Role of Jasmonic and Salicylic Acid on Enzymatic Changes in The Root of Two Alyssum Inatum Náyr. Populations Exposed to Nickel Toxicity
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Role of Jasmonic and Salicylic Acid on Enzymatic Changes
in The Root of Two Alyssum Inflatum Náyr. Populations
Exposed to Nickel Toxicity
Shiva Najafi Kakavand
Razi University of Kermanshah: Razi University
Naser Karimi
Razi University of Kermanshah: Razi University
Hamid-Reza Ghasempour
Razi University of Kermanshah: Razi University
Ali Raza
Chinese Academy of Agricultural Sciences
Mehrdad Chaichi
Hamedan Agricultural and Natural Resources Research and Education Center
Masoud Modarresi ( mmodarresi@kums.ac.ir )
Kermanshah University of Medical Sciences https://orcid.org/0000-0001-6722-3434
Research Article
Keywords: Alyssum inflatum, Antioxidant defense, detoxification, Oxidative stress, Plant hormones, toxic metals.
Posted Date: April 27th, 2021
DOI: https://doi.org/10.21203/rs.3.rs-435393/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License
Page 1/24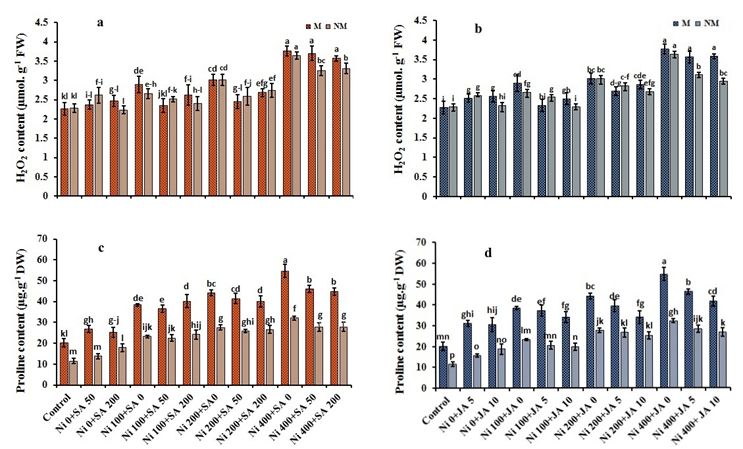
Abstract
Salicylic acid (SA) and jasmonic acid (JA) as plant growth regulators (PGRs) have the potential to ameliorate plant
development and tolerance to deleterious effects of toxic metals like nickel (Ni). Therefore, the current study was carried
out to evaluate SA and JA's interactive effect on the root antioxidative response of two Alyssum inflatum Nyár. populations
against Ni-toxicity. Two A. inflatum species under Ni-stress conditions (0, 100, 200, and 400 µM) were exposed to alone or
combined levels of SA (0, 50, and 200 µM) and JA (0, 5, and 10 µM) treatments. Results showed that high Ni doses
reduced the roots fresh weight (FW) in two populations than control; however, the use of external PGRs had ameliorated
roots biomass by mitigated Ni-toxicity. Under Ni toxicity, SA and JA, especially their combination, induced high Ni
accumulation in plants' roots. Moreover, the application of SA and JA alone, as well as combined SA + JA, was found to be
effective in the scavenging of hydrogen peroxide (H2O2) by improving the activity of superoxide dismutase (SOD),
peroxidase (POD), catalase (CAT), and ascorbate peroxidase (APX) in both populations under Ni-toxicity. Overall, our
results manifest that SA and JA's external use, especially combined SA + JA treatments, ameliorate root biomass and plant
tolerance by restricting translocation Ni to the shoot, accumulating in roots, and also enhancing antioxidant defense
systems.
Introduction
In the past decades, rapid mechanization gives rise to environmental pollution by heavy metals, which is considered an
essential universal concern currently (Ramírez et al. 2021). A considerable amount of this pollution, such as nickel (Ni), has
been entered the environment via the activities of both anthropogenic and natural origins (Ali et al. 2019). However, Ni as
trace elements exists in natural soils, except in serpentine soils (Nagajyoti et al. 2010). Besides, Ni is an essential
microelement (in the lower amount of 0.05–10 mg kg− 1 dry biomass) for plant growth. Naturally, it exists in the structure
of some metalloenzymes, including urease, glyoxalase-I, and a few superoxide dismutase (SOD), required for nitrogen
metabolism in higher plants (Shahzad et al. 2018; Hassan et al. 2019; Leškova et al. 2020). However, exceeding the
accumulation of Ni in plant tissues leads to toxicity in the plant and can negatively affect plenty of the plant's
physiological and biochemical processes (Kosakivska et al. 2021). High Ni concentrations indirectly cause increased
reactive oxygen species (ROS) production in plant cells (Silva et al. 2020). The high ROS generation in plant cells caused
redox imbalance, disrupt enzyme activities, damage the structure of lipids and proteins, oxidative stress, and eventually cell
death (Chaki et al. 2020). Subsequently, plant cells trigger antioxidant mechanisms to tolerate Ni-toxicity and combat
oxidative stress (Amjad et al. 2019). For instance, excess Ni concentration in mustard plants (Abd_Allah et al. 2019),
Brassica juncea (Khan et al. 2020), and Solanum lycopersicum L. (Jahan et al. 2020) leads to reduced growth traits by
photosynthesis inhibition, the nutrients' absorption disruption, and oxidative stress due to ROS generation. Thus, they
triggered antioxidant defense mechanisms to overcome the Ni-induced oxidative stress, including improving the activity of
the antioxidant enzymes such as SOD, catalase (CAT), and ascorbate peroxidase (APX) as well as raising proline levels as
a non-enzymatic antioxidant compound.
Likewase, when plants are exposed to abiotic stresses such as heavy metal, they use various stress tolerance strategies,
including regulating the levels of endogenous hormones and their interaction pathways (Emamverdian et al. 2020). Plant
endogenous hormones play a vital role in regulating responses to an extended range of internal and environmental stimuli
(Mubarik et al. 2021). Jasmonic acid (JA) and salicylic acid (SA), arbitrated signaling pathways have been shown as
potential tools in increasing plant tolerance to stress conditions (Caarls et al. 2015). Jasmonates [JA and methyl
jasmonate (MeJA)] are considered endogenous-growth substances that play a significant role by modulating signal
transduction mechanisms under stress conditions in numerous plant species (Raza et al. 2020). Many studies have been
shown that JA and MeJA supplementation can recover the toxic effects of heavy metals in plants that have been
described in diverse plants such as like soybean (Glycine max) (Keramat et al. 2009) and Mentha arvensis (Zaid and
Page 2/24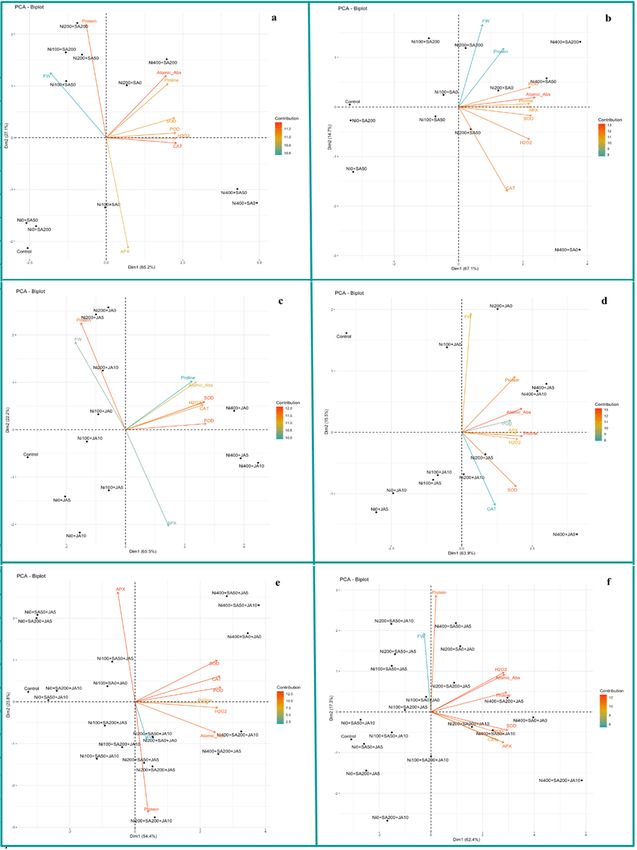
Mohammad 2018) under high cadmium (Cd) concentration, and Lemna valdiviana under arsenic (As) toxicity (Coelho et
al. 2020).
