in various parts of the brain including cerebellar
Purkinje cells (Khodakhah and Armstrong, 1997).
However, little is known about what type of cytokines
modulates the maintenance of Purkinje cell synaptic
Figure 4 Hst-1/Fgf-4 transcripts in the primary cultured cells. plasticity. Our ®ndings strengthen the need to further
Expression of Hst-1/Fgf-4 mRNAs in the cultured Purkinje cells study the possible regulatory role of HST-1/FGF-4 in
(a) and Sertoli cells (b) from 10-week-old ICR mice was synaptic plasticity of Purkinje cells.
determined by RT ± PCR analysis. DNAs were denatured and
transferred onto a nylon membrane, and then hybridized with a
The whole process of dierentiation of male germ cells
a32-labeled mouse Hst-1/Fgf-4 cDNA fragment as a probe. depends on a complex network of endocrine and
Production of the control b-actin products was observed paracrine communication involving a variety of support-
throughout the lanes with mRNAs. N indicates no RNA samples; ing cells. In the seminiferous tubule, spermatogenic cells
P indicates mRNA from L361-M3E cells as a template are in continuous contact with Sertoli cells; this means
that Sertoli cells play a critical role in germinal
dierentiation (Griswold et al., 1988). Among the FGF
Purkinje cells in adult and developing CNS in human gene family, FGF-2 was found to signi®cantly increase
and rats (Todo et al., 1998; Nakamura et al., 1999). the number of Sertoli cells and also stimulated the
FGF-19, a novel member of the human FGF family, proliferative activity of the gonocytes in vitro (Van
was expressed in fetal brain (Nishimura et al., 1999). In Dissel-Emiliani et al., 1996). The presence of continuous
the chick system, a model for FGF function in expression of Hst-1/Fgf-4 gene in Sertoli cells was a
speci®cation and patterning of the midbrain was novel ®nding, and this expression is likely to be involved
proposed (Shamin et al., 1999). Regarding FGF in speci®c signals during sperm maturation because the
receptors (FGFRs), the FGFR1 and FGFR2 genes, mature spermatogenic cycle is thought to be coordinated
which were high anity receptors for FGF-2 and HST/ in part by an exchange of signals, possibly by Sertoli
OncogeneHst-1/Fgf-4 expression in the adult mice
H Yamamoto et al
3809
cells, between germ cells at dierent phases of matura- 0.5% sodium dodecyl sulfate (SDS), 56sodium saline
tion. In fact, the FGFRs 1 and 4 are known to be phosphate EDTA (16sodium saline phosphate ED-
localized in Sertoli cells in rat testis (Le Magueresse- TA=150 mM NaCl, 10 mM sodium phosphate, 1.3 mM
Battistoni et al., 1994; Cancilla and Risbridger, 1998). EDTA), and 200 mg/ml of salmon sperm DNA at 658C for
16 h. The ®lters were then washed in 0.1% SDS and
Taken together, these data suggest that FGF is an
0.16sodium saline phosphate EDTA at 658C. The images
important factor for the spermatogenesis. Whether the were incorporated into Macintosh computer through a
Sertoli cell line established from transgenic mice STORM image scanner (Molecular Dynamics). Intensity of
expressing the human Hst-1/Fgf-4 gene (Ochiya et al., each band on the image was measured and the data were
unpublished data) showed enhanced ability to support processed by an image processing software ImageQuaNT.
sperm maturation awaits further analysis.
In conclusion, as information about the FGF gene Primary cultured cells
family expression in the adult animals accumulates, it
becomes apparent that the Hst-1/Fgf-4 gene may have Two 10-week-old mice were sacri®ced and their cerebellums
some function in cells once they undergo terminal were removed without their cortexes. They were then rinsed
with phosphate-buered saline (PBS) and cut into pieces with
dierentiation, as well as in developmental pathways.
scissors in 3 ml of digested solution (1% trypsin/0.05%
This is the ®rst demonstration that the Hst-1/Fgf-4 collagenase/70 U/ml of DNase I in PBS) and were incubated
gene is expressed in the brain and testis at adult stages, for 10 min in 378C of water bath with vigorous shaking. They
and will be an important insight into unexplored FGF were suspended into 10 ml of culture medium [50% DMEM/
functions. 50% HAM-F12/10% FBS/10 mg/ml insulin-0.55 mg/ml
transferrin-6.7 ng/ml sodium selenium (Life Technologies
Inc., MD, USA)/100 U/ml Penicillin G, 100 U/ml Streptomy-
Materials and methods cin sulfate, 0.25 mg/ml AmphotericinB (Life Technologies
Inc., MD, USA)] to stop the reaction, and the suspension was
Animals and cells passed through a nylon mesh and double sheets of lens paper.
