Leaf-Derived Jasmonate Mediates Water Uptake from Hydrated Cotton Roots under Partial Root-Zone Irrigation1 OPEN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Leaf-Derived Jasmonate Mediates Water Uptake from
Hydrated Cotton Roots under Partial
Root-Zone Irrigation1[OPEN]
Zhen Luo,a,2 Xiangqiang Kong ,a,2 Yanjun Zhang,a Weijiang Li,a Dongmei Zhang,a Jianlong Dai,a
Shuang Fang,b Jinfang Chu,b and Hezhong Dong a,c,3,4
a
Cotton Research Center, Shandong Key Laboratory for Cotton Culture and Physiology, Shandong Academy
of Agricultural Sciences, Jinan 250100, People’s Republic of China
b
National Center for Plant Gene Research (Beijing), Institute of Genetics and Developmental Biology, Chinese
Academy of Sciences, Beijing 100101, People’s Republic of China
c
College of Life Sciences, Shandong Normal University, Jinan 250014, People’s Republic of China
ORCID ID: 0000-0003-4964-1784 (H.D.).
Partial root-zone irrigation (PRI), a water-saving technique, improves water uptake in hydrated roots by inducing specific
responses that are thought to be regulated by signals originating from leaves; however, this signaling is poorly understood.
Using a split-root system and polyethylene glycol 6000 to simulate PRI in cotton (Gossypium hirsutum), we showed that increased
root hydraulic conductance (L) and water uptake in the hydrated roots may be due to the elevated expression of cotton plasma
membrane intrinsic protein (PIP) genes. Jasmonate (jasmonic acid [JA] and jasmonic acid-isoleucine conjugate [JA-Ile]) content
and the expression of three JA biosynthesis genes increased in the leaves of the PRI plants compared with those of the
polyethylene glycol-free control. JA/JA-Ile content also increased in the hydrated roots, although the expression of the three
JA genes was unaltered, compared with the control. The JA/JA-Ile contents in leaves increased after the foliar application of
exogenous JA and was followed by an increase in both JA/JA-Ile content and L in the hydrated roots, whereas the silencing of
the three JA genes had the opposite effect in the leaves. Ring-barking the hydrated hypocotyls increased the JA/JA-Ile content in
the leaves but decreased the JA/JA-Ile content and L in the hydrated roots. These results suggested that the increased JA/JA-Ile
in the hydrated roots was mostly transported from the leaves through the phloem, thus increasing L by increasing the expression
of GhPIP in the hydrated roots under PRI. We believe that leaf-derived JA/JA-Ile, as a long-distance signal, positively mediates
water uptake from the hydrated roots of cotton under PRI.
Water resources are becoming increasingly scarce in need to improve irrigation management in order to
many regions due to global climate change and in- increase water use efficiency. Partial root-zone irriga-
creasing population growth. There is thus an urgent tion (PRI) is a water-saving irrigation technique that
improves water use efficiency without reducing yield
(Kang et al., 1997; Kang and Zhang, 2004; Fu et al.,
1
This work was supported by the National Key Research and De- 2017). Previous reports showed that PRI is associated
velopment Program of China (2017YFD0201906), the National Natu- with a relatively high level of photosynthetic activity
ral Science Foundation of China (31501249), the earmarked fund for
despite the reduced irrigation and decreased transpi-
the China Agricultural Research System (CARS-18-21), and the spe-
cial fund for Taishan Scholars (tspd20150213 and tsqn201812120).
ration compared with conventional irrigation (Kirda
2
These authors contributed equally to the article. et al., 2004; Zegbe et al., 2004; Tang et al., 2005). In ad-
3
Author for contact: donghezhong@163.com. dition to decreasing transpiration, increasing water
4
Senior author uptake from the hydrated roots is another important
The author responsible for distribution of materials integral to the means of maintaining the water balance in plants.
findings presented in this article in accordance with the policy de- McLean et al. (2011) detected a rapid increase in water
scribed in the Instructions for Authors (www.plantphysiol.org) is: uptake by the hydrated roots in the riparian tree
Hezhong Dong (donghezhong@163.com). Melaleuca argentea under PRI, which allowed leaf
H.D. and Z.L. conceived the original screening and research plans; function and water status to remain unchanged. This
H.D., Z.L., and X.K. designed the experiments; H.D. and W.L. super-
compensatory increase in water uptake from the wet
vised the experiments; Z.L. performed most of the experiments; Z.L.,
X.K., J.D., D.Z., and Y.Z. performed partial experiments and analyzed
zone plays an important role in maintaining water
the data; S.F. and J.C. provided technical assistance to Z.L.; Z.L. and status and plant growth under PRI (Wakrim et al.,
X.K. conceived the project and wrote the article with contributions 2005; Li et al., 2007; Wang et al., 2010). However, the
from all the authors; H.D. supervised and complemented the writing. underlying mechanisms of the increased water uptake
[OPEN]
Articles can be viewed without a subscription. from the wet root zone under PRI are still poorly
www.plantphysiol.org/cgi/doi/10.1104/pp.19.00315 understood.
1660 Plant PhysiologyÒ, July 2019, Vol. 180, pp. 1660–1676, www.plantphysiol.org Ó 2019 American Society of Plant Biologists. All Rights Reserved.
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.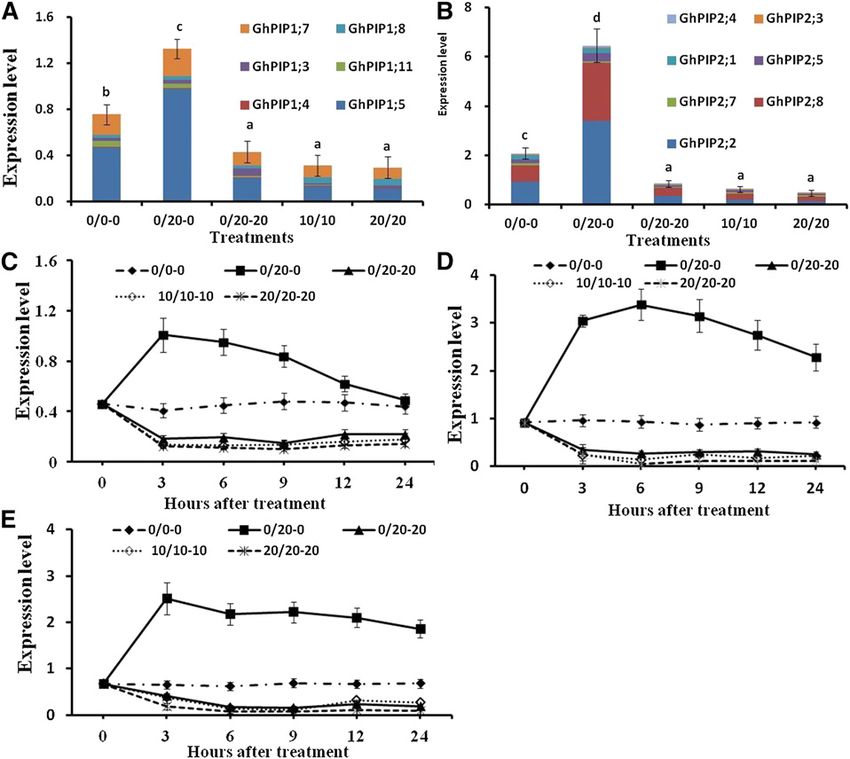
Leaf Jasmonate Mediates Water Uptake in Cotton
The water transport capacity of the roots depends on Fonseca et al., 2009; Wasternack and Hause, 2013).
the root hydraulic conductance (L), which is deter- Previous reports have indicated that JA and MeJA are
mined by root architecture and the intrinsic water the only precursors of JA-Ile, which is the bioactive
permeability of the root system (Sutka et al., 2011). compound involved in JA signaling (Staswick and
Plant roots take up water through parallel apoplastic, Tiryaki, 2004; Thines et al., 2007; Suza et al., 2010).
