Leaf δ15N, δ13C and Their Associations with Soil Fungal Biodiversity, Ectomycorrhizal and Plant Pathogenic Abundance in Forest Ecosystems of China
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Leaf δ15N, δ13C and Their Associations with Soil
Fungal Biodiversity, Ectomycorrhizal and Plant
Pathogenic Abundance in Forest Ecosystems of
China
Wenchen Song ( chenge2412@126.com )
Minzu University of China
Research
Keywords: leaf δ15N, leaf δ13C, fungal diversity, ectomycorrhizal fungi, plant pathogenic fungi, forest
ecosystems
DOI: https://doi.org/10.21203/rs.3.rs-57460/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Read Full License
Page 1/22Abstract
Leaf δ15N and δ13C are important functional traits in biogeographic studies of forest ecosystems.
However, little is known about their relationships with soil fungal biodiversity, ectomycorrhizal, and plant
pathogen abundance at large scales. In this study, leaf and soil samples were collected from 33 forest
reserves along a large range across China to explore the associations between leaf δ15N and δ13C and
soil fungal biodiversity, ectomycorrhizal, and plant pathogen relative abundance using molecular and
stable isotope techniques. We found large-scale biogeographic patterns for leaf δ15N, δ13C, soil fungal
biodiversity, and ectomycorrhizal relative abundance. The soil-plant-microbial interaction may contribute
to the variations in leaf δ15N, δ13C, and soil fungal communities across different types of forest
ecosystems. Temperature and precipitation were the main factors affecting large-scale biogeographic
patterns of latitude and longitude. Leaf δ15N was mainly affected by the relative abundance of
ectomycorrhizal fungi and leaf δ13C was affected by the relative abundance of plant pathogens. Leaf
δ15N and δ13C may be indicators re ecting soil fungal communities in forest ecosystems.
Introduction
Soil microbes can exhibit biogeographic patterns in species diversity and distribution (Hanson et al.
2012), and soil fungi are crucial components of microbial communities in forest ecosystems, where they
play fundamental ecological roles in soil formation, conservation, and regulating nutrient cycling
(Zeilinger et al. 2015). Recent studies have reported distinct distribution patterns for different fungal taxa
like ectomycorrhizal fungi (EMF) and arbuscular mycorrhizal fungi (AMF) across latitudes (Tedersoo et
al. 2012; Davison et al. 2015). However, the mechanisms underlying these biogeographic patterns are
di cult to distinguish in forest ecosystems because multiple community assembly processes may
govern the biogeographic variation in soil fungal diversity (Ricklefs et al. 2004; Kraft et al. 2011; Bahram
et al. 2018). In addition, biotic interactions within fungal communities may vary along large-scale
geographic gradients, resulting in different strengths in the assembly of mechanisms across forest
ecosystems (Hu et al. 2019). Yet, to date, few studies have focused on how these complex geographic
processes together in soil fungal diversity in different forest types of China (Huang et al. 2019).
Therefore, a more comprehensive perspective is required to understand how and why their relative
in uences vary across forest types (Powell et al. 2015).
How plant pathogens and EMF affect the biodiversity and construction of forests along large-scale
gradients remains underreported. Recent studies suggest that plant pathogens play an important role in
promoting plant species coexistence through negative plant-soil feedback at local scales in forests, which
subsequently affects the biodiversity and construction of forest ecosystems (Bever et al. 2015; Chen et al.
2019). Furthermore, plant pathogens interact with other fungal taxa such as EMF who can assist plants
to defend against pathogens (Sikes et al. 2009; Laliberté et al. 2015; Bennett et al. 2017). Additionally,
plant nutrient acquisition strategies and resistance to soil-borne pathogens strongly depend on
mycorrhizal associations (Tedersoo et al. 2020). Local plant diversity may be in uenced by varied plant
Page 2/22responses to soil fungi through the following four mechanisms: access to organic nutrients, positive
plant-soil feedback, accumulation of organic material, and allelopathic compounds (Bennett et al. 2017;
Teste et al. 2017; Tedersoo et al. 2020). However, most studies on plant-soil-fungal interactions have
focused on the variations in plant performance using potting experiments with the application of
fungicide or inoculation with several fungal isolates (e.g., plant pathogens and EMF), which may overlook
the relationships between different fungal taxa in natural conditions (Wang et al. 2019). Thus, a
comprehensive study linking the spatial distributions of fungal biodiversity and functional taxa, e.g., EMF
and plant pathogens, to plant functional indicators across large scales (i.e., across China) is urgently
needed.