Likewise, SA is an important signaling molecule contributing to the fine-tuning of multiple physiological processes in
plants. It is known as a stress messenger involved in abiotic stress signaling, as a plant defense responds to heavy metal
toxicity (Zaid et al. 2019; Rhaman et al. 2020). Numerous studies show SA has a role in dropping the heavy metals toxicity
in plants. For instance, a study by Wang et al. (2009) indicated that SA supplementation in maize (Zea mays L.) reduced
the toxic effect of Ni by increasing APX and SOD activities and decrease hydrogen peroxide (H2O2) contents. Exogenously
applied SA remarkably improved antioxidative enzymes' activities in Lemna minor under cadmium (Cd) toxicity (Lu et al.
2018) and Nymphaea tetragona (Gu et al. 2018), Sorghum bicolor L. under chromium (Cr) toxicity (Sihag et al. 2019),
Brassica campestris under lead (Pb) toxicity (Hasanuzzaman et al. 2019), and watermelon plants under boron (B) stress
(Moustafa-Farag et al. 2020).
The adjustment of plant defense mechanisms against biotic and abiotic stresses is determined by altering the
concentration ratio of various interacting endogenous hormones, also the effect of each hormone on the endogenous
concentration of others (Avalbaev et al. 2016; Mubarik et al. 2021). It is well-identified that the antagonistic effect has
between the JA and SA cross-talk in various plants against biotic stress. However, there is a JA-SA cross-talk synergistic
effect against biotic stress in rice (Oryza sativa L.) (Tong et al. 2012; Liu et al. 2015). Consequently, the adjustment of JA-
SA signaling pathways' interaction may be critical for particular defensive responses in plants (Liu et al. 2015). According
to our best knowledge and the previous report, no studies have reported SA/JA's interactive effects in plants exposed to
heavy metal toxicity (Najafi Kakavand et al. 2019).
The species of Alyssum inflatum Nyár. species belong to the Brassicaceae family, is the native plant of serpentine and non-
serpentine soil of west of Iran (Ghaderian et al. 2007). Besides, A. inflatum is considered as a Ni-hyperaccumulator plant
(can accumulate Ni > 1000 µg.g− 1 in their dry biomass of leaves) (Reeves et al. 2018; Mohseni et al. 2018). Therefore, the
current study focuses for the first time on the beneficial effects of JA and SA at different concentrations on antioxidant
enzyme activities of roots of two metallicolous (M) and non-metallicolous (NM) A. inflatum species from serpentine and
non-serpentine soils under several Ni doses.
Materials And Methods
Plant material and treatment
A. inflatum (M) seeds, is serpentine soils endemic population, were gathered from Marivan in the west of Iran, and A.
inflatum (NM) seeds, non-serpentine soils population, were harvested from Shahu area in Kermanshah, Iran (Ghaderian et
al. 2007; Najafi Kakavand et al. 2019). Overall, 108 pots for each population (a total of 216 pots for both populations) were
used to perform this experiment. Originally, each replicate contained eight seeds of A. inflatum in a pot (450-ml) filled with
perlite: sand (2:1) mixture individually. The pots were irrigated daily with tap water till germination and subsequently with
Hoagland's solution as described by Najafi kakavand et al. (2019). All A. inflatum seedling was grown in a growth chamber
environmentally controlled conditions (25 − 20°C, 16/8 h light/dark cycle, PPFD of 140 µmol m− 2 s− 1), and the nutrient
solutions were replaced by each 5-days. After six weeks of germination, the 108 pots related to each population were
randomly divided into 36 groups, and each group received one treatment (n = 3 pots for each treatment). Four doses of Ni
and three doses of SA and JA were exerted by applying various solution concentrations of Ni (0, 100, 200, and 400 µM), SA
(0, 50, and 200 µM), and JA (0, 5, and 10 µM). Treatments were performed as a mixture of Ni and SA with Hoagland
solution and JA as a foliar spray for three weeks. Plants were harvested 21 days after exposure, and then divided into
shoot and root, and roots fresh weight (FW) were measured. Eventually, roots material was kept at − 70 ᵒC for further
examination.
Page 3/24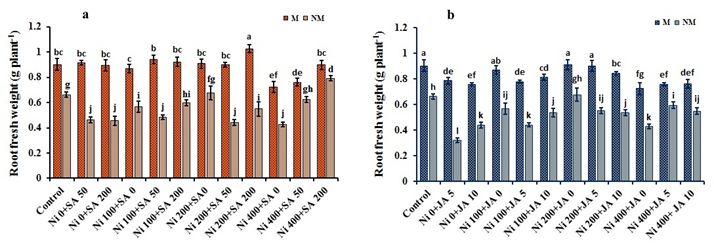
Nickel contents evaluation
Nickel concentration was measured according to Ghasemi et al. (2009). Briefly, Ni concentrations of roots were determined
by digesting 50 mg of dried material of roots in 2 ml of nitric acid (65%) overnight, followed by incubating at 90°C for 4 h.
Subsequently, 1 ml H2O2 (30%) was added to the samples, and the mixture was heated again at 90°C until the digests had
clarified. The volume of samples was adjusted to 10 ml with distilled water. Finally, the samples were analyzed for Ni
concentration measurement by atomic absorption spectrophotometry (ASS 6200, Shimadzu, Japan).
Measurement of H2O2 concentration
Hydrogen peroxide (H2O2) contents were measured according to the Sergiev et al. (2000) method. Briefly, H2O2 was
extracted from 50 mg fresh root materials by homogenization with 5 ml of cooled trichloro-acetic acid (TCA) [0.1% (W/V)].
The homogenates were centrifuged at 12000xg (15 min, 4°C). To measurement, the H2O2 content, the upper solution (0.5
ml) was mixed with 0.5 ml phosphate buffer (200 mM, pH 7.0) and 1ml potassium-iodide (1 M), and the mixture
absorbance was recorded at 390 nm.
Extraction and Measurement of Enzyme activities
The root extraction was produced by homogenizing fresh root material (100 mg) with phosphate buffer (50 mM; pH 7.5)
consisting of 1mM ethylene-diamine-tetraacetic acid (EDTA) and polyvinylpyrrolidone (PVP) [% 1(W/V)], and afterwards
centrifuged (13,000×g; 15 min; 4°C). The upper solution was kept for enzyme activity assays.
The superoxide dismutase (SOD) activity was assayed using Giannopolitis and Ries (1977) method by measuring a
decline in nitro-blue-tetrazolium (NBT) photo-reduction at 560 nm. The reaction solution (3 ml) consisted of 50 µl of
enzyme solution, phosphate buffer (50 mM; pH 7.8), 13 mM methionine, 0.2 mM riboflavin, 1 mM NBT, 10 mM EDTA.
The catalase (CAT) activity was assayed using Aebi (1983) process, the process of catalase (CAT) activity was assayed.
The assay solution included 800 µl of phosphate buffer (50 mM; pH 7.0), 100 µl of 30mM H2O2, and 100 µl of enzyme
solution. The disintegration of H2O2 was monitored by a drop in absorbance at 240 nm. The enzyme activity was stated as
U mg− 1 protein.
The peroxidase (POD) was examined by the technique of Chance and Maehly (1955). The reaction solution consisted of
100 µl enzyme solution, phosphate buffer (200 mM; pH 7), guaiacol (1%), and 1mM H2O2. The oxidation of guaiacol was
monitored at 470 nm using an extinction coefficient of 26.6 mM− 1cm− 1.
The ascorbate peroxidase (APX) activity was determined according to Nakano and Asada (1981) method. The APX assay
included enzyme solution (50µl), phosphate potassium buffer (50 mM; pH 7), 1 mM EDTA, 5 mM ascorbate, and 1mM
H2O2. The oxidation of ascorbate was monitored at 290 nm using an extinction coefficient of 2.8mM− 1cm− 1. Finally, the
enzyme activity for all enzymes examined was stated as U. mg− 1 protein.
The total protein in the roots was measured based on the method of Bradford (1976). Protein contents were measured
according to a standard curve made using Bovine-serum albumin.
Proline concentration
Proline levels in the roots were estimated based on the Bates et al. (1973) method. Initially, dry roots materials (50 mg)
were extracted with 10 ml 3% sulfosalicylic acid, and the mixture was centrifuged (1700 g; 10 min). Then, to 0.5 ml of
upper solution, 0.5 ml glacial acetic acid and 1 ml ninhydrin reagent was added and held in a water bath (100°C) for 1 hr.
