ORDERING OF THE CELLULAR ARRANGEMENT AND XYLOGENESIS IN WOUNDED SHOOTS OF WILLOW - BRILL
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Zajączkowska – Cellular
IAWA Journal
arrangement
36 (4), and
2015:
xylogenesis
387–399 in wound callus 387
Ordering of the cellular arrangement and
xylogenesis in wounded shoots of willow
Urszula Zajączkowska
Department of Forest Botany, Warsaw University of Life Sciences, 159 Nowoursynowska Street,
02-776 Warsaw, Poland
E-mail: urszula.zajaczkowska@wl.sggw.pl
Abstract
Development of living organisms is characterized by self-organization, which
results in ordered cell and tissue patterns. Xylem formation in callus tissue may
serve as a model to study these phenomena. Applying auxin on the apical trans-
verse cut surface of willow shoot segments stimulates the proliferation of callus
with an unorganized cell arrangement. In some areas of callus, the cells form an
ordered system and partly differentiate into tracheary elements. Below the cut
surface a zone of initially unorganized parenchymatous cells is produced by the
cambium. Later, some of the cells formed ordered arrangements giving rise to
differentiation in xylem rays with a subsequent layer of normal wood. Digital
image processing software based on a structure tensor revealed a more coherent
orientation of the cellular pattern in the callus region close to the cambial zone
in the cut shoot surface, compared with the areas at further distances near the
outer parts of the callus ring. Differentiation of tracheary xylem elements occurs
mostly in the regions where a higher degree of cellular ordering in parenchyma
tissue is observed. Digital image analysis is a useful tool for the quantitative
estimation of subtle changes of cellular ordering in various regions of regenerat-
ing tissue. Wider application of this tool may open new opportunities in studies
of the complex mechanisms that control morphogenetic patterns in plants.
Keywords: Callus, cambium, differentiation, image processing, morphogene-
tic pattern, Salix alba, structure tensor.
Introduction
The control mechanisms of morphogenetic processes involved in the development of
secondary xylem in woody plants can be considered from various points of view and
analyzed using different approaches. Studies on the regenerative processes that occur
in the injured plant tissues and organs are of particular interest in understanding these
mechanisms. Woody plants, similar to other living organisms, are capable of develop-
ing increasingly complex systems. This directly translates into the morphological and
anatomical structure of cells, tissues and organs (Oborny 2004; Barthelemy & Caraglio
2007). This capability is typical for regeneration processes, during which it is manifested
in its most characteristic form (Biggs 1986; de Klerk et al. 1997; Nishitani et al. 2002;
© International Association of Wood Anatomists, 2015 DOI 10.1163/22941932-20150109
Published by Koninklijke Brill NV, Leiden
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access388 IAWA Journal 36 (4), 2015
Du et al. 2006). Regeneration first of all means the restoration of tissue function, and
requires a certain form to be generated within the morphogenetic potential of a plant.
To trigger the process of regeneration, a signal inducing a chain of morphogenetic
events must be recognized.
It is thought that knowing the factors that control signal perception is essential to
understanding growth and regeneration processes in plants in general. The cambium
plays a key role in plant growth reactions (Larson 1994; Lachaud et al. 1999). Also,
in both woody and herbaceous dicot plants, the flow of phytohormones, which induce
specific divisions in the cambium, is of great importance (Sablowski 2007). Many
studies have set out to identify the specific genes and proteins responsible for morpho-
genetic reactions (Zhang et al. 2000; Plomion et al. 2000, 2001; Tocquard et al. 2014).
Research is also being conducted on the anatomical factors related to a given cambial
reaction, such as plasmodesmatal network density (Fuchs et al. 2011), which changes
at various stages and leads to different types of meristematic reactions.
