Genotypic differences in root traits to design drought-avoiding soybean ideotypes
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
OCL 2022, 29, 26
© E. Dayoub et al., Published by EDP Sciences, 2022
https://doi.org/10.1051/ocl/2022021 OCL
Oilseeds & fats Crops and Lipids
Available online at:
www.ocl-journal.org
REVIEW
Genotypic differences in root traits to design drought-avoiding
soybean ideotypes☆
Elana Dayoub1,2,* , Jay Ram Lamichhane3 , Philippe Debaeke3 and Pierre Maury1
1
INP-ENSAT Toulouse, UMR AGIR, Université fédérale de Toulouse, 31326 Castanet-Tolosan, France
2
INP-Purpan, Université fédérale de Toulouse, 75 voie du TOEC, 31076 Toulouse, France
3
AGIR, Université fédérale de Toulouse, INRAE, 31326 Castanet-Tolosan, France
Received 8 December 2021 – Accepted 17 May 2022
Abstract – Soybean (Glycine max (L.) Merr.) may contribute to the agro-ecological transition of cropping
systems in Europe, but its productivity is severely affected by summer drought. The crop is mainly grown in
southern and continental parts of Europe, whereby increasing drought and heat waves are expected in the
near future. Agronomic strategies, such as early sowing, require cultivars with enhanced early plant growth
traits under suboptimal conditions. Moreover, efficient water uptake by root delays dehydration and
promotes drought avoidance. In general, changes in root morphology and root architecture are important
pathways for plant adaptation to water stress conditions. This paper reviews the cultivar differences in
soybean for root morphological and architectural traits especially during early growth stage. Previous works
reported cultivar differences for root traits in soybean but they did not deal with cultivars commonly grown
in Europe on which little information is available to date. Genotypic differences in available early-stage root
traits can be used as a framework to design soybean ideotypes less vulnerable to drought. To this aim, high-
throughput phenotyping supported by digital methods and crop modelling offer new avenues for the
exploration of target root traits involved in drought avoidance.
Keywords: early growth / ideotype / root traits / soybean cultivars / water deficit
Résumé – Différences génotypiques dans les caractéristiques des racines de soja pour concevoir
des idéotypes évitant la sécheresse. Le soja (Glycine max (L.) Merr.) peut contribuer à la transition agro-
écologique des systèmes de culture en Europe. Cependant, sa productivité est fortement impactée par la
sécheresse estivale. Cette culture est principalement pratiquée en Europe du Sud et continentale, où
sécheresse et vagues de chaleur plus fréquentes et plus intenses sont attendues à l’avenir. Les stratégies
agronomiques, telles que le semis précoce, nécessitent des cultivars ayant une croissance précoce accrue en
conditions sous-optimales. De plus, une absorption efficace de l’eau par les racines retarde la déshydratation
et participe à l’évitement de la sécheresse. En général, les changements dans la morphologie et l'architecture
des racines sont des voies importantes d'adaptation de la plante au stress hydrique. S’appuyant sur une revue
bibliographique, cet article vise à examiner les différences génotypiques chez le soja pour ce qui concerne
les traits morphologiques et architecturaux des racines, en particulier au stade précoce. Des travaux
précédents ont mis en évidence des différences entre cultivars pour les traits racinaires du soja, mais ils ne se
rapportaient pas à ceux couramment cultivés en Europe, pour lesquels peu d'informations sont disponibles à
ce jour. Les différences génotypiques pour les traits racinaires observées au stade précoce peuvent être
exploitées pour concevoir des idéotypes de soja moins vulnérables à la sécheresse. Dans ce but, le
phénotypage à haut débit soutenu par des méthodes numériques et la modélisation des cultures offrent de
nouvelles pistes pour l'exploration des traits racinaires cibles impliqués dans l'évitement de la sécheresse.
Mots clés : phase précoce / idéotype / traits racinaires / cultivars de soja / déficit hydrique
☆
Contribution to the Topical Issue “Soybean / Soja”.
*Correspondence: elana.dayoub@purpan.fr
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0), which permits
unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.E. Dayoub et al.: OCL 2022, 29, 26
Highlights reduced stomatal conductance, by reducing leaf absorption of
radiation or by enhancing the root water uptake through a
deeper rooting system. Deepening rooting system could be a
– A wide genetic variability exits for root architectural traits major adaptation trait to climate change for both increasing
among soybean cultivars. soybean yield and decreasing annual yield variability (Battisti
– Deep taproot, numerous lateral roots and wide root angle and Sentelhas, 2017). This literature review explores
are components of drought avoiding ideotype. genotypic differences in soybean root system with a particular
– Phenotyping methods at early growth provide proxy for focus on drought avoidance.
more advanced growth stages. The root system characterization is commonly based on
– Complementarity between structural and functional root structural (morphological and architectural) and functional
traits should be considered in future research. traits (e.g., water uptake and nutrients acquisition). A trait is
defined as a morphological, physiological or phenological
feature measurable at the individual level, from the cell to the
whole-organism level, without reference to the environment or
any other level of organization (Violle et al., 2007). Root
1 Introduction morphology refers to the surface features of a single root axis
as an organ, including characteristics of the epidermis such as
The global soybean production in 2019 reached number and length of roots hairs, root diameter, root length,
334 million tons of which 80% were achieved in Brazil, root surface area, root volume and specific root length (Lynch,
USA and Argentina only (FAOstat, 2021). In the European 1995). Root system architecture (RSA), defined as the spatial
Union, soybean production has increased progressively over configuration of a root system in the soil, is used to describe the
the last 5 years, reaching 2,8 million tons on 933,000 ha in shape and structure of roots such as width, depth, ratio of roots
2021 (European Commission, 2021). However, this production width to depth (De Dorlodot et al., 2007), and root growth
is still not sufficient to satisfy the increasing needs of the angle between lateral roots and soil surface (Zhao et al., 2004).
European market for food and feed requiring importation of In addition, RSA is determined by the interactions between
soybean from the American continent. Indeed, out of genetic and environmental factors making it highly plastic and
17 million tons of crude proteins imported annually in the able to respond to rapid environmental changes such as water
European Union, 13 million tons come from soybean, stress (Xiong et al., 2020) or waterlogging. On the other hand,
corresponding to 30 million tons of equivalent grains functional traits are defined as morpho-physio-phenological
(European Commission, 2018). In France, the strategic plan characteristics, which impact plant fitness indirectly via their
of the oil–protein sector aims at increasing the soybean acreage effects on plant performance (e.g., root traits involved in water
with an objective of 300,000 ha by 2030 (versus 186,000 ha in uptake efficiency) (Violle et al., 2007). However, the
2020). An increasing interest in this crop derives from the correlation between structural and functional traits is not
quality of its grain (~41% proteins) as well as from the straightforward and depends on plant growth environment.
