EFFECT OF NATALIZUMAB TREATMENT ON METALLOPROTEINASES AND THEIR INHIBITORS IN A MOUSE MODEL OF MULTIPLE SCLEROSIS
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JOURNAL OF PHYSIOLOGY AND PHARMACOLOGY 2020, 71, 2, 265-273
www.jpp.krakow.pl | DOI: 10.26402/jpp.2020.2.11
G. PYKA-FOSCIAK, G.J. LIS, J.A. LITWIN
EFFECT OF NATALIZUMAB TREATMENT ON METALLOPROTEINASES
AND THEIR INHIBITORS IN A MOUSE MODEL OF MULTIPLE SCLEROSIS
Department of Histology, Faculty of Medicine Jagiellonian University Medical College, Cracow, Poland
Matrix metalloproteinases (MMPs) regulated by their tissue inhibitors (TIMPs) play a significant role in the
pathogenesis of multiple sclerosis (MS) and its mouse model, experimental autoimmune encephalomyelitis (EAE), as
they degrade extracellular matrix including vascular basal laminae and by damaging blood-brain barrier (BBB) facilitate
transmigration of immune cells into the central nervous system. MMPs are also involved in destruction of myelin
sheaths, leading to axonal and neuronal loss. The aim of the present study was to assess whether natalizumab, a
transmigration-inhibiting monoclonal antibody against a4b1 integrin, influences expression of MMPs and TIMPs in the
central nervous system of mice with EAE. MMP-2 and MMP-9, their respective inhibitors TIMP-2 and TIMP-1 and
laminin were assessed by quantitative immunohistochemistry in the spinal cord cryosections of C57BL/6 mice with
EAE in the successive phases of the disease (onset, peak and chronic). The percentage of immunopositive areas were
calculated in sections encompassing the whole spinal cord cross-sectional area occupied by the gray and white matter.
Results obtained in animals administered with 5 mg/kg natalizumab were compared with those collected from control
mice receiving 5 mg/kg IgG. Both studied MMPs and both TIMPs were upregulated in control EAE mice. Natalizumab
treatment significantly reduced expression of MMPs and increased expression of TIMPs in the peak and chronic phases
of the disease. This effect was accompanied by inhibition of laminin degradation in the vascular basal laminae and
reduction of inflammatory infiltration. Results of this study demonstrate that in addition to its well known anti-integrin
activity counteracting transmigration of immune cells into the central nervous system, natalizumab strengthens this
effect by its probably indirect influence on MMPs and TIMPs leading to protection of blood-brain barrier integrity.
K e y w o r d s : experimental autoimmune encephalomyelitis, blood-brain barrier, natalizumab, matrix, metalloproteinases, tissue
metalloproteinases, inhibitors, laminin
INTRODUCTION of endothelial cells induce expression of MMP-2 which in turn
facilitates T cell migration into the perivascular tissue (8).
Experimental autoimmune encephalomyelitis (EAE) is a Natalizumab, a monoclonal antibody against a-chain of
well known mouse model of multiple sclerosis (MS) (1). The a4b1 integrin (very large antigen-4, VLA-4) has potentially
factors responsible for the development of MS include an beneficial therapeutic effects in MS and Crohn disease, since it
increase in matrix metalloproteinase (MMP) activity effectively inhibits the entry of pathogenic T cells and
accompanied by a decrease in expression of tissue inhibitors of monocytes into the central nervous system and their attack on
MMPs (TIMPs) (2, 3). MMPs disrupt the blood-brain barrier myelin sheaths. Anti-VLA-4 mAb binds to the a4 subunit of
(BBB), leading to increased leukocyte migration into the CNS a4b1 integrin on the surface of lymphocytes, blocking the
and are involved in degradation of myelin proteins (4). binding of the integrin to their endothelial receptors in the brain
Additionally, infiltration of leukocytes is associated with and spinal cord blood vessels (9). Anti-VLA-4 mAb can also
abnormalities of the vascular basement membranes, which can block cellular migration and/or attenuate accumulation of
contribute to BBB disruption. Laminins, components of infiltrating cells by inhibiting the binding of a4-positive
basement membranes and substrates for MMPs, influence leukocytes with fibronectin and osteopontin (10).
infiltration of leukocytes in EAE and correlate with disease Anti-VLA-4 mAb efficacy has been demonstrated in
severity (5, 6). inhibiting the severity of EAE when it was administered during
Studies on the mechanism of leukocyte infiltration across the disease progression (9). However, discordant effects of anti-
BBB revealed that a4b1 integrin (VLA-4) is a critical cell VLA-4 mAb treatment in EAE have also been reported:
adhesion molecule that mediates Th-1 cell migration in the preclinical administration attenuated the onset and severity of
pathogenesis of EAE, and has been a target of clinical trials in the disease but administration either at the peak of acute disease
MS (7). VLA-4 influences migration of immune cells by or during remission exacerbated disease relapses (11).