Many questions in this area remain unanswered. It is known, however, that coordinat-
ing the processes of growth and differentiation requires the presence of a mechanism
that integrates morphogenetic processes at various levels of plant organization. This
mechanism can be linked to the concept of morphogenetic field, which can also be
explored by the methods of digital image analysis. The objective of the present study
was to investigate the role of positional control of tracheary element differentiation
during both callus formation and the regeneration of secondary willow shoot tissues.
Special attention was paid to the cellular ordering within the callus tissue in relation to
differentiation of tracheary elements. So far, the issue of cellular ordering in morpho-
genetic processes was raised mainly by authors studying animal and micro-organism
systems, characterized by cells with an ability to move and form various geometric
patterns (Klebe et al. 1991; Volfson et al. 2008). However, in the case of plant cells,
which are commonly thought to be immobile, intrusive growth of plant cells enables
positional adjustments of adjoining cells (Lev-Yadun 2001). In addition, although the
problem of cellular ordering during plant development has been discussed by many
authors dealing with plant anatomy (e.g. Fahn 1990; Romberger et al.1993; Hejnowicz
2002), there is a deficiency of quantitative assessments of cellular ordering.
Callus, as a parenchymatous tissue in which xylem and phloem can originate, ap-
pears to be a suitable model for investigating these phenomena. In addition, a disrup-
tion of the continuity of stem tissues can affect cells not only on the wound’s surface
but also in some tissues located at a certain distance from the wound. The present work
focuses on the cellular ordering and xylogenesis in callus formed on the apical surface
of transversely cut young shoots of Salix alba and also in the shoot region just below
the cut surface. The selection of white willow for the experiments was mainly driven
by the highly regenerative capacity of this tree species. To measure the local ordering
of cellular arrangement, the method of digital image analysis based on a structure ten-
sor can be used. It has recently been shown that this technique may be useful for the
evaluation of cellular pattern ordering during morphogenetic processes in plants. The
method was successfully applied to pattern analysis and cellular ordering in the area
where tissues were regenerating after longitudinal stem wounding of adult Scots pine
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 389
trees (Zajączkowska 2014a) and during the overgrowth process of transverse surfaces
of Douglas fir stumps (Zajączkowska 2014b).
Materials and METHODS
Stem segment cultures
White willow (Salix alba L.) shoots were collected in April. The shoots, 15–25 mm
in diameter, were cut transversely into 15-cm-long sections. Lanoline paste with auxin
(IAA at conc. 1.0 % w/w) or lanoline without auxin was applied to the apical surface
of the shoot segments, which were then covered with aluminum foil. The basal ends of
the shoot segments were placed in containers with water and kept at 21°C in dispersed
sunlight during the day and halogen light after dark. The experiment, which used 46
replicate stem segments with IAA and 10 control segments without IAA, started on
April 10. After 14 days of cultivation, 2-cm-long apical fragments were cut off of 30
shoots with IAA treatment and 10 control shoots with no IAA applied, together with
the calluses that had formed, and these were studied under the microscope. Newly-
formed tissues from below the cut surface were studied in both the shoots that were
cultivated for two weeks and also in the 6 additional shoots with IAA that were kept
in the culture for eight weeks.
Microscopic observations
Microscopic examinations were carried out on 30-μm-thick sections, cut with a
sliding microtome. The presence of lignin was tested with the phloroglucinol–HCl
reaction. An Olympus optical UV microscope with integrated Cell^P software was
used for the anatomical investigations. The photomicrographs were used to determine
the cellular ordering in developing tissues by digital image analysis.
Measurement of cellular ordering
Analyses of cellular ordering seen in the microscopic sections were based on a
structure tensor which is commonly used in the field of digital image processing (Jahne
1993; Bigun et al. 2004). The OrientationJ image processing tool, which is an ImageJ
plug-in, was used. OrientationJ was originally developed for the measurement orienta-
tion of elastin fibres in human cerebral arteries (Fonck et al. 2009). It was then used
to analyze collagen orientation in arterial adventitia (Rezakhaniha et al. 2012) as well
as to measure cellular ordering in regenerating plant tissues (Zajączkowska 2014a, b).