agronomic and environmental benefits of this crop (diversifi- Identifying ideotypes for root structural traits (morphology
cation of crop rotation, lower need for pesticides, low and architecture) involved in drought avoidance is useful in
greenhouse gas emissions, biological nitrogen fixation, etc.) guiding the development of soybean cultivars to enhance soil
(Jouffret et al., 2015). exploration and thus water acquisition under water-limited
In Europe, soybean is mainly grown in southern and conditions. Many efforts in plant breeding have been made to
continental parts, where increasing drought and heat waves are improve drought avoidance and resource acquisition. Several
expected in the near future (Dai, 2013; Rojas et al., 2019). studies highlighted the interaction between structural and
Soybean yield and its stability are constrained by drought, functional root traits. Crops characterized by a large root
which is the most important limiting abiotic stress causing diameter could have an increased ability to penetrate the hard
yield losses (up to 40%), particularly when it occurs during soil (Bengough et al., 2011). Lynch (2013) showed that roots
both the vegetative and the reproductive stages (Specht et al., growing vertically at low metabolic cost (steep, cheap and
1999). However, the most critical period of water stress in this deep) contribute to build an interesting ideotype in maize since
crop occurs from flowering stage (Meckel et al., 1984). Several root system with rapid exploitation of deep soil could optimize
agronomic practices and adaptive strategies could be planned both water and N uptake. However, in the case of phosphorus
to counteract crop losses due to water stress and to promote the (immobile resource), soybean cultivars with root phenes, such
soybean acreage (Maury et al., 2015). One of these strategies is as shallower root growth angle of basal roots and long root
the choice of cultivars adapted to water-limited environments hairs, have been a good choice to enhance P acquisition in low-
through drought escape, dehydration tolerance and drought P soils (Lynch, 2011; Lynch and Brown, 2001; Richardson
avoidance (Turner et al., 2001). Drought escape is an et al., 2011; Wang et al., 2010). In addition, a relationship was
agronomic practice corresponding to the introduction of observed between root architecture and arbuscular mycorrhizal
early-maturing cultivars or shifting sowing dates earlier. fungi colonisation in soybean. The deep root genotype had
Dehydration tolerance is the ability of plant cells to continue greater colonisation by these fungi at low P compared to the
the metabolic process at low leaf water status by various shallow root genotype (Wang et al., 2011). Previous studies
physiological adaptations such as osmotic adjustment. On the reported some specific soybean root traits involved in water
other hand, drought avoidance (or dehydration postponement) uptake particularly under drought (Boote, 2011; Valliyodan et al.,
occurs when plants are able to keep a favourable water status 2017). On the other hand, functional traits, such as N2 fixation, are
under drought either by limiting water loss from leaves through related to root nodulation ability (Prudent et al., 2016) that could
Page 2 of 14Table 1. Main growth environments and associated root system phenotyping methods for soybean.
Growth Phenotyping method Advantages Disadvantages Reference
environment
Field Root excavation Real conditions, early and Heavy extraction, challenge of visualizing Allmaras et al. (1975); Pantalone et al.
advanced stages of roots, destructive, labour-intensive (1996a, 1996b); Zhao et al. (2004);
Benjamin and Nielsen (2006); Ao et al.
(2010); Matsuo et al. (2013); Fenta et al.
(2014); Wijewardana et al. (2019)
Root excavation/rhizotrons Visualisation of RSA in 2 dimensions Kaspar et al. (1978); Kaspar et al. (1984)
Semi-field Pots/tubes Close to the field conditions Gao et al. (2020)
Lysimeters/minirhizotrons or pots (the medium used may be soil or sand) Matsuo et al. (2013)
Greenhouse Plastic tubes (mesocosms) Easy root extraction, easily repeatable Only for early stages Fried et al. (2018); Thu et al. (2014)
Rhizotubes Non-destructive method No visualisation of RSA in 3 dimensions Maslard et al. (2021)
Rhizotrons Visualisation of RSA in 2 dimensions Dayoub et al. (2017)
Pots Rapid and low-cost Mwamlima et al. (2019)
Growth Minirhizotrons Dayoub et al. (2021)
chamber Germination paper Falk et al. (2020)
Cones/tubes Manavalan et al. (2010)
Page 3 of 14
E. Dayoub et al.: OCL 2022, 29, 26E. Dayoub et al.: OCL 2022, 29, 26
these techniques may not accurately reflect rooting system
morphology and architecture under field conditions, they can
be proposed as complementary approaches to overcome field
limitations. Under greenhouse environment, new image
analysis approaches were also implemented to characterize
soybean root architectural traits at high throughput (Maslard
et al., 2021). Although root phenotyping under controlled
conditions is a rapid, low-cost and adaptable method, further
experiments are required under field conditions to evaluate the
performance of cultivars and to show how shoot and root traits
are affected by various soil, climate and management
conditions.
3 Soybean root growth
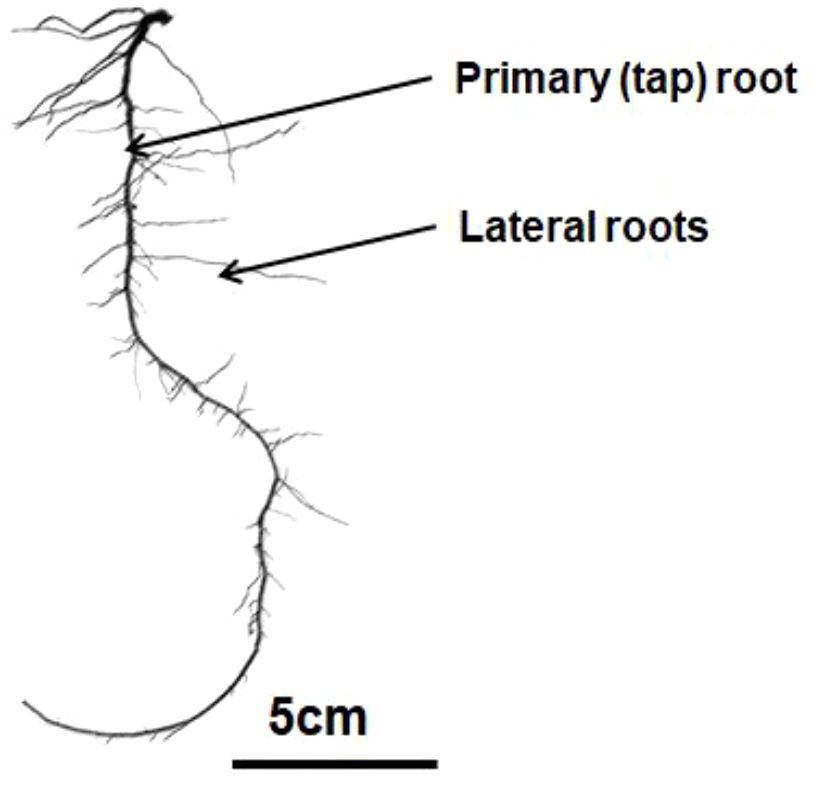
Soybean has a simple rooting system named allorhizic
(Fig. 1), consisting of a taproot and lateral roots (Ao et al.,
2010; Fenta et al., 2011, 2014). The taproot may reach a depth
Fig. 1. A characteristic allorhizic root system architecture of soybean of 200 cm and most lateral roots emerge from the upper 10 to
(cv. Isidor) at cotyledon stage (VC) (source: E. Dayoub, unpublished). 15 cm of the taproot (Lersten and Carlson, 2004). More than
50% of the roots are localised in the first 20 cm of soil layer
occur in interaction with structural root traits (morphology and (Hoogenboom et al., 1987). Moreover, 95–97% of the total
architecture) and may thus be involved in drought avoidance root weight and 85–70% of the total root surface area have
(Pantalone et al., 1996a, 1996b). been found in the upper 23 cm of soil (Benjamin and Nielsen,
For instance, the ability of roots to colonize the soil quickly 2006; Mitchell and Russell, 2010).
and effectively during crop establishment could be a major trait The soybean root consists of a simple architecture and
related to the competitiveness for resources. However, to date, morphology, which is similar to that of oilseed rape
only little is known on root traits involved in drought (Louvieaux et al., 2020). However, there is paucity of
avoidance in commonly grown soybean cultivars in Europe. information on the soybean rooting system compared to other
In the ongoing context of climate change, and with the crop species such as cereals (e.g., maize) or other leguminous
perspective of restrictive irrigation programs, new candidates species (e.g., common bean). Indeed, maize is characterized by
(ideotypes) are required in relation to root traits in soybean a complex rooting system composed of different root types viz.
cultivars under water-limited environments of Europe. The primary, crown, brace and seminal roots. Moreover, since
objectives of this paper were three-fold: lateral roots in maize are more metabolically demanding per
– to briefly report the main features of the soybean root gram of tissue than axial roots, a balance is required between
system and root phenotyping methods; the need of soil exploration and the metabolic need (Lynch,
– to review the genetic variability of root traits in soybean in 2013).
