Molecular and cytological analysis of widely-used Gal4 driver lines for Drosophila neurobiology - BMC Genetics
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96
https://doi.org/10.1186/s12863-020-00895-7
RESEARCH Open Access
Molecular and cytological analysis of
widely-used Gal4 driver lines for Drosophila
neurobiology
Anna A. Ogienko1†, Evgeniya N. Andreyeva1†, Evgeniya S. Omelina1, Anastasiya L. Oshchepkova1,2 and
Alexey V. Pindyurin1,3*
From 11th International Young Scientists School “Systems Biology and Bioinformatics” – SBB-2019
Novosibirsk, Russia. 24-28 June 2019
Abstract
Background: The Drosophila central nervous system (CNS) is a convenient model system for the study of the
molecular mechanisms of conserved neurobiological processes. The manipulation of gene activity in specific cell
types and subtypes of the Drosophila CNS is frequently achieved by employing the binary Gal4/UAS system.
However, many Gal4 driver lines available from the Bloomington Drosophila Stock Center (BDSC) and commonly
used in Drosophila neurobiology are still not well characterized. Among these are three lines with Gal4 driven by
the elav promoter (BDSC #8760, #8765, and #458), one line with Gal4 driven by the repo promoter (BDSC #7415),
and the 69B-Gal4 line (BDSC #1774). For most of these lines, the exact insertion sites of the transgenes and the
detailed expression patterns of Gal4 are not known. This study is aimed at filling these gaps.
Results: We have mapped the genomic location of the Gal4-bearing P-elements carried by the BDSC lines #8760,
#8765, #458, #7415, and #1774. In addition, for each of these lines, we have analyzed the Gal4-driven GFP
expression pattern in the third instar larval CNS and eye-antennal imaginal discs. Localizations of the endogenous
Elav and Repo proteins were used as markers of neuronal and glial cells, respectively.
Conclusions: We provide a mini-atlas of the spatial activity of Gal4 drivers that are widely used for the expression
of UAS–target genes in the Drosophila CNS. The data will be helpful for planning experiments with these drivers
and for the correct interpretation of the results.
Keywords: Drosophila, Gal4/UAS, CNS, Neurons, Glia, Driver, elav-Gal4, elavC155, Gal4repo, 69B-Gal4
* Correspondence: a.pindyurin@mcb.nsc.ru
†
Anna A. Ogienko and Evgeniya N. Andreyeva contributed equally to this
work.
1
Institute of Molecular and Cellular Biology, Siberian Branch of RAS,
Novosibirsk 630090, Russia
3
Novosibirsk State University, Novosibirsk 630090, Russia
Full list of author information is available at the end of the article
© The Author(s). 2020 Open Access This article is licensed under a Creative Commons Attribution 4.0 International License,
which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give
appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if
changes were made. The images or other third party material in this article are included in the article's Creative Commons
licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons
licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the
data made available in this article, unless otherwise stated in a credit line to the data.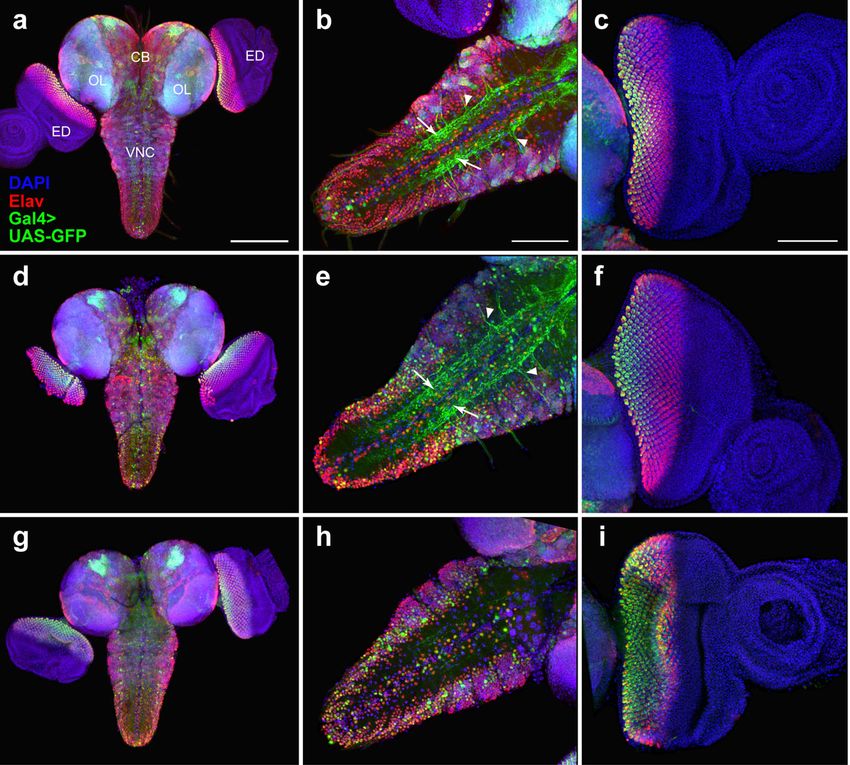
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 2 of 11
Background gene (X:523350; here and afterwards, coordinates are
Genetic and molecular studies in Drosophila can provide from Release 6 of the D. melanogaster genome assembly
valuable insight into the pathogenesis of human diseases, [17]); this insertion is also known as the elavC155 allele
due to the high conservation of key molecular mecha- [18]. Some differences between patterns/strength of ac-
nisms underlying biological processes in metazoans, and tivity of these drivers were previously described in epi-
to the fact that about 77% human disease genes have thelial tissues and embryonic CNS [19, 20]. Gal4 drivers
orthologues in Drosophila [1]. Thus, targeted expression from all the three lines are used to drive the expression
of mutant human disease genes in Drosophila can recap- of different genes and RNAi constructs in neuronal cells
itulate relevant aspects of the pathology [2, 3]. Such ex- in CNS [21, 22].
pression is frequently achieved by employing the binary In line #7415, the P{Gal4} element, presumably an
Gal4/UAS system, which allows the control of the activity enhancer-trap Gal4 construct, is inserted somewhere in
of the gene of interest in a spatiotemporal-restricted man- the reversed polarity (repo) locus (3R:18236194..18239604;
ner [4]. The specificity of Gal4 expression allows cellular the insertion is referred to as Gal4repo) and the Gal4 ex-
resolution that is particularly important for studies on the pression pattern matches that of the endogenous Repo
nervous system, in which there is a need to manipulate protein [23], which is specifically expressed in glia during
the activity of small sets of neurons or glia cells [5–8]. development [24]. Contrary to the elav-Gal4 drivers, no
Anatomically, the Drosophila larval CNS comprises the activity of Gal4repo was detected in larval epithelial tissues
brain and the ventral nerve cord (VNC), which are the [19]; the highest level of repo expression in the larval CNS
equivalents of the vertebrate brain and spinal cord, re- was observed in the optic lobes [24]. The Gal4repo driver is
spectively. The brain is composed of two main regions, commonly used to manipulate gene expression in glial
the optic lobes located at the lateral surface of the brain cells in the CNS [25, 26].