A structure tensor is defined for each pixel as the 2×2 positive matrix. The tensor is
calculated for each pixel by computing the continuous spatial derivatives in the principal
directions x and y using cubic B-spline interpolation (Unser et al. 1993). Dominant
directions in the environment of a pixel gradient and coherency coefficients for the
chosen regions of image can then be calculated (Jahne 1993). A coherency coefficient
close to 1, geometrically represented as a slender ellipse, indicates highly oriented
structures, while a coherency coefficient close to zero, represented as a circle, denotes
no preferential orientation (isotropic areas). The OrientationJ software also outputs a
colour-coded map, which shows the predominant angles of the oriented structures to
the coordinates selected for the image. In the case of this study, the angle orientations
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access390 IAWA Journal 36 (4), 2015
were measured with respect to the image frames’ horizontal axis. The OrientationJ
plug-in and ImageJ macros used in the present studies are available online at http://
bigwww.epfl.ch/demo/orientation.
Results
Callus on the shoots’ cut surface
After 14 days of cultivation, a doughnut-shaped layer of callus of 2–5 mm formed
on the wound surface at the apical end of all 30 shoots where IAA with lanoline paste
had been applied. The callus developed above the areas of the cambial zone, the phloem
and the youngest xylem layers (Fig. 1A, B). It was also noted that after two weeks of
culture, the shoot diameter gradually increased (up to 6 mm), starting at 15 mm below
A C
Figure 1. Apical wound surface of
willow shoots after two weeks of cul-
ture. – A: Callus layer on the trans-
verse cut apical wound surface of a
shoot on which IAA was applied. –
B: Radial section of the shoot seg-
ment with callus. – C: Apical wound
B surface of shoot on which no IAA
was applied; no callus is formed and
new shoots developed from buds in
the apical part of the stem segment. —
Scale bar = 10 mm.
→
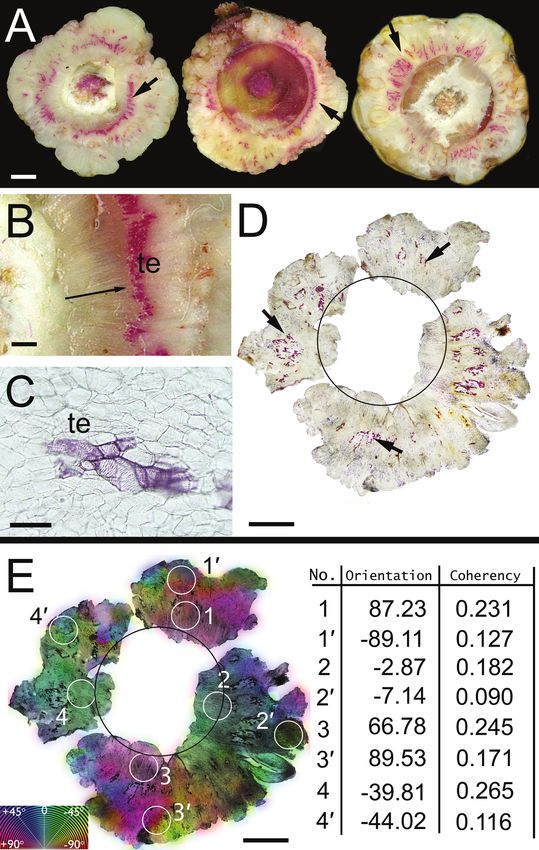
Figure 2. Cellular organization of callus formed on apical transverse cut wound surface of willow
stem shoots treated with IAA after two weeks of culture. The sections were cut perpendicularly
with respect to the shoots’ longitudinal axis and stained with phloroglucinol-HCl for lignified cell
walls. – A: Typical sections of calluses at about 150 μm above the cut surfaces of three shoots;
radial arrangements of tracheary elements’ lignified cell walls (arrows). – B: Tissue sector marked
by arrow on A (left section) under higher magnification; radially oriented clusters of tracheary
elements (te) above the shoot cambium (position indicated by arrow). – C: Cluster of tracheary
elements (te) surrounded by parenchymatous cells; the tracheary elements resemble vessel ele-
ments. – D: Section of callus further above the cut shoot surface (about 500 μm); within the re-
gion of unorganized parenchymatous callus tissue, some cells differentiated into tracheary ele-
ments with lignified cell walls (arrows); the black circle indicates the position of vascular cambium
at the shoot surface of the callus. – E: Colour-coded maps of local predominant cellular angle
orientations in the callus regions seen in D; numerical values for angle orientations (degrees) and
coherency coefficients for four pairs of sample points on the callus tissue; (1–4) and (1'–4')-sam-
ples located close and at farther distances from the cambial region on the cut shoot surface,
respectively. — Scale bars: 2500 μm in A; 1000 μm in B; 100 μm in C; 3500 μm in D and E.