Finally, the reaction was terminated by placing the sample tubes into a cold bath. The product was extracted by combining
Page 4/24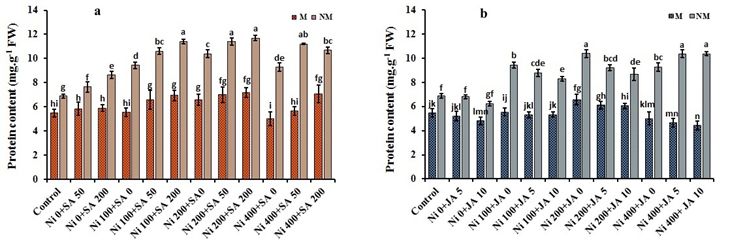
1 ml of toluene to the reaction solution by vortexing for 1 min. Finally, the upper phase's absorbance is recorded at 520 nm,
and proline concentrations are calculated from a standard graph of L-proline.
Statistical analysis
The GLM procedure of SAS was exerted for statistical analysis of the variance (one-way ANOVA) of each data set, and
Duncan’s test (at P ≤ 0.05) was exerted to compare the mean values of treatments. For a better understanding of treatment
effects, data analysis of variance was performed in three separate groups. In one group, all Ni and SA levels were
considered, while in another group, analysis of variance was performed on all Ni and JA levels treatments. The third group
of analyses included the three-way effects of Ni, SA, and JA. In this group, SA0 and JA0 were also used in all Ni levels to
perform the analysis of variance with 20 treatments. The data stated are mean ± SD and n = 3. Also, principal component
analysis (PCA) was done applying the fviz-pca function of the factoextra R package ver. 1.0.7 (Kassambara 2017) to
visually biplot treatments and variables.
Results
Root fresh weight
At 100-µM and 200-µM Ni-treatment, root fresh weight (FW) of both M and NM plants did not indicate significant
differences with control plants (p ≤ 0.05). In contrast, root FW declined by 19.8% and 35.5% at 400-µM Ni concentration in
M and NM species of A. inflatum, respectively, compared to control plants (Fig. 1a, b). Application of 200-µM SA on the
plantsʼ exposure to 400-µM Ni, roots FW incremented 1.26-and 1.85-times in M and NM species respectively, compared to
only 400-µM Ni-treated plants. Similar results were obtained with 10-µM JA in plants under 400-µM Ni-toxicity so that a
1.28-fold increase in root FW of NM plants was observed in comparison to 400-µM Ni treatment. Application of 200-µM SA
+ 10-µM JA on M population increased the roots FW by 11.74% under 400-µM-Ni stress than control plants. Conversely, on
the other hand, supplementation of 50-µM SA + 5-µM JA on the NM population enhanced the root FW by 7.25% with 400-
µM-Ni stress relative to the controls (Table 1).
Page 5/24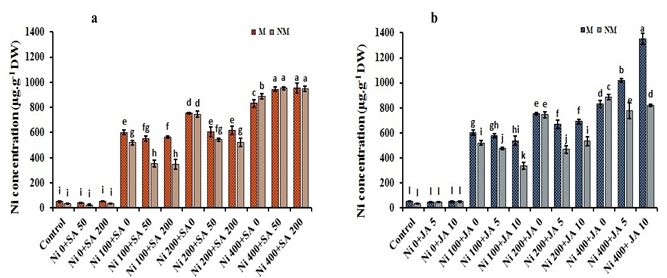
Table 1
Influence of combined SA + JA on Ni contents, FW, protein levels, H2O2 levels, and proline contents in roots of M/NM
populations of A.inflatum exposed to Ni-toxicity
Treatments Ni concentration FW Protein content H2O2 content Proline content
(µg g− 1 DW) (g plant− 1) (mg g− 1 FW) (µmol g− 1 FW) (µg g− 1 DW)
M NM M NM M NM M NM M NM
Control 50.9 34.5 ± 0.90 ± 0.66 ± 5.47 ± 6.87 ± 2.26 ± 2.28 ± 20.1 ± 11.5 ±
± 3.74q 0.04fg 0.02opq 0.34nop 0.17i − l 0.16rs 0.10rs 2.01opq 1.17u
4.03q
(Ni0 + 46.5 46.4 ± 0.68 ± 0.54 ± 4.77 ± 8.15 ± 2.35 ± 2.21 ± 19.0 ± 15.6 ±
SA50 + ± 4.03q 0.02l − o 0.02tu 0.24pq 0.33h 0.14qr 0.03rs 2.13o − 0.92t
JA5) µM 4.14q r
(Ni0 + 51.8 40.0 ± 0.67 ± 0.62 ± 5.68 ± 8.26 ± 2.73 ± 2.28 ± 20.1 ± 16.6 ±
SA50 + ± 5.56q 0.03nop 0.02qr 0.46mno 0.21hg 0.10k − o 0.03rs 1.67opq 0.36st
JA10) µM 3.81q
(Ni0 + 51.9 40.0 ± 0.81 ± 0.53 ± 4.56 ± 6.76 ± 2.17 ± 2.25 ± 25.8 ± 15.0 ±
SA200 + ± 4.58q 0.02h 0.04tu 1.10q 0.22i − l 0.07s 0.07rs 1.19jkl 1.51t
JA5) µM 3.89q
(Ni0 + 52.8 47.1 ± 1.07 ± 0.38 ± 5.41 ± 6.19 ± 2.51 ± 2.22 ± 23.2 ± 17.1 ±
SA200 + ± 4.36q 0.01b 0.05w 0.64nop 0.20j − 0.16pq 0.08rs 1.33m 0.98rst
JA10) µM 3.18q n
(Ni100 + 602 ± 520 ± 0.86 ± 0.56 ± 5.54 ± 9.43 ± 2.89 ± 2.66 ± 38.3 ± 23.2 ±
SA0 + JA0) 18.4k 19.3l 0.03g 0.04st 0.34m − 0.25def 0.21h − l 0.12m − 0.73d 0.56m
µM p p
(Ni100 + 797 ± 242 ± 0.72 ± 0.65 ± 5.23 ± 10.4 ± 2.82 ± 2.64 ± 26.0 ± 18.3 ±
SA50 + 9.77i 16.0p 0.01kl 0.03opq 0.64opq 0.39bc 0.04i − m 0.11nop 1.45jk 1.25qrs
JA5) µM
(Ni100 + 838 ± 254 ± 0.76 ± 0.32 ± 6.58 ± 10.2 ± 2.73 ± 2.73 ± 23.9 ± 16.6 ±
SA50 + 19.7gh 25.5op 0.01ijk 0.01x 0.66i − l 0.28bcd 0.07l − o 0.05l − o 1.16klm 1.06st
JA10) µM
(Ni100 + 877 ± 259 ± 0.97 ± 0.63 ± 5.74 ± 8.16 ± 2.57 ± 2.70 ± 27.1 ± 20.6 ±
SA200 + 28.8fg 24.0op 0.01de 0.01qpr 0.33mno 0.51h 0.07op 0.07mno 0.92ij 0.92op
JA5) µM
(Ni100 + 895 ± 273 ± 1.16 ± 0.49 ± 6.09 ± 7.29 ± 2.70 ± 2.78 ± 27.3 ± 23.8 ±
SA200 + 61.6f 23.0op 0.02a 0.04uv 0.49lmn 0.12i 0.04mno 0.06j − n 0.92ij 0.31lm
JA10) µM
(Ni200 + 753 ± 745 ± 0.91 ± 0.67 ± 6.58 ± 10.4 ± 3.01 ± 3.01 ± 44.1 ± 27.6 ±
SA0 + JA0) 8.05j 24.5j 0.03fg 0.05m 0.45i − l 0.33bc 0.14fgh 0.14fgh 1.46b 1.18ij
µM −p
(Ni200 + 963 ± 288 ± 0.73 ± 0.49 ± 6.96 ± 12.0 ± 3.03 ± 2.78 ± 28.6 ± 21.1 ±
SA50 + 24.9e 69.0no 0.03jk 0.01uv 0.38ij 0.66a 0.09fgh 0.12j − n 0.65hi 1.33no
JA5) µM
(Ni200 + 976 ± 357 ± 0.77 ± 0.67 ± 6.29 ± 11.8 ± 2.91 ± 2.81 ± 30.3 ± 18.8 ±
SA50 + 37.4e 12.5m 0.02hij 0.03nop 0.36j − m 0.48a 0.09hij 0.09j − n 0.73gh 0.88pqr
JA10) µM
Page 6/24
Treatments Ni concentration FW Protein content H2O2 content Proline content
(µg g− 1 DW) (g plant− 1) (mg g− 1 FW) (µmol g− 1 FW) (µg g− 1 DW)
M NM M NM M NM M NM M NM
(Ni200 + 1034 322 ± 0.79 ± 0.59 ± 6.88 ± 10.5 ± 2.90 ± 2.95 ± 31.6 ± 26.0 ±
SA200 + ± 22.5mn 0.01hi 0.02rs 0.50ijk 0.44b 0.08h − k 0.07hij 1.67fg 1.07j
JA5) µM 12.7d
(Ni200 + 986 ± 344 ± 0.93 ± 0.47 ± 8.21 ± 9.67 ± 2.88 ± 2.98 ± 33.4 ± 27.3 ±
SA200 + 18.2e 27.0m 0.02ef 0.02v 0.26h 0.38cde 0.01h − l 0.05ghi 1.76ef 1.55ij
JA10) µM
(Ni400 + 831 ± 886 ± 0.72 ± 0.42 ± 5.01 ± 9.28 ± 3.763 ± 3.64 ± 54.6 ± 32.1 ±
SA0 + JA0) 29.1hi 19.5f 0.04klm 0.02w 0.55opq 0.36ef 0.12a 0.09ab 3.10a 0.96fg
µM
(Ni400 + 1268 743 ± 0.74 ± 0.71 ± 5.07 ± 11.6 ± 3.17 ± 3.00 ± 35.6 ± 22.8 ±
SA50 + ± 9.50j 0.03jk 0.03k − 0.46opq 0.70a 0.07def 0.15gh 1.24e 1.39mn
JA5) µM 16.2b n
(Ni400 + 1310 818 ± 0.93 ± 0.53 ± 4.55 ± 8.87 ± 3.14 ± 2.92 ± 34.6 ± 22.9 ±
SA50 + ± 37.5hi 0.02ef 0.02tu 0.34q 0.47fgh 0.06efg 0.09hij 1.25e 1.04mn
JA10) µM 39.8a
(Ni400 + 1151 803 ± 0.99 ± 0.57 ± 6.70 ± 9.00 ± 3.21 ± 3.48 ± 41.8 ± 27.4 ±
SA200 + ± 25.0hi 0.01cd 0.04st 0.35i − l 0.63efg 0.188de 0.05bc 0.97c 1.68ij
JA5) µM 37.3c
(Ni400 + 1345 900 ± 1.02 ± 0.55 ± 6.17 ± 6.97 ± 3.30 ± 3.32 ± 39.4 ± 31.3 ±
SA200 + ± 21.0f 0.01c 0.03st 30.42k 0.96ij 0.11de 0.010cd 1.21d 1.58fg
JA10) µM 16.9a −n
Data are introduced as a mean of each treatment group ± SE (n = 3). Different letters presented statistical differences in
means by the Duncan test (at p ≤ 0.05).