relation to drought avoidance; During the vegetative phase, soybean rooting can reach 30
– to highlight how the cultivar differences for these traits and 40 cm depth at 23 (second unrolled trifoliate leaf: V2) and
could be used to design soybean ideotypes avoiding 30 (third unrolled trifoliate leaf: V3) days after sowing,
drought stress. respectively (Fenta et al., 2014; Torrion et al., 2012). At the
beginning of flowering stage (R1), rooting depth may vary
from 50 to 70 cm (Böhm et al., 1977; Manavalan et al., 2010;
Matsuo et al., 2013). The maximum rooting depth in soybean
2 Root system phenotyping tools and differs between cultivars and could reach from 70 to 200 cm
methods (Lersten and Carlson, 2004). An in-depth growth of rooting
system could end between R1 to R3 (Kaspar et al., 1978), or
We have updated the list of root phenotyping methods, could continue until full seed stage (R6) (Torrion et al., 2012).
recently reviewed by Zhu et al. (2011) and Wasaya et al. On cultivars with indeterminate growth habit, the taproot
(2018), by integrating newly published studies in the literature extension follows a linear model with a progression of 1,3 cm
(Tab. 1). These studies proposed different methods for per day (Torrion et al., 2012). Soybean root system could
identifying root traits (morphology and architecture) under continue to grow after the beginning of pod filling but with no
laboratory and field conditions. Phenotyping techniques under further increase in root weight from the late bloom to mid-pod
controlled environments (growth chamber, greenhouse) are fill growth stages (Mitchell and Russell, 2010). However, the
much easier than those under field conditions due mainly to the root surface area increased between these two latter stages
easier extraction of roots. Several methods and approaches under both dryland and irrigated conditions (Benjamin and
could be proposed based on the nature of growth medium (gel, Nielsen, 2006). Therefore, there are contradictory information
liquid, or filter paper), which allow to measure a wide range of in the literature about soybean root growth in relation to
plants and root traits, mainly during early growth. Although cultivars and environmental conditions.
Page 4 of 14E. Dayoub et al.: OCL 2022, 29, 26
4 Genetic variability for root traits in the soil layers since the early development of seedling growth.
soybean However, soybean cultivars differ for early seedling growth
and vigour. A significant variability between cultivars was
noticed early in the crop cycle from 21 days after sowing
A number of previous studies reported a variability in root (~stage V3) for roots traits as taproot depth and root biomass
traits among soybean cultivars. This variability was observed (Manavalan et al., 2010; Matsuo et al., 2013). Differences
during early growth (Allmaras et al., 1975; Kaspar et al., 1978, among cultivars were also found as early as 12 days after
1984; Zhao, 2004; Ao et al., 2010; Manavalan et al., 2010; sowing for taproot progression in depth (Manavalan et al.,
Matsuo et al., 2013; Fenta et al., 2014; Thu et al., 2014; Fried 2010; Matsuo et al., 2013).
et al., 2018, 2019; Mwamlima et al., 2019; Falk et al., 2020;
Gao et al., 2020; Dayoub et al., 2021; Maslard et al., 2021), at
flowering stage (Zhao et al., 2004; Mwamlima et al., 2019) or 4.2 Rooting plasticity in early growth
at maturity (Ao et al., 2010). Phenotypic differences among
soybean cultivars for root traits have been well documented Unfavourable conditions during early growth may lead to
under different conditions (drought, phosphorus availability) yield losses due to reduced root growth. Flooding conditions
across many locations worldwide (Tabs. 2 and 3). In contrast, during two weeks after emergence caused yield damage even if
only a few studies focused on soybean cultivars commonly water conditions were optimal later in the crop cycle (Bajgain
grown in Europe (Dayoub et al., 2021; Maslard et al., 2021). et al., 2015) because of the inhibition of the rooting growth in
For most investigated root traits in the literature (Tabs. 2 and depth (Matsuo et al., 2013). Water stress during the vegetative
3), the extent of variation seems to be different as a function of phase in soybean or at the beginning of reproductive phase
cultivar, phenology and growth environment. (R1–R2) induced a significant increase in root system growth
(Manavalan et al., 2009). However, soybean undergoing water
stress before the flowering stage could be able to produce
4.1 Genetic variability in early growth higher yield than the crop undergoing water stress in post-
flowering thanks to early plasticity of rooting system (taproot
A wide genetic variability has been reported for most length and rooting density) (Hirasawa et al., 1994). Soil water
investigated architectural and morphological root traits during uptake during early season may differ among soybean cultivars
early growth (~stage V3) regardless of water regime or soil because of differences in root expansion and early plant
status. Architectural root traits such as rooting depth, root growth. However, few studies focused to date on the variability
angle and root width were significantly different among for early stage plant development among soybean cultivars
cultivars, irrespective of the water regime and growth grown in Europe under contrasted soil water regimes (Dayoub
environment (Matsuo et al., 2013; Fenta et al., 2014; Falk et al., 2021)
et al., 2020; Maslard et al., 2021). Root morphological traits as
root surface, root volume, taproot length, root tips number and
total root length were also different among cultivars (Fenta 5 Interactions between root growth and root
et al., 2014; Fried et al., 2018, 2019; Falk et al., 2020; Dayoub nodulation
et al., 2021). However, this variability was less noticeable for
average root diameter (Manavalan et al., 2010; Fenta et al., During early growth, symbiosis establishment and the
2014; Fried et al., 2018). Because the cultivars used so far varietal difference for root nodulation traits in soybean were
belonged to a wide range of maturity groups and were tested poorly highlighted in the literature, which might be due to
across different environments, the information presented in either the absence of nodulation (Dayoub, unpublished) or to
Tables 2 and 3 could help target root traits involved in drought the insignificant role of N2 fixation at early stage (Dayoub
avoidance mainly during early growth. et al., 2017). For example, although root nodulation was
Early growth is a critical phase of the crop cycle, which established, more than 90% of the plant N was derived from
spans from sowing until the beginning of competition among both seed N and soil N uptake at 35 days after sowing (stage
plants (crops or weeds) for the acquisition of trophic resources V3) under low N soil condition (Dayoub et al., 2017). This
(Boiffin et al., 1992; Fayaud et al., 2014). This phasic result illustrates that the complementarity between structural
sensitivity depends on species traits and sowing conditions, (root nodule) and functional root traits (biological N2 fixation)
including seedbed moisture. The latter, for example, is a key is not relevant during early growth. However, a large variation
factor for soybean establishment in southwestern France, in nodule number and size was found among soybean cultivars
affecting both seed germination and seedling emergence during later growth stages (Serraj et al., 1998; Fenta et al.,
(Lamichhane et al., 2020a, 2020b). Therefore, species or 2014).
cultivars characterized by an early and prompt ability to uptake Little is known to date on how symbiosis modifies root
resources during the first phase could show a higher growth and architecture in soybean (Concha and Doerner,
competitive ability for the acquisition of the same resources 2020; Maslard et al., 2021). Moreover, the correlations
later in the crop cycle (Fig. 2). Early seedling establishment of between the frequency and intensity of nodulation and root
soybean via an increased shoot and root vigour could thus be growth are still poorly understood, particularly the factors that
one of the most important traits for genotype selection in order control nodule density per unit root length in the absence or
to improve crop production under water-limited environments presence of stress (Kunert et al., 2016). The cultivar, which
(Manavalan et al., 2009; Thu et al., 2014). Since water is a was characterized by a great root length, surface, volume and
mobile resource, it may be advantageous to have a primary root tips number under drought, showed an increase in nodule
(taproot) and a network of lateral roots penetrating deeper into number and size (Fenta et al., 2014). Similarly, a positive
Page 5 of 14Table 2. Intraspecific variability in soybean cultivars for investigated morphological root traits.