hemispheres and the medially-located central brain [9]. Line #1774 carries the P{GawB} element inserted
Histologically, the CNS can be subdivided into the neur- somewhere on the third chromosome. This insertion is
onal cell cortex, where all neuronal cell bodies reside, and known as 69B or 69B-Gal4 [16] and it drives expression
the neuropil, where axons and dendrites project to form of the reporter genes semi-ubiquitously. In third instar
neural circuits [10]. Currently, a lot of different Gal4 larvae, 69B-Gal4 activity was detected in the CNS, imagi-
drivers are available for targeted expression of reporter nal discs, ring gland, epidermis and testes [16, 27–29].
genes in distinct CNS cell types and subtypes. The choice In the CNS, the expression of 69B-Gal4 seems to be
of the proper driver for each experiment is crucial, since it restricted to non-glial cells [30].
can influence the results and their interpretation. How- Our previous experience indicates that transgenic
ever, for several widely-used Gal4 driver lines, the exact Drosophila lines often carry more transposon constructs
genomic location of the Gal4-bearing transgene and the than commonly assumed, and that these insertions can
precise pattern of Gal4 expression are still unknown. This potentially influence the experimental outcome [31]. In
information could be very useful (i) to learn whether the the present study, we identified the precise genomic lo-
insertion of the Gal4 transgene disrupts the expression of cations of the Gal4 transgenes in lines #8760, #8765,
an endogenous gene potentially relevant for the analysis #7415, and #1774, and confirmed the previous mapping
undertaken and (ii) for combining the driver with other of elavC155 in line #458. We also carefully characterized
transgenes or mutations, so as to construct the desired the expression pattern of a UAS-GFP reporter gene in-
genotype. Thus, we decided to identify the insertion sites duced by the Gal4 drivers of these five lines in the CNS
of the transgenes and characterize Gal4 expression pat- and eye-antennal imaginal discs of third instar larvae.
terns in larval CNS for the following five commonly used We found that the enhancer-trap Gal4 drivers from lines
CNS-specific driver lines available from BDSC: #8760, #458 and #7415 elicit the expression patterns highly
#8765, #458, #7415, and #1774. comparable to those of the endogenous Elav and Repo
In three lines, the Gal4 expression seems to be primar- proteins, respectively. In contrast, the elav-Gal4 drivers
ily under the control of the regulatory sequences of the from lines #8760 and #8765 match the Elav expression
embryonic lethal, abnormal vision (elav) gene, which is pattern only partially. In addition, our data support the
active in postmitotic neurons throughout development view that the 69B-Gal4 driver is primarily active in non-
[11–13]. The lines #8760 and #8765 carry the P{Gal4- glial cell types.
elav.L} element [14] somewhere on the third and second
chromosomes, respectively. This P-element-based trans- Results
poson contains a 3.5-kb genomic fragment including the Molecular characterization of the chromosomes bearing
elav promoter region [15] upstream of the Gal4 coding the Gal4 drivers
sequence. In line #458, an enhancer-trap Gal4 construct, We first evaluated the number of P-element-based con-
P{GawB} [16], is inserted in the promoter of the elav structs harbored by the chromosomes Chr3#8760,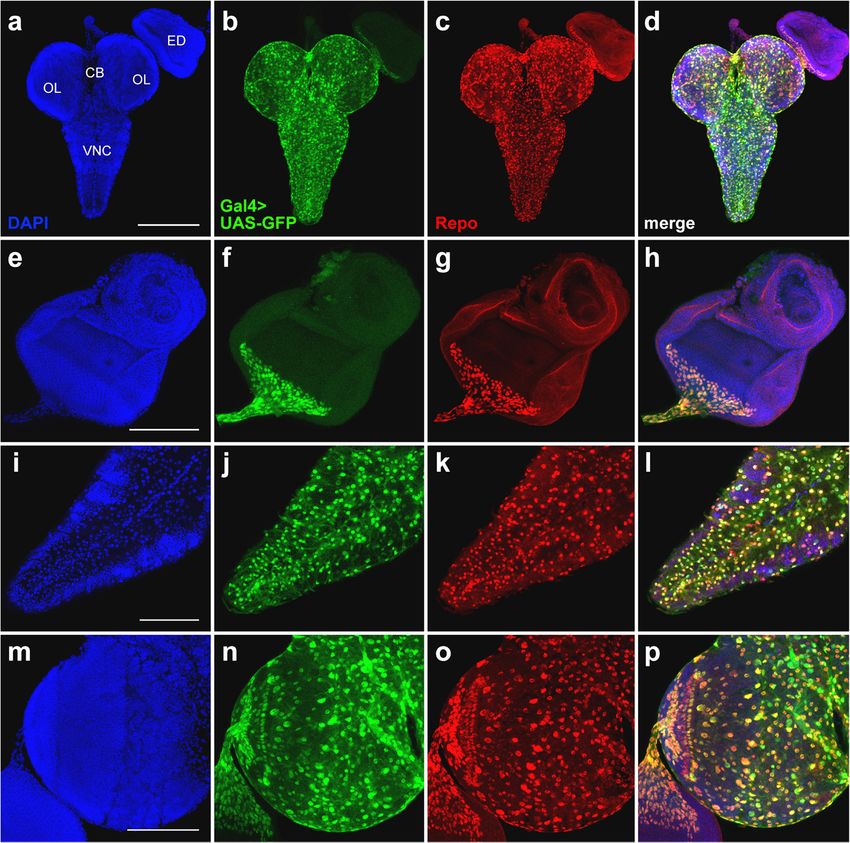
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 3 of 11
Chr2#8765, ChrX#458, Chr3#7415, and Chr3#1774 bearing the Chr3#7415, the orientation of the Gal4 coding sequence
Gal4 drivers in the five BDSC lines. To do that, we re- matches that of the elav and repo genes. In Chr3#1774,
placed all chromosomes (except the ones bearing the Gal4 the insertion site of P{GawB} element is in the promoter
constructs) in the original BDSC stocks with those from region of the corto gene (3R:5087163–5087170), but the
yw; KrIf-1/CyO; TM3, Sb1/TM6, Tb1 flies. Next, we per- orientation of the Gal4 coding sequence is opposite to
formed qPCR analysis using genomic DNA from such fly the orientation of this gene. In addition, this chromo-
lines with pairs of primers specific to the 5′ and 3′ P- some carries an internally truncated P-element sequence
element ends [31]. Notably, the minimal functional 5′ P- of ~ 1.1 kb (corresponding to positions 3152–2931 and
element end of 140 bp [32], present in the P{GawB} 1138–246 of GenBank accession number AB331393.1)
transposon and likely in P{Gal4} transposon, cannot be inserted in the distal antenna-related (danr) gene pro-
detected by these primers. qPCR indicates that there is moter region (3R:25138044–25138051). The presence
only one P-element construct in each of the following and location of all transposons was verified by PCR with
chromosomes: Chr3#8760, Chr2#8765, ChrX#458, and primers specific to the P-element ends and genomic se-
Chr3#7415 (Fig. 1). However, Chr3#1774 seems to carry not quences flanking the insertion sites (Additional file 1:
only the P{GawB} construct but also another unexpected Table S1).
transposon possessing a long 5′ P-element end (Fig. 1).