The cell suspension was centrifuged at 1400 r.p.m. for 5 min.
Male and female ICR mice (7-, 12- and 38-week-old), The cells were plated on two poly-D-lysine/laminin-coated
pregnant mice, and C57BL6J mice (66-week-old) were dishes (Becton Dickinson) and cultured in 5% CO2 at 378C for
purchased from Charles River Japan (Yokohama, Japan). 6 h. Afterwards, only adherent cells were collected by a
L361-M3E cells, which were transformed NIH3T3 cells by scraper and their mRNA was prepared by Micro-FastTrack
introducing genomic mouse Hst-1/Fgf-4 genes, were cultured 2.0 mRNA isolation kit (Invitrogen, The Netherlands). The
for positive control of murine Hst-1/Fgf-4 gene expressing adherent cells were con®rmed to be Purkinje cells by staining
cells. with Purkinje cell speci®c marker (Vandaele et al., 1991): anti-
Calbindin-D 28k antibody (Sigma).
RT ± PCR analysis Sertoli cells from 10-week-old animals were isolated
following the method of Orth and Boehm (1990) and
Adult mice (5-, 10-, 38- and 66-week-old), embryos cultured with DMEM supplemented with 10% fetal bovine
(embryonic days (E) 14.5 and 18.5) and neonates (postnatal serum. Sertoli cell cultures were also ®xed in buered
days (P) 0, 6 and 10) were sacri®ced and their organs were formaldehyde and processed for immunohistochemistry with
dissected then prepared separately. Embryonic cerebellum goat-anti-vimentin antibody (Cortex Biochem, USA) to
and testis were pooled from 10 ± 18 embryos. Total RNAs demonstrate the purity of Sertoli cells in culture (Paranko
were prepared using Isogen solution (Nippon Gene, Tokyo) et al., 1986; Guillou et al., 1990). Total RNAs from the
according to the manufacturer's protocol. Two hundred cultured Purkinje and Sertoli cells were extracted and then
nanograms of total RNA isolated from each organ of adult subjected to RT ± PCR analysis, respectively. The PCR
mice were treated with DNase (DNase I, ampli®cation grade; primers used for ampli®cation of IP3R1 (Fujino et al.,
Life Technologies Inc., MD). RT ± PCR reaction was 1995) were 5'-ATTAGTGCCCCTGTATCCTA-3' and 5'-
performed using Superscript One Step RT ± PCR system GGAGATGACACTGACTGGT-3', which produce a
(Life Technologies Inc., MD, USA) and Platinum Taq 254 bp product. b-actin cDNA was ampli®ed as an internal
antibody (Life Technologies Inc., MD, USA) with primers control using primers 5'-GACATCAAAGAGAAGCTGT-
5'-TGGTGTGACCGCAGACACGA-3' and 5'-GGTAAA- GC-3' and 5'-TAGGAGCCAGAGCAGTAATC-3'.
GAAAGGCACACCGA-3'. After 30 min at 558C for reverse
transcription reaction of 1 mg of total RNA, the reaction was
terminated at 948C for 2 min, then PCR was performed with Preparation of riboprobes
35 cycles in a Perkin-Elmer Thermal Cycler, using a cycle Dioxygenin-labeled sense or antisense riboprobes were
pro®le of: 1 min at 978C, 1 min at 608C and 40 s at 728C. synthesized by T3 or T7 RNA polymerase from cDNA
The ampli®ed DNA was recovered with a silica-gel plasmids containing entire coding sequences of mouse Hst-1/
membrane ®lter, then cloned in TA cloning vector (pGEM- Fgf-4 cDNA in the presence of digoxygenin-dUTP (Boehrin-
T Easy Vector Systems; Promega, WI, USA). The sequence ger-Mannheim).
analysis was performed using a dye deoxy primer method
(Thermo Sequenase; Amersham, Buckinghamshire, UK).