interaction with its ligand, vascular cell adhesion molecule Since MMPs play a significant role in the pathogenesis of
(VCAM-1). VLA-4 expressing T cells after binding to VCAM-1 MS/EAE and some authors reported altered plasma levels of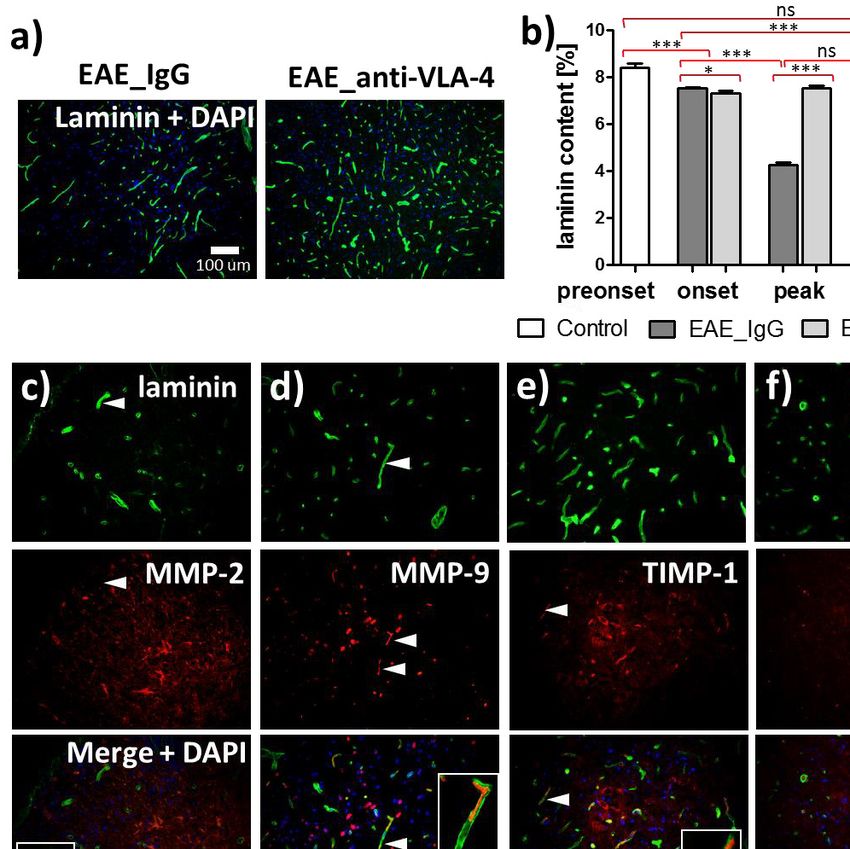
266
MMPs in natalizumab treated MS patients (12), while others did group consisted of mice injected with CFA and PTx only
not find any MMP changes in such patients (13), the aim of the (Control; n = 10).
present study was to investigate by quantitative In EAE mice injected with IgG no symptoms of EAE were
immunohistochemistry the effect of anti-VLA-4 mAb treatment on observed between post immunization days 0 – 8 days (preonset
the expression of MMPs and their inhibitors (TIMPs) in the phase of EAE). Initial symptoms of the disease appeared
successive phases of EAE. between days 9 and 14 (onset phase). The maximum scores
(peak phase) occurred between days 15 and 20 and then the mice
partially recovered (chronic phase). In EAE mice treated with
MATERIALS AND METHODS anti-VLA-4 mAb, the onset phase began later (day 11) and the
peak phase was shorter (days 15 to 18).
Animals and treatment For histological analysis, mice treated for EAE (anti-VLA-4
and IgG groups) were sacrificed at three different time points
Naive, C57BL/6 mice (female, 10 – 11 weeks old, weight 19 representing terminal days of three disease phases: onset phase
– 24 g, obtained from the Center for Experimental Medicine of (day 13; n = 10), peak phase (day 18; n = 10) and chronic phase
Bialystok Medical University, Poland) were housed under 12/12h (day 28; n = 10) (Fig. 1). Control mice were sacrificed on days
light/dark cycles in temperature-controlled environment (22 ± 6 and 28 (n = 10).
2°C, 55 ± 10% humidity) and specific pathogen-free conditions
(Animal House of the Jagiellonian Center for Experimental Spinal cord collection and processing
Therapeutics, JCET, Cracow) with free access to water and food.
The mice were habituated for 8 days before the immunization. Mice received i.p. 100 mg/kg ketamine and 10 mg/kg
All experiments were conducted in compliance with the xylazine and were transcardially perfused with ice-cold PBS for
Council Directive 2010/63EU of the European Parliament and 10 min, followed by 4% paraformaldehyde for the next 10 min.
the Council of 22 September 2010 on the protection of animals Spinal cords were carefully removed from the vertebral canal
used for scientific purposes and were approved by the Local and postfixed in the same fixative for 4 hours. After overnight
Ethics Committee of the Jagiellonian University Medical incubation in 5% sucrose at 4°C, tissue was embedded in OCT
College, Cracow, Poland. (Shandon Cryomatrix, Thermo Fisher Scientific, Rockford, IL
USA) and snap-frozen at –80°C. The examined area of the spinal
Induction and evaluation of experimental autoimmune cord included the lumbar part, a region commonly and rapidly
encephalomyelitis affected in EAE. Serial cryosections, 10 µm thick, were cut at
100 µm intervals, collected on poly-L-lysine coated slides and
On day 0, each mouse (n = 30) received subcutaneous air dried. The sections were fixed with acetone (for
injections of immunizing Hooke Kits™ EAE Emulsion (Hooke immunohistochemistry), with 4% buffered formaldehyde (for
Laboratories, Lawrence, Massachusetts USA) at two sites (100 HE staining) or left unfixed (for zymography).