Next, we used an inverse PCR approach [33] to iden- Expression patterns of the Gal4 drivers
tify the insertion sites of the P-element constructs To reveal spatial activity of the Gal4 drivers carried
present in Chr3#8760, Chr2#8765, ChrX#458, Chr3#7415, and by Chr3#8760, Chr2#8765, ChrX#458, Chr3#7415, and
Chr3#1774 (Fig. 2). In Chr3#8760, the P{Gal4-elav.L} con- Chr3#1774 in the larval CNS and eye imaginal discs,
struct is inserted within an mdg3 retrotransposon (at we used a UAS-GFP reporter. Flies carrying the Gal4
DNA sequence position 731–738, GenBank accession drivers were crossed to UAS-GFP flies and the tissues
number X95908.1), which in turn is imbedded in the dissected from the resulting progeny were immuno-
second/third intron of the CG16779 gene (immediately stained with anti-Elav or anti-Repo antibodies and an-
after position 3R:9478115). In Chr2#8765, the insertion alyzed by confocal microscopy. GFP with a nuclear
site of the P{Gal4-elav.L} construct is in the promoter localization signal [34] was used because Elav and
region of the l(2)01289 gene (2R:6740727–6740734). In Repo are nuclear proteins [12, 35].
ChrX#458, the P{GawB} construct is inserted in the pro- In the larval CNS, the expression pattern of elav-Gal4
moter region of the elav gene (X:523343–523350), con- from lines #8760 and #8765 overlaps only partially with
firming the description of this line (https://flybase.org/ Elav immunostaining. The degree of colocalization of the
reports/FBrf0211983). In Chr3#7415, the P{Gal4} con- GFP and Elav signals is higher in the VNC than in the
struct is inserted in the repo gene promoter (3R: central brain or the optic lobes (Fig. 3a-b, d-e; Add-
18236035–18236042). Notably, in ChrX#458 and itional files 2 and 3: Figures S1 and S2). We also observed
Fig. 1 Copy numbers of P-element ends in different fly lines as measured by qPCR. One copy of P-element ends corresponds to a single
transgene in the heterozygous flies. As expected, no P-element ends were detected in yw; KrIf-1/CyO; TM3, Sb1/TM6, Tb1 flies. In the BDSC lines
#8760, #8765, #458, #7415, and #1774, all chromosomes except those carrying P-element insertions (based on the annotations of these lines
available at BDSC website) were replaced by chromosomes from the line yw; KrIf-1/CyO; TM3, Sb1/TM6, Tb1 before qPCR analysis: #8760 (yw; KrIf-1/
CyO; Chr3#8760/TM3, Sb1); #8765 (yw; Chr2#8765/KrIf-1; TM3, Sb1/TM6, Tb1); #458 (ChrX#458/yw; KrIf-1/CyO; TM3, Sb1/TM6, Tb1); #7415 (yw; KrIf-1/CyO;
Chr3#7415/TM3, Sb1); #1774 (yw; KrIf-1/CyO; Chr3#1774/TM3, Sb1). In line #1774, two P-element transgenes were detected. The experiment was done
in three replicates. Error bars represent standard errors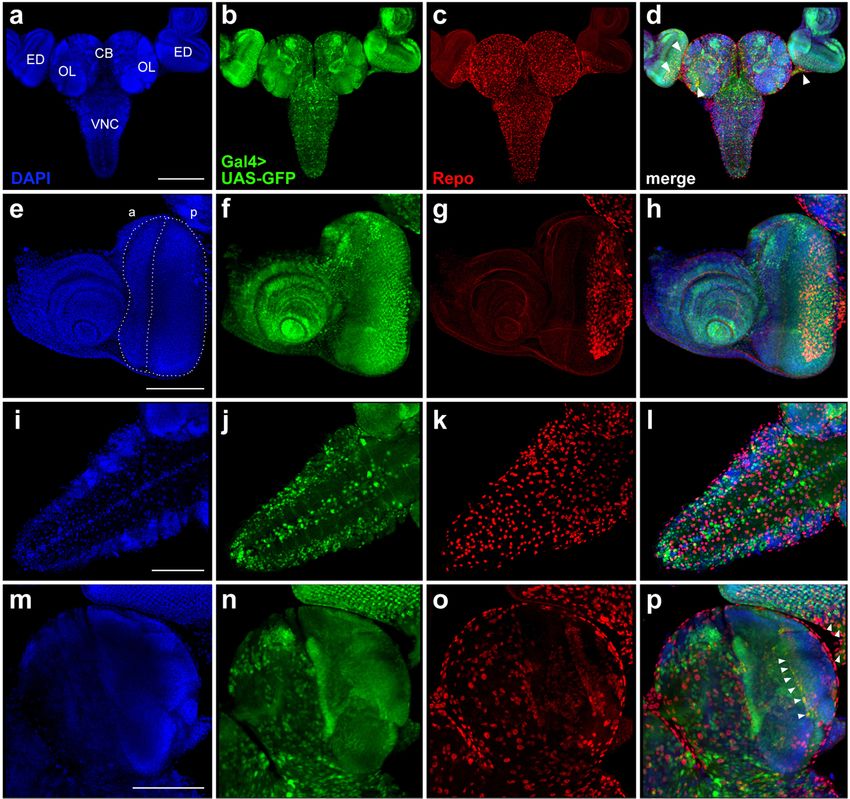
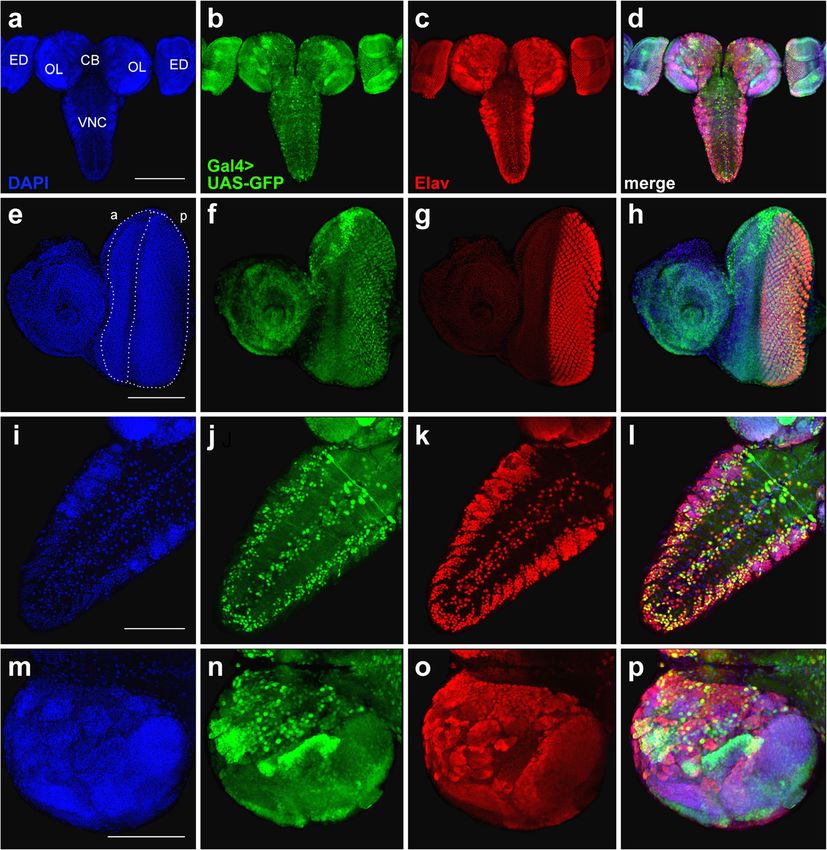
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 4 of 11 Fig. 2 Insertion sites of P-element constructs found in the BDSC lines #8760, #8765, #458, #7415, and #1774. Thick horizontal black lines show segments of chromosomes flanking the insertion sites of P-element constructs. Genes on the forward and reverse strands are shown in magenta and blue, respectively. Coding sequences, UTRs and introns are represented as wide bars, narrow bars and lines, respectively. Grey triangles represent the 5′ and 3′ P-element ends, as well as the 5′ and 3′ long terminal repeats of the mdg3 retrotransposon. Note that there are two types of 5′ P-element ends in the transposons: the “standard” ends of 585 bp and short ends of 140 bp, the latter are indicated by red asterisks. All P- element transgenes and the mdg3 retrotransposon are not shown to scale. LTR, long terminal repeat ectopic GFP expression in the neuropil and nerve roots Also, in the CNS from line #458 bearing the (Fig. 3b, e). In the eye imaginal discs, the endogenous Elav elavC155 allele, the overlapping of the GFP and Elav protein is expressed in photoreceptor neurons and there- signals is only partial. Signal colocalization is evident fore marks the developing eye area posterior to the mor- in the VNC but there is no ectopic GFP expression phogenetic furrow [36]. We found that the elav-driven in the neuropil (Fig. 3g, h; Additional file 5: Figure expression of Gal4 in lines #8760 and #8765 does not fully S4). In the eye imaginal discs of elavC155 larvae, the reproduce the pattern of Elav immunostaining, as the GFP GFP expression pattern is almost completely overlap- signals occupy a narrower area compared to the Elav sig- ping the Elav immunostaining (Fig. 3i; Additional files 4 nal (Fig. 3c, f; Additional file 4: Figure S3). and 5: Figures S3 and S4).