Root traits Cultivar Mean Genotype Water regime Water regime Other Plant stage, days Growth Cultivar Maturity Location Reference
(per plant) range or cultivar effect effect condition after sowing (DAS) environment/ or genotype group
or days after phenotyping number
germination (DAG) method
Average root 0.50–0.56 0.53 * Well-watered * 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
diameter (mm) (volumetric soil water minirhizotrons
content 28%)
0.49–0.61 0.55 * Water stress (volumetric
soil water content
16.5%)
0.32–0.49 0.41 NS 42 DAS Greenhouse/mesocosms 49 From IV to VIII USA Fried et al. (2018)
0.43–0.49 0.46 NS Well-watered NS 60 DAS (V3) Field/root excavation 3 NA South Africa Fenta et al. (2014)
(volumetric soil water
content 14%)
0.40–0.54 0.47 ** Water stress from V3
(volumetric soil water
content
0.66–0.90 0.78 NS Well-watered (soil ** Flowering Greenhouse/pots 6 From 00 to II Kenya Mwamlima et al. (2019)
moisture 80% at field
capacity)
0.52–0.62 0.57 NS Water stress (soil **
moisture 20% at field
capacity)
Root biomass 0.01–0.26 0.135 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
(dry weight, g) germination paper
0.01–0.26 0.135 *** 9 DAG
0.01–0.26 0.135 *** 12 DAG
Page 6 of 14
0.02–0.04 0.03 *** Well-watered NS 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
(volumetric soil water minirhizotrons
content 28%)
0.02–0.03 0.025 *** Water stress (volumetric
soil water content
E. Dayoub et al.: OCL 2022, 29, 26
16.5%)
0.36–0.52 0.44 * 23 DAS (V1) Greenhouse/rhizotubes/ 9 I France Maslard et al. (2021)
throughput
0.54–0.70 0.62 NS 40 DAS Greenhouse/mesocosms 10 From V to VIII USA Fried et al. (2019)
2.09–3.61 2.85 *** Well-watered (soil *** Flowering Greenhouse/pots 6 From 00 to II Kenya Mwamlima et al. (2019)
moisture 80% at field
capacity)
0.40–0.84 0.62 *** Water stress (soil
moisture 20% at field
capacity)
Root length 0.06–0.16 0.11 *** Well-watered NS 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
density (volumetric soil water minirhizotrons
(cm cm-3) content 28%)
0.07–0.16 0.115 *** Water stress (volumetric
soil water content
16.5%)
0.92–1.84 1.38 NS 21 DAS Field 2 NA Japan Matsuo et al. (2013)
0.59–1.30 0.945 NS 41 DAS (R1)
0.54–1.18 0.86 * 62 DAS (R4)
0.84–1.21 1.025 NS 85 DAS (R6)
0.03–0.19 0.11 * 21 DASTable 2. (continued).
Root traits Cultivar Mean Genotype Water regime Water regime Other Plant stage, days Growth Cultivar Maturity Location Reference
(per plant) range or cultivar effect effect condition after sowing (DAS) environment/ or genotype group
or days after phenotyping number
germination (DAG) method
Lysimeter/
minirhizotrons
0.07–0.24 0.155 * 40 DAS
0.01–0.22 0.115 * 61 DAS
1.48–1.52 1.5 * 21 DAS Pots
4.26–4.88 4.57 NS 42 DAS
8.30–8.68 8.49 NS 56 DAS
0.50–2.50 1.5 * Water stress R2 Semi field/pots/tubes 8 NA China Gao et al. (2020)
Root surface 0.0–27.5 13.75 *** 6 DAG Growth chamber/ 292 USA Falk et al. (2020)
area (cm2) germination paper
0.0–59.0 29.5 *** 9 DAG
0.0–122.7 61.35 *** 12 DAG
15.8–38.1 26.95 *** Well-watered NS 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
(volumetric soil water minirhizotrons
content 28%)
20.0–37.2 28.6 *** Water stress (volumetric
soil water content
16.5%)
59.0–271.0 165 *** 42 DAS Greenhouse/mesocosms 49 From IV to VIII USA Fried et al. (2018)
406.0–622.0 514 NS 40 DAS Greenhouse/mesocosms 10 From V to VIII USA Fried et al. (2019)
277.0–548.0 412.5 *** * R7 Field 10
6.5–8.3 7.4 NS Well-watered 60 DAS (V3) Field/root excavation 3 NA South Africa Fenta et al. (2014)
(volumetric soil water
Page 7 of 14
content 14%)
4.7–15.4 10.05 ** Water stress from V3
(volumetric soil water
content
Root tips 86–204.3 145.2 *** Well-watered 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
E. Dayoub et al.: OCL 2022, 29, 26
number (volumetric soil water minirhizotrons
content 28%)
103.8–217.6 160.7 *** Water stress (volumetric
soil water content
16.5%)
168.7–190.5 179.6 NS Well-watered * 60 DAS (V3) Field/root excavation 3 NA South Africa Fenta et al. (2014)
(volumetric soil water
content 14%)
177.8–377.8 277.8 * Water stress from V3
(volumetric soil water
content
Root volume 0.10–0.54 0.32 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
and primary germination paper
root volume 0.10–0.71 0.405 *** 9 DAG
(cm3) 1.14–1.20 1.17 *** 12 DAG
0.22–0.47 0.345 *** Well-watered NS 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
(volumetric soil water minirhizotrons
content 28%)
0.27–0.46 0.365 *** Water stress (volumetric
soil water content
16.5%)Table 2. (continued).
Root traits Cultivar Mean Genotype Water regime Water regime Other Plant stage, days Growth Cultivar Maturity Location Reference
(per plant) range or cultivar effect effect condition after sowing (DAS) environment/ or genotype group
or days after phenotyping number
germination (DAG) method
0.45–3.52 1.985 * 42 DAS Greenhouse/mesocosms 49 From IV to VIII USA Fried et al. (2018)
4.56–7.79 6.175 * 40 DAS Greenhouse/mesocosms 10 From V to VIII USA Fried et al. (2019)
5.90–16.30 11.1 *** R7 Field 10
0.07–0.10 0.085 NS Well-watered * 60 DAS (V3) Field/root excavation 3 NA South Africa Fenta et al. (2014)
(volumetric soil water
content 14%)
0.05–0.16 0.105 ** Water stress from V3
(volumetric soil water
content
Taproot 0.2–38.2 19.2 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
length (cm) germination paper
1.7–54.4 28.05 *** 9 DAG
1.2–68.3 34.75 *** 12 DAG
18.0–30.5 24.25 * 12 DAS Greenhouse/plastic 13 NA Vietnam Thu et al. (2014)
tubes
36.2–59.3 47.75 * 21 DAS (V3)
9.4–30.2 19.8 *** 12 DAS Growth chamber/cones 34 From III to VI USA Manavalan
et al. (2010)
20.0–50.0 35 *** 21 DAS (V3) Growth chamber/tubes 8
50.0–70.0 60 *** 35 DAS
Total root 292
0.2–227.2 113.7 *** 6 DAG Growth chamber/ II and III USA Falk et al. (2020)
length (cm) germination paper
2.8–566.5 284.65 *** 9 DAG
Page 8 of 14
0.6–923.3 461.95 *** 12 DAG
93.9–247.6 170.75 *** Well-watered NS 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
(volumetric soil minirhizotrons
water content 28%)
106.2–241.9 174.05 *** Water stress (volumetric
E. Dayoub et al.: OCL 2022, 29, 26
soil
water content 16.5%)
2942–4107 3524.5 NS 40 DAS Greenhouse/Mesocosms 10 From V to VIII USA Fried et al. (2019)
646–1949 1297.5 *** 42 DAS Greenhouse/mesocosms 49 From IV to VIII USA Fried et al. (2018)
51.5–56.9 54.2 NS Irrigated (volumetric * 60 DAS (V3) Field/root 3 NA South Africa Fenta et al. (2014)
soil excavation
water content 14%)
41.0–120.5 80.75 ** Not irrigated from V3
(volumetric
soil water content
60.4–110.4 85.4 *** High phosphorus 100 DAS Field 8 China Zhao et al. (2004)
70.8–110.4 90.6 *** Low phosphorus China
132.8–416.6 274.7 *** High phosphorus Maturity Field 2 NA China Ao et al. (2010)
81.71–199.1 140.405 *** Low phosphorus
NS: non-significant, NA: not available.Table 3. Intraspecific variability in soybean cultivars for investigated architectural root traits.