µl between the ears and 100 µl in hind flank). The immunizing
kits contained MOG35-55 antigen in an emulsion with Complete Histology, immunohistochemistry and zymography
Freund’s Adjuvant (CFA) including Mycobacterium tuberculosis
(H37Ra). On day 0 and 1, mice were also injected Hematoxylin and eosin (H&E) staining method was applied
intraperitoneally (i.p.) with 340 µl of Bordetella pertussis to analyze degree of inflammatory infiltration.
pertussis toxin (PTx) dissolved in phosphate-buffered saline Metalloproteinases, their inhibitors and laminin were detected
(PBS) (Hooke Laboratories). Control mice (n = 10) were in sections by immunofluorescence. The following primary
injected with CFA and PTx only (Hooke Control Kits™, Hooke antibodies were used: rabbit anti-laminin (1:500, Thermo Fisher
Laboratories, USA) according to the same schedule. Scientific, Rockford, IL USA, cat. # PA1-16730) or rat anti-
After appearance of the first clinical signs of EAE, mice laminin (1:200, Thermo Fisher Scientific, Rockford, IL USA,
were injected i.p. with 5 mg/kg of anti-VLA-4 mAb cat. # MA5-14649), rabbit anti-MMP-2 (Abcam, Cambridge,
(Natalizumab, Biogen Idec, Berkshire, UK) or, in case of EAE UK; 1:500; cat. # ab37150), rabbit anti-MMP-9 (Abcam,
control, with 5 mg/kg of IgG (Sigma-Aldrich, St. Louis, MO, Cambridge, UK; 1:500; cat. # ab38898), rat anti-TIMP1
USA). The injections were continued until the appearance of the (Abcam, Cambridge, UK; 1:200; cat. # ab86482) and goat-anti
first remission symptoms (days 9, 12, 15, 18, 21) (11, 14). TIMP2 (R&D Systems, Minneapolis, USA.; 1:200; cat #
Animals were examined on a daily basis (from day 0 to day AF971). The secondary antibodies included goat anti-rat
28) in search of clinical symptoms of EAE. Neurological Alexa488-conjugated antibodies (1:100, Jackson IR, West
symptoms of EAE were assessed according to the score table Grove, PA, cat. # 112-545-167), donkey anti-goat Alexa594-
provided by the protocols included in the Hooke Kits™ EAE conjugated antibodies (1:100, Jackson IR, West Grove, PA, cat.
Emulsion (Hooke Laboratories) for each mice separately. Disease # 112-545-167) and Cy3-conjugated goat anti-rabbit antiserum
severity was evaluated using a scale ranging from 0 to 3: (Jackson IR, West Grove, PA; 1:500; cat. # 111-225-144).
0 - no clinical disease, The spinal cord sections were preincubated for 40 min in a
0.5 - tip of tail is limp; blocking solution: PBS containing 5% normal goat serum
1 - limp tail; (Sigma-Aldrich, St. Louis, MO, USA), 0.01% sodium azide,
1.5 - limp tail and hind leg inhibition; 0.05% thimerosal, 0.1% bovine serum albumin, 0.5% Triton X-
2 - limp tail and weakness of hind legs; 100, and 2% dry milk. They were next incubated overnight at
2.5 - limp tail and dragging of hind legs; room temperature with primary antibodies and after a rinse in
3 - limp tail and complete paralysis of hind legs. PBS incubated for 90 min with the secondary antibodies. Then,
sections were washed three times in PBS and mounted in
Experimental groups glycerol/PBS solution.
To detect the gelatinase activity of matrix metalloproteinases
All immunized mice (n = 30) were divided into two main (mainly MMP-2 and MMP-9), in situ gelatin zymography was
groups: group injected with anti-VLA-4 mAb (EAE_anti-VLA- employed: unfixed cryosections were incubated for 2 hours at
4; n = 15) or injected with IgG (EAE_IgG; n = 15). The control 37°C in a dark humid chamber with 50 mg/ml FITC-labeled DQ-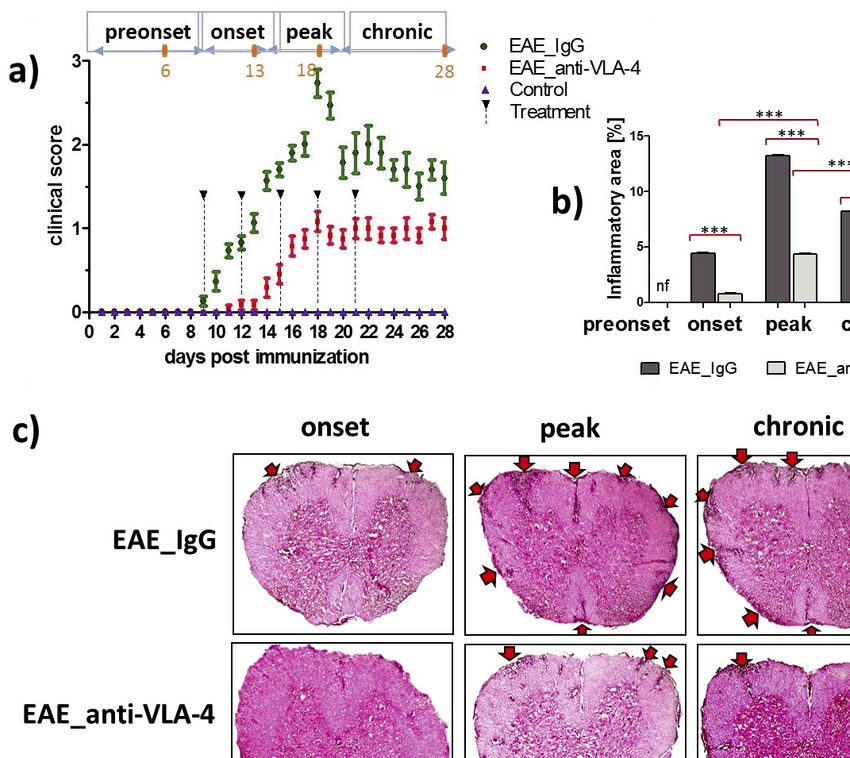
267
Fig. 1. Progression of experimental autoimmune encephalomyelitis (EAE) in mice and effect of anti-VLA-4 and IgG (control)
treatment on neurological symptoms (a) and on inflammation degree in the spinal cord (b, c). The orange points and numbers in disease
phases (a, upper part) mark days of tissue collection. Hematoxylin-eosin stained spinal cord sections (c) show inflammatory
infiltrations (arrows). Data in (a) and (b) are presented as means ± SEM; n = 5 per group. ***P < 0.001; **P < 0.01; *P < 0.05. All
micrographs are at the same magnification.
gelatin (Invitrogen, Eugene, OR). After a rinse in PBS, sections analyses. All values were expressed as mean ± standard error of
were mounted in glycerol/PBS. the mean (S.E.M.). Statistical significance of the obtained results
was verified using two-sided Mann-Whitney test, at the
Microscopy and morphometry confidence level of 0.05 (***P < 0.001; **P < 0.01; *P < 0.05;
ns - not significant).