In situ hybridization
In order to evaluate the gene expression level, the cloned
desired sequences (16101 ± 46106 copies with 10 fold serial In situ hybridization experiments using fresh-frozen tissue
increment per each reaction) were used as quantitative sections and dioxygenin-labeled riboprobes were carried out
controls. After RT ± PCR, aliquots were run on 3.5% agarose essentially as described previously (Wilkinson, 1992; Ishii et
gels, stained with ethidium bromide, and then photographed al., 1997). Adult organs and embryos were ®xed with 4%
under UV illumination. DNAs were denatured with 0.5 N parformaldehyde in phosphate buered saline (PBS) for
NaOH-1.5 M NaCl and then transferred onto a nylon 10 min at 48C for 12 h, then dehydrated in 30% sucrose in
membrane (Hybond N plus, Amersham), and hybridized PBS at 48C for 4 h with gentle agitation and embedded in
with a probe; a a32P-labeled mouse Fgf-4 cDNA fragment O.C.T. and sectioned at a thickness of 5 ± 10 mm using a
in 56Denhardt's solution (16Denhardt's=0.02% Ficoll, cryostat (Sakura Coldtome, Japan). The sections were
0.02% bovine serum albumin, 0.02% polyvinylpyrrolidone), immediately dried at 458C for 3 h. Before hybridization,
OncogeneHst-1/Fgf-4 expression in the adult mice
H Yamamoto et al
3810
slides were soaked in 0.2 N HCl for 20 min to inactivate testis and cerebellum were required until signi®cant signals
endogenous alkaline-phosphatase, rinsed with deionized appeared. As references, standard immunostaining and
water, treated with proteinase K (1 mg/ml in PBS, Boehringer hematoxylin and eosin staining were performed in serial
Mannheim) for 7 ± 15 min at 378C, hydrated with ethanol, sections.
then air dried. An appropriate amount of the probe in 50 ml
of hybridization buer consisting of 300 mM NaCl, 30 mM
sodium citrate (pH 7.0), 50% v/v deionized-formamide, 1%
w/v sodium dodecyl sulfate, 50 mg/ml heparin and 50 mg/ml
yeast RNA was applied to each slide, and hybridization was Acknowledgments
carried out at 658C for 12 ± 16 h. After hybridization, slides We gratefully acknowledge Drs Katsuhiko Mikoshiba
were washed with 300 mM NaCl, 30 mM sodium citrate (pH (Department of Molecular Neurobiology, Institute of
7.0), 50% deionized-formamide at 52 ± 558C for 1 h. Detec- Medical Science, University of Tokyo, Japan) and Teiichi
tion was carried out by anti-digoxygenin antibody conjugated Furuichi (Brain Science Institute, The Institute of Physical
with alkaline-phosphatase (1 : 500, Boehringer Mannheim). and Chemical Research, Saitama, Japan) for their kind
Colormetric reaction with nitro-blue tetrazolium and 5- assistance. We thank Ms Maki Abe for her technical work.
bromo-4-chloro-3-indolyl-1-phosphate (NBT/BCIP) was car- This work was supported in part by a Grant-in-Aid for the
ried out at room temperature: 12 h for control samples such Second-Term Comprehensive 10-Year Strategy for Cancer
as E9.5 limb and myotome and 96 h for all the samples from Control from the Ministry of Health and Welfare in Japan.
References
Bean AJ, Elde R, Cao YH, Oellig C, Tamminga C, Goldstein Niswander L and Martin GR. (1992). Development, 114,
M, Pettersson RF and Hokfelt T. (1991). Proc. Natl. Acad. 755 ± 768.
Sci. USA, 88, 10237 ± 10241. Niswander L and Martin GR. (1993). Nature, 361, 68 ± 71.
Cancilla B and Risbridger GP. (1998). Biol. Reprod., 58, Niswander L, Jerey S, Martin GR and Tickle C. (1994).
1138 ± 1145. Nature, 371, 609 ± 612.
Collignon J, Sockanathan S, Hacker A, Cohen-Tannoudji Ochiya T, Sakamoto H, Tsukamoto M, Sugimura T and
M, Norris D, Rastan S, Stevanovic M, Goodfellow PN Terada M. (1995). J. Cell Biol., 130, 997 ± 1003.
and Lovell-Badge R. (1996). Development, 122, 509 ± 520. Orth JM and Boehm R. (1990). Anat. Rec., 226, 320 ± 327.
Dahmane N and Altaba Ari. (1999). Development, 126, Ozawa K, Uruno T, Miyakawa K, Seo M and Imamura T.
3089 ± 3100. (1996). Mol. Brain Res., 41, 279 ± 288.
Drucker BJ and Goldfarb M. (1993). Mech. Dev., 40, 155 ± Paranko J Kajjajoki M, Pelliniemi LJ, Lehto V-P and
163. Virtanen I. (1986). Dev. Biol., 117, 35 ± 44.
el-Husseini A el-D, Paterson JA, Shiu RP. (1994). Mol. Cell Sakamoto H, Ochiya T, Sato Y, Tsukamoto M, Konishi H,
Endocrinol., 104, 191 ± 200. Saito I, Sugimura T and Terada M. (1994). Proc. Natl.
Fujino I, Yamada N, Miyakawa A, Hasegawa M, Furuichi T Acad. Sci. USA, 91, 12368 ± 12372.
and Mikoshiba K. (1995). Cell Tissue Res., 280, 201 ± 210. Shamin H, Mahmood R, Logan C, Doherty P, Lumsden A
Gomez-Pinilla F and Cotman CW. (1993). Brain, 606, 79 ± and Mason I. (1999). Development, 126, 945 ± 959.
