Gene expression patterns of red sea urchins (Mesocentrotus franciscanus) exposed to different combinations of temperature and pCO2 during early ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Wong and Hofmann BMC Genomics (2021) 22:32
https://doi.org/10.1186/s12864-020-07327-x
RESEARCH ARTICLE Open Access
Gene expression patterns of red sea
urchins (Mesocentrotus franciscanus)
exposed to different combinations of
temperature and pCO2 during early
development
Juliet M. Wong1,2* and Gretchen E. Hofmann1
Abstract
Background: The red sea urchin Mesocentrotus franciscanus is an ecologically important kelp forest herbivore and
an economically valuable wild fishery species. To examine how M. franciscanus responds to its environment on a
molecular level, differences in gene expression patterns were observed in embryos raised under combinations of
two temperatures (13 °C or 17 °C) and two pCO2 levels (475 μatm or 1050 μatm). These combinations mimic various
present-day conditions measured during and between upwelling events in the highly dynamic California Current
System with the exception of the 17 °C and 1050 μatm combination, which does not currently occur. However, as
ocean warming and acidification continues, warmer temperatures and higher pCO2 conditions are expected to
increase in frequency and to occur simultaneously. The transcriptomic responses of the embryos were assessed at
two developmental stages (gastrula and prism) in light of previously described plasticity in body size and
thermotolerance under these temperature and pCO2 treatments.
Results: Although transcriptomic patterns primarily varied by developmental stage, there were pronounced
differences in gene expression as a result of the treatment conditions. Temperature and pCO2 treatments led to the
differential expression of genes related to the cellular stress response, transmembrane transport, metabolic
processes, and the regulation of gene expression. At each developmental stage, temperature contributed
significantly to the observed variance in gene expression, which was also correlated to the phenotypic attributes of
the embryos. On the other hand, the transcriptomic response to pCO2 was relatively muted, particularly at the
prism stage.
(Continued on next page)
* Correspondence: juliwong@fiu.edu
1
Department of Ecology, Evolution and Marine Biology, University of
California Santa Barbara, Santa Barbara, CA 93106, USA
2
Present address: Department of Biological Sciences, Florida International
University, North Miami, FL 33181, USA
© The Author(s). 2021 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License,
which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if
changes were made. The images or other third party material in this article are included in the article's Creative Commons
licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the
data made available in this article, unless otherwise stated in a credit line to the data.Wong and Hofmann BMC Genomics (2021) 22:32 Page 2 of 21 (Continued from previous page) Conclusions: M. franciscanus exhibited transcriptomic plasticity under different temperatures, indicating their capacity for a molecular-level response that may facilitate red sea urchins facing ocean warming as climate change continues. In contrast, the lack of a robust transcriptomic response, in combination with observations of decreased body size, under elevated pCO2 levels suggest that this species may be negatively affected by ocean acidification. High present-day pCO2 conditions that occur due to coastal upwelling may already be influencing populations of M. franciscanus. Keywords: Red sea urchin, Mesocentrotus franciscanus, RNA-seq, Transcriptomics, Early development, Climate change, Warming, Ocean acidification Background and early summer months [18–20]. Therefore, in the The red sea urchin Mesocentrotus franciscanus (A. Agas- CCS, M. franciscanus embryos and larvae experience siz, 1863) is an ecologically and economically valuable combinations of temperature and pH conditions that species found along the Pacific Coast of western North may vary depending on whether spawning and upwelling America [1]. In subtidal areas, especially within kelp for- events coincide. Furthermore, these urchins may be par- ests, these echinoderms are herbivorous ecosystem engi- ticularly vulnerable to stress during early development. neers that can shape the flow of resources within marine Although planktonic embryological and larval stages of habitats [2]. Overgrazing by M. franciscanus, often in echinoids are capable of exhibiting vertical migration combination with overgrazing by the purple sea urchin [21, 22], they are likely less capable of finding refuge Strongylocentrotus purpuratus, can lead to the formation from stressful conditions than their benthic adult coun- of urchin barrens in which macroalgal communities are terparts. There is also evidence that many organisms are severely reduced or depleted [3, 4]. Red sea urchins also most vulnerable to environmental stress early in their function as prey to animals at higher trophic levels, in- life history [23–26]. Both lethal and sublethal effects that cluding spiny lobsters and sea otters [5–7]. In addition occur during early development or that carry over into to its removal by natural predators, M. franciscanus is later life stages will negatively affect the recruitment ne- widely collected as a lucrative wild fishery species. Fish- cessary to support future populations [27, 28]. eries in Mexico, the United States, and Canada harvest Given the dynamic nature of their habitat and the pro- M. franciscanus for their gonads (i.e., roe) that supply gression of ocean warming and acidification, it is im- domestic markets as well as international exports, prin- perative to understand how early stage M. franciscanus cipally to Japan [8, 9]. Over recent years (2015–2019), respond to their environment on a molecular level. A the annual revenue reported from M. franciscanus fish- limited number of studies have investigated how M. eries across the states of California, Oregon, and Wash- franciscanus responds to temperature or pCO2 stress ington averaged over $7.1 million USD/year, far more [29–31], and even fewer have done so within a multi- than all other echinoderm fishery species combined [10]. stressor context [32]. In S. purpuratus, a species whose Given the considerable ecological and economic im- habitat largely overlaps with that of M. franciscanus, portance of M. franciscanus, determining how this spe- elevated temperatures and pCO2 levels during early cies will be affected by continuing environmental change development can cause increased mortality, abnormality, in coastal oceans remains an overlooked and critical area and a reduction in size and scope for growth [20, 33– of research [11]. Due to their habitat and life history, 35]. Several studies have identified and examined these urchins are threatened by climate change impacts temperature- and pCO2-responsive genes in S. purpura- [12] such as ocean warming, which may include sudden tus embryos and larvae [36–41]. Studies such as these and extreme marine heat waves [13, 14], and ocean acid- are essential for contributing molecular-level insights ification, which may amplify the low pH conditions that into how these organisms respond, or fail to respond, to episodically occur in upwelling regions [15]. The upwell- stressful environmental conditions and may help explain ing season in the California Current System (CCS) typic- effects observed at the level of the organism or popula- ally extends from early spring until late summer or fall; tion. This is particularly pertinent for fishery species in it is characterized by fluctuations between periods of up- which accurate predictions are necessary for adaptive, welling, when cold, low pH water is transported to the climate-ready fisheries management [42]. Although a surface, and periods in which upwelling is relaxed (i.e., clear understanding of how M. franciscanus responds to wind conditions are not conducive for driving upwelling) environmental stress is lacking, suggestions have already [16, 17]. This overlaps with the natural spawning period been made to replace or offset the M. franciscanus fish- of M. franciscanus that occurs annually during spring ery with Strongylocentrotus fragilis, a sea urchin species
Wong and Hofmann BMC Genomics (2021) 22:32 Page 3 of 21
expected to be more tolerant to climate change [12]. understanding of how M. franciscanus development is
Here, both temperature and pCO2 conditions were affected by current ocean conditions, as well as our pre-
manipulated in a laboratory setting to investigate their dictive capacity of how this species will respond to fu-
influence on the gene expression patterns of M. francis- ture ocean change.
canus during its early development. To the best of our
knowledge, this is the first study to use RNA sequencing
Results
(RNA-seq) to examine the M. franciscanus stress
Summary statistics and overview of RNA-seq
response.