As important indicators for studying plant ecophysiology, stable isotopes provide fundamental insights
into how plants interact with and respond to environmental factors, improving our understanding of how
the plants adapt to their environment (Dawson et al. 2002). Leaf δ15N and δ13C were found more closely
associated with belowground communities than soil isotopic values, where they could be used to predict
the properties of belowground communities (Wang et al. 2017). In particular, leaf δ13C re ects the
balance between leaf conductance and photosynthetic rate, which can used to calculate intrinsic water
use e ciency (iWUE) created by Farquhar et al. (1989). Based on this, leaf δ13C is widely used to
measure long-term water use e ciency under different environmental conditions and reveal signi cant
functional changes in plant metabolism and adaptation to various environmental stresses (Cernusak et
al. 2013; Gautam and Lee 2016; Aguilar-Romero et al. 2017; Acosta-Rangel et al. 2018). Compared with
leaf δ13C, the factors affecting leaf δ15N are more complex. The variability in leaf δ15N observed in nature
might originate from various subtle changes in metabolic uxes or environment-driven effects, such as
multiple soil N sources with distinct isotopic values, mycorrhizal associations, temporal and spatial
variation in N availability, and human disturbance (Sah et al. 2006; Tcherkez, 2011; Hobbie and Hogberg
2012; Branoff 2017). Nevertheless, most studies believe that leaf δ15N is mainly in uenced by soil
processes, especially the N dynamics between rhizomicrobes and plants in forest ecosystems (Morford et
al. 2011; Hobbie and Hogberg 2012). EMF can supply relatively 15N-enriched N to their hosts in
rhizosphere processes (Song et al. 2020), and transfer 15N-enriched N from root to leaf (Takizawa et al.
2017), thus establishing a link between leaf δ15N and soil processes. However, although the variations in
EMF δ15N have been reported across large scales (Mayor et al. 2015), studies on the relationship between
leaf δ15N and soil fungi are scarce.
Soil fungi are crucial components of microbial communities in forest ecosystems, and play fundamental
roles in large-scale plant-soil interactions (Huang et al. 2019; Wang et al. 2019). However, we still lack the
answers to the following questions: how do soil fungal biodiversity, ectomycorrhizal, and plant pathogen
abundance vary in forest ecosystems across China? What affects these large-scale variations? Can leaf
δ15N and δ13C be used as indicators to re ect soil fungal communities in forest ecosystems? To answer
these questions, we collected leaf and soil samples from 33 forest reserves along a large range across
China to explore the associations between soil fungal communities and leaf C, N stable isotopes using
molecular and stable isotope techniques.
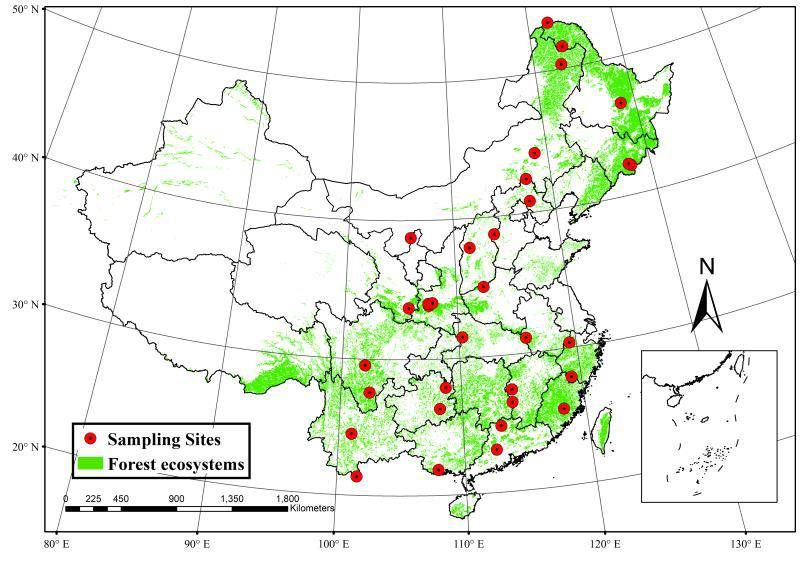
Page 3/22Methods Soil and leaf sampling We surveyed 33 mountain forest reserves located across a broad range of latitudes (21.40◦∼53.56 ºN) and longitudes (101.03º∼128.52 ºE) in China (Figure 1 and Table S1). In each forest reserve, 9–15 sampling plots were randomly chosen along the same aspect of the mountain. Five topsoil samples (within a 5 cm depth) were collected from each plot (5 m apart) and stored in icy sterilized bags. Soils were stored at -20 ºC for molecular experiments. The leaves of the dominant trees species (such as Pinaceae,Betulaceae, and Fagaceae) in each plot were sampled at the same time. Each healthy adult trees was chosen 10 leaves were sampled from each tree was sampled 10 leaves. Soil and leaf analysis Brie y, the DNA of the soil samples was extracted using the MoBio PowerSoil DNA extraction kit (Carlsbad, CA) (see Wang et al. [2019] for more detail). The second nuclear ribosomal internal transcribed spacer (ITS2) region of the rRNA operon was ampli ed to determine fungal community composition using the primer set ITS3 (5’- GCATCGATGAAGAACGCAGC-3’) and ITS4 (5’- TCCTCCGCTTATTGATATGC- 3’). Polymerase chain reaction (PCR) conditions were as follows: preheat 30 s at 98 ºC, 35 cycles 10 s at 98 ºC, annealing 25 s at 52 ºC, extension 30 s at 72 ºC, nal extension 8 min at 72 ºC. PCR products from the different samples were pooled together in equimolar concentrations and puri ed using a E.Z.N.A. Gel Extraction Kit (Omega BioTek, Doraville, USA). The pooled PCR products were sequenced on a 2 × 300 paired-end MiSeq NextGen platform (Illumina; San Diego, CA, USA). Low-quality bases with Phred quality scores