Root Ni concentration
After three weeks of treatment, M and NM populations of A. inflatum accumulated Ni significantly in the roots when treated
with 400-µM Ni concentration (Fig. 2a, b). Salicylic acid application increased Ni accumulation by about 1.2-folds in both
populations, compared to only 400-µM Ni-treated plants. Likewise, increasing JA concentration upsurges the Ni contents in
the roots of the M population. Although, in NM plants treated with 400-µM Ni + 5-µM JA and 400-µM Ni + 10-µM JA, Ni
levels were dropped by 12.3% and 7.33%, respectively, compared with Ni 400-µM treatment alone. Additionally, exogenous
SA supply and foliar application of JA at the same time have positively affected the Ni-accumulation in the roots of both
populations (Table 1). The average increase in root Ni contents, under 400-µM Ni + 200-µM SA + 10-µM JA treatment, was
recorded as 38.2% in M and 1.50% in NM plants, concerning 400-µM Ni treatment.
Protein contents in roots
A moderate Ni (200-µM) dose increased the protein contents by 16.8% and 33.8% in M and NM species' roots, respectively,
than control plants (Fig. 3a, b). With the SA and/or JA application, protein contents increased under 400-µM Ni stress in the
roots of M/NM plants. Unlike this, the exogenous JA decreased the protein level in M populations' roots with an increase in
Ni doses. Importantly, the external application of SA and JA in M/NM plants at the same time, increased the protein
content, especially under 200-µM Ni stress. For example, the highest protein levels were observed in the simultaneous
treatment of 200-µM Ni + 200-µM SA + 10-µM JA by 33.3% in the root of M population, and 200-µM Ni + 50-µM SA + 5-µM
JA by 42.9% in the root of NM populations, compared to the root of control plants (Table 1).
Page 7/24Hydrogen peroxide level in root
On day 21 of 400-µM Ni-treatment, excised roots of M/NM plants demonstrated a significant accumulation of H2O2, which
was not observed in control plants (Fig. 4a,b). Whereas the external application of SA and/or JA in plants exposed to Ni,
we observed a declining trend in H2O2 content in roots of M/NM populations. Additionally, the simultaneous application of
SA + JA in plants exposed to Ni showed a decreasing trend in H2O2 levels (Table 1). For instance, in the treatment of 400-
µM Ni + 200-µM SA + 10-µM JA, the H2O2 content decreased by 12.2% and 8.22% in M/NM plants compared with plants
treated with 400-µM Ni.
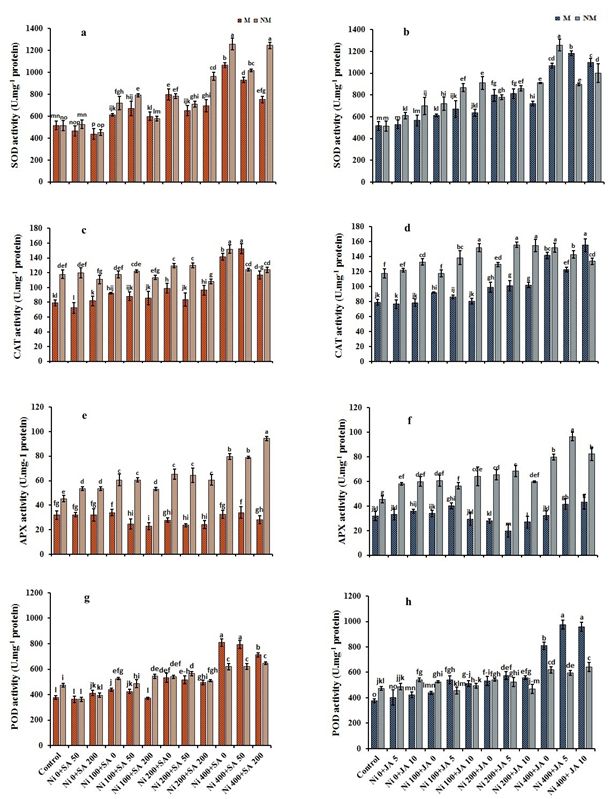
The activity of antioxidant enzymes
High doses of Ni indirectly lead to the generation of H2O2, which in turn stimulates the activity of antioxidative enzymes
(SOD, CAT, POD, and APX) under Ni stress in M/NM plants (Fig. 5). However, the AXP activity in the roots of M-population
exposed to high Ni doses did not significantly change compared to control plants (Fig. 5e,f). The highest Ni level (400-µM
Ni) caused a maximal increment in the antioxidative enzyme activities, i.e., 51.6% and 59.2% (SOD), 44.1% and 22.4%
(CAT) and 53.5% and 23.6% (POD) in M and NM plants, respectively, compared to the control plants. Likewise, the APX
activity increased by 43.1% at 400-µM Ni concentration in NM plants, and there was no significant difference between
these treatments compared to the control plants. The activities of four antioxidative enzymes in the roots of M/NM
populations exposed to different Ni concentrations were changed in response to SA and JA alone and the simultaneous
application of SA + JA. Percent of activities of SOD, POD, and CAT was remarkably improved with the application of SA
and/or JA than the control plants in both populations. The highest enzyme activity, i.e., 58.1% (SOD), 57.3% (CAT), 21.0%
(APX), and 61.9% (POD) were recorded in the roots of M plants by 400-µM Ni + 50-µM SA + 10-µM JA treatment whit
respect to the control plants. In contrast, the highest enzymes activity, i.e., 69.0% (SOD), 53.9% (CAT), 48.8% (APX), and
35.8%(POD) in the roots of NM plants were observed under 400-µM Ni + 50-µM SA + 10-µM JA treatment whit respect to the
control plants (Table 2).
