The MFHR1 Fusion Protein Is a Novel Synthetic Multitarget Complement Inhibitor with Therapeutic Potential - Plant Biotech Lab
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
BASIC RESEARCH www.jasn.org
The MFHR1 Fusion Protein Is a Novel Synthetic
Multitarget Complement Inhibitor with Therapeutic
Potential
Stefan Michelfelder,1 Friedericke Fischer,1 Astrid Wäldin,1 Kim V. Hörle,1 Martin Pohl,1
Juliana Parsons,2 Ralf Reski ,2,3 Eva L. Decker,2 Peter F. Zipfel,4 Christine Skerka,4 and
Karsten Häffner1
1
Department of General Pediatrics, Adolescent Medicine and Neonatology, Medical Center, Faculty of Medicine,
2
Plant Biotechnology, Faculty of Biology, and 3Centre for Biological Signaling Studies, University of Freiburg,
Freiburg, Germany; and 4Leibniz Institute for Natural Product Research and Infection Biology, Friedrich Schiller
University, Jena, Germany
ABSTRACT
The complement system is essential for host defense, but uncontrolled complement system activation
leads to severe, mostly renal pathologies, such as atypical hemolytic uremic syndrome or C3 glomerulop-
athy. Here, we investigated a novel combinational approach to modulate complement activation by tar-
geting C3 and the terminal pathway simultaneously. The synthetic fusion protein MFHR1 links the
regulatory domains of complement factor H (FH) with the C5 convertase/C5b-9 inhibitory fragment of
the FH-related protein 1. In vitro, MFHR1 showed cofactor and decay acceleration activity and inhibited C5
convertase activation and C5b-9 assembly, which prevented C3b deposition and reduced C3a/C5a and
C5b-9 generation. Furthermore, this fusion protein showed the ability to escape deregulation by FH-
related proteins and form multimeric complexes with increased inhibitory activity. In addition to substan-
tially inhibiting alternative and classic pathway activation, MFHR1 blocked hemolysis mediated by serum
from a patient with aHUS expressing truncated FH. In FH2/2 mice, MFHR1 administration augmented
serum C3 levels, reduced abnormal glomerular C3 deposition, and ameliorated C3 glomerulopathy. Tak-
ing the unique design of MFHR1 into account, we suggest that the combination of proximal and terminal
cascade inhibition together with the ability to form multimeric complexes explain the strong inhibitory
capacity of MFHR1, which offers a novel basis for complement therapeutics.
J Am Soc Nephrol 29: ccc–ccc, 2018. doi: https://doi.org/10.1681/ASN.2017070738
Complement activity is strictly controlled by a panel
of soluble and membrane-bound regulators that Significance Statement
intervene at several levels of the cascade. With a
Atypical hemolytic uremic syndrome (aHUS) and C3
serum concentration of 500 mg/ml, plasma factor glomerulopathy (C3G) are caused by overactivation of
the alternative pathway of complement and have poor
prognosis, often leading to ESRD. Therapeutic options
Received July 7, 2017. Accepted December 13, 2017. for these diseases are limited. This manuscript
describes a novel synthetic fusion protein, MFHR1,
Published online ahead of print. Publication date available at combining proximal and terminal cascade inhibition
www.jasn.org. activities and the ability to form multimeric complexes.
Correspondence: Dr. Karsten Häffner, Department of Pediatrics MFHR1 shows strong inhibitory capacity in vitro and
and Adolescent Medicine, University of Freiburg Medical Center, ameliorates experimental C3G in a factor H knockout
Mathildenstrasse 1, 79106 Freiburg, Germany. Email: karsten. mouse model in vivo. MFHR1 might, therefore, offer a
haeffner@uniklinik-freiburg.de novel basis for therapeutics in complement-associated
diseases.
Copyright © 2018 by the American Society of Nephrology
J Am Soc Nephrol 29: ccc–ccc, 2018 ISSN : 1046-6673/2904-ccc 1BASIC RESEARCH www.jasn.org H (FH) is the major regulator of the alternative pathway (AP). some patients, which might be explained by preserved C3 FH restricts “tick-over” C3 activation and the C3 amplifica- convertase activity.19 In addition, incomplete responses or tion loop in the fluid phase as well as on host surfaces by acting even therapeutic failures have been described to occur in other as cofactor to promote factor I (FI)–mediated proteolysis of complement-driven pathologies (e.g., paroxysmal nocturnal C3b into iC3b1 and preventing the formation or accelerating hemoglobinuria).20,21 the decay of C3 convertases.2 FHR-1 as well as FHR-2, -3, -4, Newer therapeutic strategies are focusing on the interven- and -5 (summarized as FHR family) are closely related to FH, tion of C3 activation by using small peptides (e.g., Compstatin and complement regulatory as well as host/opsonin recogni- and Amyndas), antibodies (e.g., targeting factor D, factor B, or tion features are attributed to most of them.3 FHR-1 regulates C3), or naturally occurring physiologic regulators or their terminal pathway activation by preventing C5 cleavage functional domains for designed therapeutic inhibitors.22–24 through the C5 convertase and thereby, inhibiting C5a release, Especially in FH deficiencies, replacement of FH has been pro- membrane attack complex (MAC) formation, and subsequent posed as a viable treatment option to replace dysfunctional cytolysis.4 In addition and independent of C5 convertase reg- protein and restore AP regulation,22,25–27 and the latest tech- ulation, FHR-1 inhibits assembly of C5b6(7) complexes and nologic advancements now allow the recombinant production MAC formation.4 FHR-1, FHR-2, and FHR-5 contain a con- of glycosylation-optimized FH with full bioactivity.28 Also, served dimerization interface that enables the formation of minimized factor H (mini-FH) variants,29–31 soluble CR1,32 homo- and heterodimers (e.g., FHR-1:FHR-1; FHR-2:FHR-1, or combinational approaches, like TT30, 33,34 inhibit C3 respectively), and these complexes can compete with FH activation. for C3b or host cell surface binding and thereby, reduce FH Regarding the complexity of the complement system, which surface levels, leading to local complement activation.5,6 The if not properly regulated, leads to accumulation of various plasma concentration of FHR-1 is 70–100 mg/ml.4 However, potential pathogenic intermediates, a combinational or “mul- given the regulatory as well as deregulatory properties titarget” strategy to modulate complement activation could of FHRs, it has been postulated that FHRs have evolved potentially improve therapeutic efficiency while diminishing to act in concert with FH, but their exact role in comple- unwanted side effects, and it might also be more efficient in ment regulation as well as disease has not yet been precisely controlling complex disease systems and less prone to drug elucidated. 3 resistance.35,36 Mutations in activators or inhibitors of the complement We describe here a novel fusion protein MFHR1, which cascade (e.g., FH, FI, factor B, C3, CD46) or autoantibodies combines the C3 regulatory and cell surface/opsonin targeting (e.g., anti-FH) cause uncontrolled complement activation domains of FH with the terminal pathway regulatory domains and MAC formation on host surfaces (e.g., on endothelial derived from FHR-1 displaying strong complement inhibitory cells), leading to the development of atypical hemolytic ure- activity in vitro. MFHR1 modulates complement by acting on mic syndrome (aHUS).7 In C3 glomerulopathies (C3Gs), C3 activation and amplification as well as the C5/C5b-9 level. fluid-phase activation is deregulated by mutations in comple- Our data show that targeting complement activation on mul- ment regulators, FH/CFHR genomic rearrangements,8 or tiple levels simultaneously offers a basis for an optimized com- autoantibodies (e.g., C3Nef), resulting in glomerular accu- plement inhibition, which was also shown by the ability of mulation or deposition of C3 cleavage fragments, which MFHR1 to prevent AP deregulation in clinically relevant mod- seem to be the major pathogenic factors.9 In addition to els of aHUS and C3G. MAC-induced damage, opsonin depositions (e.g., C3b) and anaphylatoxins (C3a and C5a) promote disease progression by triggering inflammation, proliferation, or infiltration of RESULTS immune cells or podocyte damage. The same factors are also thought to play a critical role in the pathomechanism Structure, Expression, and Characterization of MFHR1 of IgA nephropathy, membranous nephropathy, MFHR1 was designed to combine the terminal pathway reg- diabetic kidney disease, ANCA vasculitis, graft rejection, ulatory and dimerization properties of FHR-1 with the C3 and ischemia-reperfusion injury.9–13 regulatory and surface binding properties of FH. We di- Targeting the complement system pharmacologically is rectly linked the two N-terminal C5/C5b6 binding domains highly desired, and terminal pathway inhibition (i.e., C5 by FHR-11–2 to the regulatory (FH1–4) and surface recognition domains the humanized antibody eculizumab) has already been suc- (FH19–20) of FH (Figure 1A). MFHR1 was expressed in insect cessfully transferred into clinical practice and showed remark- cells and purified from the supernatant, yielding approxi- able improved patient survival and quality of life.14–16 mately 5 mg protein per liter culture. The purity of MFHR1 However, despite the clinical success of eculizumab, incom- was confirmed by Coomassie and silver staining (Figure 1B). plete responses by solely blocking C5 (serum concentration MFHR1 migrated at its predicted molecular mass of 59 kD. The 55–113 mg/ml) have been reported for aHUS (e.g., when re- presence of FHR-11–2, FH1–4, and FH20 was confirmed by sidual C5 activity derogates the drug’s therapeutic bene- immune detection using specific antibodies and antisera fit).17,18 In C3G, eculizumab led to a partial response only in (Figure 1C). 2 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018
www.jasn.org BASIC RESEARCH
Figure 1. Evidence for correct structure and expression of MFHR1. (A) Structural assembly of MFHR1. MFHR1 is a fusion protein
composed of FHR-11–2 N-terminally linked to FH domains FH1–4 and FH19–20. A decahistidine tag was used for means of purification.