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 5 of 11 Fig. 3 Comparison of GFP expression patterns elicited by the elav-Gal4 drivers of the BDSC lines #8760, #8765, and #458 with the Elav immunostaining pattern in the CNS and eye-antennal imaginal discs from third-instar larvae. Maximum intensity projections of confocal images of tissues stained with anti-Elav antibodies and showing GFP expression in the CNS and eye-antennal discs driven by Chr3#8760 (a-c), Chr3#8765 (d-f) or Chr3#458 (g-i). a, d, g The GFP and Elav patterns overlap only partially in the central brain and optic lobes. b, e Colocalization of the GFP and Elav signals in the VNC. Note the ectopic GFP expression in the neuropil (arrows) and in nerve roots (arrowheads). c, f In the eye imaginal discs, the pattern of the GFP expression is narrower compared to anti-Elav antibody staining. h Substantial colocalization of the GFP and Elav signals in the VNC. i In the eye imaginal discs, the patterns of GFP and Elav expression seem largely overlap. CB, central brain; OL, optic lobe; ED, eye imaginal disc; VNC, ventral nerve cord. Scale bars: a, d, g, 200 μm; b, c, e, f, h, i, 100 μm In the optic lobes and eye imaginal discs, the GFP ex- brain, optic lobes and optic stalk also express GFP. In pression pattern directed by the Gal4repodriver of line the eye-antennal disc, Repo is expressed only in the pos- #7415 is almost coincidental with the localization of the terior part of the eye disc, where it partially overlaps Repo protein (Fig. 4; Additional file 6: Figure S5). How- with the GFP signal (Fig. 6; Additional file 8: Figure S7). ever, in both the VNC and central brain, the GFP and Altogether, these results suggest that the 69B-Gal4 Repo localization patterns are slightly different (Fig. 4i-l; driver is mainly active in neuronal cells, although it is Additional file 6: Figure S5). also expressed in other cell types, including imaginal disc To characterize the expression pattern of the 69B- cells and small subsets of glial cells. Gal4 driver of line #1774, we compared the GFP localization elicited by this driver with the distribution Discussion of the Elav and Repo proteins in larval CNS and eye- The accessibility of the CNS of Drosophila third instar antennal imaginal discs. The GFP and Elav signals sub- larvae and its similarities to the developing mammalian stantially colocalize in the VNC and eye imaginal discs, nervous system make it a very attractive model system. but colocalization is limited in the central brain and the Here, we have characterized five transgenic fly lines car- optic lobes. In addition, the 69B-Gal4 driver is heavily rying commonly used Gal4 drivers for neurobiological expressed in the antennal disc, where Elav is not present studies. Specifically, we identified the genomic insertion (Fig. 5; Additional file 7: Figure S6). A comparison be- sites of the Gal4 transgenes and visualized the Gal4 ac- tween the 69B-Gal4 and Repo expression patterns re- tivity patterns using a UAS-GFP reporter. These patterns vealed that individual Repo-positive cells in the central were compared to the localization of the Elav and Repo
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 6 of 11 Fig. 4 Comparison of GFP expression patterns controlled by the Gal4repo driver from the BDSC line #7415 with the Repo expression pattern in the CNS and eye-antennal imaginal discs from third-instar larvae. Maximum intensity projections of confocal images of tissues stained with anti-Repo antibodies are shown. a-d Substantial overlap between the GFP and Repo signals in the CNS and eye imaginal discs. e-h Perfect colocalizations of the GFP and Repo signals in the eye imaginal disc and optic stalk. i-l Colocalization of the GFP and Repo expression patterns in the VNC. m-p Overlapping of the GFP and Repo signals in the central brain and optic lobes; note that the GFP expression pattern seems to be broader than the Repo immunostaining pattern. CB, central brain; OL, optic lobe; ED, eye imaginal disc; VNC, ventral nerve cord. Scale bars: a-d, 200 μm; e-p, 100 μm proteins that are believed to be specific for neuronal and maintenance of gene expression patterns throughout de- glial cell types, respectively. velopment [43]. In the CNS of third instar larvae, the ex- In lines #8760 and #8765, the functions of the pression of corto was detected in the optic lobes and the CG16779 and l(2)01289 genes might be compromised by VNC [44]. The danr gene encodes a putative transcrip- the insertion of the elav-Gal4 construct. Information on tion factor, which is involved in the retinal determin- these genes and their expression profiles is currently ation network during eye development [45, 46]. Lastly, it scarce. However, both genes were found to regulate is not clear whether the insertions of the enhancer-trap Drosophila lifespan [37, 38], suggesting some caution in Gal4 constructs in the lines #458 and #7415 somehow the interpretation the longevity experiments, in which affect the expression of the elav and repo genes, respect- the elav-Gal4 drivers were used [39–41]. In addition, the ively. Nevertheless, all these Gal4 driver lines, with the CG16779 gene is predicted to encode a factor involved exception of #8765, are homozygous or hemizygous in regulation of Choline acetyltransferase (ChAT), which viable. is responsible for the synthesis of the neurotransmitter The partial overlap of Gal4 expression from lines acetylcholine [42]. In line #1774, P-element insertions #8760 and #8765 with the localization of the Elav resulted in mutations in the corto and danr genes. The protein in the CNS and eye imaginal discs, as well as corto gene encodes an Enhancer of Trithorax and Poly- ectopic Gal4 expression in the neuropil, are not so comb (ETP) protein, which might be involved in the surprising. This finding is probably related to the
Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 7 of 11
Fig. 5 Comparison of GFP expression patterns elicited by the 69B-Gal4 driver from the BDSC line #1774 with the Elav immunostaining pattern in