The spinal cord sections were examined under Olympus
BX50 brightfield/epifluorescence microscope (Olympus, Tokyo,
Japan). Images were recorded with the use of Olympus DP71 RESULTS
digital CCD camera, stored as TIFF files and processed for
quantitative analysis using ImageJ software (NIH, Bestheda, Model of experimental autoimmune encephalomyelitis
Maryland, USA). induction
A total of at least 25 slides were analyzed per experimental
group (n = 5). The percentage of immunopositive areas were All immunized mice developed neurological symptoms of
assessed in sections encompassing the whole spinal cord cross- EAE. Control mice did not show any pathological manifestations.
sectional area occupied by the gray and white matter. Treatment with anti-VLA-4 mAb effectively inhibited clinical
symptoms of EAE compared to IgG-treated group. The four phases
Statistical analysis of EAE were characterized by the following mean clinical scores:
preonset: 0; onset: 0.08 ± 0.1 for anti-VLA-4 and 1.1 ± 0.1 for IgG
GrandPad Prism 5.0 software (GraphPad, La Jolla, group; peak: 1.1 ± 0.1 for anti-VLA-4 and 2.7 ± 0.2 for IgG group;
California, USA) was used throughout this study for statistical chronic:1.0 ± 0.1 for anti-VLA-4 and 1.6 ± 0.2 for IgG group. After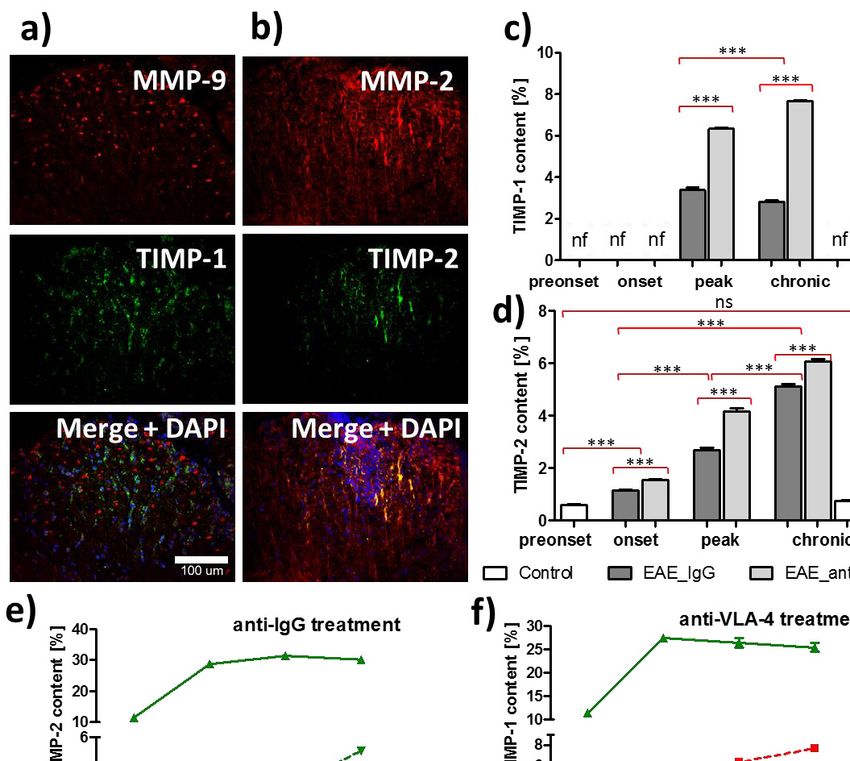
268
termination of the treatment, the disease severity manifested by indicating that inflammation did not fully retreat. Samples
clinical scores did not change in both groups of EAE mice, with obtained from control mice were free of any inflammatory
significantly lower scores observed in anti-VLA-4 group compared infiltrates (Fig. 1b).
to IgG treated mice (Fig. 1a).
Effect of anti-VLA-4 mAb treatment on gelatinase activity
Anti-VLA-4 mAb treatment reduces inflammation
Although both MMP-2 and MMP-9 can be responsible for
In order to determine whether anti-VLA-4 mAb treatment gelatinase activity, double labeling demonstrated that it was
influenced recruitment of inflammatory cells and their ability to mainly related to the MMP-2 (Fig. 2a and 2c), as MMP-9 showed
break the BBB, inflammation degree was quantified using a much weaker expression (Fig. 2b and 2d). I n EAE mice treated
morphometric approach to calculate the surface area of with IgG, gelatinase activity increased in the onset phase from 9.3
inflammatory infiltrations. Hematoxylin-eosin (H&E) stained ± 0.4% to 11.4 ± 0.4%, reached maximum (22 ± 0.5%) in the
spinal cord sections showed that the surface area of EAE lesions peak phase and decreased to 15.3 ± 0.5% in the chronic phase.