The samples used for RNA-seq were generated from
In this study, M. franciscanus embryos were raised
triplicate cultures of embryos raised at each of the four
under a combination of two temperatures (13 °C or
combined temperature and pCO2 treatments (i.e., 12
17 °C) and two pCO2 levels (475 μatm or 1050 μatm)
total cultures) (see Additional file 1). Each sample was
that reflect current and future ocean conditions in their
collected as a pool of 5000 embryos from each of the 12
natural habitat [15, 43–45]. This generated four different
cultures at both the gastrula and prism stages during de-
treatment combinations: 1) 17 °C and 1050 μatm pCO2,
velopment to produce a total of 24 samples used for
2) 17 °C and 475 μatm pCO2, 3) 13 °C and 1050 μatm
RNA extractions and library preparation. Sequencing of
pCO2, and 4) 13 °C and 475 μatm pCO2. In the highly
the 24 libraries yielded a total of 728,782,735,100-bp sin-
dynamic CCS, treatment combinations #2–4 are cur-
gle reads. After quality trimming, an average of 30.3 ±
rently measured during and between upwelling events
1.3 million reads per library remained. FASTQC reports
[15, 43–45]. Future ocean conditions are represented by
[47] of trimmed sequences showed high sequence qual-
treatment combination #1 (i.e., the simultaneous occur-
ity (> 30) with limited adapter contamination or pres-
rence of 17 °C and 1050 μatm pCO2) given continued
ence of overrepresented sequences. Per-library mapping
ocean warming and acidification. Additional detail re-
efficiency to the developmental transcriptome [48] using
garding the selection of these temperature and pCO2
RSEM [49] was at an average of 52.6%. The presence of
treatment levels is located in the Methods. Gene expres-
mitochondrial rRNA appeared to have contributed to
sion patterns were assessed at both the gastrula and
the percentage of unmapped reads, although mapping
prism embryo stages. We also discuss the gene expres-
rate may have also been affected by the completeness of
sion results within the context of previously reported
the reference transcriptome.
physiological assessments from this experiment, includ-
ing body size and thermotolerance [46]. Here, we de-
scribe the effects of the temperature and pCO2 Developmental stage influenced transcriptomic patterns
treatments at the molecular level and whether they re- A principal component analysis (PCA) of sample-to-
late to observations made at the level of the organism. sample distances showed that differences in gene
Temperature elicited a robust transcriptomic response expression profiles were primarily between the two devel-
at both developmental stages. Gene expression analyses opmental stages, gastrula and prism (Fig. 1a). Principal
indicated that the warmer temperature (i.e., 17 °C) in- Component (PC) 1 captured the majority of the variance
duced a cellular stress response, amongst other pro- (67.5%) and revealed a clear separation between gastrula
cesses. Additionally, the variation in gene expression and prism stage embryos, while PC2 only captured 3.8%
that was significantly correlated to the temperature of the variance. Indeed, a permutational multivariate
treatment was also significantly correlated to embryo ANOVA across all 24 samples with developmental stage,
body size and thermotolerance, characteristics that were temperature treatment, and pCO2 treatment as fixed fac-
neutrally or positively influenced by the warmer tors, revealed that developmental stage explained 66.4% of
temperature treatment [46]. In contrast, the transcrip- the variance (p = 0.001) (Fig. 1b). In contrast, temperature
tomic response to the pCO2 treatment was compara- treatment explained only 4.1% of the variance (p = 0.041)
tively muted. This minor molecular-level response may and pCO2 treatment explained only 2.3% of the variance
explain the reduction in embryo body size that is ob- (p = 0.207). All factor interactions were not significant
served under elevated pCO2 levels (i.e., 1050 μatm) [46]. (p > 0.05). Results from gene expression analyses across all
Overall, we examined a valuable fishery species that is samples independent of stage (i.e., gastrula and prism
capable of dramatically shaping coastal ecosystems, and stages were not analyzed separately) are available in
determined that during early development M. francisca- Additional file 2. Because we have previously explored the
nus exhibits different magnitudes of transcriptomic differences in gene expression patterns across M. francis-
plasticity in response to two climate change-related canus during early development [48] and it is not the main
stressors. This study provides much needed insight into focus of the current study, from here onward we report
a species that is important for many fisheries on the separate gene expression analyses for the gastrula and
Pacific coast of North America, facilitating our prism stages.Wong and Hofmann BMC Genomics (2021) 22:32 Page 4 of 21 Fig. 1 General gene expression patterns. Principal component analysis (PCA) plots of a all samples, c the gastrula stage only, and e the prism stage only are displayed with the two components that explained the most variance. Pie charts (b, d, and f) display the percent of variation explained by fixed factors determined using permutational multivariate ANOVAs (*p < 0.05 and ***p < 0.001). For b all samples, fixed factors included developmental stage, temperature treatment, and pCO2 treatment. The interactions of the three fixed factors have been consolidated into a single, “Interactions” pie chart segment for figure simplicity. For d the gastrula stage and f the prism stage, fixed factors only included temperature and pCO2 treatment Temperature and pCO2 affected gastrula gene expression temperature treatment (Fig. 2a). PC2 captured 12.0% of Separate PCA plots were generated for the gastrula and the variance (Fig. 1c) and was found to have a significant prism stages. At the gastrula stage, we generally ob- positive correlation with the pCO2 treatment (Fig. 2a). served that both temperature and pCO2 treatments ap- Using average embryo length measurements from this peared to drive differences in gene expression patterns experiment (previously reported in [46]), both PC1 and across samples. A PCA of only the gastrula stage showed PC2 were also found to be negatively correlated with that replicate samples grouped together (Fig. 1c). Here, gastrula body size (Fig. 2a). PC1 captured 23.8% of the variance and was found to Upon examining the PC loadings for the gastrula stage have a highly significant negative correlation to the using the PCAtools package [50], the genes most