Root traits (per Cultivar Mean Genotype or Water Water Other Plant stage, days after Growth Cultivar or Maturity group Location Reference
plant) range cultivar effect regime regime condition sowing (DAS) environment/ genotype
effect or days after phenotyping number
germination (DAG) method
Rooting depth 0.2–37 18.6 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
(cm) germination paper
0.3–49 24.65 *** 9 DAG
0–56 28 *** 12 DAG
43–56 49.5 * 35 DAS Field 2 NA Japan Matsuo et al. (2013)
62–93 77.5 * R1 Field/excavation of 7 NA USA Kaspar et al. (1978)
rhizotrons to 217 cm
deep
122–167 144.5 * R3
170–208 189 * R4
111–142 126.5 ** 54 DAS (R2 to R3) Field/excavation of 105 From I to III USA Kaspar et al. (1984)
rhizotrons
compartments
159–186 172.5 ** 75 DAS (R5)
16–27 21.5 ** High phosphorus Maturity Field 2 NA China Ao et al. (2010)
21–25 23 ** Low phosphorus
Root growth 2.0–90.0 46 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
angle (°) germination paper
Page 9 of 14
2.0–90.0 46 *** 9 DAG
2.0–90.0 46 *** 12 DAG
42.9–57.4 50.15 NS Well-watered *** 10 DAS Growth chamber/ 10 From 000 to II France Dayoub et al. (2021)
minirhizotrons
E. Dayoub et al.: OCL 2022, 29, 26
57.7–71.4 64.55 NS Water stress
23.3–64.2 43.75 ** Well-watered NS 60 DAS (V3) Field/root excavation 3 NA South Africa Fenta et al. (2014)
28.8–68.3 48.55 ** Water stress
Root 0.1–22.4 11.25 *** 6 DAG Growth chamber/ 292 II and III USA Falk et al. (2020)
width (cm) germination paper
0.1–33.8 16.95 *** 9 DAG
0.3–34.3 17.3 *** 12 DAG
15.3–24.6 19.95 * 23 DAS (V1) Greenhouse/rhizotubes/ 9 I France Maslard et al. (2021)
throughput
4.8–26.7 15.75 *** High phosphorus Maturity Field 2 NA China Ao et al. (2010)
1.33–18.33 9.83 *** Low phosphorus
NS: non-significant, NA: not available.E. Dayoub et al.: OCL 2022, 29, 26
On the other hand, root nodules are known to be crucial
sensors of drought, but their responses and their drought
tolerance features remain poorly characterized in the
literature for soybean. Root nodule number and size were
reduced 60 days after sowing for soybean cultivars under
drought (Fenta et al., 2014). Decreased N2 fixation in
response to drought leads to soybean yield losses (King and
Purcell, 2001; Sinclair et al., 2010). However, differences
exist among cultivars in sensitivity of N2 fixation to drought
(King and Purcell, 2001; Fenta et al., 2014). Some cultivars
can maintain more nodules under drought conditions, thus the
ability to form and sustain root nodules may also be an
important trait underpinning shoot productivity under drought
Fig. 2. Schematic representation of the importance of early growth (Fenta et al., 2014). Previous studies showed that a sustained
period for resource acquisition, rooting system plasticity and root nitrogen fixation is a major trait associated to drought
phenotyping ability (VE, VC and V3 indicates emergence, cotyledon tolerance in some soybean cultivars, which was due mainly to
and third-node stage, respectively (Fehr and Caviness, 1977). greater nodule size (Pantalone et al., 1996b; King and Purcell,
2001).
Soybean root traits involved in increased water acquisition
correlation was observed between an increase in the length and were defined by:
surface of fibrous roots and nodule number (Pantalone et al., – deeper rooting (and faster root growth in depth);
1996a). In contrast, the number of lateral roots was negatively – improved distribution of root length density into deeper
correlated to the rate of nodulation 23 days after transplanting soil layers;
for a range of soybean cultivars (Maslard et al., 2021). A – increased length per unit root mass;
previous study investigating the potential correlation between – increased assimilate partitioning to roots at the expense of
root architecture and nodulation found that the deep root shoot growth (constitutive);
genotype had better nodulation than the shallow root genotype – increased biomass partitioning to roots to increase root
but only under high P soil 60 days after planting (Wang et al., length density but only when induced by onset of water
2011). This trade-off between nodulation and root growth, deficit (adaptive) and delayed onset of seedling growth to
which is due mainly to the competition for carbon (Voisin increase assimilate to roots (Boote, 2011).
et al., 2003), needs to be investigated under various growing
environments.
A deeper taproot associated with a high density of lateral
roots leads to an increased total root surface area and thus
6 Root traits involved in drought avoidance water absorption from soil (Garay and Wilhelm, 1983;
Hufstetler et al., 2007; Matsuo et al., 2013). When subject
Soybean root traits play a key role for soil resource to water stress, soybean cultivars with a shallow root
acquisition and for improved crop performance under abiotic architecture (root angle 60°) (Fenta et al., 2014). Identifying
regime applied but this modification began mainly after early root traits in soybean cultivars will thus allow to find
growth period (from V3). An important increase in root tips candidates able to avoid drought that could be an essential step
number, root volume and root length was observed (Fenta in cultivar adaptations.
et al., 2014; Mwamlima et al., 2019; Dayoub et al., 2021).
However, root diameter was decreased under water stress
(Mwamlima et al., 2019). 7 Potential soybean ideotype for drought
Previous studies found significant correlations in soybean avoidance
between drought avoidance and root traits such as root
biomass, total root length, root volume and number of lateral A recent study investigating a range of soybean cultivars
roots (Liu et al., 2005; Read and Bartlett, 2006). Since soybean (Fig. 3) commonly grown in France and Europe has
yield under drought depends highly on both root depth and root proposed an ideotype (Fig. 4) in order to avoid drought
density, deeper rooting system may improve soybean yield. represented by a number of traits (morphology and
Some soybean cultivars are able to adapt to water stress architecture) (Dayoub et al., 2021). This study showed that
conditions by developing a deeper taproot and a large and cultivars characterized by high root depth and length, high
fibrous rooting system and by increasing the root/shoot root density and narrow root angle could be considered as
biomass ratio, which enable to reach deeper soil layers with good candidates to cope with water stress via better soil
available water (Manavalan et al., 2009; Boote, 2011). It is exploration. This study identified cultivar differences at the
well established that root length is one of the main traits that beginning of crop cycle (10 days after sowing); that will
support plants to tolerate the limited water condition during require a further validation later during the crop cycle and
early crop growth stage (Shao et al., 2008). under different environments.
Page 10 of 14E. Dayoub et al.: OCL 2022, 29, 26
Fig. 3. Overview of different root system morphologies for a range of soybean cultivars at cotyledon stage (VC) (Fehr and Caviness, 1977)
(credit: E. Dayoub).
Fig. 4. A framework to designing of a potential soybean ideotype for drought avoidance during early growth. The ideotype is characterized by a
high (green arrow) or low (red arrow) value depending on the considered trait.
8 Concluding remarks and perspectives 2001; He et al., 2017, 2021). Future studies should consider
these different soil situations in order to find a compromise for
root traits adapted for different growing environments.