FHR-11–2 contains a dimerization motif and inhibits C5 cleavage and MAC assembly. FH1–4 has decay acceleration, cofactor activity, and
FH19–20 binding sites for C3b/iC3b/C3d and cell surfaces. (B) SDS-PAGE and Coomassie loaded with 5 mg (left panel) or silver staining
loaded with 0.4 mg of MFHR1 purified from supernatants of baculovirus-infected Spodoptera frugiperda cells via Ni affinity and size
exclusion chromatography (right panel). MFHR1 migrates with the calculated molecular mass of 59 kD under reducing conditions
(Coomassie stain; left panel, lane I). Faster mobility of MFHR1 under nonreducing conditions (Coomassie stain; right panel, lane II)
indicates the presence of disulfide bounds. (C) Immunodetection using a-hFH or a-FH1–4 antisera or mAb C18 that binds to SCR20 or
a–FHR-1/2 antibodies indicates the correct integrity of FHR-11–2, FH1–4, and FH19–20 domains in recombinant MFHR1. Mini-FH, full-
length FHR-1, FH1–4, and hFH served as controls. MW, molecular mass marker.
MFHR1 Inhibits AP and CP Activation densitometry (Figure 3A), MFHR1, hFH, and FH 1– 4
Global complement inhibitory potency of MFHR1 was ana- mediated a dose-dependent cleavage of a9-chain fragments.
lyzed by complement ELISA measuring C5b-9 formation after hFH with an IC50 of 4 nM showed an 8.75-fold higher cofactor
LPS-induced AP activation in human serum (HS) with the activity compared with MFHR1 (IC50=35 nM) that was in a
addition of MFHR1 or controls. MFHR1 inhibited C5b-9 for- similar range to FH 1–4 (IC50=26 nM), whereas FHR-11–2
mation with an IC50 of 3.2 nM and exceeded the inhibitory showed no activity (Figure 3A, Table 2). In addition,
activities of eculizumab (5.5-fold; IC50=17.5 nM), human fac- MFHR1 and hFH showed cofactor activity when bound to
tor H (hFH; 52-fold; IC50=167 nM), and mini-FH (3.9-fold; human umbilical vein endothelial cells (Supplemental Figure
IC50=12.4 nM), whereas FHR-1 showed only AP inhibition at 2B). We then compared the decay acceleration activity of
very high concentrations (3.16 mM) (Figure 2A, Table 1). In MFHR1 and hFH by their ability to displace factor B from
addition, MFHR1 inhibited classical pathway (CP) activation preformed C3 convertases (C3bBb), where MFHR1 showed
after IgM-induced CP activation in HS and C5b-9 formation nearly identical activity to hFH (Figure 3B). After AP activa-
at an IC50=15.9 nM (Figure 2B, Table 1). tion, C3b depositions initiate and further amplify the AP and
the generation of proinflammatory anaphylatoxins (e.g., C3a).
MFHR1 Inhibits Complement Activation on the C3 Axis Therefore, we analyzed the ability of MFHR1 to block surface
To investigate whether MFHR1 maintained the functional deposition of the opsonin C3b after LPS activation in HS in the
properties of FH, we first measured the ability of MFHR1 to AP ELISA. MFHR1 strongly inhibited C3b depositions with an
bind to C3b, the key opsonin of the AP, using an ELISA-based IC50 of 12.4 nM that exceeded the inhibitory activity of hFH
binding assay. MFHR1 specifically and in a dose-dependent (IC50=513 nM) and mini-FH (IC50=40.4 nM) (Figure 3C,
manner bound C3b at levels comparable with those of hFH Table 1). The IC50 values of hFH and mini-FH were compa-
(Supplemental Figure 1A). In addition, MFHR1 bound to hu- rable with observations reported in a previous study.29 As ex-
man umbilical vein endothelial cells, which were used as a pected, eculizumab and FHR-1 had no inhibitory activity,
model of host cells37 (Supplemental Figure 1B). As analyzed because these proteins are known to regulate complement ac-
in a fluid-phase cofactor assay after SDS-PAGE (Supplemental tivity only in later stages of the cascade (Figure 3C). Because
Figure 2) and quantitative analysis of a9-chain cleavage by anaphylatoxins have proinflammatory effects and play an
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 3BASIC RESEARCH www.jasn.org
MFHR1 Binds C5 and Regulates AP Activation at the
Level of C5 and C5b-9 Formation
To test whether MFHR1 has the ability to interact with C5,
microtiter plates were coated with MFHR1, FHR-1, or hFH,
respectively (Supplemental Figure 3). After incubation of the
wells with purified C5, anti-C5 antibody strongly and dose-
dependently detected C5 bound to FHR-1 and MFHR1, but
only a weak C5 signal was detected in hFH- or BSA-coated
wells (Figure 4A). In a next step, inhibitory effects of MFHR1
on the C5 convertase were investigated. MFHR1 inhibited
C5a formation in HS after activation of the AP slightly stron-
ger than mini-FH and eculizumab and considerably stronger
than hFH (Figure 4B). Again and consistent with our results
from the C3a ELISA, FHR-1 increased C5a generation, prob-
ably as a consequence of FHR-1–mediated deregulation in
this serum–based assay. To investigate the influence of
MFHR1 on C5 cleavage in a serum-free approach, an exper-
imental cobra venom factor (CVF) convertase (CVFBb) was
generated and added to C5 preincubated with or without test
proteins, sheep erythrocytes (sEs), and C6-C9, and the lytic
activity of the samples was measured. Eculizumab38 as well as
FHR-14 are known to prevent convertase-mediated C5 cleav-
age by binding to C5. MFHR1 significantly inhibited CVF-
induced lysis at comparable levels to eculizumab and FHR-1,
whereas hFH and BSA had no significant effect (Figure 4C).
We then assayed whether MFHR1, like FHR-1, has the ability
to inhibit the C5 convertase–independent step of C5b-9 for-
mation. We incubated C5b6 with test proteins before addition
to sE and C7–C9. C5b-9 formation was measured by the rate
Figure 2. MFHR1 inhibits AP and CP activation. (A) IC50 fitting of lysis (Figure 4D). MFHR1 and FHR-1 significantly reduced
curves for inhibition of AP activation. MFHR1, hFH, mini-FH, C5b-9 formation, although FHR-1 tended to be slightly less
FHR-1, or eculizumab was added to HS, and C5b-9 complex effective, whereas neither eculizumab nor hFH and BSA in-
formation was measured by ELISA after AP activation by LPS. hibited C5b-9 at significant levels compared with controls
Heat-inactivated (hi) HS or HS added to wells not coated with LPS without regulators added (Figure 4D). These data show that
(2LPS) was used as control. Data points represent mean values
MFHR1 is a complement inhibitor combining C3 and termi-
from n=4 assays 6SEM using sera of four individual healthy do-
nal pathway regulatory properties acting on the C3/C5 and
nors. Serum without inhibitors was set to 100% AP activity for
each individual experiment. The AP inhibitory activity of MFHR1
C5b-9 axes simultaneously.