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 391
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access392 IAWA Journal 36 (4), 2015
the cut surface and extending in the direction of the stem’s wound surface. At the apical
cut surface of the control shoots, where the lanoline paste without IAA was applied,
no callus formation was observed, nor did the shoot’s diameter increase (Fig. 1C).
Some of the lateral buds in apical parts of the stem segments without IAA developed
new shoots, whereas the buds in segments treated with the synthetic auxin remained
inactive during the period of culture.
Observations of the callus usually revealed an unorganized arrangement of paren-
chyma cells, which is a typical characteristic of this tissue. In the area of callus, about
150 μm above the cut shoot surface, radial clusters of tracheary elements were observed
as shown in three typical sections in Figure 2A. The lignified tracheary elements
occurred mainly in areas closer to the cambial region of the wounded shoot (Fig. 2 B).
At distances further away from the cambium, tracheary element clusters occurred in
lower densities. These clusters were shorter in the radial direction and wider in the
tangential direction as compared to clusters near the vascular cambium (Fig. 2 C, D).
The cellular ordering within the callus tissues was measured using the image analysis
software using a representative group of eight calluses. Typical results are shown in
Figure 2 E. The measurements are presented in a colour-coded map of local predominant
cellular orientation on tissue images (Fig. 2D). Spatial distribution of the colours on the
map indicates the radial direction of the predominant cellular orientation within the width
of the callus ring above the cut shoot surface. Measurements of the coherency coeffi-
cients were performed on eight locations within the callus (Fig. 2E). Four samples were
located close to the cambial zone on the cut shoot surface (1–4); the four remaining
samples were taken at distances further from the cambium and closer to the outer bound-
ary of the callus ring (1’– 4’). The numerical values of coherency coefficients indicate
a higher degree of cellular ordering near the cambial zone, as compared to those from
the regions located at distances further from the shoots’ vascular cambium (Fig. 2E).
Shoot below the cut surface
The increase in thickness of the IAA-treated shoots near the apical cut surface
(2 mm) was the result from higher cambial activity (Fig. 3A), mainly producing rows
of unlignified parenchyma cells. Later-formed cells had lignified walls. Thus, a paren-
chymatous layer (about 10–20 cells in a radial direction) that was visible in transverse
sections separated the two layers of xylem formed before and after the time of wound-
ing (Fig. 4A). In the area closer to the apical end, where the shoot was thicker, the
→
Figure 3. Transverse sections of willow shoot segment after two weeks of culture cut at distances
of 2 mm from the apical wound surface. Shoots treated with IAA (A & B) and without IAA (C)
applied to the apical wound surface. Parenchyma zone (p), seen in A and B, separates the xylem
layer formed in the previous season (x) and the layer of new xylem (nx) produced by cambium
(c) during the two-week experiment. – B: Tissue sector marked on A under higher magnification;
vessel elements (ve) with bordered pits in new xylem at the boundary of the annual ring of wood
formed in the previous season. – C: Cambial zone aligned to the boundary of the annual ring
of wood formed during the season before the experiment started (arrow) in shoots without IAA
after two weeks of culture. — Scale bars: 350 μm in A and C; 150 μm in B.