Wide genetic variability for soybean root traits has been Root phenotyping techniques are still tedious to study
reported in the literature, particularly for root system differences in soybean cultivars under field conditions.
architecture traits involved in drought avoidance. The cultivars Consequently, little attention has been paid to date in analysing
studied were mainly those grown outside Europe and they the phenotypic differences in terms of root morphological,
belonged to later maturity groups than those grown in Europe, architectural and nodulation traits among European soybean
the latter ranging from 000(0) to II (III) from North to South of cultivars. Furthermore, the complementarity between structural
Europe. Soybean cultivars characterized by deeper taproot (morphological and architecture) and functional (water uptake
associated with a high contribution of lateral roots and wide and nutrients acquisition) traits of the root system should be
root angle could be considered as good candidates to cope with considered in future studies under different conditions.
water stress via a better soil exploration. On the other hand, Identifying root traits and classifying soybean cultivars
cultivars characterized by shallow rooting system with many according to these traits could be useful in the selection of
lateral roots could be adapted for environments with low soil cultivars adapted to water-limited environments. Phenotyping
phosphorus availability (Lynch, 2011; Lynch and Brown, methods at early growth stages in the laboratory provide
Page 11 of 14E. Dayoub et al.: OCL 2022, 29, 26
opportunities for high throughput phenotyping and may explain Battisti R, Sentelhas PC. 2017. Improvement of soybean resilience to
differences in vigour between soybean cultivars under water- drought through deep root system in Brazil. Agron J 109: 1612–
limited field conditions (Dayoub et al., 2021). Screening root 1622. https://doi.org/10.2134/agronj2017.01.0023.
traits at early stages in plant development can provide proxy for Bengough AG, McKenzie BM, Hallett PD, et al. 2011. Root
more advanced stages but such evaluation is needed on a case- elongation, water stress, and mechanical impedance: a review of
by-case basis in order to verify that early traits are related to limiting stresses and beneficial root tip traits. J Exp Bot 62: 59–68.
increased crop productivity under drought conditions (Comas https://doi.org/10.1093/jxb/erq350.
et al., 2013). Research aiming at the design of ideotypes should Benjamin JG, Nielsen DC. 2006. Water deficit effects on root
distribution of soybean, field pea and chickpea. Field Crop Res
be encouraged for European soybean cultivars to increase the
97: 248–253. https://doi.org/10.1016/j.fcr.2005.10.005.
market availability of soybean cultivars adapted for different
Böhm W, Maduakor H, Taylor HM. 1977. Comparison of five
sowing conditions. Cultivar differences in root traits reported at methods for characterizing soybean rooting density and
early stages could be used as a reference framework for ideotype development. Agron J 69: 415–419. https://doi.org/10.2134/
designing. To this aim, simple trait phenotyping methods under agronj1977.00021962006900030021x.
laboratory conditions should be developed and evaluated to Boiffin J, Durr C, Fleury A, Maillet I, Marin-Laflèche A. 1992.
assist the selection of drought avoiding soybean cultivars. Analysis of the variability of sugar beet (Beta vulgaris L) growth
Crop growth models could be suitable tools for evaluating during the early stages. I. Influence of various conditions on crop
and designing ideotypes adapted to a range of environments establishment. Agronomie 12: 515–525. https://doi.org/10.1051/
(Sinclair et al., 2010; Rötter et al., 2015). In most of the 1D agro:19920703.
field crop models, a limited number of parameters are required Boote KJ. 2011. Improving soybean cultivars for adaptation to
for describing the growth of the rooting system and its climate change and climate variability. In: Yadav, S.S., Redden, R.
dynamics under ideal and water-limited conditions (Jones J., Hatfield, J.L., Lotze-Campen, H., Hall, A.E., eds. Crop
et al., 1991; Calmon et al., 1999; Brisson et al., 2009): e.g., adaptation to climate change. Chichester, West Sussex, PO19
maximum root depth, root front velocity, root length density 8SQ, UK: John Willey and Sons Inc., pp. 370–395.
distribution with depth, fraction of biomass partitioned into Brisson N, Launay M, Mary B, Beaudoin N. 2009. Conceptual basis,
roots. However, the parameterization of these models is formalisations, and parameterization of the Stics crop model.
generally only achievable at species level (e.g., soybean) and Versailles, France: Éditions Quae.
seldom at the cultivar level. Due to the progress in phenotyping Calmon MA, Batchelor WD, Jones JW, Ritchie JT, Boote KJ,
methods, one can expect to have an easier future access to both Hammond LC. 1999. Simulating soybean root growth and soil
above- and below-ground plant parameters with an increased water extraction using a functional crop model. Trans ASAE 42:
1867–1877.https://doi.org//10.13031/2013.13352.
accuracy, a high throughput and for a wide range of inbred
Comas L, Becker S, Cruz VM, Byrne PF, Dierig DA. 2013. Root
lines and cultivars. The evaluation of root traits in young plants
traits contributing to plant productivity under drought. Front
(e.g., root angle) could bring useful information for predicting Plant Sci 4: 442. https://www.frontiersin.org/article/10.3389/
the final development of the rooting system, and thus could be fpls.2013.00442.
used for the parameterization of crop models at the cultivar Concha C, Doerner P. 2020. The impact of the rhizobia-legume
level. This will facilitate the virtual evaluation of soybean symbiosis on host root system architecture. J Exp Bot 71: 3902–
cultivars across a wider range of environments under current 3921. https://doi.org/10.1093/jxb/eraa198.
and future climates. Dai A. 2013. Increasing drought under global warming in
Acknowledgments. The authors thank all partners of the observations and models. Nature Clim Change 3: 52–58.
https://doi.org/10.1038/nclimate1811.
“Sojamip” research project and the UMT Pactole. Special
Dayoub E, Lamichhane JR, Schoving C, Debaeke P, Maury P. 2021.
thanks to Bé atrice Quinquiry, to the other technicians of the
Early-stage phenotyping of root traits provides insights into the
Vasco research team (UMR AGIR), and to Terres Inovia, drought tolerance level of soybean cultivars. Agronomy 11: 188.
LIDEA, and RAGT 2n for their kind support during this study. https://doi.org/10.3390/agronomy11010188.
Dayoub E, Naudin C, Piva G, Shirtliffe SJ, Fustec J, Corre-Hellou G.
Conflicts of interest. The authors declare that they have no 2017. Traits affecting early season nitrogen uptake in nine legume
conflicts of interest in relation to this article. species. Heliyon 3: e00244. https://doi.org/10.1016/j.heli
yon.2017.e00244.
References De Dorlodot S, Forster B, Pagès L, Price A, Tuberosa R, Draye X.
2007. Root system architecture: opportunities and constraints for
Allmaras RR, Nelson WW, Voorhees WB. 1975. Soybean and corn genetic improvement of crops. Trends Plant Sci 12: 474–481.
rooting in southwestern Minnesota: II. Root distributions and https://doi.org/10.1016/j.tplants.2007.08.012.
related water inflow. Soil Sci Soc Am J 39: 772–777. https://doi. European Commission. 2018. Report from the Commission to the
org/10.2136/sssaj1975.03615995003900040046x. Council and the European Parliament on the development of
Ao J, Fu J, Tian J, Yan X, Liao H. 2010. Genetic variability for root plant proteins in the European Union. COM/2018/757 final.
morph-architecture traits and root growth dynamics as related to Document 52018DC0757. Available from https://eur-lex.europa.
phosphorus efficiency in soybean. Funct Plant Biol 37: 304–312. eu/homepage.html.
https://doi.org/10.1071/fp09215. European Commission. 2021. Crops market observatory: oilseeds
Bajgain R, Kawasaki Y, Akamatsu Y, et al. 2015. Biomass production and protein crops statistics. Available from https://ec.europa.eu/
and yield of soybean grown under converted paddy fields with info/food-farming-fisheries/farming/facts-and-figures/markets/
excess water during the early growth stage. Field Crop Res 180: overviews/market-observatories/crops/oilseeds-and-protein-crop
221–227. https://doi.org/10.1016/j.fcr.2015.06.010. s_en (March 28, 2022).