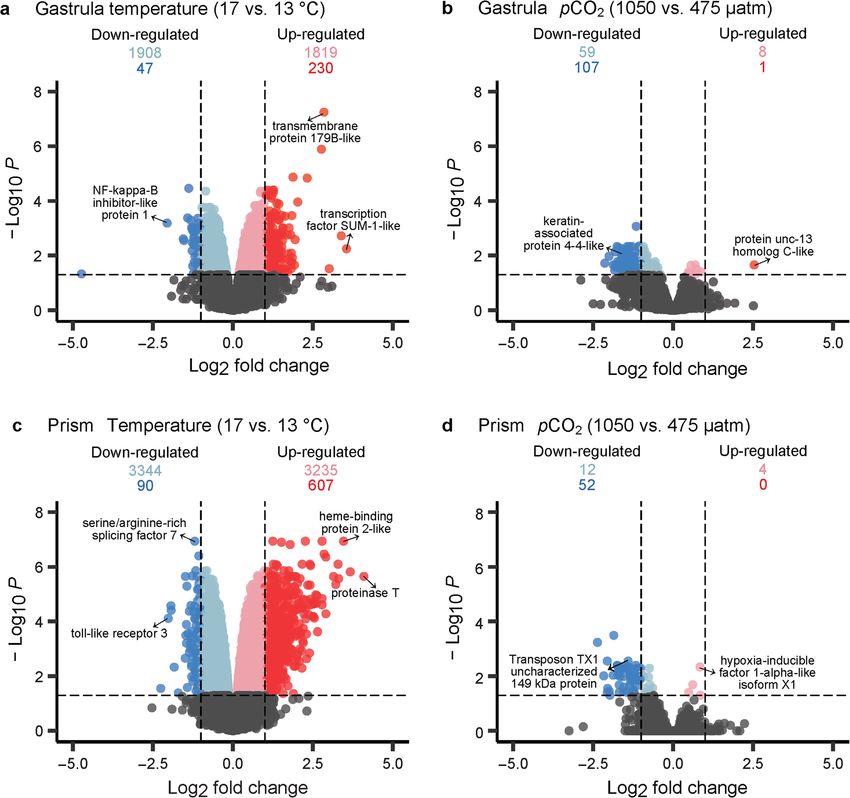
Wong and Hofmann BMC Genomics (2021) 22:32 Page 5 of 21 Fig. 2 Correlations at a the gastrula stage and b the prism stage between PC1-PC8 (columns), which contribute > 80% of the explained variation in gene expression, and metadata variables (rows) of the experiment treatments (i.e., temperature and pCO2), body size (i.e. embryo length in mm), and thermotolerance (i.e. LT50 in °C, prism stage only). The orange-purple color scale represents the strength of the Pearson’s correlation (1 to − 1). *p < 0.05, **p < 0.01, and ***p < 0.001 responsible for variation along PC1 included an elong- identify GO categories that were enriched by up- ation factor 1-alpha gene and a transcription factor regulated or down-regulated genes. Terms across SUM-1-like gene (Additional file 3). Genes contributing molecular function (MF), biological process (BP), and variation to PC2 included a poly(A)-specific ribonuclease cellular component (CC) GO categories were identified PARN gene and a putative DNA polymerase gene. A using moderated t-test values from the full list of genes rank-based gene ontology (GO) analysis was performed (i.e., not exclusively DE genes with adjusted p < 0.05). following the GO_MWU package [51] in R. Using GO categories significantly enriched with up-regulated complete sets of loading values calculated from the PCA genes influenced by temperature included DNA recom- (Additional file 3), this analysis identified GO categories bination, DNA metabolic process, cation channel, and G enriched by genes contributing variance to the PCs. GO protein-coupled receptor signaling pathway (Fig. 4a, terms related to regulation of gene expression, ion bind- Additional file 5a). ing, and DNA recombination were enriched by variable A total of 1955 genes were down-regulated in embryos genes in PC1 (Additional file 4a). Genes contributing raised at 17 °C relative to embryos raised at 13 °C (Fig. 3a). variation to PC2 enriched GO categories associated with These included a NF-kappa-B inhibitor-like protein 1 gene heat shock protein binding, peptide metabolic process, (log2 FC = − 2.06, adj. p < 0.001), a heat shock 70 kDa pro- and amide biosynthetic process (Additional file 4b). tein cognate 5 gene (log2 FC = − 0.27, adj. p = 0.019), and a A permutational multivariate ANOVA revealed that at heat shock 70 kDa protein 14 gene (log2 FC = − 0.40, adj. the gastrula stage, 20.3% of the variance was explained p = 0.003) (Additional file 3). GO categories enriched with by temperature treatment (p = 0.001) (Fig. 1d). Differen- down-regulated genes included regulation of gene expres- tial expression (DE) analyses conducted in limma [52] sion, chromatin organization, histone modification, and identified differentially expressed genes (relatively up- ion binding (Fig. 4a, Additional file 5a). and down-regulated) between gastrula raised under dif- The pCO2 treatment also affected gene expression pat- ferent temperature treatments. A total of 2049 genes terns at the gastrula stage, explaining 13.2% of the were significantly up-regulated in embryos raised at observed variance (p = 0.021) (Fig. 1d). Only 9 genes 17 °C relative to embryos raised at 13 °C (adjusted p < were up-regulated when comparing the 1050 μatm to 0.05) (Fig. 3a). These up-regulated genes included a the 475 μatm pCO2 treatment at the gastrula stage, transcription factor SUM-1-like gene (log2 fold change including a protein unc-13 homolog C-like gene (log2 (FC) = 3.39, adj. p = 0.002), a transmembrane protein FC = 2.54, adj. p = 0.022) (Fig. 3b, Additional file 3). GO 179B-like gene (log2 FC = 2.85, adj. p < 0.001), a cell analyses identified terms significantly enriched (p < 0.05) death protein 3 gene (log2 FC = 1.58, adj. p = 0.031), and with genes affected by the pCO2 treatment. GO categor- a heat shock 70 kDa protein 12A-like gene (log2 FC = ies enriched with up-regulated genes included macro- 0.661, adj. p = 0.023) (Additional file 3). molecule catabolic process, ion binding, and active Following DE analysis, gene ontology (GO) analyses transmembrane transporter (Fig. 4b, Additional file 5b). were performed using the GO_MWU package [51] to A total of 166 genes were down-regulated in embryos
Wong and Hofmann BMC Genomics (2021) 22:32 Page 6 of 21 Fig. 3 Temperature, and to a lesser degree, pCO2 treatments caused differential gene expression at a, b the gastrula stage and c, d the prism stage of early development. Genes that were not differentially expressed are displayed in gray while significant DE genes (adjusted p-value < 0.05) are displayed in color with a few selected genes labelled. Significant DE genes that were up-regulated are shown in pink (0 < log2 FC < 1) and red (log2 FC ≥ 1) and significant DE genes that were down-regulated are shown in light blue (− 1 < log2 FC < 0) and blue (log2 FC ≤ − 1) raised at 1050 μatm relative to embryos raised at 475 μatm Temperature was the primary factor affecting prism gene (Fig. 3b), including a keratin-associated protein 4–4-like expression gene (log2 FC = − 1.62, adj. p = 0.008) and a carbonic Similar to the gastrula stage, the PCA of only the prism anhydrase 14-like isoform X3 gene (log2 FC = − 0.89, adj. stage showed a separation of samples by treatment with p = 0.047) (Additional file 3). Enriched GO categories in- sample replicates grouping together (Fig. 1e). PC1, which cluded macromolecule biosynthetic process, macromol- captured 27.6% of the variance, was found to have highly ecule metabolic process, and nucleic acid binding (Fig. 4b, significant negative correlations to the temperature Additional file 5b). The interaction between temperature treatment, prism body size, and thermotolerance (Fig. and pCO2 factors explained 7.1% of the variance observed 2b). Prism body size for each sample was estimated at the gastrula stage, but the interaction was not signifi- using average embryo length and LT50 (i.e., the cant (p = 0.440) (Fig. 1d). temperature at which 50% mortality occurred)