Page 8/24Table 2
Influence of combined SA + JA on antioxidative enzymes activity in roots of M/NM populations of A.inflatum exposed to
Ni-toxicity
SOD activity APX activity POD activity CAT activity
(U mg− 1protein) (U mg− 1protein) (U mg− 1protein) (U mg− 1protein)
Treatments M NM M NM M NM M NM
Control 516 ± 513 ± 31.9 ± 45.3 ± 376 ± 473 ± 79.1 ± 117 ±
41.1st 45.8st 3.57mno 2.72ij 13.8st 15.0l − p 3.84r 6.01h − l
(Ni0 + SA50 + JA5) 664 ± 399 ± 38.8 ± 52.8 ± 415 ± 500 ± 84.8 ± 113 ±
µM 17.6n − q 34.9n 2.68kl 1.11gh 30.8p − s 31.5i − m 8.51pqr 4.74jkl
(Ni0 + SA50 + 504 ± 495 ± 28.3 ± 49.0 ± 442 ± 408 ± 78.4 ± 130 ±
JA10) µM 60.8t 30.0t 3.29nop 4.62hi 47.4m − r 36.4q − t 11.2r 13.4fg
(Ni0 + SA200 + 569 ± 747 ± 39.7 ± 55.6 ± 441 ± 395 ± 84.1 ± 114 ±
JA5) µM 63.9rst 102k − n 1.91k 2.45fg 52.0n − r 27.1rst 4.31qr 9.98jkl
(Ni0 + SA200 + 627 ± 683 ± 33.1 ± 64.9 ± 454 ± 474 ± 79.3 ± 121 ±
JA10) µM 21.1o − r 81.2nop 3.56mn 0.69de 39.3m − q 21.6k − p 6.92r 8.50g − k
(Ni100 + SA0 + 609 ± 719 ± 33.9 ± 60.6 ± 438 ± 526 ± 92.2 ± 117 ±
JA0) µM 12.4pqr 60.4lmn 2.52lm 4.65ef 13.5n − r 8.32h − l 0.85opq 4.77h − l
(Ni100 + SA50 + 725 ± 572 ± 33.5 ± 52.8 ± 557 ± 471 ± 99.4 ± 127 ±
JA5) µM 25.8k − n 39.6rst 2.75m 2.26gh 60.7ghi 23.0l − p 8.13mno 7.82gh
(Ni100 + SA50 + 562 ± 738 ± 23.9 ± 51.7 ± 475 ± 417 ± 96.6 ± 123 ±
JA10) µM 67.7rst 49.8k − n 1.07pqr 4.03gh 36.3k − p 23.9o − s 9094nop 2.08g − j
(Ni100 + SA200 + 594 ± 896 ± 28.2 ± 61.4 ± 572 ± 400 ± 95.9 ± 113 ±
JA5) µM 23.9qrs 30.1ghi 3.84nop 4.41de 42.9gfh 13.5q − t 5.26n − q 4.03jkl
(Ni100 + SA200 + 702 ± 979 ± 27.6 ± 65.8 ± 569 ± 496 ± 97.6 ± 112 ±
JA10) µM 32.0mno 23.5fg 2.79opq 2.60d 66.7gfh 30.6j − n 4.85mno 6.70jkl
(Ni200 + SA0 + 797 ± 780 ± 27.9 ± 65.3 ± 532 ± 540 ± 99.0 ± 129 ±
JA0) µM 53.2jkl 21.8j − m 1.77opq 4.12de 35.1g − k 12.1g − j 6.56mno 2.90fgh
(Ni200 + SA50 + 728 ± 713 ± 23.0 ± 46.5 ± 587 ± 440 ± 108 ± 109 ±
JA5) µM 66.0k − n 82.4l − o 1.63qr 0.81i 35.0gf 23.9n − r 15.1lmn 10.8klm
(Ni200 + SA50 + 812 ± 730 ± 23.5 ± 54.8 ± 585 ± 357 ± 121 ± 109 ±
JA10) µM 47.2ijk 91.1k − n 1.37pqr 3.03g 52.4gf 13.5t 8.14g − k 15.4klm
(Ni200 + SA200 + 784 ± 893 ± 21.8 ± 64.8 ± 573 ± 556 ± 114 ± 155 ±
JA5) µM 50.6j − m 35.9ghi 1.61r 2.96de 30.5gfh 23.0ghi 1.62i − l 4.76d
(Ni200 + SA200 + 712 ± 1169 ± 21.9 ± 62.4 ± 616 ± 531 ± 111 ± 191 ±
JA10) µM 62.9l − o 31.3cd 1.81r 4.50de 20.5f 25.4g − j 6.87kl 7.87bc
(Ni400 + SA0 + 1067 ± 1257 ± 32.5 ± 79.7 ± 809 ± 619 ± 141 ± 151 ±
JA0) µM 21.6ef 52.7bc 3.46mno 2.38b 29.7d 26.1f 4.37ef 5.85de
(Ni400 + SA50 + 1154 ± 953 ± 40.4 ± 65.2 ± 825 ± 537 ± 160 ± 127 ±
JA5) µM 54.8de 110g 5.02jk 3.14de 31.7cd 32.0g − j 7.99d 6.49ghi
Page 9/24SOD activity APX activity POD activity CAT activity
(U mg− 1protein) (U mg− 1protein) (U mg− 1protein) (U mg− 1protein)
(Ni400 + SA50 + 1233 ± 1268 ± 35.3 ± 73.5 ± 987 ± 619 ± 185 ± 161 ±
JA10) µM 69.2bcd 93.1b 1.41klm 5.00c 68.6a 32.74f 4.41c 7.95d
(Ni400 + SA200 + 863 ± 1241 ± 24.3 ± 62.4 ± 881 ± 570 ± 129 ± 202 ±
JA5) µM 20.4hij 40.7bcd 1.48pqr 5.45de 39.5bc 41.2gfh 3.19fgh 5.15b
(Ni400 + SA200 + 911 ± 1657 ± 25.1 ± 88.7 ± 910 ± 737 ± 156 ± 255 ±
JA10) µM 42.1gh 43.2a 1.54pqr 4.97a 57.7b 45.1e 6.77d 6.24a
Data are introduced as a mean of each treatment group ± SE (n = 3). Different letters presented statistical differences in
means by the Duncan test (at p ≤ 0.05).
Root Proline levels
The contents of an important non-enzymatic antioxidant compound, proline, altered remarkably under Ni-toxicity. However,
Ni toxicity increased the proline concentration in a dose-depended manner. The proline levels in the roots of both M/NM
populations increased by 63.5% exposed to Ni 400-µM, compared to the control plants (Fig. 4c, d). However, exogenous SA
and JA increased proline accumulation under high Ni doses in M/NM plants, compared to the control. Despite this, the use
of exogenous SA and/or JA in the roots of both populations exposed to high Ni doses showed a decreasing trend of about
1.25-fold compared to 400-µM Ni-stress alone. Simultaneous treatment with SA and JA in both populations had improved
Ni toxicity by increasing proline levels at 400 µM Ni. For example, the highest proline contents (51.7% and 63.2%) were
observed in the simultaneous treatment of 400-µM Ni + 200-µM SA + 5-µM JA in the root of M population and 400-µM Ni +
200-µM SA + 5-µM JA in the root of NM species, respectively, compared to the root of control plants (Table 1).
Principal Component Analysis (PCA)
The responses of M and NM of A. inflatum populations to SA, JA, and SA + JA treatments under Ni-toxicity were assessed
through the principal component analysis (PCA) score plot. The PCA results of A. inflatum in M and NM population, treated
with SA under Ni toxicity, showed that the first two components explain 92.3% and 81.8% of the total variation, respectively
(Fig. 6a, b). In the M population of A. inflatum, proline, CAT, POD, SOD, H2O2, and Ni concentration were positively
correlated. In contrast, FW and protein showed a slight correlation with the mentioned traits and were negatively correlated
with APX (Fig. 6a). In the NM population, a more positive correlation can be seen among the evaluated traits, but it is
noteworthy that all of these traits are located in the sections where the high levels of Ni are also located, or in the other
word, in the opposite direction to the control (Fig. 6b)
Biplot of A. inflatum in M and NM-population treated with JA under Ni toxicity revealed that the first two components
explain almost 87.7% and 79.4% of the total variance. In both populations, the variables in this evaluation followed a
similar pattern as SA treatment. All enzymes, H2O2, proline, and Ni, positively correlated with each other and strongly
influenced the first component (Fig. 6c, d).
Considering the combination of SA + JA treatment under Ni toxicity, it was shown that the first two components explain
78.2% and 79.7% of the total variance in M and NM populations, respectively (Fig. 6e, f). The highest Ni accumulation in
both populations' roots occurred after the combination of SA + JA treatments, which was associated with the highest
enzymatic activity, proline accumulation, and H2O2.
Discussion
In the current study, the effect of JA and SA supplementation, alone and combined, was assessed for its capability to
restrict Ni's toxic effect on the physiological and biochemical features of both M/NM populations of A. inflatum under Ni-
Page 10/24toxicity. High Ni accumulation in various plant tissues has toxic effects on various growth and physiological processes,
such as inhibits growth and photosynthetic capability, leading to a decrease in biomass (Ghori et al. 2019; Sharma et al.