symplastic, and transcellular pathways (Steudle, 1994). Insectivory induced a high level of jasmonate accu-
Although anatomical changes affect apoplastic L over mulation in Drosera capensis leaves, and a good corre-
longer time scales, the rapid changes in the water per- lation was observed between JA and JA-Ile levels in the
meability of the roots are mainly regulated by aqua- plant tissues. By contrast, wounding induced a high
porins, especially the plasma membrane intrinsic transient increase in JA and only a low accumulation of
protein (PIP; Enstone et al., 2003; Maurel et al., 2010). JA-Ile, suggesting different responses between JA and
For instance, McLean et al. (2011) demonstrated that the JA-Ile to different stresses (Mithöfer et al., 2014). The
increased water uptake from the wet root zone under (9S,13S)-cis-oxophytodienoic acid reductase (OPR;
PRI was due to increased aquaporin levels. In land Schaller and Weiler, 1997), allene oxide synthase (AOS;
plants, aquaporins are classified into five homologous von Malek et al., 2002), and lipoxygenase (LOX;
subfamilies (Johanson et al., 2001; Sade et al., 2009), Bannenberg et al., 2009) enzymes are pivotal enzymes
with the PIP subfamily being the largest (Maurel et al., involved in the generation of jasmonates. It has been
2008). PIP subfamily aquaporins, including the PIP1 established that JAs regulate defense responses and
and PIP2 subgroups, play a key role in regulating root developmental processes (Campos et al., 2014; Howe
water uptake, and PIP2 proteins have a higher water et al., 2018), but their exact role in regulating L under
transport capacity than PIP1 proteins (Chaumont et al., PRI has been poorly studied.
2000; Siefritz et al., 2002; Tournaire-Roux et al., 2003; Abscisic acid (ABA) has long been recognized as the
Sakurai et al., 2005). primary plant stress hormone, regulating a wide range
Long-distance signaling plays an important role in of processes leading to enhanced stress tolerance
regulating plant development and stress responses. The (Cutler et al., 2010; Danquah et al., 2014). ABA is known
regulation of stomatal conductance by long-distance to modify L through up-regulating PIP gene expression
chemical signals from the roots to the leaves under or reducing the activation energy of the PIP protein
PRI has been well documented (Blackman and Davies, (Zhang et al., 1995; Wan et al., 2004; Aroca, 2006; Aroca
1985; Sobeih et al., 2004). However, the regulatory et al., 2008b; Ruiz-Lozano et al., 2009; Sánchez-Romera
mechanism of increased root L in the hydrated roots et al., 2014). SlPIP1-4 gene expression was up-regulated
under PRI has rarely been investigated. There is evi- by exogenous ABA under well-watered conditions in
dence that leaf-to-root signals may regulate root L by S. lycopersicum (Aroca et al., 2008a). Mahdieh and
mediating the quantity or activity of aquaporins Mostajeran (2009) found that PIP protein abundance
(Kudoyarova et al., 2011; Sakurai-Ishikawa et al., 2011). increased in tobacco (Nicotiana tabacum) roots when
Previous studies suggested that hydraulic signals, such ABA was added to the root medium. Exogenous ABA
as cell turgor pressure (Rygol et al., 1993), water po- enhanced the expression of the PIP1;7, PIP2;2, and
tential, and the osmolality of the xylem sap (Vandeleur PIP2;10 genes in the nonsaline side of cotton (Gossypium
et al., 2014), or chemical signals, such as plant hormones hirsutum) under nonuniform root salinity (Kong et al.,
(Ljung et al., 2002; Shah, 2009; Tungngoen et al., 2009; 2016). In addition to increasing protein abundance,
Sánchez-Romera et al., 2014), might regulate root water ABA may also regulate L at the posttranslational level.
uptake under biotic/abiotic stress. Osmotic stress can It might bind to PIP proteins, thus reducing the acti-
reduce root water content and increase the levels of the vation energy required for the change from closed to
compatible osmolyte Pro, which not only alters the cell open water channel states (Wan et al., 2004).
turgor pressure (Wilson et al., 2014) but also has other Water uptake is significantly increased in the hy-
significant roles, such as in signaling. drated root side under PRI, which may be regulated by
Chemical signals, including plant hormones and some signaling from the leaves or the dehydrated root
hydrogen peroxide, are believed to be involved in the side (McLean et al., 2011; Kong et al., 2016). Previous
regulation of L and water uptake (Aroca et al., 2005, studies also showed that specific hydraulic and/or
2008a, 2012; Benabdellah et al., 2009; Mahdieh and chemical signals may be involved in the regulation of
Mostajeran, 2009; Kong et al., 2016). Lee et al. (1996) water uptake in the hydrated root side under PRI (Boari
reported an improvement in the root bleeding rate of and Malone, 1993; Lee et al., 2008; Shah, 2009; Sánchez-
rice (Oryza sativa) following the application of exoge- Romera et al., 2014). However, the associated signaling
nous methyl jasmonate (MeJA), while Sánchez-Romera molecules and their sources are still unclear. In this
et al. (2014) suggested that jasmonic acid (JA) or MeJA study, a split-root system was established through
regulated L in Phaseolus vulgaris, Solanum lycopersicum, grafting to simulate PRI and to determine the physio-
and Arabidopsis (Arabidopsis thaliana) roots. Jasmonates logical and molecular bases of the increased water up-
(JA and jasmonic acid-isoleucine conjugate [JA-Ile]) is a take in the hydrated roots of cotton. We focused on
plant hormone that belongs to the octadecanoid family. determining (1) the exact signaling molecules involved
It can be converted into numerous conjugates and de- in regulating water uptake in the hydrated root side, (2)
rivatives, such as MeJA and JA-Ile (Pauwels et al., 2008; the source of the signaling molecules and the associated
Plant Physiol. Vol. 180, 2019 1661
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
gene expression patterns in the hydrated root sides, and Both the stomatal conductance and transpiration
(3) the manner in which the signaling molecules reach under PRI, MDI, and HDI decreased significantly at 24
the hydrated roots. HAT compared with that of the PEG-free control (Fig. 1,
C and D). The stomatal conductance and transpiration
of PRI were decreased by 26.4% and 23.9% compared
RESULTS with that of the PEG-free control, respectively. The
Increased Water Uptake and L of the Hydrated Roots stomatal conductance and transpiration of MDI and
under PRI HDI were significantly lower than those of PRI, and
there was no significant difference in the stomatal
As shown in Figure 1, in the treatment where the root conductance and transpiration between MDI and HDI
portions were subjected to 0% and 20% (w/v) poly- (Fig. 1, C and D).