the CNS and eye-antennal imaginal discs from third-instar larvae. Maximum intensity projections of confocal images of tissues stained with anti-
Elav antibodies are shown. a-d Overlapping of the GFP expression pattern and the Elav localization in the CNS and eye imaginal discs. e-h In the
eye-antennal disc, the Elav staining is largely coincident with the GFP signal, but the GFP expression is much broader than that of the Elav
protein and includes the anterior area of the eye disc and the antennal disc. i-l Partial colocalization of the GFP and Elav expression patterns in
the VNC. m-p Partial overlap between the GFP and Elav signals in the central brain and optic lobes. Note that GFP is expressed only in subset of
Elav-positive cells in the VNC. CB, central brain; OL, optic lobe; ED, eye imaginal disc; a, anterior part of the disc; p, posterior part of the disc; VNC,
ventral nerve cord. Scale bars: a-d, 200 μm; e-p, 100 μm
absence of important regulatory sequences controlling genes, in which they are integrated. These drivers do in-
the expression pattern of the endogenous elav gene deed show substantial, but not complete, overlap be-
within the P{Gal4-elav.L} transposon. Particularly, it tween the Gal4 expression and Elav and Repo
was recently reported that the elav gene is ubiqui- immunostaining, respectively. This indicates that some
tously transcribed and post-transcriptionally repressed regulatory signals, that normally control the activity of
in non-neural tissues via its 3′ UTR sequence [47], the elav and repo genes, became inactive or improperly
which is absent in the P{Gal4-elav.L} construct. At functional upon the insertion of the transgenes. The
the same time, the influence of the genomic location presence of the P-transposase promoter and the mini-
on the activity of the elav-Gal4 construct in lines white reporter gene in the P{GawB} and P{Gal4} ele-
#8760 and #8765 seems to be negligible, as both ments might be responsible for the phenomenon.
drivers elicit almost identical Gal4 expression The unique expression pattern of 69B-Gal4 driver is
patterns. most likely the result of interactions of the P-transposase
More interesting are the observations made for the promoter located upstream of the Gal4 coding sequence
enhancer-trap Gal4 drivers elavC155 and Gal4repo that in the P{GawB} construct with the enhancers and some
were expected to mimic the expression patterns of the other regulatory elements from the corto locus.Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 8 of 11
Fig. 6 Comparison of GFP expression patterns elicited by 69B-Gal4 driver from the BDSC line #1774 with the Repo expression pattern in the CNS
and eye-antennal imaginal discs from third-instar larvae. Maximum intensity projections of confocal images of tissues stained with anti-Repo
antibodies are shown. a-d In the CNS and eye-antennal imaginal discs, the GFP expression pattern overlaps with that of Repo just in individual
groups of cells (arrowheads). e-h In the eye-antennal imaginal discs, the broad GFP expression pattern overlaps with the Repo protein expression
pattern only in the optic stalk and in the posterior part of the eye disc. i-l The GFP and Repo expression patterns in the VNC are not overlapping.
m-p The GFP and Repo signals minimally overlap in the central brain and optic lobes; cells showing both signals are indicated by arrowheads.
CB, central brain; OL, optic lobe; ED, eye imaginal disc; a, anterior part of the disc; p, posterior part of the disc; VNC, ventral nerve cord. Scale bars:
a-d, 200 μm; e-p, 100 μm
Conclusions expression patterns of Gal4 drivers, should be taken into
Most of the currently available Gal4 drivers used in account in both experimental design and the interpre-
Drosophila neurobiology elicit expression patterns that tation of the results.
only partially overlap those of the endogenous proteins.
Transgenes encoding Gal4 under the control of pro-
moters of particular genes may lack important regulatory Methods
elements that are normally located either outside the Fly stocks
genes or in their exon-intron sequences. In addition, ex- Flies were raised and crossed on standard cornmeal agar
pression of such Gal4 transgenes may depend on their media at 25 °C. The fly stocks carrying driver constructs
genomic location. The enhancer-trap Gal4 constructs, used in this study were obtained from the BDSC
although showing expression patterns similar to those of (Bloomington, IN, USA; https://bdsc.indiana.edu/):
the genes in which they are integrated, also seem to lack #8760 (w*; P{w+mC = Gal4-elav.L}3); #8765 (P{w+mC =
some regulatory expression signals. Thus, a perfect copy- Gal4-elav.L}2/CyO); #458 (P{w+mW.hs = GawB}elavC155);
ing of the endogenous protein expression patterns by #7415 (w1118; P{w+m* = Gal4}repo/TM3, Sb1); #1774 (w*;
Gal4 drivers appears to be a very complex task. To ac- P{w+mW.hs = GawB}69B); #4775 (w1118; P{w+mC = UAS-
complish this task, it would be necessary to identify all GFP.nls}14). Line yw; KrIf-1/CyO; TM3, Sb1/TM6, Tb1
factors affecting the activity of the gene studied. Until was provided by the “Molecular and Cellular Biology”
this is achieved, all available information on the core facility of the IMCB SB RAS.Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 9 of 11
Genomic DNA extraction and determination of P-element pBS-F1 (5′-cagggttttcccagtcacgac-3′) and pBS-R1
transgene copy number (5′-ggctttacactttatgcttcc-3′).