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 393
A
B
C
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access394 IAWA Journal 36 (4), 2015
A
B
C
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 395
larger radial dimension had a wider zone of both parenchyma and xylem cells. The
parenchyma zone became thinner with increasing distance from the cut surface, and
at about 15 mm below the wound it disappeared, while the amount of newly formed
xylem cells was also less. In certain cases, the transverse sections close to the cut shoot
surface showed that the first formed vessels were radially elongated and had bordered
pits in some wall regions. This may indicate that the first formed vessel elements were
oriented obliquely with respect to the shoot axis (Fig. 3B). In cases where no IAA was
applied to the apical cut surface of the shoot, no cambial activity or differentiation of
xylem elements were observed in the shoot regions below the cut surface (Fig. 3 C).
In the parenchyma layer, some of the cells differentiate into the xylem rays (Fig.
4A & B). Applying the digital image analysis software for measurement, the cellular
ordering within the parenchyma layer revealed low coherency coefficient values in the
region of unorganized cells (areas 1 and 2 in Fig 4 C) that were formed at the beginning
of the experiment. The higher values of this characteristic were found in the area where
the xylem rays start to differentiate (areas 3 and 4).
Discussion
The application of auxin (1.0% IAA) on the apical transverse cut surface in the stem
stimulated an intensive proliferation of the callus. The tissue was composed primarily of
unorganized parenchyma cells. Tracheary elements with a characteristic spatial pattern
differentiated from the callus tissue: directly above the cambial zone in the transverse
cut surface, these elements formed radially-aligned clusters. The further the clusters
were from the vascular cambium, closer to the outer surface of the callus ring, the
sparser they became. The image processing software based on a structure tensor that
was used to measure the angular orientation of cellular structures showed that these
structures were predominantly oriented radially along the entire width of the callus
ring. It should be noted that the results of angular orientation measurements concern
all types of cells present in the analyzed tissue. This means that the observed radial
orientation of tracheary elements is consistent with the predominant angular orienta-
tions of all types of cells present in a given region, including parenchyma cells, which
constituted the dominant type of cells.
←
Figure 4. A: Transverse section of a willow shoot segment after eight weeks of culture cut at a
distance of 2 mm from the apical wound surface, as seen under the UV microscope. – B: Tissue
section marked (on A) under higher microscopic magnification. – C: Coherency coefficients
represented geometrically as ellipses at four sample points located in the microscopic image
of tissue seen on B; numerical values for angle orientations (degrees) and coherency coeffi-
cients of four sample points in the parenchymatous tissue regions shown in C were as follows:
(1) -14.9° and 0.085, (2) -8.7° and 0.081, (3) -45.1° and 0.340, (4) -76.7° and 0.190; angle
orientations were measured with respect to the image frames’ horizontal axis. The parenchyma
zone (p) separates xylem layer formed during the previous season (x) and the layer of new xylem
(nx) with rays (dashed lines) produced by cambium during the eight-week experiment. — Scale
bars: 250 μm in A; 50 μm in B and C.
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access396 IAWA Journal 36 (4), 2015
Measurements of the coherency coefficients of cellular structures in the region of
the callus showed that cells above the cambial region were ordered to a greater degree
than cells in regions located farther away, closer to the outer layers of callus rings.
A decrease in coherency coefficient values was accompanied by a less frequent pres-
ence and a less clear radial orientation of tracheary element clusters. This was related
to the fact that these radially oriented clusters were shorter and wider as compared to
the more elongated clusters closer to the cambium.