ethylene glycol 6000 (PEG6000) treatment (PRI: 0/20),
water uptake was significantly affected 24 h after
treatment (HAT). In comparison with the PEG-free Induced Expression of GhPIP Genes in the Hydrated Roots
control (0/0-0), water uptake from the dehydrated of PRI Plants
roots under PRI (0/20-20) and either side of the roots
under the medium deficit irrigation (MDI; 10/10-10) The expression of the 13 GhPIP genes (six GhPIP1 and
and high deficit irrigation (HDI; 20/20-20) decreased seven GhPIP2) at 6 HAT is shown in Figure 2. Most of
significantly, whereas the water uptake from the hy- these genes were decreased by the PEG treatment. The
drated roots (0/20-0) increased considerably (Fig. 1A). expression of seven GhPIP genes (three GhPIP1 and
The total water uptake decreased by 15.2%, 62%, and four GhPIP2) in the dehydrated root side under PRI and
72.6% under PRI, MDI, and HDI, respectively, com- 11 GhPIP genes (four GhPIP1 and seven GhPIP2) in ei-
pared with the PEG-free control at 24 HAT. Under PRI, ther root side under MDI or HDI decreased more than
82.6% of the total water was absorbed from the hy- 2-fold (Fig. 2, A and B). In the hydrated root side of the
drated roots (Fig. 1A). plants under PRI, nine GhPIP genes (five GhPIP1 and
The L of the dehydrated roots under PRI and on ei- four GhPIP2) were up-regulated, of which four GhPIP
ther root side under MDI and HDI decreased signifi- genes (one GhPIP1 and three GhPIP2) were expressed
cantly from 3 to 24 HAT compared with that in the more than twofold compared with those in the control
PEG-free control, and there was no significant differ- (Fig. 2, A and B). The total expression levels of all the
ence in L among the three treatments (Fig. 1B). How- differentially expressed GhPIP1 and GhPIP2 genes in-
ever, the L of the hydrated root side under PRI was 2.2-, creased 1.8- and 2.9-fold in the hydrated root side but
2.64-, 2.8-, 3.5-, 3.6-, 3.4-, and 3.3-fold higher than that of decreased significantly in the dehydrated root side, and
the dehydrated root side and 1.06-, 1.29-, 1.5-, 1.9-, 1.7- similarly, the expression of these genes in either root
,1.6-, and 1.7-fold higher than that of either root side of side under MDI and HDI was also significantly de-
the PEG-free control at 1, 2, 3, 6, 9, 12, and 24 HAT, creased compared with that under the PEG-free control
respectively (Fig. 1B; Supplemental Fig. S1A). (Fig. 2, A and B).
Figure 1. Effects of PRI on water uptake,
root L, stomatal conductance (Cond),
and transpiration (Tr) under different
dehydration conditions. The two root
sides of each seedling were exposed to
different levels of PEG6000 (g g21) as
different treatments. A and B, Water
uptake by different root sides (A) at 24
HAT and L of the roots (B) in either root
side of the control (0/0-0), MDI plants
(10/10-10), and HDI plants (20/20-20)
as well as the hydrated root side (0/20-0)
and the dehydrated root side (0/20-20)
under PRI (0/20) at 3, 6, 9, 12, and 24
HAT. C and D, Cond (C) and Tr (D) of the
control (0/0), PRI (0/20), MDI plants (10/
10), and HDI plants (20/20). Values are
means 6 SD (n 5 6). Different letters in
A, C, and D indicate statistically signif-
icant differences (P , 0.05) after ANOVA
and the LSD tests. DW, Dry weight.
1662 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Leaf Jasmonate Mediates Water Uptake in Cotton
Figure 2. Changes in GhPIP gene ex-
pression in the roots under different de-
hydration conditions. Total GhPIP1 (A)
and GhPIP2 (B) expression at 24 HAT
(each GhPIP gene expression is shown
relative to the expression of ACTIN) and
expression of GhPIP1;5 (C), GhPIP2;2
(D), and GhPIP2;8 (E) are shown in ei-
ther root side of the control (0/0-0) or
under MDI (10/10-10) and HDI (20/20-
20) and the hydrated (0/20-0) and
dehydrated root side (0/20-20) under
PRI (0/20) as monitored by quantitative
real-time PCR (RT-qPCR) at 0, 3, 6, 9, 12,
and 24 HAT. Values are means 6 SD (n 5
12). Different letters in A and B indicate
statistically significant differences (P ,
0.05) based on ANOVA and the LSD tests.
GhPIP1;5 (EX164868.1), GhPIP2;2 (EX168689.1), and the contrary, no difference in Pro content was found
GhPIP2;8 (ES826026.1) were the three most abundant between the hydrated root under PRI and the root un-
GhPIP genes in this experiment (Fig. 2, A and B). der PEG-free control (Fig. 3C). In accordance with the
Analysis of the expression patterns of these three GhPIP Pro content, the expression of the Pyrroline 5 carboxylate
genes showed that their expression was down- synthetase (GhP5CS) gene, which is a key Pro biosyn-
regulated in the dehydrated roots under PRI and ei- thesis gene, was up-regulated in the dehydrated root
ther root side under MDI or HDI from 1 to 24 HAT side under PRI and in either root side under MDI and
(Fig. 2, C–E; Supplemental Fig. S1, B–D). On the con- HDI but was unaltered in the hydrated root side under
trary, these three GhPIP genes in the hydrated root side PRI (Fig. 3D).
under PRI were up-regulated and exhibited a gradually
increasing trend from 0 to 3 HAT (Fig. 2, C–E;
Supplemental Fig. S1, B–D). The expression of GhPIP2;2 Expression of JA Biosynthesis Genes and JA/JA-Ile
and GhPIP2;8 peaked at 3 and 6 HAT in the hydrated Contents under PRI
root side under PRI and then remained at 2.48- to 3.62-
fold and 2.73- to 3.51-fold higher than that in the PEG- The expression of JA biosynthesis genes (GhOPR11,
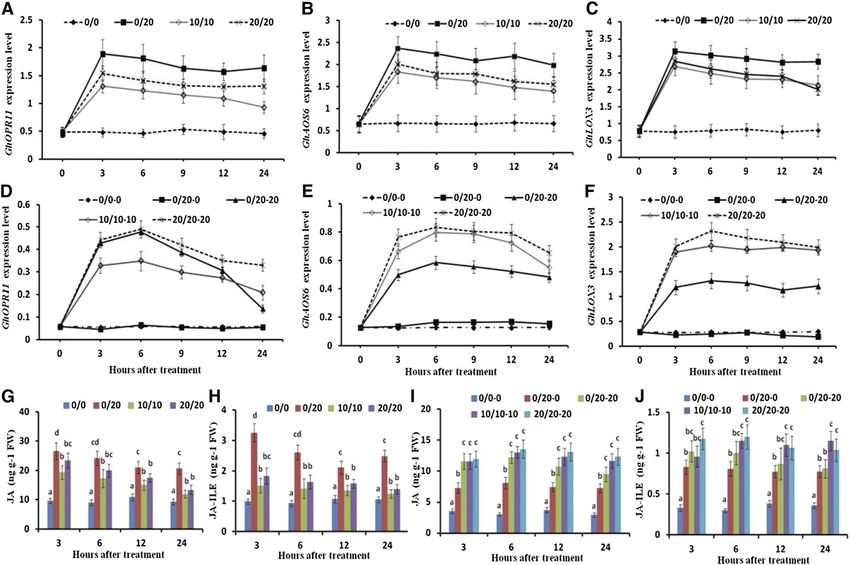
free control until 24 HAT. The expression of GhPIP1;5 GhAOS6, and GhLOX3) in the leaves under PRI was up-
peaked at 3 HAT and then decreased rapidly in the regulated and remained 2.86- to 4.25-fold, 2.51- to 4.27-
hydrated root side under PRI (Fig. 2, B–D). fold, and 3.01- to 4.76-fold higher than that in the leaves
of the control from 1 to 24 HAT, respectively (Fig. 4,
A–C; Supplemental Fig. S2, A–C). The expression
Root Water Content, Pro Content, and Root Water Potential of GhOPR11, GhAOS6, and GhLOX3 was also up-
regulated and remained at 1.84- to 3.08-fold, 1.85- to
The water content and water potential in the dehy- 3.26-fold, and 1.91- to 3.89-fold higher in the leaves of
drated root side under PRI and in either root side under MDI and at 2.46- to 3.21-fold, 2.33- to 3.96-fold, and
MDI or HDI decreased gradually following PEG treat- 2.52- to 4.15-fold higher in the leaves of HDI than that
ment, whereas these did not change in the hydrated in the PEG-free control from 1 to 24 HAT, respec-
root side under PRI compared with that in the PEG-free tively (Fig. 4, A–C; Supplemental Fig. S2, A–C). The JA
control (Fig. 3, A and B). The Pro content in the dehy- content in the leaves under PRI, MDI, and HDI in-
drated root side under PRI and either root side under creased consistently with the increased expression of
MDI and HDI increased following PEG treatment. On the three JA biosynthesis genes (Fig. 4, A–C and G). The
Plant Physiol. Vol. 180, 2019 1663
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
Figure 3. Water content, Pro content,
water potential, and GhP5CS expression
in the roots under different dehydration
conditions. Root water content (A), root
water potential (B) and root Pro content
(C) at 3, 6, 9, 12, and 24 HAT and the
expression of GhP5CS (D) at 24 HAT are
shown in either root side of the control
(0/0-0), MDI (10/10-10), and HDI plants
(20/20-20) as well as the hydrated (0/20-
0) and dehydrated root side (0/20-20)
under PRI (0/20). Values are means 6 SD
(n 5 6). Different letters in D indicate
statistically significant differences (P ,
0.05) based on ANOVA and the LSD tests.