Genomic DNA was isolated from 30 to 50 flies accord-
ing to the protocol reported earlier [48]. For detection of
PCR genotyping
P-element-based transgene copy number by qPCR, we
PCR was performed using Hot-Start Taq DNA polymer-
used the reference plasmid pP5′-Vps36-759bp-P3′ de-
ase (Biolabmix) according to the manufacturer’s recom-
scribed previously [31]. The following three primer pairs
mendations. Details of primer pairs used for genotyping
were used: Vps36-realtime-F and Vps36-realtime-R spe-
all P-element transposons are provided in Add-
cific for the Vps36 gene, qP5-F1 and qP5-R1 for the 5′
itional file 1: Table S1. The PCR products were analyzed
P-element end, and qP3-F1 and qP3-R1 for the 3′ P-
on 1% agarose gel along with an appropriate DNA
element end (for primer sequences, see [31]). Import-
marker.
antly, primers qP5-F1 and qP5-R1 can detect only the
“standard” 5′ P-element end of 585 bp in length, but not
the minimal functional 5′ P-element end of 140 bp in Immunofluorescence staining and confocal microscopy
length that is present in enhancer-trap Gal4 transposons. The CNS and attached eye-antennal imaginal discs were
qPCR was performed with 100 ng of genomic DNA or 5 dissected from third instar larvae. For direct GFP detec-
pg of the reference plasmid pP5′-Vps36-759bp-P3′, 400 tion (without GFP immunostaining) and antibody stain-
nM of each primer in a 25-μl reaction mixture using the ing, tissues were fixed in phosphate-buffered saline
HS-qPCR SYBR Blue Master Mix (Biolabmix), and the (PBS) containing 4% formaldehyde (Merck), and then
CFX96 Touch Real-Time PCR Detection System (Bio- washed 3 times (5 min each) with 0.5% Triton X-100 in
Rad) under the following conditions: incubation at 95 °C PBS. The primary monoclonal rat anti-Elav (DSHB
for 5 min, followed by 39 cycles of 95 °C for 15 s, 60 °C #7E8A10) and monoclonal mouse anti-Repo (DSHB
for 30 s, and 72 °C for 30 s. Data analysis was performed #8D12) antibodies were used at concentration 1 μg/ml.
using CFX Manager™ Software v3.0 (Bio-Rad). The 5′ They were detected by goat anti-rat IgG antibodies con-
and 3′ P-element copy numbers per diploid genome jugated to AlexaFluor568 (1:800; Invitrogen #A-11077)
were calculated according to [31]. and by goat anti-mouse IgG antibodies conjugated to
AlexaFluor568 (1:800; Invitrogen #A-11031). Finally, tis-
Mapping P-element transgene insertion sites sues were stained with 0.4 μg/ml DAPI dissolved in PBS.
Mapping of P-element transgene insertion sites was All samples were imaged at the same settings using con-
done by inverse-PCR [33] according to “Inverse PCR & focal microscope LSM 710 (Carl Zeiss) with 10×/0.45
Cycle Sequencing of P Element Insertions for STS Gen- plan-apo and 20×/0.8 plan-apo lenses. Optical sections
eration” protocol of E.J. Rehm (Berkeley Drosophila were combined using the LSM Image Browser version
Genome Project; www.fruitfly.org/about/methods/in- 4.2 software (Carl Zeiss).
verse.pcr.html) with the following modifications. One
microgram of genomic DNA was digested with HhaI
(New England Biolabs) or Kzo9I (SibEnzyme) or MspI Supplementary information
Supplementary information accompanies this paper at https://doi.org/10.
(SibEnzyme) or SalI (New England Biolabs) restriction 1186/s12863-020-00895-7.
enzyme for 6 h at 37 °C in a volume of 50 μl. Restriction
fragments were circularized by incubation with T4 DNA Additional file 1: Table S1. Primers used for PCR verification of
ligase (Thermo Scientific) and then purified using transposon insertion sites and details of PCR products.
Microcon Ultracel YM-30 centrifugal filter (Millipore), Additional file 2: Figure S1. Representative series of z-stack confocal
images of the CNS and eye-antennal imaginal discs from a third-instar
followed by elution of DNA in 40 μl of nuclease-free larva expressing GFP under the control of the elav-Gal4 driver from the
water. Five microliters of each sample was used as a BDSC line #8760. The tissues are stained with anti-Elav antibodies.
template for PCR amplification of fragments containing Additional file 3: Figure S2. Representative series of z-stack confocal
the P-element end (5′ or 3′) and flanking genomic images of the CNS and eye-antennal imaginal discs from a third-instar
larva expressing GFP under the control of the elav-Gal4 driver from the
DNA. PCR reactions were performed using Phusion® BDSC line #8765. The tissues are stained with anti-Elav antibodies.
High-Fidelity DNA Polymerase (New England Biolabs) Additional file 4: Figure S3. Comparison of GFP expression patterns
and the following two primer pairs: Plac1 and Plac4 elicited by the Gal4 drivers from the BDSC lines #8760, #8765, and #458
specific for the 5′ P-element end of 585 bp (but not of with the Elav immunostaining pattern in the eye-antennal imaginal discs
from third-instar larvae. CB, central brain; OL, optic lobe. Scale bar:
140 bp) in length, and Pry1 and Pry2 specific for the 3′ 100 μm.
P-element end (for primer sequences, see Add- Additional file 5: Figure S4. Representative series of z-stack confocal
itional file 1: Table S1). The gel-purified PCR products images of the CNS and eye-antennal imaginal discs from a third-instar
were cloned into pBluescript II SK+ vector (Promega) larva expressing GFP under the control of the elavC155 driver from the
BDSC line #458. The tissues are stained with anti-Elav antibodies.
for subsequent sequencing using universal primersOgienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 10 of 11
Additional file 6: Figure S5. Representative series of z-stack confocal Published: 22 October 2020
images of the CNS and eye-antennal imaginal discs from a third-instar
larva expressing GFP under the control of the Gal4repo driver from the References
BDSC line #7415. The tissues are stained with anti-Repo antibodies. 1. Reiter LT, Potocki L, Chien S, Gribskov M, Bier E. A systematic analysis of
Additional file 7: Figure S6. Representative series of z-stack confocal human disease-associated gene sequences in Drosophila melanogaster.
images of the CNS and eye-antennal imaginal discs from a third-instar Genome Res. 2001;11(6):1114–25.
larva expressing GFP under the control of the 69B-Gal4 driver from the 2. Xu Z, Tito AJ, Rui Y-N, Zhang S. Studying polyglutamine diseases in
BDSC line #1774. The tissues are stained with anti-Elav antibodies. Drosophila. Exp Neurol. 2015;274(Pt A):25–41.