corresponded to severity of clinical manifestations (Fig. 1b and Anti-VLA-4 mAb treatment significantly inhibited gelatinase
1c). In IgG treated group, the largest inflammatory area was activity in the peak and chronic phases of EAE (Fig. 2e).
observed in the peak phase (13.3 ± 0.1%) and it decreased in the
chronic phase (8.2 ± 0.1%). Anti-VLA-4 mAb treatment Anti-VLA-4 mAb treatment lowers matrix metalloproteinase
significantly reduced the inflammatory area (4.4 ± 0.1% in the expression
peak phase and 3.8 ± 0.1% in the chronic phase). In both groups,
the size of inflammatory area in the chronic phase was During progression of EAE, MMP-2 expression rapidly
intermediate between those found in onset and peak phases, increased in the onset phase and remained relatively stable in the
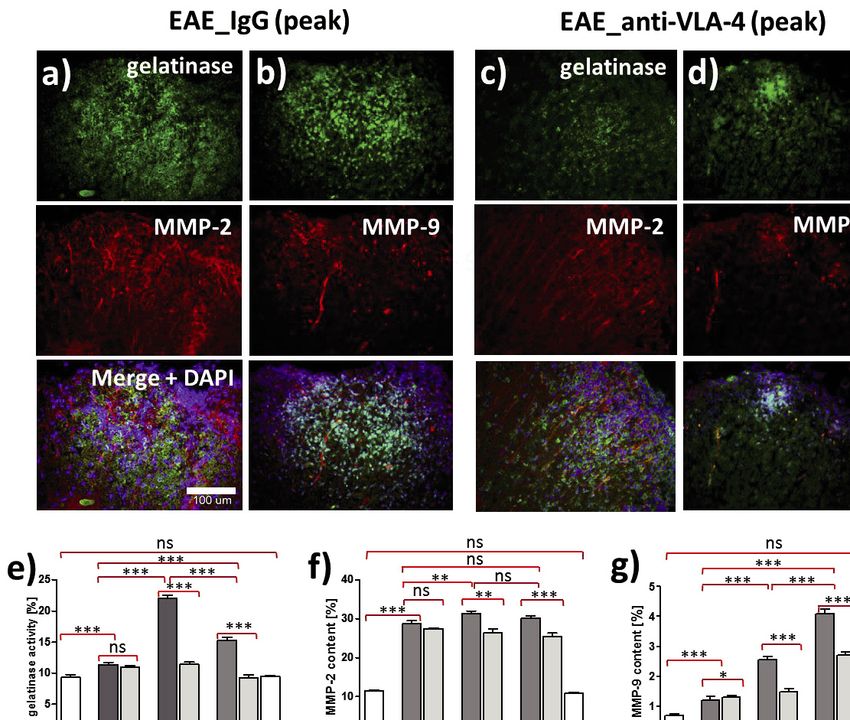
Fig. 2. Gelatinase activity (a, b, c, d, top panel), immunohistochemical demonstration of MMP-2 (a, c, middle panel), MMP-9 (b, d,
middle panel) and overlapping fluorescence of gelatinase and MMPs (a, b, c, d, bottom panel), as well as their quantification (e, f, g)
in spinal cords of IgG and anti-VLA-4 treated mice. Percentage values concern cross-sectioned spinal cord surface area occupied by
immunofluorescent structures. Data are presented as means ± SEM; n = 5 per group. ***P < 0.001; **P < 0.01; *P < 0.05; ns - not
significant). All micrographs are at the same magnification.269 successive phases, whereas MMP-9 expression gradually Effect of anti-VLA-4 mAb treatment on matrix increased, reaching the maximum in the chronic phase, both in metalloproteinase inhibitors anti-VLA-4 and IgG treated mice (Fig. 2f and 2g). Generally, MMP-2 expression was significantly higher as compared to TIMP-1 expression in spinal cords of EAE mice was MMP-9, with maximum values of 31.4 ± 0.6% for MMP-2 and detected only in peak and chronic phases of the disease. In IgG 4.1 ± 0.2% for MMP-9 (Fig. 2f and 2g). This difference can also treated animals it significantly decreased between these phases be seen in the micrographs (Fig. 2a and 2c). Anti-VLA-4 mAb from 3.4 ± 0.1% to 2.8 ± 0.1%. Anti-VLA-4 mAb treatment treatment markedly reduced MMP-9 expression in peak and elevated TIMP-1 expression to 6.3 ± 0.1% in the peak phase and chronic phases (peak phase: from 2.5 ± 0.1% to 1.5 ± 0.1%, to 7.7 ± 0.1% in the chronic phase (Fig. 3c). chronic phase: from 4.1 ± 0.2% to 2.7 ± 0.1%, respectively, for Double labeling of MMP-2 and TIMP-2 showed their co- IgG and anti-VLA-4 mAb treated groups, respectively) (Fig. localization (Fig. 3b). With progression of the disease, TIMP-2 2g). The decrease in MMP-2 expression observed in the peak expression gradually increased from 1.1 ± 0.04% to 5.1 ± 0.1% and chronic phases was much weaker, although statistically in IgG group and from 1.5 ± 0.03% to 6.1 ± 0.1% in anti-VLA-4 significant (Fig. 2f). group, reaching the maximum during the chronic phase of the Fig. 3. Double immunofluorescence labeling for MMP-9, TIMP-1 (a), MMP-2,TIMP-2 (b) and overlapping fluorescence in the peak phase of EAE treated anti-VLA-4 group, as well as their quantitative measurements in successive phases of the disease (c, d, e, f). Percentage values concern cross-sectioned spinal cord surface area occupied by immunofluorescent structures. Data are presented as means ± SEM; n = 5 per group; nf - not found. ***P < 0.001; **P < 0.01; *P < 0.05; ns - not significant). All micrographs are at the same magnification.