was significantly different from mini-FH, hFH, and eculizumab
(two-way ANOVA; P,0.001). (B) Inhibition of CP after adding
MFHR1 to HS and measuring activity in processed serum using Multimer Formation Potentiates AP Inhibitory Activity
WIESLAB Complement System CP ELISA. Bars represent mean of MFHR1
values from n=3 assays 6SEM using sera of three individual In the preparative size exclusion chromatography, the elution
healthy donors. HS or hi HS was used as control. Positive kit profile of MFHR1 from the purified protein stocks migrated
control was set to 100% for each individual experiment. Asterisks as a broad peak, whereas hFH purified from human plasma was
indicate statistical significance versus untreated serum (one-way found to migrate in a single and narrow peak fraction at ap-
ANOVA with Bonferroni test). **P,0.01; ***P,0.001.
proximately 300 kD (Figure 5A, middle panel). Collected frac-
tions of MFHR1 were analyzed via SDS-PAGE and silver
important role in disease progression, we investigated the in- stained (Figure 5B). The six fractions contained decreasing
hibitory potency of MFHR1 on C3a generation in activated MFHR1 concentrations as indicated by the OD at 280 nm in
sera. Here again, MFHR1 and mini-FH exhibited the highest the chromatogram. All fractions were pure for MFHR1, with
efficiency for preventing C3a generation followed by hFH only minor low molecular mass bands that might contain
(Figure 3D). Eculizumab did not result in any reduction of MFHR1 degradation products. As analyzed by AP ELISA, ei-
C3a levels, and FHR-1 at 0.316 and 1 mM increased C3a levels, ther 10 nM MFHR1 collected in fraction I or purified MFHR1
presumably as a consequence of FHR-1–mediated deregula- completely inhibited AP activity compared with heat-inactivated
tion in this serum-based assay. HS, whereas the inhibitory activity was severely reduced
4 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018www.jasn.org BASIC RESEARCH
Table 1. Calculated IC50 values for complement inhibitory capacity The addition of MFHR1 protected sE from
Complement Assay and Inhibitor IC50 Values, nM 2
R 95% Confidence Interval complement-mediated lysis more effec-
AP ELISA (C5b-9) tively than eculizumab (Figure 7A). We
MFHR1 3.2 0.87 2.73 to 3.8 also investigated the efficacy of MFHR1 in
Eculizumab 17.5 0.86 12.3 to 25 an animal model of C3G, namely FH
Mini-FH 12.4 0.83 8.4 to 18 knockout mice. FH2/2 mice display ab-
hFH 167 0.88 126 to 222 normal glomerular C3 accumulation and
FHR-1 2748 0.14 666 to 11,300 low serum C3/C5 levels.40,41 Administra-
CP ELISA (C5b-9) tion of a single dose of MFHR1 increased
MFHR1 15.9 0.82 6.4 to 40 serum C3 levels at all analyzed time points,
AP ELISA (C3b deposition)
reaching a peak of approximately 26% of
MFHR1 12.4 0.84 8.3 to 18.6
wild-type levels after 12 hours, whereas
Eculizumab No activity — —
Mini-FH 40.4 0.70 25 to 64
hFH increased serum C3 to comparable
hFH 513 0.82 356 to 739 levels to MFHR1 after 12 hours but led
FHR-1 No activity — — to a further increase, reaching approxi-
mately 53% of wild-type levels after 24
hours (Figure 7B). Serum C5 was detect-
able 24 hours after injection of MFHR1 or
hFH, whereas it was not present in PBS-in-
in fractions II–VI (Figure 5C). These data suggest that MFHR1 jected mice (Figure 7C). In addition, glomerular C3 staining
migrates predominantly in a multimeric state in the fluid was significantly reduced at a comparable degree in mice in-
phase. Preparations containing multimeric MFHR1 complexes jected with MFHR1 or hFH, although no changes in hema-
have higher inhibitory activity than monomeric MFHR1 frac- toxylin- and eosin–stained samples were detected (Figure 7, D
tions. and E). Injected proteins MFHR1 and hFH were detected in
the glomeruli of treated mice (Figure 7E, Supplemental Figure
MFHR1 Is Resistant to Competition by FHR-1 and 4). These data show that MFHR1 has the ability to reverse an
FHR-5 inherent complement defect in vivo or in vitro.
FHR-1 and FHR-5 act as competitive agonists for FH binding to
C3b and thereby, act as modulators of complement activation
(i.e., antagonists of FH-directed complement regulation). We DISCUSSION
speculated that dimeric or multimeric MFHR1 preparations
might influence its susceptibility to FHR competition. There- Hereditary or acquired complement defects may lead to con-
fore, we performed C3b binding ELISAs, where equimolar stitutive overactivation of the complement system downstream
amounts of MFHR1 or hFH were incubated with increasing of the defective effector protein. Therapeutic complement in-
amounts of FHR-1 or FHR-5 on C3b-coated microtiter plates. hibition focusing on C5 inhibition as a single target has im-
MFHR1 binding to C3b was completely resistant to competi- proved disease progression and survival, especially in patients
tion by FHR-1 or FHR-5 (up to 100-fold molar excess), with aHUS and patients with paroxysmal nocturnal hemoglo-
whereas hFH binding to C3b was affected by an already 3.1- binuria, but it only provides partial complement control and
fold molar excess of either FHR-1 or FHR-5 (Figure 6A). To misses full efficiency in C3G and other diseases associated with
determine whether MFHR1-mediated inhibition of comple- complement dysregulation.17–21
ment activation on cell surfaces is also resistant to competition Because the complement cascade produces active effector
by FHRs, we incubated sE with FH-depleted HS, leading to proteins on several levels, a multitargeted strategy could be
subsequent AP complement activation and hemolysis. beneficial for better control of a complex disease–causing im-
MFHR1 or hFH were added together with increasing amounts balance between complement activation and regulation.35,36
of FHR-5. Here again, MFHR1 was completely resistant to We, therefore, combined relevant complement regulatory
FHR-5–mediated competition up to 100-fold molar excess, domains of FHR-1 and FH to design a complement inhibitor
whereas hFH-mediated protection of sE was inhibited after that operates on different levels of the cascade (Supplemental
the addition of tenfold molar excess of FHR-5 (Figure 6B). Figure 5). This new fusion protein, MFHR1, strongly inhibited
complement activation as shown by distinct reductions of AP-
MFHR1 Controls Complement Activation In Vitro and and CP-mediated C5b-9 complex formations and consider-
Shows Therapeutic Benefit in C3G In Vivo ably exceeded the AP inhibitory activity of hFH (.50-fold),
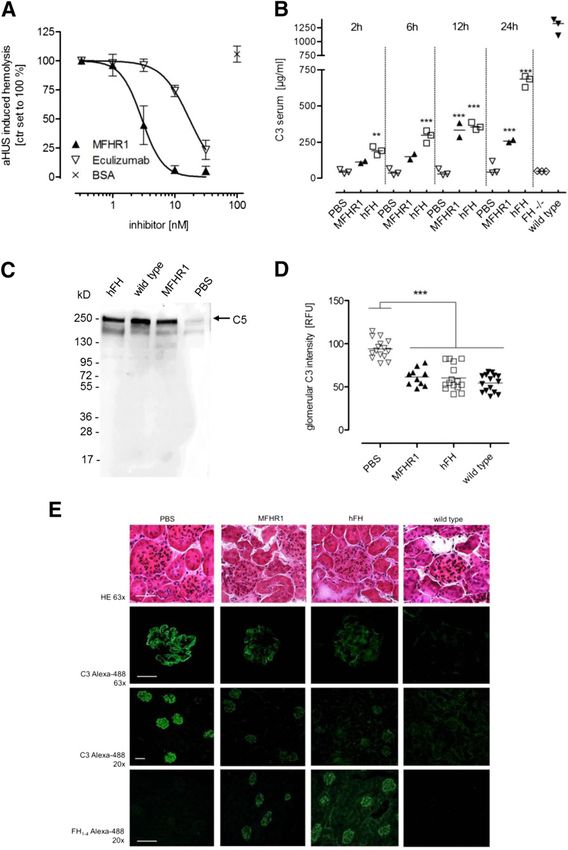
We then explored the capacity of MFHR1 to suppress comple- mini-FH (.3.5-fold), and the C5inhibitor eculizumab
ment activation under pathophysiologic conditions. Serum (greater than fivefold). It also prevented the release of the
derived from a patient with aHUS carrying a C-terminal FH potent immune modulators C3a and C5a in HS and com-
deletion39 caused complement overactivation and lysis of sE. pletely reversed complement overactivation after in vitro