86. Suzuki HR, Sakamoto H, Yoshida T, Sugimura T, Terada M
Griswold M, Morales C and Sylvester S. (1988). Oxford Rev. and Solursh M. (1992). Dev. Biol., 150, 219 ± 222.
Reprod. Biol., 10, 124 ± 161. Theillet C, Le Roy X, De Lapeyriere O, Grosgeorges J,
Guillou F, Monet-Kuntz C, Fountain I and Flechon JE. Adnane J, Raynaud SD, Simony-Lafontaine J, Goldfarb
(1990). Cell Tissue Res., 260, 395 ± 401. M, Escot C, Birnbaum D and Gaudray P. (1989).
Hannon K, Kudla AJ, McAvoy MJ, Clase KL and Olwin Oncogene, 4, 915 ± 922.
BB. (1996). J. Cell Biol., 132, 1151 ± 1159. Todo T, Kondo T, Nakamura S, Kirino T, Kurokawa T and
Heikinheimo O, Dong KW, Lanzendorf SE, Gordon K, Ikeda K. (1998). Brain Res., 783, 179 ± 187.
Toner JP and Gibbons WE. (1995). Mol. Reprod. Dev., 42, Tsuda H, Hirohashi S, Shimosato Y, Hirota T, Tsugane S,
397 ± 406. Yamamoto H, Miyajima N, Toyoshima K, Yamamoto T,
Heuer JG, von Bartheld CS, Kinoshita Y, Evers PC and Yokota J, Yopshida T, Sakamoto H and Terada M.
Bothwell M. (1990). Neuron, 5, 283 ± 296. (1989). Cancer Res., 49, 3104 ± 3108.
Ishii Y, Fukuda K, Saiga H, Matsushita S and Yasugi S. Yang Y and Niswander L. (1995). Cell, 24, 939 ± 947.
(1997). Dev. Growth Dier., 39, 643 ± 653. Yoshida T, Tsutsumi M, Sakamoto H, Miyagawa K,
Khodakhah K and Armstrong CM. (1997). Proc. Natl. Acad. Teshima S, Sugimura T and Terada M. (1988). Biochen.
Sci. USA, 94, 14009 ± 14014. Biophys. Res. Commun., 155, 1324 ± 1329.
Konishi H, Ochiya T, Sakamoto H, Tsukamoto M, Saito I, Yoshida T, Miyagawa K, Sakamoto H, Sugimura T and
Muto T, Sugimura T and Terada M. (1995). J. Clin. Terada M. (1991). Methods Enzymol., 198, 124 ± 138.
Invest., 95, 1125 ± 1130. Yoshida T, Ishimaru K, Sakamoto H, Yokota J, Hirohashi
Konishi H, Ochiya T, Yasuda Y, Sakamoto H, Muto T, S, Igarashi K, Sudo K and Terada M. (1994). Cancer Let.,
Sugimura T and Terada M. (1996). Oncogene, 13, 9 ± 19. 83, 261 ± 268.
Le Magueresse-Battistoni B, Wol J, Morera AM and Yuan H, Corbi N, Basilico C and Dailey L. (1995). Gene
Benahmed M. (1994). Endocrinology, 135, 2404 ± 2411. Dev., 9, 2635 ± 2645.
Matsuda S, Okumura N, Yoshimura H, Koyama Y and Vandaele S, Nordquist DT, Feddersen RM, Tretjako I,
Sakanaka M. (1992). Neuroscience, 50, 99 ± 106. Peterson AC and Orr HT. (1991). Gene Dev., 5, 1136 ±
Matsuda S, Ii Y, Desaki J, Yoshimura H, Okumura N and 1148.
Sakanaka M. (1994). Neuroscience, 59, 651 ± 662. Van Dissel-Emiliani FM, De Boer-Brouwer M and De Rooij
McGaugh JL. (2000). Science, 287, 248 ± 251. DG. (1996). Endocrinology, 137, 647 ± 654.
Nakamura S, Todo T, Motoi Y, Haga S, Aizawa T, Ueki A Wilcox BJ and Unnerstall JR. (1991). Neuron, 6, 397 ± 409.
and Ikeda K. (1999). Glia, 28, 53 ± 65. Wilkinson DG. (1992). In situ hybridization: A practical
Nishimura T, Utsunomiya Y, Hoshikawa M, Ohuchi H and approach. IRL Press: London.
Itoh N. (1999). Biochim. Biophys. Acta, 18, 148 ± 151.
OncogeneYou can also read

















