270
disease (Fig. 3d). Hence, anti-VLA-4 mAb treatment augmented DISCUSSION
TIMP expression.
MMP-associated mechanisms which contribute to MS
Laminin expression pathology include blood-brain barrier (BBB) breakdown,
facilitated inflammatory infiltration into CNS (15), induction of
Localization of laminin was associated with the distribution inflammatory cytokines promoting tissue damage (16) and
of vascular basement membranes (Fig. 4a). Anti-VLA-4 mAb degradation of myelin sheaths leading to axonal damage (17).
treatment inhibited the degradation of laminin observed in IgG Upregulation of various MMPs and their tissue inhibitors
treated group in which laminin expression was 7.5 ± 0.1% in the (TIMPs) in the brain and spinal cord tissues of EAE mice has
onset phase and decreased to 4.3 ± 0.1% and 4.0 ± 0.1%, been demonstrated in several studies (18-22). As compared with
respectively, in the peak and chronic phases (Fig. 4b). Anti- wild-type mice, young MMP-9-deficient animals are more
VLA-4 mAb treatment maintained laminin expression at a stable resistant to EAE (23), whereas TIMP-1 null mice display a more
level in all phases (from 7.3 ± 0.1% to 7.5 ± 0.1%) (Fig. 4b). severe disease (24). Even though the MMP/TIMP regulation in
Double immunostaining showed that laminin co-localized with EAE is well documented, there are few data concerning its
MMP-9 (Fig. 4d), TIMP-1 (Fig. 4e) and TIMP-2 (Fig. 4f). temporal dynamics. We assessed expression of MMP-2, MMP-
Summarized results of the study are graphically presented 9, TIMP-1 and TIMP-2 during progression of EAE, which
in Fig. 5. according to clinical scores can be divided into three successive
Fig. 4. Immunohistochemical localization of laminin associated with the distribution of vascular basement membranes (a), its
quantitative measurements in the successive phases of EAE (b) and colocalization with MMPs and TIMPs (c, d, e, f). Percentage
values in (b) concern cross-sectioned spinal cord surface area occupied by immunofluorescent structures. Micrographs represent the
peak phase of EAE. Data are presented as means ± SEM; n = 5 per group. ***P < 0.001; **P < 0.01; *P < 0.05; ns - not significant).
In each micrograph group (a, c - f, insets) magnification is the same (indicated by scale bars).271
Fig. 5. Changes in the studied parameters of EAE plotted as a function of disease progression.
phases: onset, peak and chronic (recovery) phase. Anti-VLA-4 In control, IgG treated EAE mice, increase in expression of
mAb delayed the clinical disease onset and reduced clinical both, MMP-2 and MMP-9 was observed already in the onset
scores in the subsequent phases. phase. Further dynamics was different: MMP-2 expression only
The degree of the inflammatory infiltration strongly slightly increased in the peak phase and retained that level in the
correlates with disease severity (25). In our experiments, degree chronic phase, whereas the increase in MMP-9 expression in the
of inflammation in EAE IgG-treated mice increased during the peak phase was much stronger (twofold) and achieved maximum
first two phases of EAE and remained at the elevated level in the in the chronic phase. Quantitative PCR studies on MMP-9
chronic phase. These results agree with our earlier studies mRNA in the spinal cord homogenates of EAE animals yielded
showing similar trend of changes in the number of total discrepant results: the maximal values were found in the onset
leukocytes, T cells, macrophages and microglial cells (26). Anti- phase (20) and in the peak phase (21). However, we studied
VLA-4 mAb treatment significantly suppressed inflammatory expression of the protein and the duration of its synthesis can be
infiltration compared to IgG treated group. responsible for later occurrence of maximal expression.
Expression of the studied MMPs and TIMPs was observed In the course of MS, MMP-9 and MMP-2 seem to be
in both, gray matter and white matter, so we assessed the overall reciprocally associated with the stages of the disease: increased
expression in the entire spinal cord sections. We found that MMP-9 levels and production was observed in the active phase
MMP-2 was the dominant metalloproteinase as far as of MS (30), whereas elevated MMP-2 was characteristic for MS
quantitative expression of the protein was concerned and this remission (31). Results of the present study confirmed an
observation remains in agreement with other studies (27). increase in MMP-9 expression in the peak phase of EAE, but
However, its role in the pathogenesis of EAE seems to be less MMP-2 did not significantly change between peak and chronic
significant compared to MMP-9. MMP-9 causes more severe phases. This discrepancy could result from incomplete adequacy
axonal injury than MMP-2 (28). In our material, anti-VLA-4 of the human disease and its animal model.
mAb treatment only slightly inhibited MMP-2 expression but MMP-2 has been detected in the early stages of the ischemic
significantly down-regulated MMP-9 and the decrease in its event and appears to be involved in reversible early opening of
expression was accompanied by inhibition of inflammation. It is BBB (32). Our study demonstrated a rapid increase in its
not clear, whether the effect of anti-VLA-4 mAb treatment is expression already in the onset phase. These findings highlight
direct or indirect: MMP and TIMP expression is influenced by the potential early role of MMP-2 in the degradation of vascular
proinflammatory cytokines (29), hence it seems probable that basement membranes, even though its effect in the further
the observed changes result from the inflammatory status of the phases seems to be weaker than that of MMP-9. The latter has
spinal cord in the successive phases of EAE. been demonstrated to be an inflammatory modulator associated272
with adverse coronary and carotid ischemic events in patients blood-brain barrier in experimental autoimmune
with the symptomatic internal carotid artery stenosis (33). encephalomyelitis. J Cell Biol 2001; 153: 933-945.