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 5BASIC RESEARCH www.jasn.org Figure 3. MFHR1 displays cofactor and decay acceleration activity and blocks C3b surface deposition and C3a release (A) IC50 fitting curves for inhibition of a9-chain cleavage by MFHR1, FH1–4, FHR-11–2, or hFH. C3b and FI were incubated with increasing concen- trations of recombinant proteins or hFH (5–250 nM) for 30 minutes at 37°C. For quantification, intact C3b a9-chain densitometry was expressed relative to b-chain in each sample. Control without cofactor was set to 100% intact C3b a9-chain. Data represent mean values 6SD from n=5 experiments for MFHR1 and hFH or n=2 for FH1–4 and FHR-11–2. The cofactor activity of hFH was significantly different from MFHR1 (two-way ANOVA; P,0.001), whereas the activities of MFHR1 and FH1–4 were similar. (B) MFHR1 dissociates C3bBb. Convertase complexes were assembled on microtiter plates in the presence of C3b, factor B (FB), and factor D (FD), and MFHR1 or hFH was added and incubated at 37°C. Intact C3bBb and dissociation of these complexes were measured by the relative amount of FB. OD at 450 nm measured for control wells without regulator (C3b+FB+FD) was set to 100% FB. Negative control was performed without adding FD (C3b+FB). Data are mean6SD from n=3 experiments. The decay acceleration activities of MFHR1 and hFH were not significantly different from each other (two-way ANOVA). (C) IC50 fitting curves for inhibition of C3b deposition. MFHR1, hFH, FHR-1, or eculizumab was added to HS, and C3b depositions were measured after AP activation by LPS. HS or heat-inactivated (hi) HS added to wells not coated with LPS (2LPS) was used as control. Data points represent mean values from n=4 assays 6SEM using sera of four individual healthy donors. Serum without inhibitors was set to 100% C3b deposition for each individual experiment. The inhibitory activity of MFHR1 was significantly different from mini-FH (P,0.001; two-way ANOVA). (D) MFHR1 blocks generation of C3a as assayed in the supernatant of AP-activated sera using C3a ELISA (Quidel); hi HS was used as negative control. Data points are mean values from n=3 assays 6SEM using sera of three individual healthy donors. Untreated HS was set to 100% relative C3a. The C3a inhibitory activity of MFHR1 was not significantly different from mini-FH (two-way ANOVA). supplementation to serum of a patient with aHUS, showing FI.31,42 MFHR1 bound to the effector molecule C5 inhibited the effectiveness of MFHR1 in a disease model and proving the experimental C5 convertase and C5b-9–mediated lysis of sE concept of our multitarget approach. Furthermore, MFHR1 under serum-free conditions, presumably by inhibiting C5 partially inhibited serum AP activation and reduced patho- cleavage and binding to C5b6 complexes on cell surfaces as logic C3 depositions in FH2/2 mice at a comparable degree described for FHR-1.4,43 These results confirm that the fusion to recombinant FH,28 mini-FH,31 soluble CR1,32 and TT30,34 of FHR-11–2 to FH1–4 and FH19–20 is tolerated well without despite their individual mechanisms of action. In comparison steric hindrances, because full C5/C5b-9 and C3 regulatory with these recombinant proteins, plasma-derived hFH out- functional properties were both maintained by MFHR1. performs the in vivo efficacy, which might be explained by The strong inhibitory potency of MFHR1 to inhibit serum aspects of serum t1/2 and glycosylation. MFHR1 retained the AP–mediated C3b deposition on LPS-coated surfaces might C3 regulatory activity of FH as well as its host cell binding partly be explained by more accessible C3b/C3dg binding sites features. However, cofactor activity of MFHR1 was similar in FH19–20, targeting complement regulation to sites of ongo- to FH1–4 fragment but lower compared with hFH. This is in ing AP activation as was shown for mini-FH,29–31 but is likely line with earlier studies and confirmed that full-length FH dependent on the formation of multimers. Like native FHR-1, provides better structural rigidities than FH1–4 for optimally FHR-2, and FHR-5, MFHR1 contains a dimerization interface stabilizing the conformational state of C3b to be cleaved by localized within its N-terminal FHR-1–derived SCR1–2 6 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018
www.jasn.org BASIC RESEARCH
Table 2. Calculated IC50 values for cofactor activity in modulating FH regulation, especially on pathogen surfaces,
Inhibitor IC50 Values, nM R 2
95% Confidence Interval but they also have a pathologic role. In some subtypes of C3G,
hFH 4 0.83 2 to 7 gene rearrangements leading to duplication of the dimeriza-
MFHR1 35 0.85 17 to 71 tion domains and abnormal FHR proteins are held responsible
FH1–4 26 0.89 14 to 51 for a deregulation of complement activation on host surfaces
FHR-11–2 No activity — — and disease development.44–47 The dimerization motif in
MFHR1 mediates binding to FHR-1 and FHR-2 in vitro (Sup-
plemental Figure 6); however, complement regulation does
domain, and MFHR1 forms multimers that increase the in- not seem to be influenced negatively by these potential
hibitory activity of MFHR1. This is particularly remarkable, MFHR1-FHR heterodimers under physiologic conditions
because homo-, hetero-, or multimeric FHR complexes result (Figures 2, 3, B and C, and 4B). We further show that
in increased binding on cell surfaces or to complement acti- MFHR1 is resistant to competition by FHR-1 and FHR-5.
vation products (i.e., C3b and C3d).6,43 Alterations in local Therefore, the unique design of MFHR1 is expected to resist
composition and levels are believed to play a physiologic role inhibitory influences in subgroups of patients with C3G and
Figure 4. MFHR1 binds C5 and regulates terminal pathway activation by inhibition of C5 cleavage and MAC formation. (A) MFHR1
binds to C5 as determined by ELISA. Equimolar amounts of MFHR1, FHR-1, hFH, or BSA were immobilized to Nunc plates and in-
cubated with increasing concentrations of C5. Binding was detected using monoclonal C5 antibodies and HRP-labeled secondary
antibodies. Data are mean6SD from n=3 experiments. ***Statistical significance versus C5 bound to hFH (one-way ANOVA with
Bonferroni test; P,0.001). (B) MFHR1 blocks generation of C5a as measured in the supernatant of AP-activated sera using C5a ELISA
(Quidel). Heat-inactivated (hi) HS was used as negative control. Data points are mean values from n=3 assays 6SEM using sera of three
individual healthy donors. Untreated HS was set to 100% relative C5a. The C5a inhibitory activity of MFHR1 was significantly different
from mini-FH (P,0.10; two-way ANOVA), whereas the activities of MFHR1 and eculizumab were similar. (C) MFHR1 inhibits C5
cleavage and lysis of sE. CVFBb convertase was generated on sE after adding factor B (FB) and factor D (FD). Hemolysis was induced
after adding C5-C9 and detected at 414 nm. Preincubation of C5 with FHR-1, MFHR1, or eculizumab significantly inhibited hemolysis,
whereas hFH or BSA showed no inhibition. Control reactions (ctr) were performed without the addition of C5 (no C5b to generate lytic
complexes) or without FB/FD (no CVFBb convertase is formed to cleave C5 into C5a and activated C5b). C5 activation control without
regulators (w/o reg) was set to 100%. ***Statistical significance versus w/o reg (P,0.001; one-way ANOVA with Bonferroni test). (D)
MFHR1 inhibits formation of the MAC on sE. MAC formation on sE was induced by incubation with C5b6 and C7–C9 components and
detected by hemolysis of cells. Preincubation of C5b6 with MFHR1 or FHR-1 inhibited hemolysis at significant levels compared with
hFH, eculizumab, or BSA. MAC formation control without regulators (w/o reg) was set to 100%. Samples without C9 did not induce
hemolysis. All data represent mean values 6SD of three independent experiments. ***Statistical significance w/o reg (one-way ANOVA
with Bonferroni test; P,0.001).