Page 12 of 14E. Dayoub et al.: OCL 2022, 29, 26
Falk KG, Jubery TZ, O’Rourke J.A., et al. 2020. Soybean root system Kaspar TC, Stanley CD, Taylor HM. 1978. Soybean root-growth during
architecture trait study through genotypic, phenotypic, and shape- reproductive stages of development. Agron J 70: 1105–1107.
based clusters. Plant Phenom 2020: 1925495. https://doi.org/ https://doi.org/10.2134/agronj1978.00021962007000060051x.
10.34133/2020/1925495. King CA, Purcell LC. 2001. Soybean nodule size and relationship to
Fayaud B, Coste F, Corre-Hellou G, Gardarin A, Dürr C. 2014. nitrogen fixation response to water deficit. Crop Sci 41: 1099–
Modelling early growth under different sowing conditions: a tool 1107. https://doi.org/10.2135/cropsci2001.4141099x.
to predict variations in intercrop early stages. Eur J Agron 52: Kaspar TC, Taylor HM, Shibles RM. 1984. Taproot-elongation rates
180–190. https://doi.org/10.1016/j.eja.2013.09.009. of soybean cultivars in the glasshouse and their relation to field
Fehr WR, Caviness CE. 1977. Stages of soybean development. Iowa rooting depth. Crop Sci 24: 916–920. https://doi.org/10.2135/
Coop Ext Serv Iowa Agric Home Econ Exp Stn Spec Rep 80: 11. cropsci1984.0011183x002400050021x.
http://lib.dr.iastate.edu/specialreports/87. Kunert KJ, Vorster BJ, Fenta BA, Kibido T, Dionisio G, Foyer CH.
Fenta BA, Barlow K, Burridge J, et al. 2014. Field phenotyping of 2016. Drought stress responses in soybean roots and nodules.
soybean roots for drought stress tolerance. Agronomy 4: 418–435. Front Plant Sci 7: 1015. https://doi.org/10.3389/fpls.2016.01015.
https://doi.org/10.3390/agronomy4030418. Lamichhane JR, Aubertot J-N, Champolivier L, Debaeke P, Maury P.
Fenta BA, Schlüter U, Garcia BM, DuPlessis M, Foyer CH, Kunert KJ. 2020a. Combining experimental and modeling approaches to
2011. Identification and application of phenotypic and molecular understand genotype × sowing date × environment interaction
markers for abiotic stress tolerance in soybean. In: Krezhova, D., ed. effects on emergence rates and grain yield of soybean. Front Plant
Soybean—Genetics and novel techniques for yield enhancement. Sci 11: 558855 https://doi.org/10.3389/fpls.2020.558855.
Shanghai, China: InTech, pp. 181–200. Lamichhane JR, Constantin J, Schoving C, et al. 2020b. Analysis of
Fried HG, Narayanan S, Fallen B. 2018. Characterization of a soybean germination, emergence, and prediction of a possible
soybean (Glycine max L. Merr.) germplasm collection for root northward establishment of the crop under climate change. Eur J
traits. PLoS One 13: e0200463. https://doi.org/10.1371/journal. Agron 113: 125972. https://doi.org/10.1016/j.eja.2019.125972.
pone.0200463. Lersten NR, Carlson JB. 2004. Vegetative morphology. In: Boerma,
Fried HG, Narayanan S, Fallen B. 2019. Evaluation of soybean H.R., Specht, J.E., eds. Soybeans: improvement, production, and
[Glycine max (L.) Merr.] genotypes for yield, water use efficiency, uses, 3rd ed. Agronomy Monograph 16. Madison, WI, USA:
and root traits. PLoS One 14: e0212700. https://doi.org/10.1371/ ASA-CSSA-SSSA, pp. 15–57.
Liu F, Andersen MN, Jacobsen SE, Jensen CR. 2005. Stomatal control
journal.pone.0212700.
and water use efficiency of soybean (Glycine max L. Merr.) during
Garay AF, Wilhelm W. 1983. Root system characteristics of two
progressive soil drying. Environ Exp Bot 54: 33–40. https://doi.
soybean isolines undergoing water stress condition. Publ. from
org/10.1016/j.envexpbot.2004.05.002.
USDA-ARS/UNL Fac. 134.
Louvieaux J, Spanoghe M, Hermans C. 2020. Root morphological
Gao X-B, Guo C, Li F-M, Li M, He J. 2020. High soybean yield and traits of seedlings are predictors of seed yield and quality in winter
drought adaptation being associated with canopy architecture,
oilseed rape hybrid cultivars. Front Plant Sci 11: 568009. https://
water uptake, and root traits. Agronomy 10(4): 608. https://doi. doi.org/10.3389/fpls.2020.568009.
org/10.3390/agronomy10040608. Lynch JP. 1995. Root architecture and plant productivity. Plant
He J, Jin Y, Du Y-L, et al. 2017. Genotypic variation in yield, yield Physiol 109: 7–13. https://doi.org/10.1104/pp.109.1.7.
components, root morphology and architecture, in soybean in Lynch JP. 2011. Root phenes for enhanced soil exploration and
relation to water and phosphorus supply. Front Plant Sci 8: 1499. phosphorus acquisition: tools for future crops. Plant Physiol 156:
https://doi.org/10.3389/fpls.2017.01499. 1041–1049. https://doi.org/10.1104/pp.111.175414.
He J, Jin Y, Siddique KHM, Li F-M. 2021. Trade-off between Lynch JP. 2013. Steep, cheap and deep: an ideotype to optimize water
root efficiency and root size is associated with yield and N acquisition by maize root systems. Ann Bot 112: 347–357.
performance of soybean under different water and phosphorus https://doi.org/10.1093/aob/mcs293.
levels. Agriculture 11: 481. https://doi.org/10.3390/agricul Lynch JP, Brown KM. 2001. Topsoil foraging – An architectural
ture11060481. adaptation of plants to low phosphorus availability. Plant Soil
Hirasawa T, Tanaka K, Miyamoto D, Takei M, Ishihara K. 1994. 237: 225–237. https://doi.org/10.1023/A:1013324727040.
Effects of pre-flowering soil moisture deficits on dry matter Manavalan LP, Guttikonda SK, Nguyen VT, Shannon JG, Nguyen HT.
production and ecophysiological characteristics in soybean plants 2010. Evaluation of diverse soybean germplasm for root growth and
under drought conditions during grain filling. Jpn J Crop Sci 63: architecture. Plant Soil 330: 503–514. https://doi.org/10.1007/
721–730. https://doi.org/10.1626/jcs.63.721. s11104-009-0222-8.
Hoogenboom G, Huck MG, Peterson CM. 1987. Root growth rate of Manavalan LP, Guttikonda SK, Phan Tran LS, Nguyen HT. 2009.
soybean as affected by drought stress Agron J 79: 607–614. Physiological and molecular approaches to improve drought
https://doi.org/10.2134/agronj1987.00021962007900040004x. resistance in soybean. Plant Cell Physiol 50: 1260–1276. https://
Hufstetler EV, Boerma HR, Carter TE, Earl HJ. 2007. Genotypic doi.org/10.1093/pcp/pcp082.
variation for three physiological traits affecting drought tolerance Maslard C, Arkoun M, Salon C, Prudent M. 2021. Root architecture
in soybean. Crop Sci 47: 25–35. https://doi.org/10.2135/ characterization in relation to biomass allocation and biological
cropsci2006.04.0243. nitrogen fixation in a collection of European soybean genotypes.
Jones CA, Bland WL, Ritchie JT, Williams JR. 1991. Simulation of OCL 28: 48. https://doi.org/10.1051/ocl/2021033.
root growth. In: Hanks, R.J., Ritchie, J.T., eds. Modeling plant Matsuo N, Takahashi M, Fukami K, Tasaka K, Tsuchiya S. 2013. Root
and soil systems. Agronomy Monographs 31. Madison, Wi growth of two soybean [Glycine max (L.) Merr.] cultivars grown
(USA): ASA/CSSA/SSSA Publishers, pp. 91–123. https://doi. under different groundwater level conditions. Plant Prod Sci 16:
org/10.2134/agronmonogr31. 374–382. https://doi.org/10.1626/pps.16.374.