Wong and Hofmann BMC Genomics (2021) 22:32 Page 7 of 21 Fig. 4 GO results of genes expressed at the gastrula stage. Analysis determined significant enrichment within GO categories of genes up- regulated (red text) and down-regulated (blue text) due to a temperature and b pCO2 treatments in gastrula embryos. Font sizes of the category names indicate the level of statistical significance as noted in the legend. The fraction preceding each category name is the number of genes with moderated t-statistic absolute values > 1 relative to the total number of genes belonging to the category. GO categories of molecular function (MF) and biological process (BP) are shown
Wong and Hofmann BMC Genomics (2021) 22:32 Page 8 of 21 measurements from this experiment that were previ- (log2 FC = 0.84, adj. p = 0.005) (Additional file 3). GO ously reported in [46]. PC2 captured 9.9% of the vari- categories enriched in up-regulated genes were related ance (Fig. 1e) and had a significant negative correlation to ATPase coupled to transmembrane movement of with the pCO2 treatment (Fig. 2b). ions, and regulation of gene expression (Fig. 5b, Add- Loadings within PC1 showed that genes responsible itional file 5d). A total of 64 genes were down-regulated for most of the variation included a heme-binding pro- in prism embryos raised at 1050 μatm pCO2 relative to tein 2-like gene and putative tolloid-like protein 1 genes those raised at 475 μatm (Fig. 3d), including a trans- (Additional file 3). Genes contributing variance to PC2 poson TX1 uncharacterized 149 kDa protein gene (log2 included an elongation factor 1-alpha gene, a F-box/WD FC = − 1.41, adj. p = 0.003). Enriched GO terms were re- repeat-containing protein 7-like gene, and a FK506- lated to oxidoreductase and G protein-coupled receptor binding protein 5-like gene (Additional file 3). Using (Fig. 5b, Additional file 5d). Lastly, the interaction be- GO_MWU and the loading values for the complete set tween temperature and pCO2 factors explained only of genes, enriched GO categories for PC1 were identi- 7.2% of the variance at the prism stage and was not sig- fied. These included GO terms related to ion transport, nificant (p = 0.362) (Fig. 1f). regulation of gene expression, methylated histone bind- ing, RNA methyltransferase, pseudouridine synthesis, Discussion antioxidant, cellular response to DNA damage stimulus, In this study, we examined how the gene expression pat- and response to oxidative stress (Additional file 4c). terns of M. franciscanus gastrula and prism embryos Genes contributing variation to PC2 enriched GO cat- varied by the developmental temperature and pCO2 con- egories associated with ion binding, oxidoreductase, and ditions under which they were raised. We also assessed metabolic process (Additional file 4d). whether the transcriptomic results aligned with the mor- A permutational multivariate ANOVA revealed that at phometric and physiological results previously reported the prism stage, 27.2% of the variance was explained by in [46]. Although both temperature and pCO2 can influ- the temperature treatment (p = 0.001) (Fig. 1f). DE ana- ence rates of sea urchin development [34, 53], any po- lysis showed a total of 3842 genes were up-regulated in tential differences in developmental timing should not embryos raised at 17 °C relative to those raised at 13 °C have impacted the results of this study because samples (Fig. 3c). These up-regulated genes included a proteinase were collected based on developmental progression to T gene (log2 FC = 4.10, adj. p < 0.001), a heme-binding the desired embryonic stages as detailed in the Methods, protein 2-like gene (log2 FC = 3.46, adj. p < 0.001), a rather than by hours post-fertilization. Overall, we found putative tolloid-like protein 1 gene (log2 FC = 3.21, adj. that while transcriptomic patterns varied by develop- p < 0.001), and a heat shock 70 kDa protein 12A-like mental stage, temperature had a dominant effect on gene (log2 FC = 0.91, adj. p < 0.001) (Additional file 3). changes in gene expression while pCO2 elicited a more GO analysis identified GO categories enriched in these subtle transcriptomic response that was largely limited up-regulated genes, which included oxidoreductase, re- to the gastrula stage. Experimental conditions impacted sponse to oxidative stress, ion transmembrane trans- genes related to the cellular stress response, transmem- porter, and ATP metabolic process (Fig. 5a, Additional brane transport, metabolic processes, and the regulation file 5c). A total of 3434 genes were down-regulated in of gene expression. embryos raised at 17 °C relative to those raised at 13 °C In terms of experimental design, embryos were ob- (Fig. 3c), including a toll-like receptor 3 gene (log2 FC = tained by evenly pooling eggs from five females and fer- − 2.02, adj. p < 0.001), a serine/arginine-rich splicing fac- tilizing them with sperm from a single male to produce tor 7 gene (log2 FC = − 1.19, adj. p < 0.001), a heat shock all full or half siblings. Admittedly, there are caveats to 70 kDa protein cognate 5 gene (log2 FC = − 0.21, adj. p = this approach. The results presented here may only be 0.021), and a heat shock 70 kDa protein 14 gene (log2 representative of a small subset of the population, or FC = − 0.48, p < 0.001) (Additional file 3). Enriched GO they may be driven by the quality of the particular male categories included regulation of gene expression, RNA selected to fertilize the eggs. Upon including data from modification, chromatin organization, cellular response our previous study that examined gene expression pat- to DNA damage stimulus, and metabolic process (Fig. terns during M. franciscanus early development [48], a 5a, Additional file 5c). PCA showed that, although samples primarily grouped The pCO2 treatment only explained 9.3% of the vari- by developmental stage, there is a clear distinction be- ance at the prism stage and was not significant (p = tween the embryos of the two studies (Additional file 6). 0.091) (Fig. 1f). In fact, only 4 genes were up-regulated This is likely due to a combination of genetic and envir- when comparing the 1050 μatm to the 475 μatm pCO2 onmental differences between the two source popula- treatment at the prism stage (Fig. 3d), including a tions, as the adult urchins were collected from different hypoxia-inducible factor 1-alpha-like isoform X1 gene sites and during different years. Indeed, in the purple sea