In this study we examined TIMP-2 - tissue inhibitor of 6. Oki T, Takahashi S, Kuwabara S, et al. Increased ability of
MMP-2 and TlMP-1 inhibiting most MMPs including MMP-9 peripheral blood lymphocytes to degrade laminin in multiple
(34). The temporal dynamics of the expression of TIMPs in EAE sclerosis. J Neurol Sci 2004; 222: 7-11.
mice shows some delay in relation to the dynamics of respective 7. Rice GP, Hartung HP, Calabresi PA. Anti-alpha4 integrin
MMPs: TIMP-1 is first observed in the peak phase, although therapy for multiple sclerosis: mechanisms and rationale.
elevation of MMP-9 occurs already in the onset phase and Neurology 2005; 64: 1336-13342.
TIMP-2 shows the gradual increase from the onset to the chronic 8. Madri JA, Graesser D, Haas T. The roles of adhesion
phase, whilst MMP-2 rapidly rises in the onset phase and then molecules and proteinases in lymphocyte transendothelial
remains at the similar level (Fig. 3e and 3f). This delay suggests migration. Biochem Cell Biol 1996; 74: 749-757.
that EAE primarily influences MMPs and increase in TIMP 9. Yednock TA, Cannon C, Fritz LC, Sanchez-Madrid F,
expression is at least partly a secondary, compensatory effect. Steinman L, Karin N. Prevention of experimental
Generally, anti-VLA-4 mAb treatment lowered MMP autoimmune encephalomyelitis by antibodies against a4b1
expression and elevated TIMP expression. In case of MMPs, this integrin. Nature 1992; 356: 63-66.
effect was delayed: it was absent (MMP-2) or slight (MMP-9) in 10. Polman CH, O’Connor PW, Havrdova E, et al. A randomized,
the onset phase and evident in the successive phases. Such delay placebo-controlled trial of natalizumab for relapsing multiple
was not observed in case of TIMPs, suggesting that anti-VLA-4 sclerosis. N Engl J Med 2006; 354: 899-910.
mAb treatment upregulate TIMPs which in turn inhibit 11. Theien BE, Vanderlugt CL, Eagar TN, et al. Discordant
expression of MMPs, although a parallel, direct effect of anti- effects of anti-VLA-4 treatment before and after onset of
VLA-4 mAbs on MMPs can not be excluded. relapsing experimental autoimmune encephalomyelitis. J Clin
Blood-brain barrier (BBB) damage leads to the leakage and Invest 2001; 107: 995-1006.
elevated plasma levels of neuronal proteins, which in stroke 12. Iannetta M, Zingaropoli MA, Latronico T, et al. Dynamic
patients are important for disease prognosis (35). changes of MMP-9 plasma levels correlate with JCV
Proteolytic disruption of the BBB occurs when the MMPs reactivation and immune activation in natalizumab-treated
attack vascular basal laminae (15). It was reported that MMP- multiple sclerosis patients. Sci Rep 2019; 9: 311. doi:
9 degraded laminin, the main component of the basal laminae 10.1038/s41598-018-36535-5
and that this degradation not only increased BBB permeability, 13. Castellazzi M, Bellini T, Trentini A, et al. Serum gelatinases
but also induced neuronal apoptosis (36). We found that anti- levels in multiple sclerosis patients during 21 months of
VLA-4 mAb inhibited laminin degradation by decreasing natalizumab therapy. Dis Markers 2016; 2016: 8434209.
MMP expression. Interestingly, MMP-9, but not MMP-2 doi: 10.1155/2016/8434209
colocalized with laminin and this could be regarded as an 14. Marques F, Mesquita SD, Sousa JC, et al. Lipocalin 2 is
additional argument for the superior role of MMP-9 in the present in the EAE brain and is modulated by natalizumab.
pathogenesis of EAE. Front Cell Neurosci 2012; 6: 33. doi:
In summary, our findings demonstrate that anti-VLA-4 10.3389/fncel.2012.00033
mAbs not only inhibit a4b1 integrin-dependent transmigration 15. Rosenberg GA. Matrix metalloproteinases and
of immune cells across vascular wall, but also - probably neuroinflammation in multiple sclerosis. Neuroscientist
indirectly - downregulate MMP-2 and MMP-9, as well as 2002; 8: 586-595.
upregulate TIMP-1 and TIMP-2, counteracting disintegration of 16. Chandler S, Miller KM, Clements JM, et al. Matrix
the vascular basal laminae. metalloproteinases, tumor necrosis factor and multiple
sclerosis: an overview. J Neuroimmunol 1997; 72: 155-161.
Acknowledgments: The study was supported by a grant 17. Diaz-Sanchez M, Williams K, DeLuca GC, Esiri MM.
N41/DBS/000564 from the Jagiellonian University Medical Protein co-expression with axonal injury in multiple
College to GFP. sclerosis plaques. Acta Neuropathol 2006; 111: 289-299.
18. Toft-Hansen H, Nuttall RK, Edwards DR, Owens T. Key
Conflict of interests: None declared. metalloproteinases are expressed by specific cell types in
experimental autoimmune encephalomyelitis. J Immunol
2004; 173: 5209-5218.
REFERENCES 19. Pagenstecher A, Stalder AK, Kincaid CL, Shapiro SD,
Campbell IL. Differential expression of matrix
1. Constantinescu CS, Farooqi N, O’Brien K, Gran B. metalloproteinase and tissue inhibitor of matrix
Experimental autoimmune encephalomyelitis (EAE) as a metalloproteinase genes in the mouse central nervous system
model for multiple sclerosis (MS). Br J Pharmacol 2001; in normal and inflammatory states. Am J Pathol 1998; 152:
164: 1079-1106. 729-741.