JA content in the leaves under PRI was 2.99-, 3.77-, 2.76-, Effects of Silencing GhOPR11, GhAOS6, and GhLOX3 in
2.67-, 1.92-, and 2.22-fold higher than that in the leaves the Leaves under PRI
of the PEG-free control at 1, 2, 3, 6, 12, and 24 HAT,
respectively (Fig. 4G; Supplemental Fig. S2D). Simi- To determine if the leaf-biosynthesized jasmonate
larly, the JA-Ile content in the leaves under PRI, MDI, affected the JA-Ile content in the hydrated roots under
and HDI was higher than that in the leaves of the PEG- PRI, a tobacco rattle virus (TRV)-based virus-induced
free control (Fig. 4H; Supplemental Fig. S2E). The JA-Ile gene silencing (VIGS) vector was constructed and
content in the leaves under PRI was 2.75-, 3.55-, 3.27-, used to silence the GhOPR11, GhAOS6, and GhLOX3
2.8-, 1.96-, and 2.36-fold higher than that in the leaves of genes in the cotton leaves. The expression of GhOPR11,
the PEG-free control at 1, 2, 3, 6, 12, and 24 HAT, re- GhAOS6, and GhLOX3 in the leaves of their respective
spectively (Fig. 4H; Supplemental Fig. S2E). silenced plants decreased by 44.1%, 40.1%, and 41.9%,
The expression of GhOPR11, GhAOS6, and GhLOX3 respectively, compared with the VIGS-green fluores-
in the dehydrated root side under PRI and either root cent protein (GFP) control (nonsilenced plants; Fig. 5,
side under MDI and HDI was higher than that in either A–C). The JA-Ile content in the leaves of the VIGS-
root side of the PEG-free control (Fig. 4, E and F). The JA GhOPR11, VIGS-GhAOS6, and VIGS-GhLOX3 plants
content in the dehydrated root side under PRI and ei- decreased by 41.6%, 38.7%, and 35.5%, respectively,
ther root side under MDI and HDI increased consis- compared with the VIGS-GFP control (nonsilenced
tently with the increasing expression of the three JA plants) under PRI (Fig. 5D). Interestingly, the JA-Ile
biosynthesis genes (Fig. 4, E, F, and I). Interestingly, the content in the hydrated roots also decreased signifi-
JA content in the hydrated root side of the plants under cantly (by 37.3%, 37.5%, and 32.8%, respectively) in the
PRI was also 1.35-, 1.87-, 2.04-, 2.65-, 1.99-, and 2.47-fold VIGS-GhOPR11, VIGS-GhAOS6, and VIGS-GhLOX3
higher than that in either root side of the PEG-free plants compared with that in the hydrated roots of
control at 1, 2, 3, 6, 12, and 24 HAT, although the ex- the VIGS-GFP control (nonsilenced plants) under PRI
pression of GhOPR11, GhAOS6, and GhLOX3 was not (Fig. 5E). However, the expression of the GhOPR11,
significantly changed compared with that in either root GhAOS6, and GhLOX3 genes was not significantly
side of the PEG-free control (Fig. 4, F–I; Supplemental changed in the hydrated and dehydrated roots fol-
Fig. S2I). Similar to JA, the JA-Ile content in the dehy- lowing VIGS treatment (Supplemental Fig. S3, A and
drated root side under PRI and either root side under B). The expression of the GhPIP2;2 and GhPIP2;8 genes
MDI and HDI also increased (Fig. 4, H and J). The JA-Ile as well as L in the hydrated roots of the VIGS-GhOPR11,
content was 1.38-, 2.09-, 2.51-, 2.69-, 2.02-, and 2.15-fold VIGS-GhAOS6, and VIGS-GhLOX3 plants under PRI
higher in the hydrated root side and 2.65-, 4.13-, 3.09-, also decreased compared with the nonsilenced plants
3.34-, 2.27-, and 2.26-fold higher in the dehydrated root (Fig. 5, G–I). Silencing the GhOPR11, GhAOS6, and
side under PRI than that in either root side of the PEG- GhLOX3 genes decreased the expression of GhOPR11,
free control at 1, 2, 3, 6, 12, and 24 HAT (Fig. 4, H and J; GhAOS6, and GhLOX3 genes, and the expression of the
Supplemental Fig. S2, E and J). Also, the ratio of JA-Ile three JA biosynthesis genes and JA-Ile content in leaves
to JA was significantly higher in the hydrated root side of the PEG-free control and PRI plants were all signif-
(0.13–0.15) than in the dehydrated root side (0.08–0.09; icantly decreased compared with the VIGS-GFP con-
Fig. 4, I and J; Supplemental Fig. S2, I and J). trol; the JA-Ile content in either side of the roots of the
1664 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Leaf Jasmonate Mediates Water Uptake in Cotton
Figure 4. Expression of JA biosynthesis genes and JA and JA-Ile contents in the roots and leaves under different dehydration
conditions. A to C, Expression of JA biosynthetic genes (GhOPR11, GhAOS6, and GhLOX3) as monitored by RT-qPCR at
0, 3, 6, 9, 12, and 24 HAT in leaves of control (0/0), PRI (0/20), MDI (10/10), and HDI plants (20/20). D to F, Expression of JA
biosynthetic genes (GhOPR11, GhAOS6, and GhLOX3) as monitored by RT-qPCR at 0, 3, 6, 9, 12, and 24 HAT in either root
side of control (0/0-0), MDI (10/10-10), and HDI plants (20/20-20) as well as the hydrated (0/20-0) and dehydrated root side
(0/20-20) under PRI (0/20). G and H, JA and JA-Ile contents at 3, 6, 12, and 24 HAT in leaves of control (0/0), PRI (0/20), MDI
(10/10), and HDI plants (20/20). I and J, JA and JA-Ile contents at 3, 6, 12, and 24 HAT in either root side of control (0/0-0),
MDI (10/10-10), and HDI plants (20/20-20) as well as the hydrated (0/20-0) and dehydrated root side (0/20-20) under PRI
(0/20). Values are means 6 SD (n 5 12). Different letters in G to J indicate statistically significant differences (P , 0.05) based
on ANOVA and the LSD tests. FW, Fresh weight.