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 7BASIC RESEARCH www.jasn.org Figure 5. Multimeric complexes increase AP regulatory activity of MFHR1. (A) Size exclusion chromatography (SEC) analysis of MFHR1, hFH, and BSA. The three compositions of BSA mixture presented different retention volumes on the basis of molecular mass, which was BSA trimer (I; 198 kD, 10.4 ml), BSA dimer (II; 132 kD, 11.5 ml), and BSA monomer (III; 66 kD, 13.4 ml). Under the same condition, hFH (9.3 ml) showed that the peak of protein species migrates as dimeric proteins at approximately 300 kD. MFHR1 showed a peak (I) at retention volume of 10 ml, indicating that MFHR1 migrates predominantly in a multimeric state in the fluid phase. Theoretical trimer (II), dimer (III), dimer intermediate (IV and V), and monomeric (VI) MFHR1 are indicated in the elution profile. (B) Analysis of MFHR1 after elution from the SEC column as performed in A. Purified MFHR1 (100 ng) or 1 ml protein from SEC fractions was loaded on a 10% SDS- PAGE and silver stained. (C) Single or pooled MFHR1 SEC fractions were added to HS at 10 nM, and AP activation was induced by incubation in LPS-coated wells. Regulatory efficiency of MFHR1 fractions was analyzed by measuring C5b-9 complex formation. Heat- inactivated (hi) HS or HS added to wells not coated with LPS (2LPS) was used as control. Bars represent single data points from n=1 assay. Serum without MFHR1 was set to 100% AP activity. might also benefit these patients; this, however, needs to overactivation, this multitarget strategy could serve as a blue- be proven. We hypothesize that MFHR1 uses the ability for print for developing new complement therapeutics. dimerization/multimerization of FHRs, increasing local concentrations and ligand avidity to increase its regulatory performance. CONCISE METHODS However, comparing their individual IC50 ratios for the C3b inhibition relative to C5b-9 inhibition, hFH (IC50 C3b- Cloning, Expression, Purification, and Characterization ELISA/IC50 C5b-9-ELISA =3.07) and mini-FH (3.25), both of Recombinant Proteins primarily acting at the C3 level, showed comparable quotients. For production of the recombinant fusion protein MFHR1 Because of its proportionately lower IC50 C5b-9 values than (FHR-11–2^FH1–4^FH19–20), mini-FH,30 FHR-1, FHR-2, FHR-5, IC50 C3b values, the quotient for MFHR1 (3.88) was higher, and truncated FHR-11–2 and FH1–4, appropriate gene fragments reflecting a proportionately higher terminal pathway inhibi- were cloned into a modified vector pFastBac-gp67-10His, and proteins tory activity. Therefore, MFHR1 acts on multiple therapeuti- were expressed in Spodoptera frugiperda cells following the manufac- cally relevant targets in the complement cascade. turer’s instructions (Bac-to-Bac System; Invitrogen, Life Technologies, In summary, we show that regulators affecting multiple Darmstadt, Germany). An N-terminally located decahistidine tag was effector sites simultaneously in the complement cascade, included, facilitating the purification of the recombinant protein. De- such as C3 inhibition, inhibition of C5 cleavage, and preven- tailed descriptions for the cloning of pFastBac-gp67-10His and re- tion of C5b-9 formation, have advantages compared with solely combinant protein fragments are provided in Supplemental Material. C3- or C5-targeted approaches. Particularly, because MFHR1 The supernatants were adjusted with 53 binding buffer (100 mM prevents the formation of proinflammatory effectors C3a and NaH2PO4 [Merck, Darmstadt, Germany], 2.5 M NaCl [VWR, Darm- C5a and the accumulation of C3 cleavage products, it is ex- stadt, Germany], and 100 mM imidazole [Roth, Steinheim, Germany], pected to also positively affect local inflammation and disease pH 7.4) and filtered using a 0.22-mm vacuum-driven bottle top filter progression in other complement-associated diseases. In ad- (Stericup; Merck Millipore, Darmstadt, Germany) before purification dition, resistance to competition and its ability for multime- of recombinant proteins was performed using HisTrap FF columns rization are unique features of MFHR1 and suggest a superior following the manufacturer’s instructions (GE Healthcare, Munich, effect of MFHR1 not only in FHR-related C3Gs. In addition to Germany) on an ÄktaBasic10 using unicorn 5.0 software (GE Health- the therapeutic potential of MFHR1 to regulate complement care). Protein fractions were concentrated using a vivaspin 8 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018
www.jasn.org BASIC RESEARCH
Figure 6. MFHR1 is resistant to deregulation by FHR-1 and FHR-5. (A) MFHR1 binding to C3b is not competed off by FHR-1 or FHR-5.
MFHR1 (triangles) or hFH (squares; termed test protein) was added to C3b-coated microtiter plates alone or with increasing con-
centrations of FHR-1 (black line) or FHR-5 (gray line) ranging from equimolar amounts to 100-fold excess. MFHR1 or hFH bound to C3b
was detected using specific antibodies. Average of n=3 assays 6SD is shown. *P,0.05; ***P,0.001. (B) MFHR1-mediated protection
of sE from serum-induced AP activation is not attenuated by FHR-5, whereas AP regulatory function of hFH was dose dependently
deregulated by FHR-5. MFHR1 or hFH was used at concentrations that reduce FH-depleted serum-induced lysis of sE to 50%, and
increasing concentrations of FHR-5 ranging from equimolar amounts to 100-fold excess were added. Hemolysis was determined at 414
nm, and data are expressed as relative hemolysis increase over samples where no FHR-5 has been added. Data represent mean values
from n=3 assays 6SD. Asterisks mark treatments significantly different from hFH or MFHR1 alone (one-way ANOVA with Bonferroni
test). **P,0.01.
polyethersulfon membrane filter (Sartorius AG, Goettingen, Ger- tubes (Sarstedt, Nurnbrecht, Germany), and immediately frozen at
many) with a 10-kD molecular mass cutoff. Buffer desalting was 280°C. This study was approved by the Ethics Committee Freiburg,
done by HiTrap desalting column (GE Healthcare) using NaPH buffer and written informed consent was provided for sample collection and
(50 mM NaH2PO4 and 150 mM NaCl) after volume reduction in subsequent analysis. FH-deficient serum was purchased from Com-
vivaspin concentration devices. Size exclusion chromatography for plement Technologies. The patient is a 15-year-old boy with aHUS
qualitative distribution of MFHR1 molecular species was evaluated that manifested in his first year. He shows a heterozygous C-terminal
in comparison with the retention time of hFH (Complement Tech- deletion of FH.39 He receives FFP every second week to substitute
nologies) and BSA (Sigma-Aldrich, Taufkirchen, Germany) on defective FH and has shown stable renal function over the last years
Superdex 200 10/300 GL (GE Healthcare) columns according to the without sign of active thrombotic microangiopathy events. Serum
manufacturer’s instructions. Proteins were separated by SDS-PAGE samples were collected before infusion of FFP.
and visualized by Pierce silver-staining kit (Thermo Fisher Scientific)
or Coomassie R-250 (Serva, Heidelberg, Germany) staining. Immu- Inhibition of AP/CP Activity in Human Serum
noblotting was performed by standard methods using anti-hFH For determination of AP activity, 96-well MaxiSorp plates were coated
(Complement Technologies), anti-FH1–4,48 anti-FH C18,37 anti– with 25 mg/ml LPS (Salmonella enteritidis; Sigma-Aldrich) diluted in
FHR-1/2 (Hycult Biotech, Uden, The Netherlands), and anti-His an- DPBS (Gibco). After two washing steps with PBS containing 0.05%
tibodies (QIAGEN, Hilden, Germany). Secondary antibodies used Tween (Sigma-Aldrich), wells were blocked with PBS/1% BSA
were anti-mouse HRP (GE Healthcare), anti-goat HRP (DAKO, Ech- (Thermo Scientific) at 37°C for 1 hour. Test proteins were diluted
ing, Germany), or anti-rabbit HRP (GE Healthcare) before detection in 80 ml GVB/Mg2+/EGTA (Complement Technologies) before 25 ml
with enhanced chemiluminescence (GE Healthcare) in the Fusion SL HS was added. After another washing step, 100 ml of test protein
Chemiluminescence system (Vilber Lourmat). The protein concen- serum mixture was transferred to the plate and incubated at 37°C
tration was determined by Pierce BCA Assay Kit (Thermo Fisher Sci- for 1 hour. For the C3b AP ELISA, wells were washed five times before
entific). Recombinant proteins were aliquoted and stored at 280°C. incubation with HRP-labeled C3b antibody (1:10,000 in PBS:1%
BSA; MP Biomed) for 30 minutes at room temperature, whereas
Proteins and Sera serum supernatants were directly transferred to 280°C for subse-
hFH, C3b, FI, C5, C5b6, C7, C8, and C9 were purchased from Com- quent analysis of C3a and C5a levels using MicroVue kits (Quidel).