2. Boziki M, Grigoriadis N. An update on the role of matrix 20. Clements JM, Cossins JA, Wells GM, et al. Matrix
metalloproteinases in the pathogenesis of multiple sclerosis. metalloproteinase expression during experimental
Med Chem 2018; 14: 155-169. autoimmune encephalomyelitis and effects of a combined
3. Ozenci V, Rinaldi L, Teleshova N, et al. Metalloproteinases matrix metalloproteinase and tumour necrosis factor-alpha
and their tissue inhibitors in multiple sclerosis. J Autoimmun inhibitor. J Neuroimmunol 1997; 74: 85-94.
1999; 12: 297-303. 21. Kieseier BC, Kiefer R, Clements JM, et al. Matrix
4. Candelario-Jalil E, Thompson J, Taheri S, et al. Matrix metalloproteinase-9 and -7 are regulated in experimental
metalloproteinases are associated with increased blood-brain autoimmune encephalomyelitis. Brain 1998; 121: 159-166.
barrier opening in vascular cognitive impairment. Stroke 22. Weaver A, Goncalves da Silva A, Nuttall RK, et al. An elevated
2011; 42: 1345-1350. matrix metalloproteinase in experimental autoimmune
5. Sixt M, Engelhardt B, Pausch F, Hallmann R, Wendler O, encephalomyelitis is protective by affectingTh1/Th2
Sorokin LM. Endothelial cell laminin isoforms, laminins 8 polarization. FASEB J 2005; 19: 1668-1670.
and 10, play decisive roles in T cell recruitment across the273
23. Dubois B, Masure S, Hurtenbach U, Paemen L, et al. intrathecal synthesis of active matrix metalloproteinase-2
Resistance of young gelatinase B-deficient mice to (MMP-2) as markers of disease remission in patients with
experimental autoimmune encephalomyelitis and multiple sclerosis. Mult Scler 2009; 15: 547-554.
necrotizing tail lesions. J Clin Invest 1999; 104: 1507-1515. 32. Heo JH, Lucero J, Abumiya T, Koziol JA, Copeland BR, del
24. Thorne M, Moore CS, Robertson GS. Lack of TIMP-1 Zoppo GJ. Matrix metalloproteinases increase very early
increases severity of experimental autoimmune during experimental focal cerebral ischemia. J Cereb Blood
encephalomyelitis: effects of darbepoetin alfa on TIMP-1 null Flow Metab 1999; 19: 624-633.
and wild-type mice. J Neuroimmunol 2009; 211: 92-100. 33. Badacz R, Podolec J, Przewlocki T, et al. The role of
25. Stromnes IM, Goverman JM. Active induction of chemokine CCL5/RANTES and metalloproteinase-9 as
experimental allergic encephalomyelitis. Nat Protoc 2006; inflammatory modulators in symptomatic internal carotid
1: 1810-1819. artery stenosis. J Physiol Pharmacol 2019; 70: 545-555.
26. Pyka-Fosciak G, Stasiolek M, Litwin JA. 34. Maghsood F, Mirshafiey A, Farahani MM, Modarressi MH,
Immunohistochemical analysis of spinal cord components in Jafari P, Motevaseli E. Dual effects of cell free supernatants
mouse model of experimental autoimmune encephalomyelitis. from Lactobacillus acidophilus and Lactobacillus rhamnosus
Folia Histochem Cytobiol 2018; 56: 151-158. GG in regulation of MMP-9 by up-regulating TIMP-1 and
27. Rosenberg GA. Matrix metalloproteinases and their multiple down-regulating CD147 in PMA differentiated THP-1 cells.
roles in neurodegenerative diseases. Lancet Neurol 2009; 8: Cell J 2018; 19: 559-568.
205-216. 35. Lasek-Bal A, Jedrzejowska-Szypulka H, Student S, et al.
28. Newman TA, Woolley ST, Hughes PM, Sibson NR, The importance of selected markers of inflammation and
Anthony DC, Perry VH. T-cell- and macrophage-mediated blood-brain barrier damage for short-term ischemic stroke
axon damage in the absence of a CNS-specific immune prognosis. J Physiol Pharmacol 2019; 70: 209-217.
response: involvement of metalloproteinases. Brain 2001; 36. Gu Z, Cui J, Brown S, et al. A highly specific inhibitor of
124: 2203-2214. matrix metalloproteinase-9 rescues laminin from proteolysis
29. Clark IM, Swingler TE, Sampieri CL, Edwards DR. The and neurons from apoptosis in transient focal cerebral
regulation of matrix metalloproteinases and their inhibitors. ischemia. J Neurosci 2005; 25: 6401-6408.
Int J Biochem Cell Biol 2008; 40: 1362-1378.
30. Fainardi E, Castellazzi M, Bellini T, et al. Cerebrospinal R e c e i v e d : March 18, 2020
fluid and serum levels and intrathecal production of active A c c e p t e d : April 30, 2020
matrix metalloproteinase-9 (MMP-9) as markers of disease
activity in patients with multiple sclerosis. Mult Scler 2006; Author’s address: Dr. Grazyna Pyka-Fosciak, Department of
12: 294-301. Histology, Jagiellonian University Medical College, 7
31. Fainardi E, Castellazzi M, Tamborino C, et al. Potential Kopernika Street, 31-034, Cracow, Poland.
relevance of cerebrospinal fluid and serum levels and E-mail: gpfosciak@cm-uj.krakow.plYou can also read

















