2020). The toxic effect of Ni on growth inhibition further enhances with an increment in high Ni levels (Hassan et al. 2019).
The present study showed a reverse relation between Ni accumulation and root FW of the plant. By augmenting the Ni
concentration to 400-µM, high Ni accumulation in the roots of two populations of A. inflatum, and alternatively, lead to a
reduction in roots FW. Besides, it was found that the roots of plants suffer the most damage compared to the shoots,
where the roots are exposed to the highest doses of Ni, which has reverse effects on root growth and biomass (Ameen et
al. 2019). For example, Ghasemi et al. (2009a) observed a remarkable reduction in the root biomass of A. inflatum
seedlings treated with 350-µM Ni. A study on Oryza sativa exposed to different Ni-treatment showed the highest Ni
accumulation, was associated with the highest reduction in the root FW in the 200-µM Ni-treatment (Rizwan et al. 2018).
The same result was reported in Taraxacum officinale exposed to Ni stress (Kováčik et al. 2019). Although Ni has been
considered as a micronutrient in plants, the amount of Ni in plants is about < 5 mg kg− 1 of dry weight (DW), when growth
on ordinary soils and at least 0.1 mg kg− 1 DW Ni need to prevent deficiency of Ni in plants (Welch 1995; Chaney et al.
2008). Nevertheless, when the Ni concentration in plants reaches above 50 to 100 mg kg− 1 (Hassan et al. 2019), plant
architecture was demolished, and limited plants to uptake minerals resulted in reduced plant growth (Fashola et al. 2016).
One of the reasons for the reduction in plant growth with high Ni concentrations is owing to the lack of other essential
elements such as Fe, Cu, and Mn due to their similar chemical properties and competition for uptake through the root
(Najafi Kakavand et al. 2019). For example, many transporters involved in Fe absorption and distribution, such as IRON-
REGULATED TRANSPORTER 1 (IRT1), can also uptake Ni by roots from the soil and promote Ni accumulation in plants.
Additionally, Fe-ligands like nicotianamine involved in Fe translocation from root to shoot can make strong complexes with
Ni (Leškova et al. 2020). However, Ni-hyperaccumulator plants can accumulate over 1–3% DW Ni concentration in their
tissues (Reeves et al. 2018). For the first time, Ghaderian et al. (2007) reported that A. inflatum plants that grow up on the
serpentine soils with 1350 µg Ni g− 1 soils could accumulate more than 3700 µg Ni g− 1 its shoot DW. In contrast, the
current study demonstrated that SA and JA's external use leads to increased Ni concentration in roots and enhanced roots
FW of M/NM populations under high Ni-treatment. Similar to our results, some investigations illustrated that JA
supplementation moderated Ni-toxicity by enhancing chlorophyll content, CO2 fixation, and photosynthetic yield, leads to
increased plant biomass in G. max (Sirhindi et al. 2016) and Cd-stress in G. max (Keramat et al. 2009). It was also found
that exogenous JA application improved Ni inhibitory effect on mitotic division and reduced the destruction of the root
structure leding to improved growth traits in G. max (Mir et al. 2018). Likewise, Sirhindi et al. (2015) reported that the
external use of JA to Ni toxic soybean plants cause restricted Ni uptake via roots and inhibited the Ni interference with
other necessary ionic metals needed for physiological processes improved the biomass of plants. Besides, SA
supplementation moderated Ni-toxicity and the amelioration of biomass in Triticum aestivum (Siddiqui et al. 2013) and B.
juncea (Zaid et al. 2019). They suggested that exogenous SA treatment led to increased growth and plant biomass by
reducing Ni uptake, improving photosynthetic pigments involved in photosynthesis reaction, increasing nitrogen
metabolism and mitotic activities. It is demonstrated that SA reduced abiotic stress, such as heavy metal, through-
mediated regulation of main plant-metabolic processes (Khan et al. 2015). Also, SA signaling pathways often cross-talk
with other hormone signaling pathways such as JA as a reaction to stress in plants. The interaction between JA and SA
signaling pathways can be synergistically or antagonistically, depending on specific stress (Sytar et al. 2019). The external
application of SA + JA increased root DW of Zea mays under drought stress. (Tayyab et al. 2020). Interestingly, exogenous
application SA and JA in Ni-exposure plants probably restrict Ni translocation root-to-shoot by preventing Ni absorption
and Ni storage in roots of plants and ameliorating Ni-toxicity effects resulted in biomass accretion (Zaid et al. 2019;
Mubarik et al. 2021).
Proline, as a vital osmolyte, has multiple roles in protecting and tolerating plants to abiotic stresses like heavy metals
(Petrovic and Krivokapic 2020). Proline is a compatible osmolyte that plays a considerable role in osmotic adjusting,
protein stability, membrane integrity, conservation of subcellular structures, and cellular redox-balancing (El-Beltagi et al.
Page 11/242020). Plants have expended variant protective strategies, including enzymatic/non-enzymatic antioxidant defense
systems, to alleviate or obliterate ROS's destructive effects in plant cells. It was suggested that proline has a ROS-
scavenging role and elevates antioxidative enzyme activity in plants (Hayat et al. 2012; Kaur and Asthir 2015; Altieri and
Nicholls 2020). Rendering to our results, the increase in proline content due to oxidative stress induced by 400-µM Ni in the
roots of M/NM A. inflatum plants is considerably reduced in plant's roots under SA and JA or SA/JA treatments. However,
the proline levels in the plant's roots showed a considerable enhancement compared to control plants. Similarly, research
has demonstrated that 100-µM Ni-induced proline accumulation in Vigna mungo L. (Gurpreet et al. 2012) and Sesuvium
portulacastrum L. (Fourati et al. 2020). Also, SA amended the proline contents in Catharanthus roseus L. exposed to
various Ni doses. However, SA efficaciously decreased Ni-influenced C. roseus plants' proline content that grows on
different Ni doses medium (Idrees et al. 2013). Likewise, SA and JA's application improved the Ni-tolerance mechanism
respectively, in G. max (Sirhindi et al. 2016) and Eleusine coracana L. (Kotapati et al. 2017) exposed to Ni-stress conditions
by increasing proline content. Furthermore, Tayyab et al. (2020) found that external application of combined SA + JA can
effectively mitigate drought stress in maize by ameliorating the proline content. The application of SA, JA, and SA + JA
triggers proline accumulation, which consecutively elevates the osmotic potential and balances cell redox status, as well
as, and improves the antioxidant system function, and finally restrict adverse effects of heavy metals stress in plants
(Nazar et al. 2015; Raza et al. 2020). These phiso-biochemical responses are probably in the response of M/NM A.
inflatum populations to Ni-toxicity, and the signaling cascade is with mediated JA and SA.
Nickel stress considerably alleviates proteins' levels in several plant species, owing to decreased protein synthesis and
hydrolyze (Hassan et al. 2019). Decreased protein levels due to heavy metals stress (such as Ni) through various
mechanisms, including; (i) high doses of Ni indirectly cause ROS generation, which eventually harms the proteins; (ii) Ni
can alter the conformation of proteins by binding functional groups of proteins such as sulfhydryl-groups and
consequently blocked of enzymes activity; (iii) as well as Ni-stress, leads to the accumulation of various amino acids like
histidine, in the cells of various plant tissues to Ni-detoxify, which resulted in reduced protein synthesis (Dutta et al. 2018;
Hassan et al. 2019). The current study showed that the total protein level was reduced in Ni-stressed M A. inflatum species
over the control plants' roots. However, the supplement of SA, JA, and/or SA + JA with Ni in two A. inflatum species showed
promotion of the total protein levels in comparison to Ni-treated plants alone. According to previous reports, total protein
levels of roots in rice (Rizwan et al. 2018) and wheat (Gajewska et al. 2009) exposed to high doses of Ni showed a 50%
reduction than plants without treatments. Alternatively, Sirhindi et al. (2016) found that JA's addition to G. Max under Ni-
stress resulted in an approximately 60% increment in protein level than Ni-treated plants. Likewise, SA triggered a
remarkable improvement of the protein levels in the roots of Cu-stressed Helianthus annuus L. (El-Tayeb et al. 2006). It was
also explained that the use of exogenous SA + JA in maize reduced the damaging effects of drought stress resulted in an
increase in protein levels in comparison to maize plants grown under drought stress without PGR-treatments (Tayyab et al.
2020). A positive effect of exogenously SA, JA, and/or SA + JA treatments on protein contents in roots of M/NM A.
inflatum populations is probably due to the inhibition of the destructive effect of ROS induced-Ni on the protein structures
and their activity.