PEG-free control did not change significantly following Fig. S4B). Similarly, exogenous application of JA to the
VIGS treatment (Fig. 5, A–E). leaf also increased the ABA content in the hydrated
roots but had no effect on ABA content in the dehy-
drated roots (Fig. 6C). Foliar JA application to the leaf
Effects of Exogenous JA and Hypocotyl Ring-Barking increased the L in the hydrated roots by 20% and in-
under PRI creased the expression of the GhPIP2;2 and GhPIP2;8
genes in the hydrated roots by 30.4% and 29.9% under
Exogenous JA was foliar applied to the leaf or applied PRI at 24 HAT (Fig. 6, D–F). However, the JA-Ile content
to the dehydrated roots to determine the effect of ex- in the leaves and hydrated roots as well as the L in the
ogenous JA on L, GhPIP gene expression, and JA-Ile dehydrated roots did not change significantly, al-
content in the hydrated roots under PRI. Exogenous though exogenous JA applied to the dehydrated roots
application of JA to the leaf increased the expression increased the JA-Ile content in the dehydrated roots
of JA biosynthesis genes (GhOPR11, GhAOS6, and under PRI (Supplemental Fig. S5). On the contrary, the
GhLOX3) and the JA-Ile content in the leaf (Fig. 6A; JA-Ile content and the expression of the GhPIP2;2 and
Supplemental Fig. S4A). The JA-Ile content in the hy- GhPIP2;8 genes as well as the L in the hydrated roots
drated roots was also increased by the exogenous were significantly increased by the exogenous JA ap-
application of JA to the leaf, although the expression of plied to the hydrated roots (Fig. 7). Foliar-applied JA
the three JA biosynthesis genes was not affected in treatment increased the JA-Ile content in the leaf but did
the hydrated roots under PRI (Fig. 6B; Supplemental not significantly alter the expression of GhPIP2;2 and
Plant Physiol. Vol. 180, 2019 1665
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
Figure 5. Silencing of JA biosynthesis genes in the leaves decreased the L of the hydrated roots under PRI. The expression level of
GhOPR11 (A), GhAOS6 (B), and GhLOX3 (C), JA-Ile level in the leaves (D) and the roots (E), ABA content (F), the expression levels
of GhPIP2;2/EX168689.1 (G) and GhPIP2;8/ES826026.1 (H), and L (I) of the hydrated roots at 24 HAT are shown. Values are
means 6 SD (n 5 27). Different letters indicate statistically significant differences (P , 0.05) based on ANOVA and the LSD tests.
DW, Dry weight; FW, fresh weight.
GhPIP2;8 as well as L in the roots of the PEG-free control the plants under PRI to determine if the JA-Ile in the
(Fig. 6). Application of JA to one root side of the PEG- hydrated roots was transported from the leaves via the
free control increased the JA-Ile content, the expression phloem. The JA-Ile content, L, and the expression of
of the GhPIP2;2 and GhPIP2;8 genes, and the L of the JA GhPIP2;2 and GhPIP2;8 genes in the girdled root side
directly treated root side, but not on the other root side were similar to those in the nongirdled roots in the
(Fig. 7). PEG-free control plants (Fig. 7). However, ring-barking
Ring-barking was performed on either root side of increased the JA-Ile content in the leaf by 44.1% but
the PEG-free control and on the hydrated root side of decreased the JA-Ile content in the hydrated roots by
1666 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Leaf Jasmonate Mediates Water Uptake in Cotton
Figure 6. Effects of exogenous JA (200 mM) sprayed on leaves of the PEG-free control or applied to the hydrated roots under PRI.
The JA-Ile contents in the leaves (A) and roots (B), ABA contents (C), the expression levels of GhPIP2;2/EX168689.1 (D)
and GhPIP2;8/ES826026.1 (E), and root L (F) of differently treated roots at 24 HAT are shown. CS indicates the control
solution (containing 0.5% [v/v] ethanol). Values are means 6 SD (n 5 6). Different letters indicate statistically significant dif-
ferences (P , 0.05) based on ANOVA and the LSD tests. FW, Fresh weight.
64.6% under PRI (Fig. 7, A and B), although the ex- In addition, ring-barking was also performed on the
pression of the GhOPR11, GhAOS6, and GhLOX3 dehydrated root side of the plants under PRI to deter-
genes in the leaves and the hydrated roots of the mine if the JA-Ile in the dehydrated roots was produced
plants under PRI were not altered significantly locally or was shoot sourced. The results showed that
(Supplemental Fig. S5). Ring-barking decreased the ring-barking on the dehydrated root side did not sig-
expression of the GhPIP2;2 and GhPIP2;8 genes and nificantly change the JA-Ile content in the dehydrated
L in the girdled hydrated root side by 64.2%, 67.6%, roots under PRI irrespective of the foliar application of
and 46.5% at 24 HAT under PRI (Fig. 7, D–F). The JA- exogenous JA (Supplemental Fig. S6).
Ile content in the roots of the PEG-free control and the
hydrated root under PRI was increased by the exoge-
nous application of JA, and the expression of the Involvement of ABA in the Regulation of JA-Ile on L in the
GhPIP2;2 and GhPIP2;8 genes as well as the L in- Hydrated Roots under PRI
creased at 24 HAT following treatment (Fig. 7). The
decreased JA-Ile content, GhPIP2;2 and GhPIP2;8 gene The ABA content changed along with the JA-Ile
expression, and L in the hydrated root side by ring- content in the hydrated roots under PRI. It decreased
barking were partially compensated by exogenous JA or increased with JA-Ile content in the hydrated roots,
application to the hydrated root side under PRI decreasing in the VIGS-treated and girdled plants but
(Fig. 7). increasing with exogenous application of JA to the
Plant Physiol. Vol. 180, 2019 1667
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
Figure 7. Effects of ring-barking and JA (200 mM) applied to the hydrated side under PRI. The JA-Ile contents in the leaves (A) and
roots (B), ABA contents (C), the expression levels of GhPIP2;2/EX168689.1 (D) and GhPIP2;8/ES826026.1 (E), as well as the root
L (F) of the different roots at 24 HAT are shown. RB indicates ring-barking; VC indicates vertical cuttings as the control by re-
moving pieces of bark (5 mm 3 2 mm). Values are means 6 SD (n 5 6). Different letters indicate statistically significant differences
(P , 0.05) based on ANOVA and the LSD tests. FW, Fresh weight.
leaves or hydrated roots (Figs. 5D, 6C, and 7C). Exog- the hydrated root side (Wakrim et al., 2005; Li et al.,
enous ABA and its specific inhibitor fluridone were 2007; Wang et al., 2010); however, the underlying
applied to the hydrated root side of the plants under mechanisms of the increased water uptake remain un-
PRI to determine if the JA-Ile content, L, and PIP genes clear. Using a grafted split-root PRI system, we con-
in the hydrated roots were induced by ABA. Neither firmed that shoot-sourced JA/JA-Ile, as a long-distance
ABA nor fluridone significantly affected the JA-Ile signal, positively mediates water uptake from the hy-
content and expression levels of the GhPIP2;2 and drated roots of cotton under PRI. Not only do our
GhPIP2;8 genes (Fig. 8, A–C). Exogenous ABA in- findings further elucidate the role of JA/JA-Ile as a
creased L in the hydrated root side under PRI by 36.9%, long-distance signaling molecule in the regulation of
but L was decreased by fluridone (Fig. 8D). water uptake, they also provide new and robust evi-
dence to support the involvement of JA in water uptake
under PRI.