plement Technologies, and factor B and factor D were purchased from After washing five times with PBS/0.05% Tween, detection of C3
Merck. CVF and C6 were purchased from Quidel (San Diego, CA), and deposition was performed by adding TMB substrate and H2SO4 to
eculizumab (Soliris) was obtained from remnants of infusions. stop the reaction. Finally, the absorbance was read at 450 nm using an
HS was obtained from healthy donors, and serum from a patient Epoch Microplate Spectrophotometer and Gen5 Software (BioTek,
with aHUS was collected by vein punctuation, allowed to clot in serum Bad Friedrichshall, Germany). For the C5b-9 AP ELISA, wells were
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 9BASIC RESEARCH www.jasn.org Figure 7. MFHR1 controls complement activation in vitro and shows therapeutic benefit in C3G in vivo. (A) IC50 fitting curves for inhibition of aHUS-induced lysis of sE. MFHR1 added to serum derived from a patient with aHUS (C-terminal depletion in FH39) protects sE from AP-mediated lysis with higher efficiency than eculizumab. Data represent mean values 6SEM from n=4 experiments. The inhibitory activity of MFHR1 was significantly different from eculizumab (P,0.001; two-way ANOVA). (B) C3 serum levels of FH2/2 mice after intraperitoneal injection of MFHR1 (n=2), hFH (n=3), or PBS (n=3). Mean values are shown with plotted individual data points. Serum C3 of wild-type mice was 12756122 mg/ml (n=3). (C) MFHR1 or hFH treatment of FH2/2 mice but not PBS treatment restores serum C5 as analyzed by Western blotting of serum after 24 hours. Serum of wild-type mice (FH+/+) was used as positive control. (D) MFHR1 and hFH reduce abnormal glomerular C3 depositions in FH2/2 mice. Glomerular C3 fluorescence immunostaining intensity was determined 24 hours after administration of MFHR1, hFH, or PBS to treated FH2/2 mice. Sections of untreated wild-type mice were used as negative control. Means are shown with plotted individual data points obtained from five glomeruli per section expressed as relative fluorescence units (RFUs). (E) Sections of glomeruli from MFHR1- or hFH-treated mice after 24 hours. Light microscopy images from hematoxylin and eosin (HE)–stained sections (HE 633) and representative immunofluorescence images of glomerular C3 depositions (C3 Alexa-488 633 and 203) and bound MFHR1 or hFH both detected with FH antibody (anti-FH1–4 Alexa-488 203). No abnormality could be assessed by HE staining on glomeruli from FH–/– mice treated with PBS, MFHR1, or hFH or wild-type mice at an age of 2 months. Immunofluorescence microscopy shows capillary wall and mesangial deposition of C3 in FH–/– mice, whereas 10 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018
www.jasn.org BASIC RESEARCH
washed three times before incubation with alkaline phosphatase– The inhibition of C5b-9 formation was followed using a modified
conjugated mAb (WIESLAB AP ELISA; Euro-Diagnostica, Malmo, protocol.4 In brief, C5b6 (0.7 nM) was incubated with 1.3 mM test
Sweden), recognizing the C9 neoantigen formed during C5b-9 as- protein for 5 minutes and added to C7 (9 nM), C8 (7 nM), C9
sembly, followed by incubation with alkaline phosphatase substrate (15 nM), and 53107 sEs. To measure the ability of MFHR1 to protect
(WIESLAB AP ELISA) solution for 30 minutes, and the absorbance sE from serum-induced cell lysis, MFHR1 or eculizumab in GVB was
was read at 405 nm. OD values for serum without regulatory proteins diluted to a final concentration ranging from 0.3 to 100 nM. Next,
were set to 100% activity; heat-inactivated HS or serum without LPS 53107 sEs were added, and the application of 20% serum of a patient
was used as the negative control. with aHUS or FH-deficient serum started the reaction. For all lytic
For analysis of CP activity, we used the WIESLAB CP ELISA (Euro- assays, reactions were stopped after incubation for 30 minutes at 37°C
Diagnostica) following the manufacturer’s instructions. Briefly, with GVB/EDTA buffer, and the supernatant was measured at 414 nm
MFHR1 was added to HS and incubated in wells precoated with in a microplate reader.
human IgM using CP-specific buffer conditions and C5b-9 forma-
tion, and the absorbance was read at 405 nm. Animals and Treatment Procedures
FH2/2 mice, provided by Matthew Pickering, were generated as de-
Cofactor and Decay Acceleration Activity Assay scribed.41 Animal procedures were performed in accordance with the
The cofactor activity was measured in a fluid-phase assay, and decay guide for the care and use of laboratory animals published by the US
acceleration activity was measured by ELISA as previously described.28 National Institutes of Health and the German animal protection
code, and they were approved by local authorities (Regierungspräsi-
dium Freiburg G-13/86). Mice were injected intraperitoneally with
Microtiter Binding and Competition ELISAs
For C5 ELISA, FHR-1, MFHR1, or hFH was immobilized to microtiter 0.5 mg MFHR1 or hFH or an identical volume of PBS; blood samples
plates (MaxiSorb) at equimolar concentrations (133 nM). After wash- were taken at serial time points, and renal tissue was taken after 24
ing with PBS/0.05% Tween and blocking with PBS/2% BSA, increasing hours. Serum C3 levels, immunostainings, and quantitative immu-
amounts of C5 were incubated. Bound C5 was detected using anti- nofluorescence were performed as previously described.28 Hematox-
human C5 antibody (Complement Technologies) followed by incu- ylin and eosin staining of kidney sections was performed using
bation with an HRP-conjugated rabbit anti-mouse serum (DAKO). standard staining procedures. MFHR1 or hFH was detected using
The C3b competition ELISA was adapted from the work by Goicoe- rabbit anti-FH1–448 after rabbit anti-goat Alexa 488–conjugated an-
chea de Jorge et al.6 Briefly, 25 mg/ml of C3b was coated on MaxiSorb tibodies (Invitrogen). For immune detection of C5 in mouse serum,
plates in a carbonate buffer (pH 9.6). After washing with PBS/0.01% 20 ml serum diluted 1:50 in PBS was resolved by SDS-PAGE under
Tween and blocking with PBS/2% BSA for 2 hours, 10 nM MFHR1 or nonreducing conditions and immunoblotting. C5 was detected with
hFH was incubated alone or with increasing concentrations of FHR-1 anti-C5 (Quidel) antisera after HRP-conjugated anti-goat antibodies.
or FHR-5 (10, 31.6, 100, 316, and 1000 nM), respectively, and applied
to the plate. After 2 hours at room temperature, plates were washed Statistical Analyses
four times, and C3b-bound MFHR1 or hFH was detected using anti- All graphs and statistics were created using Prism5 (GraphPad Soft-
hFH for FHR-5 competitive ELISA or anti-FH1–4 for FHR-1 compet- ware, San Diego, CA). The IC50 values were calculated by x=log(x)
itive ELISA. After 1 hour, detection was performed using anti-goat or data transformation, and curves were fitted using the nonlinear re-
anti-rabbit Ig HRP. After washing, ELISAs were developed, and the gression log (inhibitor) versus response (four parameters) setting.
OD at 450 nm was measured after stopping the reaction with 10% The goodness of the fit is indicated by R2 followed by the 95% con-
H2SO4 (Roth). fidence interval. One- or two-way ANOVA with Bonferroni multiple
comparisons test was used for statistical evaluation as indicated. Sta-
tistical significance was defined as ***P,0.001, **P,0.01, and
Hemolytic Assays *P,0.05.
To test C5 convertase inhibition, 25 nM CVF, 25 nM factor B, and
2.5 nM factor D were diluted in PBS/5 mM MgCl2 (Sigma-Aldrich) and
incubated for 1 hour at 37°C to build up experimental C5 convertase
(CVFBb). C5 (200 nM) was mixed with a 16-fold molar excess of test ACKNOWLEDGMENTS
proteins and incubated for 30 minutes at room temperature, and lytic
activity of the samples was measured after adding preassembled We thank Dr. Florian Grahammer and Prof. Dr. Tobias Huber for
CVF convertases to C5/inhibitor complexes to a mixture containing support of this work; Karin Sutter and Andrea Hartmann for technical
C6 (15 nM), C7 (15 nM), C8 (11 nM), C9 (24 nM), and 53107 sEs assistance; Prof. Dr. Matthew Pickering (Centre for Complement and
(Fiebig, Idar-Oberstein, Germany) in GVB/Mg2+/EGTA buffer. Inflammation Research, Imperial College London, London, United
abnormal C3 staining is reduced in MFHR1- or hFH-treated mice. No glomerular deposition of C3 is seen in wild-type mice. Asterisks mark
significant difference between the treatments and PBS group analyzed by one-way ANOVA with Bonferroni test. Magnification, 363 in rows
1 and 2; 320 in rows 3 and 4. Scale bars, 50 mm in rows 1 and 2; 100 mm in rows 3 and 4. **P,0.01; ***P,0.001.