Heavy metal toxicity induces ROS generation in the root cells by disrupting the electron-transfer-chain in mitochondria and
apoplastic space (Farvardin et al. 2020; Hasanuzzaman et al. 2020). Therefore, the over-generation of ROS and
consequent oxidative stress in plants can cause great damage to plant cells (Petrov et al. 2015). The high content of Ni
indirectly stimulates oxidative stress and enzymatic activity inhibition (Ghori et al. 2019), so that a toxic effect of Ni is
related to the ROS regeneration and, as a result, an imbalanced redox state (Georgiadou et al. 2018). Besides, Ni-induced
ROS generation induces lipid-peroxidation, protein-oxidation, pigment damage, and harm to DNA (Ameen et al. 2019). On
the other hand, accumulated ROS due to Ni-stress can act as a signaling molecule that stimulates phytohormones such as
SA and JA, which in turn stimulate the plant's defense responses to Ni-stress, including antioxidant enzymatic system (like
SOD, CAT, POD, and APX) and non-enzymatic system (like glutathione and proline) (Sewelam et al. 2016). Besides,
antioxidant enzymes are powerful ROS scavengers, so enhancing their activities with increasing Ni concentration is a good
Page 12/24indication of stimulating the antioxidant defense pathway (Giannakoula et al. 2021). The current study showed that with
increasing Ni levels, at the same time, H2O2 content enhanced, and also SOD, POD, CAT, and APX activities improved in
both M/NM populations; however, APX activity did not change in the M population. When plants are subjected to heavy
metals, they trigger an antioxidative defense pathway to reduce oxidative stress's negative effects. Interestingly, depending
on the genotype and plant species, the kind of stress, and the stress period, the antioxidant defense response varies in
different plants (Hasanuzzaman et al. 2020). For example, Kandelia candel and Bruguiera gymnorrhiza plants were
exposed to lead (Pb), Cd, and mercury (Hg) stress, showed an increasing trend in SOD, CAT, and POD activities in roots
(Zhang et al. 2007). Also, O. sativa exposed to different doses of Ni; increasing the concentration of Ni enhanced the H2O2
level in the roots, which subsequently reduces the oxidative stress, improved the enzymatic activity of CAT, POD, and APX,
while SOD activity was found to be decreased (Rizwan et al. 2018). Moreover, under Ni-stressed wheat plants, the activity
of SOD decreased with increasing H2O2 content in roots while CAT and APX and POD activity did not significantly change
than control plants (Gajewska and Skłodowska 2008). On the other hand, SA and JA's external use on plants exposed to
abiotic stress conditions causes different physio-biochemical responses (Tayyab et al. 2020). According to our results, the
external application of SA and JA and/or combined (SA + JA) in roots of both populations of A. inflatum treated with 400-
µM Ni caused ameliorated oxidative stress by enhancing SOD, CAT, POD activities, which resulted in reduced H2O2 levels
than roots of plants without treatments. When plants are exposed to abiotic stress, SA and JA are induced as key
signaling-molecules in the biochemical response pathways for defense mechanisms in plants. Besides, SA and JA's
signaling to fine-tune abiotic stress depends on the character, intensity, and duration of stress exposure (Sewelam et al.
2016). For instance, Yusuf et al. (2012) reported that the ameliorative role of SA on Ni-induced oxidative stress in B. juncea
and also illustrated that SA supplementation reduced the reverse effects of Ni revealed in the form of the less dose of the
abridged generation of H2O2. Furthermore, in barley plants exposed to Cd-toxicity, the CAT and APX activities were
enhanced, while these enzymes' activity mitigated in the presence of SA. It was also found that the endogenous SA level
was incremented to mitigate the damage caused by oxidative stress induced by Cd-toxicity (Metwally 2003). Contrarily, SA
elevated the SOD, POD, and CAT activities in C. roseus exposed to Ni-stressed (Idrees et al. 2013), M. arvensis L. exposed to
Cd-treated (Zaid et al. 2020), and watermelon plants under B-toxicity (Moustafa-Farag et al. 2020). Recent molecular
researches have proved that SA can regulate genes involved in encoding antioxidative defense mechanisms, and therewith
able to ameliorate abiotic stress such as heavy metal (Khan et al. 2015). Moreover, JA is a signaling molecule tightly
related to plant defense against biotic and abiotic stresses (Ruan et al. 2019). For instance, in H. annuus plants, 50 ml of
methyl jasmonate (MeJA) treatment enhanced the ROS content in the root apoplastic space, followed by an increment in
the activities of the ROS-scavenging enzymes (Parra-Lobato et al. 2009). Furthermore, Sirhindi et al. (2016) displayed that
JA increased the SOD, POD, CAT, and APX activities in G. max plants exposed to Ni toxicity. Mir et al. (2018) also reported
that JA's supplement has an ameliorating effect on Ni toxicity in G. max plants exposed to NiCl2 (4-mM) by improving SOD
and CAT activities. So far, there have been several reports on the protective role of JA against metal stress in plants,
including G. max under Ni toxicity (Sirhindi et al. 2015), Lycopersicon esculentum (Bali et al. 2019b), and Arabidopsis
thaliana (Lei et al. 2020) under Cd stress, Puccinellia tenuiflora under B-toxicity toxicity (Zhao et al. 2019), tomato under Pb
toxicity (Bali et al. 2019a), and L. valdiviana exposed to As stress (Coelho et al. 2020). It is found that JA as a signaling
molecule can increase genes expression involved in encoding antioxidative enzymes in plant cells for decreasing heavy
metals toxicity by reducing ROS contents. Therefore, JA can protect the macromolecules structure such as proteins as well
as cell membranes by reducing the content of H2O2 and malondialdehyde in plant cells. (Emamverdian et al. 2020). In the
current study, it was determined that the external use of SA + JA resulted in a significantly reduced H2O2 content by
enhancing antioxidative enzyme activity in the roots of both M/NM populations. However, the high activity of the APX
enzyme in the M-population's root was observed only along whit the lower doses of SA and JA. The same results were
obtained by Tayyab et al. (2020) on Z. mays plants under drought stress. Notably, SA and JA play a substantial role in
triggering the expression of genes associated with defense mechanisms by causing oxidation to alter the signaling
pathway constituents (Fobert and Després 2005; Kalaivani et al. 2016; Raza et al. 2020, 2021). These investigations
illustrated that external SA and JA application could efficiently mitigate heavy metals toxicity (e.g., Ni) in plants.
Page 13/24Additionally, PCA plot results show that two populations of A. inflatum display similar trends in physiological and
biochemical reactions in response to SA, JA, and SA + JA treatment under Ni-toxicity conditions. These results propose that
SA and JA, especially SA + JA, under Ni-toxicity is more efficient in enhancing antioxidant enzymatic capacity and proline
content to decrease oxidative stress induced by Ni-toxicity. Although there are many investigations on SA and/or JA's
positive role to restrict toxic effect-induced by heavy metals such as Ni in plants; however, no study has been reported on
the beneficial effects of SA + JA combination on heavy metal toxicity. Hence, this study serves as the main purpose to
evaluate the influential role of SA + JA combination in enhancing Ni-tolerance in M/NM A. inflatum populations.
Conclusion
Although Ni is considered a micronutrient for plants, high concentrations of Ni can lead to metabolic disorders, ionic
imbalance, and oxidative stress in the plant and ultimately inhibit plant growth and development. Nevertheless,
supplementation of SA and JA in the present study, shown ameliorating effects and play an essential role in overcoming
Ni-toxicity. Our findings demonstrated that exogenous use of JA and SA, especially combined of SA + JA, in both M/NM A.
inflatum populations had substantial effects on improving the root biomass, antioxidant defense responses, and proline
contents (as an osmolyte), which in turn mitigate the oxidative stress due to Ni-toxicity (Fig. 7). In conclusion, the SA, JA,
and/or SA + JA supplementation can partially exclude the damaging effects of Ni-induced oxidative stress in two M/NM
populations of A. inflatum by adjusting defense signaling pathways and antioxidant mechanisms. However, the
mechanisms of interaction between heavy metals and hormones affecting abiotic stresses such as SA and JA is not yet
crystal clear and needs further investigation.
Abbreviations
APX, Ascorbate peroxidase; CAT, Catalase; FW, Fresh weight; JA, Jasmonic acid; M, Metallicolous; Ni, Nickel; NM, Non-
metallicolous; POD, Peroxidase; PGR, Plant growth regulator; ROS, Reactive oxygen species; SA, Salicylic acid; SOD,
Superoxide dismutase
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Data availability
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable
request.