Since L is influenced by stomatal conductance and
DISCUSSION
transpiration, we measured these parameters and
PRI is a promising water-saving irrigation technique found that the stomatal conductance of the leaves de-
with many advantages over traditional full-water or creased significantly and the transpiration rate decreased
deficit irrigation approaches (Kang and Zhang, 2004; by 23.9%, causing the water uptake (consumption) by the
Costa et al., 2007; Kirda et al., 2007). In addition to de- whole plant to decrease by 15.2% under PRI compared
creasing transpiration, it increases water uptake from with the PEG-free control at 24 HAT. We also showed that
1668 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Leaf Jasmonate Mediates Water Uptake in Cotton
Figure 8. Effects of ABA (10 mM) and fluridone (10 mM) applied to the hydrated side under PRI. The JA-Ile content (A), the ex-
pression levels of GhPIP2;2/EX168689.1 (B) and GhPIP2;8/ES826026.1 (C), as well as the L (D) of the different roots at 24 HAT are
shown. Values are means 6 SD (n 5 6). Different letters indicate statistically significant differences (P , 0.05) based on ANOVA
and the LSD tests. FW, Fresh weight.
the water uptake from the dehydrated root side under and GhPIP2;8 were the three most abundant GhPIP
PRI decreased by 70.2% but increased by 41.3% in genes of the 13 measured GhPIPs (Fig. 2, A and B). The
the hydrated roots at 24 HAT compared with either expression of GhPIP1;5 in the hydrated root side under
root side of the control (Fig. 1A). This is consistent PRI increased and peaked at 3 HAT and then decreased
with previous reports that substantially more water is rapidly to levels comparable to that on either side of the
absorbed from the hydrated roots than the dehydrated PEG-free control at 24 HAT (Fig. 2, B–D), whereas the
roots under PRI (English and Raja, 1996; Kang et al., expression of GhPIP2;2 and GhPIP2;8 remained at 2.48-
2003; Mingo et al., 2004; Hu et al., 2011). and 2.73-fold higher than on either side of the PEG-free
The uptake of water by the roots is largely mediated control at 24 HAT (Fig. 2, C–E). In contrast to our
by PIPs, which represent the most abundant aqua- findings, a previous study showed that PIP1 abundance
porins in the plant plasma membrane (Vandeleur increased after 24 h in the hydrated root portion under
et al., 2005; Maurel et al., 2008). Generally, PIP2s have partial root-zone drying in the riparian species M.
high water permeability, while PIP1s have little or no argentea, which was followed by a decline in PIP2s
measurable permeability (Chaumont et al., 2000; (McLean et al., 2011). The difference in PIP aquaporin
Kaldenhoff et al., 2008). Some studies have indicated response to drought stress between that previous report
that the isomeric isotetramer formed by the interaction and this study might be due to the variations in plant
of PIP1 and PIP2 significantly improved the total per- species or stress levels (Šurbanovski et al., 2013). The
meability (Fetter et al., 2004; Mahdieh et al., 2008; results of our study suggested that the increased L and
Vandeleur et al., 2009). However, a reduction in PIPs water uptake could mainly be attributed to the elevated
under drought conditions was observed in the roots of expression of GhPIP2;2 and GhPIP2;8 genes in the hy-
olive (Olea europaea) and tobacco (Vandeleur et al., 2005; drated root side under PRI.
Secchi et al., 2007; Mahdieh et al., 2008). In our study, L There is evidence that leaf-to-root signals may regu-
and water uptake increased in the hydrated root side late root L by regulating the quantity or activity of PIPs
but decreased in the dehydrated root side under PRI, (Vandeleur et al., 2005; Kudoyarova et al., 2011;
which was consistent with the increased expression Sakurai-Ishikawa et al., 2011). A rapid decrease in tur-
of GhPIP1 and GhPIP2 genes in the hydrated root side gor in root cortex cells following shoot topping resulted
and decreased expression of these genes in the dehy- in the regulation of root L by xylem-mediated hydraulic
drated root side (Figs. 1B and 2). GhPIP1;5, GhPIP2;2, signals from the leaves (Vandeleur et al., 2014). Osmotic
Plant Physiol. Vol. 180, 2019 1669
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
stress can reduce root water content and elevate the were used to further determine if the increased JA-Ile
levels of the compatible osmolyte Pro, which can alter content in the hydrated root side was transported
cell turgor pressure (Wilson et al., 2014). To determine from the leaves under PRI in this study. VIGS treat-
whether the hydrated root of PRI suffered from osmotic ment down-regulated the expression of the GhOPR11,
stress, the Pro content, water content, and water po- GhAOS6, and GhLOX3 genes and decreased the JA-Ile
tential of the roots under different dehydration condi- content in the leaves under PRI (Fig. 5, A–D). The JA-Ile
tions were measured (Fig. 3). The results indicated that content in the hydrated roots also decreased signifi-
treatment with PEG increased the Pro content but de- cantly, although the expression of the three JA biosyn-
creased the root water content and root water potential thesis genes was not changed considerably in the
in either root side of the plants under MDI and HDI, or hydrated roots after silencing the GhOPR11, GhAOS6,
the dehydrated root side under PRI, but had little effect and GhLOX3 genes in the leaf (Fig. 5E; Supplemental
on the hydrated root side under PRI (Fig. 3, A–C). A Fig. S3). On the contrary, exogenous foliar application
previous study found that water deficit also increased of JA increased the JA-Ile content in the leaves and the
the expression of GhP5CS (Silva-Ortega et al., 2008). hydrated root under PRI despite the expression of the
The expression of GhP5CS in the dehydrated root side three JA biosynthesis genes being unaffected by exog-
under PRI and either root side under MDI and HDI was enous JA (Fig. 6; Supplemental Fig. S5). These results
up-regulated but was unaltered in the hydrated root strongly indicated that the increased JA-Ile content in
side under PRI at 24 HAT (Fig. 3D). This suggested that the hydrated root side was transported from the leaves
the PRI treatment did not cause osmotic stress in the under PRI.
hydrated root side, and the increased L and GhPIP ex- To determine if the leaf-sourced JA-Ile was trans-
pression is thus likely induced by other signals rather ported to the root via the phloem, the phloem of the
than hydraulic signals. hydrated root side and either root side of the control
Leaf-to-root signals transported via the phloem reg- plants were girdled. Ring-barking on the hydrated root
ulate root growth and induce stress responses side increased the JA-Ile content in the leaves and de-
(Macháčková et al., 1992; Zhang and Baldwin, 1997; creased the JA-Ile content in the hydrated roots but did
Ljung et al., 2002; Shah, 2009; Kudoyarova et al., 2011). not affect the expression of the three JA biosynthesis
Zhang and Baldwin (1997) found that wound-induced genes either in the leaves or roots (Fig. 7, A and B;
JA in the leaves was transported to the roots and Supplemental Fig. S5). This indicated that the increased
increased in de novo nicotine synthesis in the roots accumulation of JA-Ile in the hydrated roots should be a
following leaf wounding. In this study, JA-Ile accu- result of its synthesis in the leaves and subsequent
mulation exhibited a similar pattern to JA accumulation transportation to the roots via the phloem.
and the expression of JA biosynthesis genes GhOPR11, In order to determine whether the accumulated JA in
GhAOS6, and GhLOX3 under drought stress. The in- the dehydrated root side was produced locally or was
creased JA/JA-Ile content in the roots under MDI and leaf sourced, the phloem of the dehydrated root side
HDI, as well as in the leaves and the dehydrated root was also girdled. The JA (JA/JA-Ile) content increased
side under PRI, was possibly attributed to the increased consistently with the increased expression of JA bio-
expression of the JA biosynthesis genes GhOPR11, synthesis genes in the dehydrated roots of the PRI
GhAOS6, and GhLOX3 (Fig. 4). The JA/JA-Ile content plants (Fig. 4, D–F, I, and J), but it was not affected by
was also increased, although the three JA biosynthesis the VIGS silencing or the foliar application of exoge-
genes were not up-regulated in the hydrated root side nous JA (Figs. 5D and 6B). It was also noted that ring-
under PRI (Fig. 4, D–F, I, and J), suggesting a possible barking on the dehydrated root side did not affect the
transportation of JA/JA-Ile from the leaves or the JA-Ile content in the dehydrated roots under PRI re-
dehydrated root side to the hydrated root side under gardless of the foliar application of exogenous JA
PRI. Exogenous JA added to the dehydrated root side (Supplemental Fig. S6). The overall results indicated
increased its JA-Ile content but did not affect the JA-Ile that JA in the dehydrated roots was produced locally.
content in the leaves and the hydrated root side The role of jasmonate in the water stress response,
(Supplemental Fig. S4). These results indicated that the particularly as a long-distance signal, is of interest.