Additional file 8: Figure S7. Representative series of z-stack confocal 3. McGurk L, Berson A, Bonini NM. Drosophila as an in vivo model for human
images of the CNS and eye-antennal imaginal discs from a third-instar neurodegenerative disease. Genetics. 2015;201(2):377–402.
larva expressing GFP under the control of the 69B-Gal4 driver from the 4. Duffy JB. GAL4 system in Drosophila: a fly geneticist's Swiss army knife.
BDSC line #1774. The tissues are stained with anti-Repo antibodies. Genesis. 2002;34(1–2):1–15.
5. Venken KJT, Simpson JH, Bellen HJ. Genetic manipulation of genes and cells
in the nervous system of the fruit fly. Neuron. 2011;72(2):202–30.
6. Griffith LC. Identifying behavioral circuits in Drosophila melanogaster:
Abbreviations
moving targets in a flying insect. Curr Opin Neurobiol. 2012;22(4):609–14.
BDSC: Bloomington Drosophila Stock Center; CB: Central brain; CNS: Central
7. Jenett A, Rubin GM, Ngo T-TB, Shepherd D, Murphy C, Dionne H, Pfeiffer
nervous system; DSHB: Developmental Studies Hybridoma Bank;
BD, Cavallaro A, Hall D, Jeter J, et al. A GAL4-driver line resource for
Elav: Embryonic lethal, abnormal vision; GFP: Green fluorescent protein;
Drosophila neurobiology. Cell Rep. 2012;2(4):991–1001.
LTR: Long terminal repeats; PBS: Phosphate-buffered saline; Repo: Reverse
8. Manning L, Heckscher ES, Purice MD, Roberts J, Bennett AL, Kroll JR, Pollard
polarity; VNC: Ventral nerve code; UAS: Upstream activation sequence;
JL, Strader ME, Lupton JR, Dyukareva AV, et al. A resource for manipulating
UTR: Untranslated region
gene expression and analyzing cis-regulatory modules in the Drosophila
CNS. Cell Rep. 2012;2(4):1002–13.
Acknowledgments 9. Sousa-Nunes R, Cheng LY, Gould AP. Regulating neural proliferation in the
We thank Maurizio Gatti for critical reading of the manuscript. Microscopy Drosophila CNS. Curr Opin Neurobiol. 2010;20(1):50–7.
analysis and DNA sequencing were carried out at the “Molecular and Cellular 10. Freeman MR. Drosophila central nervous system glia. Cold Spring Harb
Biology” core facility of the IMCB SB RAS. Perspect Biol. 2015;7(11):a020552.
11. Robinow S, White K. The locus elav of Drosophila melanogaster is expressed
in neurons at all developmental stages. Dev Biol. 1988;126(2):294–303.
About this supplement 12. Robinow S, White K. Characterization and spatial distribution of the ELAV
This article has been published as part of BMC Genetics Volume 21 protein during Drosophila melanogaster development. J Neurobiol. 1991;
Supplement 1, 2020: Selected Topics in “Systems Biology and Bioinformatics” 22(5):443–61.
- 2019: genetics. The full contents of the supplement are available online at 13. Berger C, Renner S, Lüer K, Technau GM. The commonly used marker ELAV
https://bmcgenet.biomedcentral.com/articles/supplements/volume-21- is transiently expressed in neuroblasts and glial cells in the Drosophila
supplement-1. embryonic CNS. Dev Dyn. 2007;236(12):3562–8.
14. Luo L, Liao YJ, Jan LY, Jan YN. Distinct morphogenetic functions of similar
Authors’ contributions small GTPases: Drosophila Drac1 is involved in axonal outgrowth and
Conceived and designed the experiments: AVP. Performed the experiments: myoblast fusion. Genes Dev. 1994;8(15):1787–802.
ENA, AAO, ESO, ALO. Analyzed the data: ENA, AAO, ESO, AVP. Wrote the 15. Yao K-M, White K. Neural specificity of elav expression: defining a Drosophila
paper: AAO, ESO, ENA, AVP. All authors read and approved the final promoter for directing expression to the nervous system. J Neurochem.
manuscript. 1994;63(1):41–51.
16. Brand AH, Perrimon N. Targeted gene expression as a means of altering cell
fates and generating dominant phenotypes. Development. 1993;118(2):401–15.
Funding 17. Hoskins RA, Carlson JW, Wan KH, Park S, Mendez I, Galle SE, Booth BW,
This work was mainly supported by the Russian Science Foundation grant Pfeiffer BD, George RA, Svirskas R, et al. The Release 6 reference sequence of
18–74-00050 and in part of qPCR experiments by a project of the the Drosophila melanogaster genome. Genome Res. 2015;25(3):445–58.
Fundamental Scientific Research Program of the Siberian Branch of the 18. Lin DM, Goodman CS. Ectopic and increased expression of Fasciclin II alters
Russian Academy of Sciences (0310–2019-0005). Publication costs are funded motoneuron growth cone guidance. Neuron. 1994;13(3):507–23.
by the Russian Science Foundation grant 18–74-00050. The funders had no 19. Casas-Tintó S, Arnés M, Ferrús A. Drosophila enhancer-Gal4 lines show
role in study design, data collection and analysis, decision to publish, or ectopic expression during development. R Soc Open Sci. 2017;4(3):170039.
preparation of the manuscript. 20. Kim MD, Kolodziej P, Chiba A. Growth cone pathfinding and filopodial
dynamics are mediated separately by Cdc42 activation. J Neurosci. 2002;
Availability of data and materials 22(5):1794–806.
All materials are available upon request. 21. Liu J, Li T, Yang D, Ma R, Moran TH, Smith WW. Synphilin-1 alters metabolic
homeostasis in a novel Drosophila obesity model. Int J Obes. 2012;36(12):
1529–36.
Ethics approval and consent to participate 22. Kang J, Shin S, Perrimon N, Shen J. An evolutionarily conserved role of
Not applicable. Presenilin in neuronal protection in the aging Drosophila brain. Genetics.
2017;206(3):1479–93.
23. Sepp KJ, Schulte J, Auld VJ. Peripheral glia direct axon guidance across the
Consent for publication
CNS/PNS transition zone. Dev Biol. 2001;238(1):47–63.