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
SNK performed the experiment, writing, editing, statistical analysis, figure construction, and finalize the manuscript; NK
supervised the research; HRG review and editing; AR reviewed, edited, writing, figure construction, and finalize the
Page 14/24manuscript; MC performed the statistical analysis, figure construction, reviewed, and writing;, MM supervised the research,
review, and writing.
Acknowledgment
We would like to thank the research deputy of Kermanshah University of Medical Sciences (project number 990252; ethical
code IR.KUMS.REC.1399.321) and Razi University for providing the necessary facilities for carrying out this research.
Funding
Not applicable.
References
1. Abd_Allah EF, Hashem A, Alam P, Ahmad P (2019) Silicon Alleviates Nickel-Induced Oxidative Stress by Regulating
Antioxidant Defense and Glyoxalase Systems in Mustard Plants. J Plant Growth Regul 38:1260–1273.
doi:10.1007/s00344-019-09931-y
2. Aebi H (1983) Catalase. In: Bergmeyer HU (ed) Methods in Enzymatic Analysis. Academic Press, New York, pp 273–
286
3. Ali H, Khan E, Ilahi I (2019) Environmental Chemistry and Ecotoxicology of Hazardous Heavy Metals: Environmental
Persistence, Toxicity, and Bioaccumulation. J Chem 2019:1–14. doi:10.1155/2019/6730305
4. Altieri M, Nicholls CI (2020) Sustainable Agriculture Reviews 41. Springer International Publishing, Cham
5. Ameen N, Amjad M, Murtaza B et al (2019) Biogeochemical behavior of nickel under different abiotic stresses: toxicity
and detoxification mechanisms in plants. Environ Sci Pollut Res 26:10496–10514. doi:10.1007/s11356-019-04540-4
6. Amjad M, Raza H, Murtaza B et al (2019) Nickel Toxicity Induced Changes in Nutrient Dynamics and Antioxidant
Profiling in Two Maize (Zea mays L.). Hybrids Plants 9:5. doi:10.3390/plants9010005
7. Avalbaev A, Yuldashev R, Fedorova K et al (2016) Exogenous methyl jasmonate regulates cytokinin content by
modulating cytokinin oxidase activity in wheat seedlings under salinity. J Plant Physiol 191:101–110.
doi:10.1016/j.jplph.2015.11.013
8. Bali S, Jamwal VL, Kaur P et al (2019a) Role of P-type ATPase metal transporters and plant immunity induced by
jasmonic acid against Lead (Pb) toxicity in tomato. Ecotoxicol Environ Saf 174:283–294.
doi:10.1016/j.ecoenv.2019.02.084
9. Bali S, Jamwal VL, Kohli SK et al (2019b) Jasmonic acid application triggers detoxification of lead (Pb) toxicity in
tomato through the modifications of secondary metabolites and gene expression. Chemosphere 235:734–748.
doi:10.1016/j.chemosphere.2019.06.188
10. Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–
207. doi:10.1007/BF00018060
11. Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the
principle of protein-dye binding. Anal Biochem 72:248–254. doi:10.1016/0003-2697(76)90527-3
12. Caarls L, Pieterse CMJ, Van Wees SCM (2015) How salicylic acid takes transcriptional control over jasmonic acid
signaling. Front Plant Sci 6:1–11. doi:10.3389/fpls.2015.00170
13. Chaki M, Begara-Morales JC, Barroso JB (2020) Oxidative Stress in Plants. Antioxidants 9:481.
doi:10.3390/antiox9060481
14. Chance B, Maehly AC (1955) Assay of catalases and peroxidases. In: Methods in Enzymology. pp 764–775
15. Chaney RL, Chen K-Y, Li Y-M et al (2008) Effects of calcium on nickel tolerance and accumulation in Alyssum species
and cabbage grown in nutrient solution. Plant Soil 311:131–140. doi:10.1007/s11104-008-9664-7
Page 15/2416. Coelho DG, de Andrade HM, Marinato CS et al (2020) Exogenous jasmonic acid enhances oxidative protection of
Lemna valdiviana subjected to arsenic. Acta Physiol Plant 42:97. doi:10.1007/s11738-020-03086-0
17. Dutta S, Mitra M, Agarwal P et al (2018) Oxidative and genotoxic damages in plants in response to heavy metal stress
and maintenance of genome stability. Plant Signal Behav 13:1–49. doi:10.1080/15592324.2018.1460048
18. El-Beltagi HS, Mohamed HI, Sofy MR (2020) Role of Ascorbic acid, Glutathione and Proline Applied as Singly or in
Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under
Different Levels of Irrigation Intervals. Molecules 25:1702. doi:10.3390/molecules25071702
19. El-Tayeb MA, El-Enany AE, Ahmed NL (2006) Salicylic acid-induced adaptive response to copper stress in sunflower
(Helianthus annuus L.). Plant Growth Regul 50:191–199. doi:10.1007/s10725-006-9118-2
20. Emamverdian A, Ding Y, Xie Y (2020) The Role of New Members of Phytohormones in Plant Amelioration under Abiotic
Stress with an Emphasis on Heavy Metals. Polish J Environ Stud 29:1009–1020. doi:10.15244/pjoes/108687
21. Farvardin A, González-Hernández AI, Llorens E et al (2020) The Apoplast: A Key Player in Plant Survival. Antioxidants
9:604. doi:10.3390/antiox9070604
22. Fashola M, Ngole-Jeme V, Babalola O (2016) Heavy Metal Pollution from Gold Mines: Environmental Effects and
Bacterial Strategies for Resistance. Int J Environ Res Public Health 13:1047. doi:10.3390/ijerph13111047
23. Fobert PR, Després C (2005) Redox control of systemic acquired resistance. Curr Opin Plant Biol 8:378–382.
doi:10.1016/j.pbi.2005.05.003
24. Fourati E, Vogel-Mikuš K, Wali M et al (2020) Nickel tolerance and toxicity mechanisms in the halophyte Sesuvium
portulacastrum L. as revealed by Ni localization and ligand environment studies. Environ Sci Pollut Res 27:23402–
23410. doi:10.1007/s11356-019-05209-8
25. Gajewska E, Skłodowska M (2008) Differential biochemical responses of wheat shoots and roots to nickel stress:
antioxidative reactions and proline accumulation. Plant Growth Regul 54:179–188. doi:10.1007/s10725-007-9240-9
26. Gajewska E, Wielanek M, Bergier K, Skłodowska M (2009) Nickel-induced depression of nitrogen assimilation in wheat
roots. Acta Physiol Plant 31:1291–1300. doi:10.1007/s11738-009-0370-8
27. Georgiadou EC, Kowalska E, Patla K et al (2018) Influence of Heavy Metals (Ni, Cu, and Zn) on Nitro-Oxidative Stress
Responses, Proteome Regulation and Allergen Production in Basil (Ocimum basilicum L.) Plants. Front Plant Sci 9:1–
16. doi:10.3389/fpls.2018.00862
28. Ghaderian SM, Mohtadi A, Rahiminejad R et al (2007) Hyperaccumulation of nickel by two Alyssum species from the
serpentine soils of Iran. Plant Soil 293:91–97. doi:10.1007/s11104-007-9221-9
29. Ghasemi R, Ghaderian SM, Krämer U (2009a) Interference of nickel with copper and iron homeostasis contributes to
metal toxicity symptoms in the nickel hyperaccumulator plant Alyssum inflatum. New Phytol 184:566–580.
doi:10.1111/j.1469-8137.2009.02993.x
30. Ghasemi R, Ghaderian SM, Krämer U (2009b) Accumulation of Nickel in Trichomes of a Nickel Hyperaccumulator
Plant, Alyssum inflatum. Northeast Nat 16:81–92. doi:10.1656/045.016.0507
31. Ghori N-H, Ghori T, Hayat MQ et al (2019) Heavy metal stress and responses in plants. Int J Environ Sci Technol
16:1807–1828. doi:10.1007/s13762-019-02215-8
32. Giannakoula A, Therios I, Chatzissavvidis C (2021) Effect of lead and copper on photosynthetic apparatus in citrus
(Citrus aurantium l.) plants. the role of antioxidants in oxidative damage as a response to heavy metal stress. Plants
10:1–14. doi:10.3390/plants10010155
33. Giannopolitis CN, Ries SK (1977) Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol 59:309–314.
doi:10.1104/pp.59.2.309
34. Gu C-S, Yang Y-H, Shao Y-F et al (2018) The effects of exogenous salicylic acid on alleviating cadmium toxicity in
Nymphaea tetragona Georgi. South African J Bot 114:267–271. doi:10.1016/j.sajb.2017.11.012
Page 16/24You can also read

















