Wong and Hofmann BMC Genomics (2021) 22:32 Page 9 of 21 Fig. 5 GO results of genes expressed at the prism stage. Analysis determined significant enrichment within GO categories of genes up-regulated (red text) and down-regulated (blue text) due to a temperature and b pCO2 treatments in prism embryos. Font sizes of the category names indicate the level of statistical significance as noted in the legend. The fraction preceding each category name is the number of genes with moderated t-statistic absolute values > 1 relative to the total number of genes belonging to the category. GO categories of molecular function (MF) and biological process (BP) are shown
Wong and Hofmann BMC Genomics (2021) 22:32 Page 10 of 21
urchin S. purpuratus, genetic variation has been shown temperature and pCO2 treatments. For instance, many
to influence transcriptomic responses to temperature more genes were differentially expressed due to
and pCO2 stress during early development [36, 54]. temperature at the prism stage than at the gastrula stage
Given that the data presented here represents a limited (Fig. 3). Additionally, the pCO2 treatment explained a
selection of the genetic variation that exists in this spe- significant amount of variance in gene expression in gas-
cies, the results should be interpreted with caution. We trula embryos, but not later at the prism stage (Fig. 1d
therefore recommend that additional studies be per- and f). Similarly, the morphometric response to
formed within other M. franciscanus populations and temperature and pCO2 treatments varied by stage, in
with multiple male-female crosses to determine if our which pCO2, but not temperature, affected gastrula
results are unique to this study. Nevertheless, this ap- embryos by reducing body size under elevated pCO2
proach was implemented in an effort to limit genetic conditions (i.e., 1050 μatm) [46]. On the other hand,
variability and male-female interactions that may have temperature was the dominant factor at the prism stage,
otherwise confounded the molecular results. with warmer conditions (17 °C) increasing body size,
All samples used for RNA extractions were each com- offsetting the stunting effect of high pCO2 [46]. The ob-
posed of a pool of 5000 individuals and should thus served patterns between gene expression and body size
represent the same mixture of genotypes. Therefore, we will be described in greater detail later in the Discussion.
do not expect differences in gene expression patterns to Different life stages are predicted to have different sen-
be due to genetic variability between embryo cultures, par- sitivities to stress [23]. The variability between gastrula
ticularly because a low incidence of mortality was and prism stress responses may be explained by a differ-
observed during the experiment, although it was not ence in stage-specific vulnerability. During the gastrula
directly measured. In the absence of selection, the ob- stage, the archenteron is formed from invagination of
served variability in gene expression, body size, and ther- the embryo’s vegetal plate [55], a fundamental process
motolerance between embryos raised under different known as gastrulation that is essential for successful devel-
experimental treatments reflect plasticity exhibited by M. opment in metazoans [56]. At the prism stage, the embryo
franciscanus during its early development. We discuss this differentiates its digestive tract and develops skeletal rods,
plasticity, and how it may relate to embryo performance which are vital structures required for the embryos to
under different conditions that M. franciscanus are likely eventually become feeding, planktotrophic larvae [57, 58].
to experience in their natural environments currently and Accordingly, differences in responses to environmental
in the future under ocean change scenarios. conditions between these two stages are likely reflective of
the distinct processes undergone by these embryos to en-
Gene expression varied by developmental stage: general sure their continued developmental progression.
patterns The variability between stages could also be due to the
Developmental stage (gastrula or prism embryos) was timing and duration of exposure to stress. The effects of
the primary factor driving differences in gene expression a stressor can become increasingly deleterious as the
patterns across samples (Fig. 1a and b). In a past study, length of exposure continues, and organisms not permit-
we raised cultures of M. franciscanus embryos in a single ted adequate time to recover may exhibit increasingly
laboratory environment that mimicked average, non- poor performance. Furthermore, during development
stressful conditions in situ (i.e., 15 °C and 425 μatm there may be negative carry-over effects that persist into
pCO2) and documented significant transcriptomic differ- later life stages [59, 60]. Alternatively, organisms may ac-
ences between gastrula and prism stages [48]. Therefore, climate to stressful conditions over time, and are there-
there are many alterations in gene expression between fore less adversely affected by a stressor following the
these stages that occur as a result of development and are initial exposure. For example, in the coral Acropora hya-
independent of differences in environmental temperature cinthus, the immediate transcriptomic response to heat
and/or pCO2 conditions. This is also evident in Fig. 1a in stress was much higher than the transcriptomic response
which gastrula samples do not cluster with prism samples following 20 h of exposure to warmed conditions [61].
that share the same experimental treatment. Thus, it remains important to acknowledge that organ-
Because comparing gastrula versus prism gene expres- isms may responds differently to various environmental
sion patterns was not a goal of this study, no direct stressors depending on their life history as well as the
differential expression analyses were performed between timing and duration of the exposure.