JA-Ile content in the leaves and the hydrated root side Vandeleur et al. (2014) concluded that foliar-applied
was not affected by the JA-Ile content in the dehydrated MeJA is not involved in water deficit response,
root side, and the increased JA-Ile content in the hy- whereas Sánchez-Romera et al. (2014) discovered that
drated root side appears to have been transported from MeJA increased the L of P. vulgaris, S. lycopersicum, and
the leaves under PRI. Arabidopsis roots in the absence of osmotic stress.
Gao et al. (2011) successfully inoculated Agro- Similarly, using grafted S. lycopersicum, De Ollas et al.
bacterium tumefaciens cultures carrying TRV-VIGS vec- (2018) showed that root-derived JA is involved in the
tors into cotton cotyledons. This VIGS method is shoot response under water deficit. In our study, the
limited to the silencing of target genes in leaves. The expression of GhPIP2;2 and GhPIP2;8 as well as root L
plant silenced by VIGS-GhCLA1 (cotton cloroplastos increased or decreased along with the increase or de-
alterados1) exhibited whitened parts only above the crease in JA-Ile content in the hydrated roots of the
cotyledon node (Supplemental Fig. S7). Therefore, plants under PRI. Hypocotyl ring-barking on the
VIGS experiments and exogenous foliar-applied JA phloem of the hydrated root side and VIGS silencing of
1670 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Leaf Jasmonate Mediates Water Uptake in Cotton
JA-biosynthesis genes in the leaves decreased the JA-Ile JA-Ile content in the roots of the PEG-free control did
content in the hydrated roots and then down-regulated not change significantly (Figs. 5 and 6). Ring-barking on
the GhPIP2;2 and GhPIP2;8 genes and decreased the either root side of the control plants did not affect the
root L (Figs. 5, E and G–I, and 7, B and D–F). Con- JA-Ile content in the leaves and roots, nor did it evoke a
versely, the exogenous application of JA to the girdled reduction in root L (Fig. 7, A and B). This corroborates
hydrated roots or to the leaves without ring-barking previous research (Vandeleur et al., 2014) where the
increased the JA-Ile content in the hydrated roots, application of JA to the shoots had no effect on the L of
thereby up-regulating the GhPIP2;2 and GhPIP2;8 genes soybean (Glycine max) in lanolin paste. In a more recent
and increasing the root L (Fig. 6, B and D–F). All of these study by Sánchez-Romera et al. (2016), it was demon-
results suggest that JA-Ile plays a key role in increasing strated that L did not change in P. vulgaris under well-
the root L by up-regulating the GhPIP genes in the hy- watered conditions following MeJA application, but
drated roots under PRI. MeJA increased L under drought conditions. It seems
Plants need to be equipped for survival in a complex that the involvement of jasmonate in water uptake
environment where they are continuously challenged might thus be dependent on abiotic stress. Our results
by biotic/abiotic stresses (Santino et al., 2013). Plant further indicated that without water deficit stress on
hormones and their cross talk participate in various one side of the roots, there would be no transport of
stress responses. A previous study showed that MeJA jasmonate from the leaves to the wet side of the roots.
could increase the synthesis of ABA (Adie et al., 2007), This suggests that water stress on one side of the
which typically increases L (Aroca, 2006; Mahdieh and root system triggers or induces the entire process, the
Mostajeran, 2009). In our study, the ABA content in- mechanism of which requires further study.
creased in the hydrated roots under PRI and increased When directly exposed to water deficit stress, the root
or decreased following the increase or decrease in JA-Ile system rapidly accumulates chemical substances and
content after VIGS silencing of JA biosynthesis genes, signaling molecules, including ABA and jasmonate,
exogenous JA application, or hypocotyl ring-barking coinciding with a decrease in stomatal conductance as
(Figs. 5F, 6C, and 7C). Our findings demonstrated an early avoidance response to drought stress (Harb
that ABA changed along with the change in JA-Ile et al., 2010; Wasternack and Strnad, 2019). This is fol-
content in the hydrated roots under PRI. The L of the lowed by the adjustment of the cell wall and a late stage
hydrated root side was also increased by exogenous of new homeostasis with reduced plant growth (Harb
ABA and decreased by its inhibitor fluridone. How- et al., 2010). In this process, reduced L and water uptake
ever, ABA or fluridone did not affect the expression of in the stressed roots constitute common phenomena
the GhPIP2;2 and GhPIP2;8 genes (Fig. 8, B–D). Hose (Vandeleur et al., 2005, 2014; Hu et al., 2011). In our
et al. (2000) speculated that ABA affects the expression study, it was noted that the expression of GhPIP genes
or activity of aquaporins. Wan et al. (2004) surmised and L in the dehydrated root side under PRI decreased,
that ABA may bind to aquaporins and then reduce the while the jasmonate content increased in the dehy-
activation energy in order to change from closed to drated root side (Fig. 4, I and J). When exogenous JA
open water channel states, but in a more recent study, it was applied to the roots, the root L of the PEG-free
was reported that ABA increases the water transport control increased, whereas the L of the dehydrated
activity of PIP2;1 through phosphorylation of PIP2;1 by roots of PRI did not change significantly (Fig. 7F;
Open stomata1 (Grondin et al., 2015). Zhang et al. Supplemental Fig. S4C). These results do not seem to
(2019) found that lectin receptor-like kinases under support our hypothesis that jasmonate accumulation
ABA induction could regulate aquaporin activity. In improves L and water uptake. However, they do sug-
our study, the disparity between L and the expression gest the potential diversity of jasmonate responses to
of the GhPIP2;2, and GhPIP2;8 genes following ABA drought stress and also imply that root jasmonate may
and fluridone treatment suggests that the increased L be affected by both the root-zone environment and the
may be regulated by other mechanisms, such as the jasmonate components (ratio of JA-Ile to JA). Via
reduction in PIP protein activation energy directly in- jasmonate signaling, the cotton plants improved the L
duced by ABA or the increased PIP protein activity of the hydrated root side rather than the dehydrated
indirectly induced by ABA. The expression of PIPs root side in the PRI system. Based on our findings, we
significantly decreased in the hydrated root under PRI suggest that the accumulation of jasmonate in the
at 12 HAT (Fig. 2, C–E), whereas L did not change dehydrated root side was probably a direct response to
significantly (Fig. 1B), which might be due to the in- drought stress, whereas the leaf-sourced jasmonate that
creased PIP protein activity induced by ABA. Regard- accumulated in the hydrated root was associated with
less of the interpretation, the interaction of JA-Ile as a the regulation of water uptake. Moreover, the different
long-distance signal along with ABA in the regulation roles of the JAs in the hydrated and dehydrated sides
of water uptake in the hydrated root side is worth might be due to the cross talk of JAs with other sig-
further investigation under PRI. naling pathways synergistically or antagonistically ei-
Notably, the JA-Ile content in the leaves of the PEG- ther on the hydrated or dehydrated root side that
free control was decreased by the silencing of the JA were involved in the regulation of hydraulic conduc-
biosynthesis genes in the VIGS experiments and was tance, just like the cross talk between salicylic acid and
increased by exogenous foliar-applied JA, whereas the JA pathways can be either mutually antagonistic or
Plant Physiol. Vol. 180, 2019 1671
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Luo et al.
synergistic in response to biotic stresses (Santino et al., Seedlings with uniform split roots were selected, and each root portion was
placed into one side of the boxes. PEG6000 was used to simulate drought stress.