Jouffret P, Labalette F, Parachini E. 2015. Analyse multicritère de la Maury P, Andrianasolo FN, Alric F, et al. 2015. Le semis très précoce :
production de soja dans des exploitations agricoles contrastées du une stratégie agronomique pour améliorer les performances
Sud-Ouest de la France. OCL 22: D505. https://doi.org/10.1051/ du soja en France ? OCL 22: D503. https://doi.org/10.1051/ocl/
ocl/2015029. 2015028.
Page 13 of 14E. Dayoub et al.: OCL 2022, 29, 26
Meckel L, Egli DB, Phillips RE, Radcliffe D, Leggett JE. 1984. Effect of Thu NBA, Nguyen QT, Hoang XLT, Thao NP, Tran L-SP. 2014.
moisture stress on seed growth in soybeans. Agron J 76: 647–650. Evaluation of drought tolerance of the Vietnamese soybean
https://doi.org/10.2134/agronj1984.00021962007600040033x. cultivars provides potential resources for soybean production and
Mitchell RL, Russell WJ. 2010. Root development and rooting genetic engineering. BioMed Res Int 2014: 809736. https://doi.
patterns of soybean (Glycine max (L.) Merrill) evaluated under org/10.1155/2014/809736.
field conditions. Agron J 63: 313. https://doi.org/10.2134/ Torrion JA, Setiyono TD, Cassman KG, Ferguson RB, Irmak S,
agronj1971.00021962006300020034x. Specht JE. 2012. Soybean root development relative to vegetative
Mwamlima LH, Ouma JP, Cheruiyot EK. 2019. Soybean (Glycine and reproductive phenology. Agron J 104: 1702–1709. https://doi.
max (L) Merrill) root growth and nodulation responses to different org/10.2134/agronj2012.0199.
soil moisture regimes. J Crop Sci Biotechnol 22: 153–159. https:// Turner NC, Wright GC, Siddique KHM. 2001. Adaptation of grain
doi.org/10.1007/s12892-019-0045-0. legumes (pulses) to water-limited environments. Adv Agron 71:
Pantalone VR, Burton JW, Carter TE. 1996a. Soybean fibrous root 193–231. https://doi.org/10.1016/s0065-2113(01)71015-2.
heritability and genotypic correlations with agronomic and seed Valliyodan B, Ye H, Song L, Murphy M, Grover Shannon J, Nguyen HT.
quality traits. Crop Sci 36: 1120–1125. https://doi.org/10.2135/ 2017. Genetic diversity and genomic strategies for improving
cropsci1996.0011183X003600050008x. drought and waterlogging tolerance in soybeans. J Exp Bot 68:
Pantalone VR, Rebetzke GJ, Burton JW, Carter TE. 1996b. 1835–1849. https://doi.org/10.1093/jxb/erw433.
Phenotypic evaluation of root traits in soybean and applicability Violle C, Navas ML, Vile D, et al. 2007. Let the concept of trait be
to plant breeding. Crop Sci 36: 456–459. https://doi.org/10.2135/ functional. Oikos 116: 882–892. https://doi.org/10.1111/
cropsci1996.0011183X003600020039x. j.0030-1299.2007.15559.x.
Prudent M, Vernoud V, Girodet S, Salon C. 2016. How nitrogen Voisin AS, Salon C, Jeudy C, Warembourg FR. 2003. Symbiotic N2
fixation is modulated in response to different water availability fixation activity in relation to C economy of Pisum sativum L. as a
levels and during recovery: a structural and functional study at the function of plant phenology. J Exp Bot 54: 2733–2744. https://doi.
whole plant level. Plant Soil 399: 1–12. https://doi.org/10.1007/ org/10.1093/jxb/erg290.
s11104-015-2674-3. Wang X, Pan Q, Chen F, Yan X, Liao H. 2011. Effects of co-
Read DJ, Bartlett EM. 2006. The physiology of drought resistance in inoculation with arbuscular mycorrhizal fungi and rhizobia on
the soybean plant (Glycine max). I. The relationship between soybean growth as related to root architecture and availability of
drought resistance and growth. J Appl Ecol 9: 487–499. https:// N and P. Mycorrhiza 21: 173–181. https://doi.org/10.1007/
doi.org/10.2307/2402447. s00572-010-0319-1.
Richardson AE, Lynch JP, Ryan PR, et al. 2011. Plant and microbial Wang X, Yan X, Liao H. 2010. Genetic improvement for phosphorus
strategies to improve the phosphorus efficiency of agriculture. Plant efficiency in soybean: a radical approach. Ann Bot 106: 215–222.
Soil 349: 121–156. https://doi.org/10.1007/s11104-011-0950-4. https://doi.org/10.1093/aob/mcq029.
Rojas M, Lambert F, Ramirez-Villegas J, Challinor AJ. 2019. Wasaya A, Zhang X, Fang Q, Yan Z. 2018. Root phenotyping for
Emergence of robust precipitation changes across crop production drought tolerance: a review. Agronomy 8: 241. https://doi.org/
areas in the 21st century. Proc Natl Acad Sci 116: 6673–6678. 10.3390/agronomy8110241.
https://doi.org/10.1073/pnas.1811463116. Wijewardana C, Alsajri FA, Irby JT, et al. 2019. Water deficit effects
Rötter RP, Tao F, Hohn JG, Palosuo T. 2015. Use of crop simulation on soybean root morphology and early-season vigor. Agronomy 9:
modelling to aid ideotype design of future cereal cultivars. J Exp 836. https://doi.org/10.3390/agronomy9120836.
Bot 66: 3463–3476. https://doi.org/10.1093/jxb/erv098. Xiong R, Liu S, Considine MJ, Siddique KHM, Lam HM, Chen Y. 2020.
Serraj R, Sinclair TR, Allen LH. 1998. Soybean nodulation and N2 Root system architecture, physiological and transcriptional traits of
fixation response to drought under carbon dioxide enrichment. soybean (Glycine max L.) in response to water deficit: a review.
Plant Cell Environ 21: 491–500. https://doi.org/10.1046/j.1365– Physiol Plant 172: 405–418. https://doi.org/10.1111/ppl.13201.
3040.1998.00298.x. Zhao J. 2004. Characterization of root architecture in an applied core
Shao H-B, Chu L-Y, Jaleel CA, Zhao C-X. 2008. Water-deficit stress- collection for phosphorus efficiency of soybean germplasm.
induced anatomical changes in higher plants. C R Biol 331: 215– Chinese Sci Bull 49: 1611–1620. https://doi.org/10.1360/
225. https://doi.org/10.1016/J.CRVI.2008.01.002. 04wc0142.
Sinclair TR, Messina CD, Beatty A, Samples M. 2010. Assessment Zhao J, Fu J, Liao H, et al. 2004. Characterisation of root architecture
across the United States of the benefits of altered soybean drought in an applied core collection for phosphorus efficiency of soybean
traits. Agron J 102: 475–482. https://doi.org/10.2134/ germplasm. Chinese Sci Bull 49: 1611–1620. https://doi.org/
agronj2009.0195. 10.1360/04wc0142.
Specht JE, Hume DJ, Kumudini S V. 1999. Soybean yield potential – Zhu J, Ingram PA, Benfey PN, Elich T. 2011. From lab to field, new
A genetic and physiological perspective. Crop Sci 39: 1560–1570. approaches to phenotyping root system architecture. Curr Opin
https://doi.org/10.2135/cropsci1999.3961560x. Plant Biol 14: 310–317. https://doi.org/10.1016/j.pbi.2011.03.020.
Cite this article as: Dayoub E, Lamichhane JR, Debaeke P, Maury P. 2022. Genotypic differences in root traits to design drought-avoiding
soybean ideotypes. OCL 29: 26.
Page 14 of 14You can also read

















