Xylogenetic processes in the callus observed in this study occurred in the tissue
induced to grow by the application of IAA. Callus did not form in samples that were
only wounded (i.e., those for which no exogenous auxin was applied). However,
perhaps the observed effects should not be analyzed only from the perspective of a
direct effect of IAA on morphogenetic processes. Taking into account that the applied
synthetic auxin concentration was high (1.0%), some of the observed xylogenetic proc-
esses could be expected to have been related to the effects of auxin-induced ethylene
production (Yoshii & Imaseki 1981; Hansen & Grossmann 2000). In addition, the po-
tential effects of wound-induced production of ethylene (Fahn 1988; Morgan & Drew
1997) and jasmonate (Koo & Howe 2009) can also be involved. Directed division and
growth of cells play a major role in the development of callus tissue and cell ordering
processes (Fahn 1990; Hejnowicz 2002). Some authors state that these processes may
be related to mechanical stresses that can occur due to interactions between cells in
developing tissues and organs (Brown & Sax 1962; Brown 1964; Makino et al. 1983;
Dumais 2007; Hejnowicz 2011). The importance of this factor in morphogenetic proc-
esses in regenerating tissue of woody plants may still be questioned due to insufficient
experimental evidence on mechanical stresses within a developing tissue. The present
study did not involve a biomechanical analysis. Nonetheless, it can be assumed that the
radial pattern of the predominant angular orientation may correspond to a hypothetical
field of mechanical stresses in the callus tissue that formed a ring on the cut surface of
the stem. It should be noted, however, that spatial patterns of numerous physical fields
representing extremely different physical phenomena are often similar and described by
the same mathematical equations, because the common element to all these phenomena
is ‘space’, the framework of physics and biology (Feynman et al. 1964).Therefore, so
far there are no sufficient grounds to claim that mechanical stresses were engaged in
the regulation of ordering and xylogenesis in the analyzed calluses.
In stems for which auxin was applied exogenously below the surface of the cut
(2 mm), the first non-lignified cells at the border of the growth ring are parenchymatous,
with only the subsequent cells showing a structure of tracheary elements typical for
the secondary xylem of willow. Similar parenchymatous structures were described in
the xylem of wounded stems in other tree species (Lev-Yadun & Aloni 1991, 1993;
Lev-Yadun 1994, 2002). The results obtained in these studies may suggest that the
appearance of a parenchyma cell layer in this region was related to the wounding it-
self. Thus, in the case of willow we may expect that the parenchyma layer was due to
wounding rather than to an apically applied synthetic auxin. Image processing software
based on a structure tensor in this region of the tissue showed that before the typical
xylem tissue began to develop in the parenchyma layer, the ordering degree of cells
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 397
increased, as indicated by higher values of coherency coefficients. Higher coherency
coefficient values found for the region of parenchyma cells near the rays in the second-
ary xylem formed outside the parenchyma layer indicate that cellular ordering precedes
the regrowth of secondary xylem.
The obtained results showed that digital image analysis software based on a struc-
ture tensor appears to be a useful tool for measuring the degree of cellular ordering in
various regions of regenerating plant tissue. It seems that a wider application of this
tool may create new opportunities for a more comprehensive approach in research on
complex mechanisms responsible for the development of cell and tissue patterns in
plants.
Acknowledgements
The author wishes to thank Professor Pieter Baas and an anonymous reviewer for the constructive
comments and very kind co-operation which substantially improved the quality of the manuscript.
References
Barthelemy D & Caraglio Y. 2007. Plant architecture: a dynamic, multilevel and comprehensive
approach to plant form, structure and ontogeny. Ann. Bot. 99: 375–407.
Biggs AR. 1986. Phellogen regeneration in injured peach tree bark. Ann. Bot. 57: 463–470.
Bigun J, Bigun T & Nilsson K. 2004. Recognition by symmetry derivatives and the general-
ized structure tensor. IEEE Trans. Pattern Anal. Mach. Intell. 26: 1590–1605.