2013). We also found that the ratio of JA-Ile to JA was sig- The two root portions of each seedling were concurrently exposed to different
nificantly higher in the hydrated root side (0.15) than in the PEG6000 (g g21) concentrations. The osmotic potentials of the nutrient solutions
dehydrated root side (0.09). As JA-Ile is the active mole- containing 0%, 10%, (w/v) and 20% (w/v) PEG6000 were 20.05, 20.7, and
cule of the jasmonate family, a lower ratio of JA-Ile to JA 21.02 MPa, respectively. The two root portions under 0% and 20% PEG6000
might be another possible reason for the decrease in hy- treatment were denoted as PRI (0/20). The treatment with the two root portions
in 0% PEG6000 was denoted as the PEG-free control (0/0). Treatments with
draulic conductivity in the dehydrated root side under PRI. both root portions in 10% (w/v) and 20% (w/v) PEG6000 were respectively
denoted as MDI (10/10) and HDI (20/20; Supplemental Fig. S8). In the PRI
treatment (0/20), the PEG6000-free side was denoted as the wet (hydration)
CONCLUSION side (0 or 0/20-0), while the 20% (w/v) PEG6000 side was denoted as the dry
(dehydration) side (0/20-20).
PRI increased the JA/JA-Ile content in the leaves by
increasing the expression of the GhOPR11, GhAOS6, Exogenous JA Treatment
and GhLOX3 genes. The leaf-sourced JA/JA-Ile was
then transported to the hydrated roots via the phloem, Five milliliters of exogenous JA (200 mM, dissolved in 0.5% [v/v] ethanol) was
foliar applied to the plants using a hand sprayer, whereas an equivalent volume
thereby increasing the ABA content. As a result, the L of double distilled water containing 0.5% (v/v) ethanol was sprayed as the
and water uptake increased in the hydrated root side control. As for the application of JA in root treatments, exogenous JA (200 mM,
through up-regulation of the GhPIP2;2 and GhPIP2;8 dissolved in 0.5% [v/v] ethanol) was supplied to the nutrient solutions, and the
genes under PRI. Leaf-sourced JA/JA-Ile, as a long- nutrient solution containing 0.5% (v/v) ethanol was used as the control. After 3,
6, 9, 12, and 24 h of treatment, the roots from both sides and the leaves of six
distance signal, positively mediated water uptake in seedlings from each treatment were sampled, washed three times with distilled
the hydrated root side of cotton under PRI. The results water, blotted dry with filter paper, frozen in liquid nitrogen, and then stored at
have improved our understanding of the role of JA/JA- 280°C.
Ile as a long-distance signaling molecule in the regula-
tion of water uptake. In future research, the different Measurements of Root Relative Water Contents, Water
roles of JA/JA-Ile in the hydrated and dehydrated root Uptake, Stomatal Conductance, and Transpiration
sides under PRI should be studied.
Root relative water contents (RRWC) were determined from 0.5 to 1 g of
sample collected in the morning (9–10 AM). The roots from both sides of six
seedlings of each treatment were washed clean, surface dried thoroughly on
MATERIALS AND METHODS paper towels, weighed (wi), immersed into distilled water at 4°C overnight,
weighed again (wf), freeze dried in a vacuum freezing machine (Christ), and
Establishment of a Split-Root System then weighed a third time (wd). RRWC was calculated as: RRWC [%] 5 100
(wi – wd)/(wf – wd).
A commercial cotton (Gossypium hirsutum cv ‘K836’), which was developed
To measure water uptake, the roots of the grafted plants were positioned in
by our lab, was used in the experiments. One kilogram of cotton fuzz seeds was
the split-root box containing 2 L of nutrient solution in each compartment.
placed in a plastic container, treated with 100 mL of concentrated sulfuric acid,
Three grafted plants were positioned into one split-root box. Six split-root boxes
and stirred for 1 to 2 min for acid delinting. The acid-delinted seeds were then
containing equal volumes of nutrient solutions without plants served as con-
washed six times and sun dried for 2 to 3 d. The seeds were sown at 2 cm depth
trols to eliminate natural evaporation. The boxes and nutrient solutions were
in plastic boxes (60 cm 3 45 cm 3 15 cm) containing sterilized wet sand. The
measured 24 HAT, and the water uptake was determined. The water uptake of
boxes were placed in a greenhouse with a light/dark regime of 16/8 h, a light
one split-root box was denoted as one biological replicate, and six boxes were
intensity of 400 mmol m22 s21 photosynthetically active radiation, and a tem-
set up for each treatment.
perature of 30°C 6 2°C. Seedlings were thinned to 100 plants per box at full
Stomatal conductance and transpiration of the second leaf on the main stem
emergence. At the two-true-leaf stage, uniform seedlings were carefully re-
from the terminus were measured using an LI-6400 portable photosynthesis
moved from the sand and washed with distilled water. Split-root systems were
system (Li-Cor) at 9 to 11 AM. Six plants were measured from each treatment.
established by grafting with these seedlings as described by Kong et al. (2012,
2017). Briefly, a slash (/)-shaped incision was made with a blade on the hy-
pocotyl 2 cm below the two cotyledons, leaving about one-third of the hypo- Pro Content
cotyl tissues intact. The top of the rootstock was cut to form a deep triangle (△)
at the same position of the hypocotyl from another seedling. The triangle section Pro was assayed using the ninhydrin assay according to the modified pro-
was then inserted into the slash incision of the plant and firmly wrapped with cedure described by Bates (1973). Roots were powdered under liquid nitrogen
Parafilm. Grafted seedlings were transferred into plastic boxes containing and mixed with 100 mM 5-sulfosalicylic acid dihydrate buffer using 1 mL of
aerated nutrient solution and sprayed with water to prevent wilting. The nu- buffer per 100 mg of tissue powder and extracted in boiling water for 10 min.
trient solution was topped up with deionized water as required and renewed After cooling to room temperature, 2 mL of supernatant was incubated with
weekly. The solution consisted of 1.25 mM Ca(NO3)2, 1.25 mM KNO3, 0.5 mM 2 mL of acetic acid and 3 mL of 150 mM ninhydrin buffer in boiling water for
MgSO4, 0.25 mM NH4H2PO4, 0.05 mM EDTA-FeNa, 10 mM H3BO3, 0.5 mM 40 min. The absorbance (520 nm) was quantified using a spectrophotometer
ZnSO4, 0.1 mM CuSO4, 0.5 mM MnSO4, and 0.0025 mM (NH4)6Mo7O24 and was (TU-1901; Purkinje). Six roots from each treatment were measured.
adjusted to pH 6 with KOH. When a new leaf emerged from the grafted
seedling 2 weeks after grafting, the Parafilm was removed. Grafted seedlings
with two uniform split-root systems were selected and allowed to grow in a Root Water Potential and Root L
greenhouse under a 14/10-h (light/dark) photoperiod at 30°C/26°C and rela-
Root water potential and root L in both the split-root compartments of the
tive humidity of 60%/80% for 30 d. Nutrient solutions were renewed daily.
box were measured by pressurizing the roots in a pressure chamber (PMS 670)
Strong seedlings with uniform split roots were selected for further study.
as described previously (Sánchez-Romera et al., 2014). All measurements were
conducted between 11 AM and 3 PM. The stem of each side of the split-root
PRI Treatment system was cut with a blade 1 cm below the grafted position. The root sys-
tems were sealed into a pressure chamber with the cut stump protruding
Plastic boxes (26 cm 3 16 cm 3 15 cm) were used to establish the PRI through the lid and the roots surrounded by the nutrient solution containing
treatment. The boxes were divided into two equal parts with a plastic board in PEG and other chemicals, where applicable, according to the different treat-
the middle of their inner space to restrict water flow between the two sides. ment requirements. Pressure was applied slowly when the first drop of liquid
1672 Plant Physiol. Vol. 180, 2019
Downloaded on February 9, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