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 11BASIC RESEARCH www.jasn.org
Kingdom) for providing the FH2/2 mice; and the Center for Ex- 9. Noris M, Remuzzi G: Glomerular diseases dependent on complement
perimental Models and Transgenic Services at the University of activation, including atypical hemolytic uremic syndrome, mem-
branoproliferative glomerulonephritis, and C3 glomerulopathy: Core
Freiburg for excellent animal care. We are grateful to Dr. Henning
curriculum 2015. Am J Kidney Dis 66: 359–375, 2015
Seismann and Prof. Dr. Martin Trepel (Department of Oncology and 10. Morgan BP, Harris CL: Complement, a target for therapy in in-
Hematology, University Medical Center Hamburg–Eppendorf) for flammatory and degenerative diseases. Nat Rev Drug Discov 14: 857–
supporting us with the baculovirus system and Anne Katrin Prowse 877, 2015
for proofreading of the manuscript. 11. Morigi M, Locatelli M, Rota C, Buelli S, Corna D, Rizzo P, Abbate M,
Conti D, Perico L, Longaretti L, Benigni A, Zoja C, Remuzzi G: A pre-
This study was supported by research funding from Greenovation
viously unrecognized role of C3a in proteinuric progressive nephrop-
Biotech GmbH (Heilbronn, Germany), the Excellence Initiative of the athy. Sci Rep 6: 28445, 2016
German Federal and State Governments (EXC 294), and contract 12. Locatelli M, Buelli S, Pezzotta A, Corna D, Perico L, Tomasoni S, Rottoli
research Glykobiologie/Glykomik of Baden–Württemberg Stiftung D, Rizzo P, Conti D, Thurman JM, Remuzzi G, Zoja C, Morigi M: Shiga
P-BWS-Glyko/13. P.F.Z. received funding from the European Com- toxin promotes podocyte injury in experimental hemolytic uremic
syndrome via activation of the alternative pathway of complement.
munity’s Seventh Framework Program grant 2012-305608 “Euro-
J Am Soc Nephrol 25: 1786–1798, 2014
pean Consortium for High-Throughput Research in Rare Kidney 13. Wada T, Nangaku M: Novel roles of complement in renal diseases and
Diseases (EURenOmics).” The study was supported by German Re- their therapeutic consequences. Kidney Int 84: 441–450, 2013
search Foundation Deutsche Forschungsgemeinschaft, SFB 1192 14. Riedl M: Recombinant human C1 esterase inhibitor in the management
Immune mediated glomerular diseases, project B06. of hereditary angioedema. Clin Drug Investig 35: 407–417, 2015
15. Legendre CM, Licht C, Muus P, Greenbaum LA, Babu S, Bedrosian C,
The funders had no role in study design, data collection and
Bingham C, Cohen DJ, Delmas Y, Douglas K, Eitner F, Feldkamp T,
analysis, decision to publish, or preparation of the manuscript. Fouque D, Furman RR, Gaber O, Herthelius M, Hourmant M, Karpman
D, Lebranchu Y, Mariat C, Menne J, Moulin B, Nürnberger J, Ogawa M,
Remuzzi G, Richard T, Sberro-Soussan R, Severino B, Sheerin NS,
DISCLOSURES
Trivelli A, Zimmerhackl LB, Goodship T, Loirat C: Terminal complement
Parts of this work contributed to an international patent application by the inhibitor eculizumab in atypical hemolytic-uremic syndrome. N Engl
University of Freiburg with S.M. and K.H. as inventors. S.M. and K.H. received J Med 368: 2169–2181, 2013
research funding from Greenovation Biotech GmbH. C.S. received funding 16. Al-Ani F, Chin-Yee I, Lazo-Langner A: Eculizumab in the management
from the Deutsche Forschungsgemeinschaft, Sk46. The remaining authors of paroxysmal nocturnal hemoglobinuria: Patient selection and special
declare no competing financial interests. considerations. Ther Clin Risk Manag 12: 1161–1170, 2016
17. Harder MJ, Kuhn N, Schrezenmeier H, Höchsmann B, von Zabern I,
Weinstock C, Simmet T, Ricklin D, Lambris JD, Skerra A, Anliker M,
Schmidt CQ: Incomplete inhibition by eculizumab: Mechanistic evi-
REFERENCES dence for residual C5 activity during strong complement activation.
Blood 129: 970–980, 2017
1. Gordon DL, Kaufman RM, Blackmore TK, Kwong J, Lublin DM: Identi- 18. Schalk G, Kirschfink M, Wehling C, Gastoldi S, Bergmann C, Hoppe B,
fication of complement regulatory domains in human factor H. Weber LT: A complicated case of atypical hemolytic uremic syndrome
J Immunol 155: 348–356, 1995 with frequent relapses under eculizumab. Pediatr Nephrol 30: 1039–
2. Weiler JM, Daha MR, Austen KF, Fearon DT: Control of the amplifica- 1042, 2015
tion convertase of complement by the plasma protein beta1H. Proc 19. Bomback AS, Smith RJ, Barile GR, Zhang Y, Heher EC, Herlitz L, Stokes
Natl Acad Sci U S A 73: 3268–3272, 1976 MB, Markowitz GS, D’Agati VD, Canetta PA, Radhakrishnan J, Appel
3. Skerka C, Chen Q, Fremeaux-Bacchi V, Roumenina LT: Complement GB: Eculizumab for dense deposit disease and C3 glomerulonephritis.
factor H related proteins (CFHRs). Mol Immunol 56: 170–180, 2013 Clin J Am Soc Nephrol 7: 748–756, 2012
4. Heinen S, Hartmann A, Lauer N, Wiehl U, Dahse HM, Schirmer S, Gropp 20. Lin Z, Schmidt CQ, Koutsogiannaki S, Ricci P, Risitano AM, Lambris JD,
K, Enghardt T, Wallich R, Hälbich S, Mihlan M, Schlötzer-Schrehardt U, Ricklin D: Complement C3dg-mediated erythrophagocytosis: Impli-
Zipfel PF, Skerka C: Factor H-related protein 1 (CFHR-1) inhibits com- cations for paroxysmal nocturnal hemoglobinuria. Blood 126: 891–894,
plement C5 convertase activity and terminal complex formation. Blood 2015
114: 2439–2447, 2009 21. Nishimura J, Yamamoto M, Hayashi S, Ohyashiki K, Ando K, Brodsky
5. Fritsche LG, Lauer N, Hartmann A, Stippa S, Keilhauer CN, Oppermann AL, Noji H, Kitamura K, Eto T, Takahashi T, Masuko M, Matsumoto T,
M, Pandey MK, Köhl J, Zipfel PF, Weber BH, Skerka C: An imbalance of Wano Y, Shichishima T, Shibayama H, Hase M, Li L, Johnson K,
human complement regulatory proteins CFHR1, CFHR3 and factor H Lazarowski A, Tamburini P, Inazawa J, Kinoshita T, Kanakura Y: Genetic
influences risk for age-related macular degeneration (AMD). Hum Mol variants in C5 and poor response to eculizumab. N Engl J Med 370:
Genet 19: 4694–4704, 2010 632–639, 2014
6. Goicoechea de Jorge E, Caesar JJ, Malik TH, Patel M, Colledge M, 22. Ricklin D, Lambris JD: Progress and trends in complement therapeutics.