Brown CL. 1964. The influence of external pressure on the differentiation of cells and tissues
cultured in vitro. In: Zimmermann MH (ed.), The formation of wood in forest trees: 389–
404. Academic Press, New York.
Brown CL & Sax K. 1962. The influence of pressure on the differentiation of secondary tissues.
Amer. J. Bot. 49: 683–691.
de Klerk G, Arnold-Schmitt B, Lieberei R & Neumann KH. 1997. Regeneration of roots, shoots
and embryos: physiological and biochemical aspects. Biol. Plant. 39: 53–66.
Du Juan, Xie Hong-Li, Zhang De-Qiang, He Xin-Qiang, Wang Min-Jie, Li Ying-Zhang, Cui
Ke-Ming & Lu Meng-Zhu. 2006. Regeneration of the secondary vascular system in poplar
as a novel system to investigate gene expression by a proteomic approach. Proteomics 6:
881–895.
Dumais J. 2007. Can mechanics control pattern formation in plants? Curr. Opin. Plant Biol.
10: 58–62.
Fahn A. 1988. Secretory tissues and factors influencing their development. Phyton 28: 13–26.
Fahn A. 1990. Plant anatomy. Ed. 4. Pergamon Press, Oxford.
Feynman RP, Leighton RB & Sands M. 1964. The Feynman lectures on physics. Vol. 2. Part 12.
Addison-Wesley, Reading, Mass.
Fonck E, Feigl GG, Fasel J, Sage D, Unser M, Rufenacht DA & Stergiopulos N. 2009. Effect
of aging on elastin functionality in human cerebral arteries. Stroke 40: 2525–2556.
Fuchs M, Bel AJE & Ehlers K. 2011. Do symplasmic networks in cambial zones correspond
with secondary growth patterns? Protoplasma 248: 141–151.
Hansen H & Grossmann K. 2000. Auxin induced ethylene triggers abscisic acid biosynthesis
and growth inhibition. Plant Physiol. 124: 1437–1448.
Hejnowicz Z. 2002. Anatomia i histogeneza roślin naczyniowych (Anatomy and histogenesis of
vascular plants). Wydawnictwo Naukowe PWN, Warszawa [in Polish].
Downloaded from Brill.com02/21/2022 09:07:16PM
via free access398 IAWA Journal 36 (4), 2015
Hejnowicz Z. 2011. Plants as mechano-osmotic transducers. In: Wojtaszek P (ed.), Mechanical
integration of plant cells and plants: 241–267. Springer, Berlin, Heidelberg.
Jahne B. 1993. Spatio-temporal image processing: theory and scientific application. Springer,
Berlin.
Klebe RJ, Overfelt TM, Magnuson VL, Steffensen B, Chen DL & Zardeneta G. 1991. Quan-
titative assay for morphogenesis indicates the role of extracellular matrix components and
G proteins. Proc. Natl. Acad. Sci. 88: 9588– 9592.
Koo AJ & Howe GA. 2009. The wound hormone jasmonate. Phytochemistry 70: 1571–1580.
Lachaud S, Catesson AM & Bonnemain JL. 1999. Structure and function of vascular cambium.
Life Sci. 322: 633–650.
Larson PR. 1994. The vascular cambium: development and structure. Springer, Berlin, Heidel-
berg, New York.
Lev-Yadun S. 1994. Experimental evidence for the autonomy of ray differentiation in Ficus
sycomorus L. New Phytol. 126: 499–504.
Lev-Yadun S. 2001. Intrusive growth – the plant analog to dendrite and axon growth of animals.
New Phytol. 150: 508–512.
Lev-Yadun S. 2002. The distance to which wound effects influence the structure of secondary
xylem of decapitated Pinus pinea. J. Plant Growth Regul. 21: 191–196.