stages, although gene expression analyses performed
independently of stage (i.e., without analyzing gastrula Temperature influenced gastrula embryos on a molecular
and prism stages separately) are reported in Additional level
file 2. Nevertheless, embryos at each developmental Temperature was the dominant factor influencing
stage exhibited different transcriptomic responses to changes in gene expression at the gastrula stage.Wong and Hofmann BMC Genomics (2021) 22:32 Page 11 of 21 Temperature explained 20.3% of the observed variance response, transmembrane transport, metabolism, and (p = 0.001) with 2049 genes up-regulated and 1955 genes gene expression regulation in M. franciscanus gastrula down-regulated in embryos raised under 17 °C relative embryos. to those raised under 13 °C (Figs. 1 and 3). Furthermore, PC1, which captured 23.8% of the gene expression vari- Cellular stress response (gastrula temperature) ance at the gastrula stage, was significantly correlated to Response to stress at the cellular level often includes the temperature treatment (Fig. 2a). In general, the ob- processes to mitigate or remove cell damage [62]. Cell served temperature effects on gene expression at the death protein 3, which encodes a protease involved in gastrula stage were approximately akin to those reported apoptosis [63], was up-regulated in gastrula embryos for the purple sea urchin S. purpuratus [36], whose bio- raised at 17 °C, indicating that the warmer temperature geographical distribution overlaps with that of M. fran- may have caused stress-induced programmed cell death. ciscanus. Here, DE analysis revealed that gastrula raised DE analysis also provided evidence of DNA damage and in the higher temperature treatment (i.e., 17 °C) repair. This is similar to observations in Acropora corals expressed genes associated with a cellular stress re- in which heat stress caused an up-regulation of DNA sponse. Gastrula embryos of S. purpuratus that were replication and repair genes [61, 64]. Here, GO terms re- raised under an 18 °C temperature treatment exhibited a lated to DNA metabolic process and DNA recombin- comparable cellular stress response by up-regulating ation were enriched with up-regulated genes under genes associated with cellular responses to reactive oxy- warmer temperature conditions (Fig. 4a). DNA meta- gen species and unfolded proteins [36]. We also found bolic processes can include both DNA synthesis and that M. franciscanus gastrula embryos raised under the degradation for the purposes of replication and repair. warmer treatment exhibited transcriptomic patterns in- Furthermore, DNA recombination in somatic cells has dicative of increased transmembrane transport, while been identified as a critical mechanism for DNA damage embryos from the colder treatment appeared to decrease repair [65, 66]. Taken together, 17 °C temperature condi- metabolic processes. Similarly, S. purpuratus increased tions appear to induce stress within the gastrula em- expression of ion channel, cell-cell signaling, and bryos, which undergo response mechanisms to combat metabolism genes at warmer temperatures [36]. Lastly, cellular damage. temperature appeared to impact the regulation of gene ex- pression in M. franciscanus gastrula embryos, including Transmembrane transport (gastrula temperature) genes related to epigenetic mechanisms. Temperature also Gastrula embryos raised under warmer temperatures appeared to influence how gene expression was regulated also increased expression of genes related to transmem- in S. purpuratus gastrula embryos with higher tempera- brane transport, potentially both within and between tures leading to a down-regulation of genes related to cells (i.e., cell-cell communication). For instance, genes transcription and RNA processing [36]. related to G protein-coupled receptor, cation channel, Despite similarities in how temperature influenced cell surface receptor signaling pathway, and vesicle- gastrula gene expression, unlike in M. franciscanus, mediated transport were up-regulated in gastrula em- there was an effect of temperature on S. purpuratus bryos raised under 17 °C relative to those raised under morphology. Specifically, S. purpuratus gastrula em- 13 °C (Fig. 4a). This is also supported by PC loadings, in bryos raised at warmer temperatures were significantly which genes related to ion binding and cation channel smaller in size [36], whereas M. franciscanus gastrula contributed variance to PC1 (Additional file 4a), which embryos did not significantly differ in size as a result of was significantly correlated to the temperature treat- temperature [46]. This could reflect differences in ex- ment (Fig. 2a). Increased transport of materials, par- perimental design between the studies (e.g., treatment ticularly ions, across cell membranes may indicate temperatures, breeding designs, and urchin collection osmoregulation and maintenance of homeostasis. This sites), or it could reflect differences that exist at the spe- aligns with reports in juvenile sea urchins of the spe- cies level. Unlike S. purpuratus, the temperature re- cies Loxechinus albus, in which gene expression alter- sponse of M. franciscanus gastrula embryos that ations under elevated temperatures provided evidence occurred at the molecular level was not reflected at the of increased active transmembrane transport of so- organismal level. We postulate that the transcriptomic dium and potassium ions [67]. differences between gastrula raised at 17 °C and 13 °C served to compensate for direct temperature effects and Metabolism (gastrula temperature) allowed the embryos to maintain the same size despite In S. purpuratus, gastrula embryos raised under warmer the temperature treatments. Below, we explore with temperatures up-regulated metabolic genes [36]. Here, greater detail how temperature affected the expression gastrula raised under the lower temperature treatment patterns of genes associated with the cellular stress expressed genes associated with metabolic depression.
Wong and Hofmann BMC Genomics (2021) 22:32 Page 12 of 21
Specifically, GO terms identified as negative regulation our data support that these modifications occurred in re-
of biological process and negative regulation of meta- sponse to the environment, additional approaches are re-
bolic process were significantly enriched by genes down- quired to determine the precise modifications, their
regulated in gastrula embryos raised under 17 °C relative locations, and their impact on gene expression. In this
to those raised under 13 °C (Fig. 4a). In this study, meta- study, there were more up-regulated than down-
bolic rates of embryos raised under different treatments regulated genes in the 17 °C versus the 13 °C gastrula
were not measured at the gastrula stage, but we may ex- treatment, and the up-regulated genes appeared to have
pect that, given the effect of temperature on biochemical a greater log2 FC in expression (Fig. 3a). Nevertheless,
reaction kinetics, metabolic rate should increase predict- future studies pairing comparative epigenetic analyses
ably with temperature [68]. Generally, higher metabolic with transcriptomic approaches are required to elucidate
rates have been recorded at warmer temperatures in how these various mechanisms influence gene expres-
marine ectotherms [69–72]. This positive correlation be- sion in response to different environmental conditions
tween temperature and metabolism has been observed in M. franciscanus.
in M. franciscanus at the adult stage [73].