Not applicable.
24. Xiong W-C, Okano H, Patel NH, Blendy JA, Montell C. repo encodes a glial-
specific homeo domain protein required in the Drosophila nervous system.
Competing interests Genes Dev. 1994;8(8):981–94.
The authors declare no competing financial interests. 25. Ren Q, Awasaki T, Wang Y-C, Huang Y-F, Lee T. Lineage-guided Notch-
dependent gliogenesis by Drosophila multi-potent progenitors.
Author details Development. 2018;145(11):dev160127.
1
Institute of Molecular and Cellular Biology, Siberian Branch of RAS, 26. Tamura T, Sone M, Yamashita M, Wanker EE, Okazawa H. Glial cell lineage
Novosibirsk 630090, Russia. 2Institute of Chemical Biology and Fundamental expression of mutant ataxin-1 and huntingtin induces developmental and
Medicine, Siberian Branch of RAS, Novosibirsk 630090, Russia. 3Novosibirsk late-onset neuronal pathologies in Drosophila models. PLoS One. 2009;4(1):
State University, Novosibirsk 630090, Russia. e4262.Ogienko et al. BMC Genetics 2020, 21(Suppl 1):96 Page 11 of 11
27. Rimmington G, Dalby B, Glover DM. Expression of N-terminally truncated Publisher’s Note
cyclin B in the Drosophila larval brain leads to mitotic delay at late Springer Nature remains neutral with regard to jurisdictional claims in
anaphase. J Cell Sci. 1994;107(Pt 10):2729–38. published maps and institutional affiliations.
28. Huang X, Suyama K, Buchanan J, Zhu AJ, Scott MP. A Drosophila model
of the Niemann-pick type C lysosome storage disease: dnpc1a is
required for molting and sterol homeostasis. Development. 2005;
132(22):5115–24.
29. Loveall BJ, Deitcher DL. The essential role of bursicon during Drosophila
development. BMC Dev Biol. 2010;10:92.
30. Palgi M, Greco D, Lindström R, Auvinen P, Heino TI. Gene expression
analysis of Drosophilaa Manf mutants reveals perturbations in membrane
traffic and major metabolic changes. BMC Genomics. 2012;13:134.
31. Ogienko AA, Yarinich LA, Fedorova EV, Lebedev MO, Andreyeva EN,
Pindyurin AV, Baricheva EM. New slbo-Gal4 driver lines for the analysis of
border cell migration during Drosophila oogenesis. Chromosoma. 2018;
127(4):475–87.
32. Mullins MC, Rio DC, Rubin GM. Cis-acting DNA sequence requirements for
P-element transposition. Genes Dev. 1989;3(5):729–38.
33. Ochman H, Gerber AS, Hartl DL. Genetic applications of an inverse
polymerase chain reaction. Genetics. 1988;120(3):621–3.
34. Shiga Y, Tanaka-Matakatsu M, Hayashi S. A nuclear GFP/β-galactosidase
fusion protein as a marker for morphogenesis in living Drosophila. Develop
Growth Differ. 1996;38(1):99–106.
35. Halter DA, Urban J, Rickert C, Ner SS, Ito K, Travers AA, Technau GM. The
homeobox gene repo is required for the differentiation and maintenance of
glia function in the embryonic nervous system of Drosophila melanogaster.
Development. 1995;121(2):317–32.
36. Fan Y, Wang S, Hernandez J, Yenigun VB, Hertlein G, Fogarty CE, Lindblad
JL, Bergmann A. Genetic models of apoptosis-induced proliferation
decipher activation of JNK and identify a requirement of EGFR signaling for
tissue regenerative responses in Drosophila. PLoS Genet. 2014;10(1):
e1004131.
37. Nakayama M, Ishibashi T, Ishikawa HO, Sato H, Usui T, Okuda T, Yashiro H,
Ishikawa H, Taikou Y, Minami A, et al. A gain-of-function screen to identify
genes that reduce lifespan in the adult of Drosophila melanogaster. BMC
Genet. 2014;15:46.
38. Rival T, Page RM, Chandraratna DS, Sendall TJ, Ryder E, Liu B, Lewis H,
Rosahl T, Hider R, Camargo LM, et al. Fenton chemistry and oxidative stress
mediate the toxicity of the β-amyloid peptide in a Drosophila model of
Alzheimer’s disease. Eur J Neurosci. 2009;29(7):1335–47.
39. Balan V, Miller GS, Kaplun L, Balan K, Chong Z-Z, Li F, Kaplun A,
VanBerkum MFA, Arking R, Freeman DC, et al. Life span extension and
neuronal cell protection by Drosophila nicotinamidase. J Biol Chem.
2008;283(41):27810–9.
40. Morrow G, Samson M, Michaud S, Tanguay RM. Overexpression of the small
mitochondrial Hsp22 extends Drosophila life span and increases resistance
to oxidative stress. FASEB J. 2004;18(3):598–9.
41. Slade FA, Staveley BE. Arm-Gal4 inheritance influences development and
lifespan in Drosophila melanogaster. Genet Mol Res. 2015;14(4):12788–96.
42. Konstantinides N, Kapuralin K, Fadil C, Barboza L, Satija R, Desplan C.
Phenotypic convergence: distinct transcription factors regulate common
terminal features. Cell. 2018;174(3):622–35.
43. Salvaing J, Nagel AC, Mouchel-Vielh E, Bloyer S, Maier D, Preiss A, Peronnet
F. The enhancer of Trithorax and Polycomb Corto interacts with Cyclin G in
Drosophila. PLoS One. 2008;3(2):e1658.
44. Harvie PD, Filippova M, Bryant PJ. Genes expressed in the ring gland, the
major endocrine organ of Drosophila melanogaster. Genetics. 1998;149(1):
217–31.
45. Curtiss J, Burnett M, Mlodzik M. distal antenna and distal antenna-related
function in the retinal determination network during eye development in
Drosophila. Dev Biol. 2007;306(2):685–702.
46. Hoang CQ, Burnett ME, Curtiss J. Drosophila CtBP regulates proliferation and
differentiation of eye precursors and complexes with Eyeless, Dachshund,
Dan, and Danr during eye and antennal development. Dev Dyn. 2010;
239(9):2367–85.
47. Sanfilippo P, Smibert P, Duan H, Lai EC. Neural specificity of the RNA-
binding protein Elav is achieved by post-transcriptional repression in non-
neural tissues. Development. 2016;143(23):4474–85.
48. Pindyurin AV. Genome-wide cell type-specific mapping of in vivo chromatin
protein binding using an FLP-inducible DamID system in Drosophila.
Methods Mol Biol. 2017;1654:99–124.You can also read

















