Lev-Yadun S & Aloni R. 1991. An experimental method of inducing “hazel” wood in Pinus
halepensis (Pinaceae). IAWA Bull. n.s. 12: 445–451.
Lev-Yadun S & Aloni R. 1993. Effect of wounding on the relations between vascular rays and
vessels in Melia azedarach L. New Phytol. 124: 339–344.
Makino R, Kuroda H & Shimaji K. 1983. Callus formation, and effects of applied pressure to
the cultured cambial explant of sugi (Cryptomeria japonica D.Don). Wood Research: Bull.
Wood Research Institute Kyoto University 69: 1–11.
Morgan PW & Drew MC. 1997. Ethylene and plant responses to stress. Physiol. Plant. 100:
620–630.
Nishitani C, Demura T & Fukuda H. 2002. Analysis of early processes in wound-induced vas-
cular regeneration using TED3 and ZeHB3 as molecular markers. Plant Cell Physiol. 43:
79–90.
Oborny B. 2004. External and internal control in plant development. Complexity 9: 22–28.
Plomion C, Leprovost G & Stokes A. 2001. Wood formation in trees. Plant Physiol. 127:
1513–1523.
Plomion C, Pionneau C, Brach C, Costa P & Baillères H. 2000. Compression wood-responsive
proteins in developing xylem of maritime pine (Pinus pinaster Ait). Plant Physiol. 123:
959–969.
Rezakhaniha R, Agianniotis A, Schrauwen JTC, Griffa A, Sage D, Bouten CVC, van de Vosse
FN, Unser M & Stergiopulos N. 2012. Experimental investigation of collagen waviness and
orientation in the arterial adventitia using confocal laser scanning microscopy. Biomech.
Model Mechanobiol. 11: 461– 473. doi: 10.1007/s10237-011-0325.
Romberger JA, Hejnowicz Z & Hill JF. 1993. Plant structure: function and development, a trea-
tise on anatomy and vegetative development, with special references to woody plants.
Springer, Berlin. xix + 298 pp.
Sablowski R. 2007. The dynamic plant stem cell niches. Curr. Opin. Plant Biol. 10: 639–644.
Tocquard K, Lopez D, Decourteix M, Thibaut B, Julien J-L, Label P, Leblanc-Fournier N,
Roeckel-Drevet P. 2014. The molecular mechanisms of reaction wood formation. In:
Gardiner B, Barnett J, Saranpää P & Grill J (eds.), The biology of reaction wood: 107–138.
Springer, Berlin, Heidelberg.
Unser M, Aldroubi A & Eden M. 1993. B-spline signal processing. Part I. Theory. IEEE Trans.
Signal. Process. 41: 821–833.
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessZajączkowska – Cellular arrangement and xylogenesis in wound callus 399
Volfson D, Cookson S, Masty J & Tsmiring LS. 2008. Biomechanical ordering of dense cell
population. Proc. Natl. Acad. Sci. 105: 15346–15354.
Yoshii H & Imaseki H. 1981. Biosynthesis of auxin-induced ethylene. Effects of indole-3-acetic
acid, benzyladenine and abscisic acid on endogenous levels of 1-aminocyclopropane-1-
carboxylic acid (ACC) and ACC synthase. Plant Cell Physiol. 22: 369–379.
Zajączkowska U. 2014a. Regeneration of Scots pine stem after wounding. IAWA J. 35: 270–
280.
Zajączkowska U. 2014b. Overgrowth of Douglas fir (Pseudotsuga menziesii Franco) stumps
with regenerative tissue as an example of cell ordering and tissue reorganization. Planta
240: 1203–1211.
Zhang Y, Sederoff RR & Allona I. 2000. Differential expression of genes encoding cell wall
proteins in vascular tissues from vertical and bent loblolly pine trees. Tree Physiol. 20:
457– 466.
Accepted: 7 September 2015
Downloaded from Brill.com02/21/2022 09:07:16PM
via free accessYou can also read