pCO2 influenced gene expression of gastrula embryos
Regulation of gene expression (gastrula temperature) The pCO2 treatment influenced gene expression pat-
Temperature also had an evident effect on the regulation terns at the gastrula stage, although to a lesser degree
of gene expression in M. franciscanus gastrula embryos. than temperature, and the interaction between the two
GO terms enriched by genes relatively down-regulated factors was not significant (Fig. 1d). Gastrula pCO2 con-
in gastrula embryos raised under 17 °C relative to those ditions explained 13.2% of the observed variance (p =
raised under 13 °C (i.e., genes were comparatively up- 0.021) with 9 genes up-regulated and 166 genes down-
regulated in the colder temperature treatment) were regulated in embryos raised under 1050 μatm pCO2 rela-
identified as regulation of transcription by RNA poly- tive to those raised under 475 μatm pCO2 (Figs. 1 and
merase II, translation regulator, and regulation of gene 3). Additionally, PC2, which captured 12.0% of the gene
expression (Fig. 4a). Histone binding, histone modifica- expression variance at the gastrula stage, was signifi-
tion, chromatin remodeling, and chromatin organization cantly correlated to the pCO2 treatment (Fig. 2a). In this
genes were also associated with the 13 °C gastrula treat- study, we anticipated that the 475 μatm pCO2 treatment
ment. Histone and post-translational modifications are was not stressful, as it represented the average ambient
examples of epigenetic mechanisms. These, and other pCO2 levels M. franciscanus regularly experience in their
epigenetic modifications can act to regulate gene func- natural habitat [44]. Evidence suggests that calcifying
tion without altering the DNA sequence, promoting marine organisms such as M. franciscanus are sensitive
phenotypic plasticity and potentially modulating the re- to declines in ocean pH (i.e., increases in pCO2 levels)
sponse to different environmental conditions [74–76]. [84–86], and while M. franciscanus may periodically ex-
Histone variants and modifications may activate or re- perience elevated pCO2 conditions in nature during up-
press transcription processes by altering chromatin welling events [15, 43, 45, 87], the 1050 μatm pCO2
structures, impacting the regions of the genome that are treatment was expected to induce a stress response.
available for transcription [77], and have been shown to While the effect of pCO2 on gastrula gene expression
mediate responses to changing environmental conditions patterns was less than the effect of temperature, there
in marine organisms [78–80]. Additional analyses such was a pronounced impact of pCO2 conditions on gas-
as ChIP-seq (i.e., chromatin immunoprecipitation se- trula body size. Gastrula raised under elevated pCO2
quencing) to locate regions targeted by histone modifica- conditions (i.e., 1050 μatm) were significantly smaller
tions and DNA-binding proteins [81, 82] or ATAC-seq than those raised under 475 μatm [46]. Therefore, pCO2
(i.e., assay for transposase-accessible chromatin with appeared to have a greater influence at the organismal
high-throughput sequencing) to assess genome-wide level but elicited a relatively muted transcriptomic re-
chromatin accessibility [83] are required to profile spe- sponse compared to temperature. Below we discuss the
cific histone variants or modifications and their impact expression patterns of genes affected by the gastrula
on gene expression. pCO2 treatment, which included those related to metab-
Although evidence of transcription regulation was ob- olism and ion transport.
served, it is difficult to conclude if this led to an increase
or decrease of gene expression, particularly with respect Metabolism and ion transport (gastrula pCO2)
to the functional significance of histone modifications Gastrula raised under the lower pCO2 treatment (i.e.,
and chromatin remodeling. These mechanisms are much 475 μatm) expressed genes that significantly enriched
less studied in marine invertebrates than other modifica- GO terms related to several macromolecule biosynthetic
tions such as DNA methylation [74, 75], and although processes (Fig. 4b). In contrast, those raised under theWong and Hofmann BMC Genomics (2021) 22:32 Page 13 of 21
elevated pCO2 treatment (i.e., 1050 μatm) expressed and the response to resulting cellular damage, due to el-
genes that enriched GO terms associated with macro- evated temperatures have been documented across a
molecule catabolic processes (Fig. 4b). This may, in part, wide variety of taxa, including algae [90], plants [91],
explain the difference in body size that was observed as mollusks [92], and fishes [93, 94]. GO enrichment from
a result of the pCO2 conditions. Gastrula in the 475 the DE analysis identified terms including oxidoreduc-
μatm pCO2 treatment appeared to construct proteins tase, antioxidant, response to oxidative stress, and re-
and other macromolecules to maintain their growth and sponse to reactive oxygen species that were enriched
body size, while gastrula in the 1050 μatm pCO2 treat- with genes up-regulated in response to the warmer
ment underwent catabolic processes, possibly to obtain temperature treatment (Fig. 5a). At the gastrula stage,
the energy required to respond to elevated pCO2 levels. embryos in the 17 °C treatment expressed genes associ-
The pCO2 stress response may include an increase of ated with DNA repair, but there was no evidence of in-
ion transport as a means of maintaining acid-base equi- creased macromolecule repair gene expression at the
librium given elevated H+ concentrations under high prism stage. This could indicate that while both stages
pCO2 conditions. GO terms related to ion binding, ac- initiated a cellular stress response due to warmer tem-
tive transmembrane transporter, and ATPase coupled to peratures, there was less cellular damage of nucleic acids
movement of substances were enriched by genes up- incurred by the prism stage.
regulated in gastrula raised in the 1050 μatm treatment In general, global change biology research in marine
(Fig. 4b). Similarly, increased expression of ion transport systems has focused on the negative consequences of in-
genes has been observed in gastrula embryos of S. pur- creasing temperatures associated with ocean warming
puratus exposed to moderately elevated pCO2 levels [95]. However, given the expected rise in variable and
(e.g., ~ 800 μatm) [39], although the transcriptomic re- extreme weather events, the impact of decreased tem-
sponse of S. purpuratus embryos to pCO2 stress can be peratures is also an important consideration, especially
influenced by maternal effects [38]. in regions dominated by upwelling. DE analysis indicated
that prism embryos in the 13 °C treatment increased ex-
Temperature was the dominant factor at the prism stage pression of genes related to DNA damage and repair.
At the prism stage, temperature accounted for 27.2% of Specifically, identified GO terms included cellular re-
the observed variance (p = 0.001) with 3842 genes up- sponse to DNA damage stimulus, DNA metabolic
regulated and 3434 genes down-regulated in embryos process, DNA binding, and catalytic, acting on DNA
raised under 17 °C relative to those raised under 13 °C (Fig. 5a). Increased DNA damage as a result of low
(Figs. 1 and 3). Furthermore, PC1 for the prism stage temperature stress has been recorded in the Pacific
captured 27.6% of variance in gene expression and was white shrimp Litopenaeus vannamei [96]. Although
significantly correlated to the temperature treatment 13 °C is within the range of temperatures that M. francis-
(Fig. 2b). Unlike at the gastrula stage, responses to canus experience in the Santa Barbara Channel (SBC)
temperature that were measured at the molecular level where urchins were collected for this study, it is lower
were also observable at the organismal level. Develop- than the annual average of ~ 15 °C [44] and may have
ment at the warmer 17 °C treatment led to an increase generated stress and cellular damage in the prism em-
in prism body size as well as a modest increase in prism bryos, leading to the activation of repair mechanisms.