Johnson S, Hakobyan S, Morgan BP, Harris CL, Pickering MC, Lea SM: Adv Exp Med Biol 735: 1–22, 2013
Dimerization of complement factor H-related proteins modulates 23. Mastellos DC, Yancopoulou D, Kokkinos P, Huber-Lang M,
complement activation in vivo. Proc Natl Acad Sci U S A 110: 4685– Hajishengallis G, Biglarnia AR, Lupu F, Nilsson B, Risitano AM, Ricklin D,
4690, 2013 Lambris JD: Compstatin: A C3-targeted complement inhibitor reaching
7. Mastellos DC, Ricklin D, Yancopoulou D, Risitano A, Lambris JD: its prime for bedside intervention. Eur J Clin Invest 45: 423–440, 2015
Complement in paroxysmal nocturnal hemoglobinuria: Exploiting our 24. Ricklin D, Lambris JD: Therapeutic control of complement activation at
current knowledge to improve the treatment landscape. Expert Rev the level of the central component C3. Immunobiology 221: 740–746,
Hematol 7: 583–598, 2014 2016
8. Barbour TD, Ruseva MM, Pickering MC: Update on C3 glomerulopathy. 25. Licht C, Weyersberg A, Heinen S, Stapenhorst L, Devenge J, Beck B,
Nephrol Dial Transplant 31: 717–725, 2016 Waldherr R, Kirschfink M, Zipfel PF, Hoppe B: Successful plasma
12 Journal of the American Society of Nephrology J Am Soc Nephrol 29: ccc–ccc, 2018www.jasn.org BASIC RESEARCH
therapy for atypical hemolytic uremic syndrome caused by factor H 38. Jore MM, Johnson S, Sheppard D, Barber NM, Li YI, Nunn MA, Elmlund
deficiency owing to a novel mutation in the complement cofactor H, Lea SM: Structural basis for therapeutic inhibition of complement
protein domain 15. Am J Kidney Dis 45: 415–421, 2005 C5. Nat Struct Mol Biol 23: 378–386, 2016
26. Licht C, Heinen S, Józsi M, Löschmann I, Saunders RE, Perkins SJ, 39. Gerber A, Kirchhoff-Moradpour AH, Obieglo S, Brandis M, Kirschfink
Waldherr R, Skerka C, Kirschfink M, Hoppe B, Zipfel PF: Deletion of Lys224 M, Zipfel PF, Goodship JA, Zimmerhackl LB: Successful (?) therapy of
in regulatory domain 4 of Factor H reveals a novel pathomechanism for hemolytic-uremic syndrome with factor H abnormality. Pediatr Nephrol
dense deposit disease (MPGN II). Kidney Int 70: 42–50, 2006 18: 952–955, 2003
27. Häffner K, Michelfelder S, Pohl M: Successful therapy of C3Nef-positive 40. Pickering MC, Warren J, Rose KL, Carlucci F, Wang Y, Walport MJ,
C3 glomerulopathy with plasma therapy and immunosuppression. Cook HT, Botto M: Prevention of C5 activation ameliorates spontane-
Pediatr Nephrol 30: 1951–1959, 2015 ous and experimental glomerulonephritis in factor H-deficient mice.
28. Michelfelder S, Parsons J, Bohlender LL, Hoernstein SNW, Proc Natl Acad Sci U S A 103: 9649–9654, 2006
Niederkrüger H, Busch A, Krieghoff N, Koch J, Fode B, Schaaf A, 41. Pickering MC, Cook HT, Warren J, Bygrave AE, Moss J, Walport MJ,
Frischmuth T, Pohl M, Zipfel PF, Reski R, Decker EL, Häffner K: Moss- Botto M: Uncontrolled C3 activation causes membranoproliferative
produced, glycosylation-optimized human factor H for therapeutic glomerulonephritis in mice deficient in complement factor H. Nat
application in complement disorders. J Am Soc Nephrol 28: 1462– Genet 31: 424–428, 2002
1474, 2017 42. Harder MJ, Anliker M, Höchsmann B, Simmet T, Huber-Lang M,
29. Schmidt CQ, Bai H, Lin Z, Risitano AM, Barlow PN, Ricklin D, Lambris Schrezenmeier H, Ricklin D, Lambris JD, Barlow PN, Schmidt CQ:
JD: Rational engineering of a minimized immune inhibitor with unique Comparative analysis of novel complement-targeted inhibitors, min-
triple-targeting properties. J Immunol 190: 5712–5721, 2013 iFH, and the natural regulators factor H and factor H-like protein 1 re-
30. Hebecker M, Alba-Domínguez M, Roumenina LT, Reuter S, Hyvärinen veal functional determinants of complement regulation. J Immunol
S, Dragon-Durey MA, Jokiranta TS, Sánchez-Corral P, Józsi M: An en- 196: 866–876, 2016
gineered construct combining complement regulatory and surface- 43. Hannan JP, Laskowski J, Thurman JM, Hageman GS, Holers VM:
recognition domains represents a minimal-size functional factor H. Mapping the Complement Factor H-Related Protein 1 (CFHR1):C3b/
J Immunol 191: 912–921, 2013 C3d interactions. PLoS One 11: e0166200, 2016
31. Nichols EM, Barbour TD, Pappworth IY, Wong EK, Palmer JM, Sheerin 44. Chen Q, Wiesener M, Eberhardt HU, Hartmann A, Uzonyi B, Kirschfink
NS, Pickering MC, Marchbank KJ: An extended mini-complement M, Amann K, Buettner M, Goodship T, Hugo C, Skerka C, Zipfel PF:
factor H molecule ameliorates experimental C3 glomerulopathy. Kid- Complement factor H-related hybrid protein deregulates complement
ney Int 88: 1314–1322, 2015 in dense deposit disease. J Clin Invest 124: 145–155, 2014
32. Zhang Y, Nester CM, Holanda DG, Marsh HC, Hammond RA, Thomas 45. Tortajada A, Yébenes H, Abarrategui-Garrido C, Anter J, García-
LJ, Meyer NC, Hunsicker LG, Sethi S, Smith RJ: Soluble CR1 therapy Fernández JM, Martínez-Barricarte R, Alba-Domínguez M, Malik TH,
improves complement regulation in C3 glomerulopathy. J Am Soc Bedoya R, Cabrera Pérez R, López Trascasa M, Pickering MC, Harris CL,
Nephrol 24: 1820–1829, 2013 Sánchez-Corral P, Llorca O, Rodríguez de Córdoba S: C3 glomerul-
33. Fridkis-Hareli M, Storek M, Mazsaroff I, Risitano AM, Lundberg AS, opathy-associated CFHR1 mutation alters FHR oligomerization and
Horvath CJ, Holers VM: Design and development of TT30, a novel C3d- complement regulation. J Clin Invest 123: 2434–2446, 2013
targeted C3/C5 convertase inhibitor for treatment of human complement 46. Gale DP, de Jorge EG, Cook HT, Martinez-Barricarte R, Hadjisavvas A,
alternative pathway-mediated diseases. Blood 118: 4705–4713, 2011 McLean AG, Pusey CD, Pierides A, Kyriacou K, Athanasiou Y,
34. Ruseva MM, Peng T, Lasaro MA, Bouchard K, Liu-Chen S, Sun F, Yu ZX, Voskarides K, Deltas C, Palmer A, Frémeaux-Bacchi V, de Cordoba SR,
Marozsan A, Wang Y, Pickering MC: Efficacy of targeted complement Maxwell PH, Pickering MC: Identification of a mutation in complement
inhibition in experimental C3 glomerulopathy. J Am Soc Nephrol 27: factor H-related protein 5 in patients of Cypriot origin with glomeru-
405–416, 2016 lonephritis. Lancet 376: 794–801, 2010
35. Melis JP, Strumane K, Ruuls SR, Beurskens FJ, Schuurman J, Parren PW: 47. Józsi M, Tortajada A, Uzonyi B, Goicoechea de Jorge E, Rodríguez de
Complement in therapy and disease: Regulating the complement Córdoba S: Factor H-related proteins determine complement-activating
system with antibody-based therapeutics. Mol Immunol 67[2 Pt A]: surfaces. Trends Immunol 36: 374–384, 2015
117–130, 2015 48. Kühn S, Skerka C, Zipfel PF: Mapping of the complement regula-
36. Korcsmáros T, Szalay MS, Böde C, Kovács IA, Csermely P: How to de- tory domains in the human factor H-like protein 1 and in factor H1.
sign multi-target drugs. Expert Opin Drug Discov 2: 799–808, 2007 J Immunol 155: 5663–5670, 1995
37. Oppermann M, Manuelian T, Józsi M, Brandt E, Jokiranta TS, Heinen S,
Meri S, Skerka C, Götze O, Zipfel PF: The C-terminus of complement
regulator Factor H mediates target recognition: Evidence for a compact
conformation of the native protein. Clin Exp Immunol 144: 342–352, This article contains supplemental material online at http://jasn.asnjournals.
2006 org/lookup/suppl/doi:10.1681/ASN.2017070738/-/DCSupplemental.
J Am Soc Nephrol 29: ccc–ccc, 2018 MFHR1 Is an Effective Complement Inhibitor 13You can also read

















