thermotolerance [46]. Indeed, prism body size and ther-
motolerance variables were also highly correlated to PC1 Transmembrane transport and metabolism (prism
(Fig. 2b). Therefore, the transcriptomic response to temperature)
temperature appears to have influenced both growth and Similar to the gastrula stage, warmer temperature condi-
resistance to heat stress in M. franciscanus prism em- tions caused an increased expression of genes related to
bryos. Prism embryos raised at 17 °C exhibited increased transmembrane transport in prism embryos. Specifically,
expression of genes related to the cellular stress re- GO terms of ion binding, ion transmembrane trans-
sponse, transmembrane transport, and metabolic pro- porter, cation channel, and sodium and potassium ion
cesses, while genes related to DNA repair and the transmembrane transporters were enriched with up-
regulation of gene expression were associated with the regulated genes (Fig. 5a), which may indicate osmoregu-
colder temperature treatment (i.e., 13 °C). lation and the maintenance of homeostasis. Prism
embryos in the 17 °C treatment also increased expression
Cellular stress response (prism temperature) of genes related to energetic processes (e.g., ATP meta-
Environmental stress can lead to the production of re- bolic process, organic acid metabolic process, lipid meta-
active oxygen species (ROS), which can cause oxidative bolic process, cyclic nucleotide metabolic process, and
stress if ROS production exceeds the organism’s antioxi- carbohydrate metabolic process) possibly to generate the
dant or damage repair capacity [88, 89]. Oxidative stress, energy required to support active transmembraneWong and Hofmann BMC Genomics (2021) 22:32 Page 14 of 21
transport of ions and other materials. The up-regulation It is possible that while there was a clear phenotypic
of genes related to energy production may have also difference in prism embryos raised under high versus
supported the increased growth of prism embryos under low pCO2 conditions, the transcriptomic changes under-
warmer temperatures. In contrast, DE analysis revealed lying this difference were much more subtle. Alterna-
an increase in expression of genes related to the negative tively, the prism stage may simply lack a robust
regulation of biological and metabolic processes in prism transcriptional response to the 1050 μatm pCO2 treat-
embryos raised at 13 °C. This supports the predicted ex- ment. For instance, the Mediterranean sea urchin Para-
pectation that organisms exhibit decreased metabolism centrotus lividus exhibits different transcriptomic
under colder temperatures [68–72]. responses depending on the magnitude of the pH stres-
sor [101]. Decreased pH conditions caused P. lividus
embryos to increase their expression of calcification
Regulation of gene expression (prism temperature)
genes, but not once the pH dropped below a certain
At the prism stage, the colder temperature treatment ex-
threshold [101]. A similar result was observed in S. pur-
hibited an enrichment of genes related to translation
puratus in which embryos raised under a high pCO2
regulator, transcription coregulator, regulation of gene
treatment designed to reflect near-future levels exhibited
expression, and regulation of transcription by RNA poly-
a muted transcriptomic response relative to those raised
merase II (Fig. 5a). Additionally, GO terms identified as
under a more moderate pCO2 treatment designed to re-
histone binding, chromatin organization, RNA modifica-
flect present-day low pH conditions [39]. The authors
tion, and methylation-dependent protein binding were
speculated that the transcriptional response required for
enriched in genes down-regulated at 17 °C relative to
acclimating to a more extreme pCO2 level was too meta-
13 °C (Fig. 5a). Thus, gene expression in prism embryos
bolically expensive, and the embryos instead opted to
raised at 13 °C appeared to be epigenetically regulated by
conserve energy to ensure short-term survival, perhaps
histone and RNA modifications. This differential expres-
until environmental conditions became more favorable
sion of genes related to gene expression regulation (i.e.,
[39]. While a failure of embryos to respond at the tran-
down-regulated at the colder temperature relative to the
scriptomic level may permit continued successful devel-
warmer temperature) is similar to the pattern observed
opment under high pCO2 conditions, there may be
at the gastrula stage.
important physiological consequences such as the ob-
served reduction in body size [46]. Thus, the lack of a
The prism stage exhibited a limited transcriptomic transcriptomic response to high pCO2 may have import-
response to pCO2 ant fitness consequences for M. franciscanus.
In other studies, echinoderms raised under elevated In the green sea urchin Strongylocentrotus droeba-
pCO2 conditions have exhibited altered expression of chiensis, a quantitative genetic breeding design imple-
genes related to skeletogenic pathways, spicule matrix mented by Runcie and colleagues demonstrated that
proteins, cellular stress response, ion regulation and changes in gene expression as a result of differences in
transport, apoptosis, metabolism and ATP production pH exposure were minor relative to gene expression dif-
[38, 39, 97–100]. Here, the pCO2 treatment had a rela- ferences as a result of parentage [54]. Thus, minimal
tively minimal effect on prism gene expression patterns. transcriptomic responses to pCO2 may be due to the
The pCO2 treatment explained only 9.3% of the ob- genetic structure of the sea urchins used in this experi-
served variance at this stage and was not significant (p = ment. Furthermore, the environmental exposure history
0.091) (Fig. 1f). This contrasts with observations made at of the adult urchins may have generated non-genetic
the organism level in which elevated pCO2 resulted in parental effects (i.e., transgenerational plasticity), which
smaller prism embryos, although this could be offset by can also generate a limited transcriptomic response to
the positive effect of temperature, which acted as the high pCO2 [38]. While all embryo cultures for this ex-
dominant factor influencing body size [46]. It is interest- periment were composed of the same mixture of pro-
ing that the transcriptomic response to elevated pCO2 geny from a cross between one male and five females, it
was more evident in gastrula than in prism embryos par- is possible that the sea urchins collected for this experi-
ticularly because at the prism stage, skeletal rod forma- ment may be from a population with a relatively muted
tion occurs. Although GO terms identified as ion transcriptomic response to high pCO2 conditions.
binding and ATPase coupled to transmembrane move- It has been proposed that selection and local adapta-
ment of ions were enriched with genes up-regulated at tion act on populations that are regularly exposed to
the elevated pCO2 level (Fig. 5b), we also expected to high pCO2 conditions, such as those that often experi-
observe increased stress associated with prism embryos ence upwelling conditions within the CCS [15], and that
undergoing calcification processes under lowered pH these populations may harbor genotypes that are resist-
conditions, but we detected no evidence of this. ant to low pH conditions [40, 98, 102–105]. In S.You can also read

















