presented as mean ± SE. The distribution map of forest reserves was drawn using ArcGIS 10.2 (ESRI Inc.,
CA, USA). Statistical analyses were performed using IBM SPSS Statistics 23.0 (IBM Inc., NY, USA).
Results
Biogeographic variations
About 420 EMF OTUs and only 15 AMF OTUs were found in the dataset, suggesting that the forests in
China are mainly ectomycorrhizal systems. Soil fungal diversity (Shannon index) was found to
signi cantly and parabolically vary with latitude and peaked at approximately 40 ºN, which is the
boundary between the North and Northeast zones of China (Figure 2A). There was no signi cant
relationship between the relative abundance of plant pathogens and biogeographic factors. However, the
relative abundance of ectomycorrhizas increased signi cantly with latitude, except for Changbai
Mountain, which showed a considerably higher relative abundance of ectomycorrhizas than the other
forest reserves (Figure 2B). Leaf δ15N increased signi cantly with latitude (Figure 2C). The relationship
between leaf δ13C and latitude was different for the South (tropical and subtropical) and North
(temperate and boreal) zones of China (the boundary is at located at approximately 35 ºN). Leaf δ13C
decreased and then increased signi cantly with latitude, and the lowest point was at approximately 30 ºN
in the South, while leaf δ13C decreased signi cantly in North China (Figure 2D).
There was no signi cant relationship between the fungal Shannon index and longitude in the South, but it
decreased signi cantly in North China with increasing longitude (Figure 3A). The relative abundance of
ectomycorrhizas was decreased and then increased related to longitude signi cantly, and the lowest
point was at approximately 110 ºE, which is the boundary between the West and East zones of China
(Figure 3B). There was no signi cant relationship between leaf δ15N and longitude (Figure 3C). Leaf δ13C
increased signi cantly with longitude in South China, while there was no signi cant relationship between
Leaf δ13C and longitude in North China (Figure 3D).
The fungal Shannon index was signi cantly and unimodally related to mean annual temperature (MAT),
and peaked at about 7 oC (Figure 4A). The relative abundance of ectomycorrhizas decreased signi cantly
with MAT, except for Changbai Mountain, which had a considerably greater relative abundance of
ectomycorrhizas than the other forest reserves (Figure 4B). Leaf δ15N decreased signi cantly as MAT
increased (Figure 4C). Leaf δ13C was decreased and then increased related to MAT signi cantly, and the
lowest point was at approximately 17.5 oC in South China. Leaf δ13C increased signi cantly as MAT
increased in North China (Figure 4D).
The fungal Shannon index decreased signi cantly as mean annual precipitation (MAP) increased (Figure
5A). The relative abundance of ectomycorrhizas was decreased and then increased related to MAP
signi cantly, and the lowest point was at approximately 1340 mm, except for Changbai Mountain, which
had a considerably greater relative abundance of ectomycorrhizas than the other forest reserves (Figure
5B). Leaf δ15N and δ13C decreased signi cantly as MAP increased (Figures 5C, D).
Page 5/22Relationships betweenleaf δ15N, δ13C and soil fungal community
Leaf δ15N was signi cantly positively correlated with the soil fungal Shannon index (Figure 6A).
Moreover, leaf δ15N was signi cantly negatively correlated with the relative abundance of plant
pathogens, but signi cantly positively correlated with the relative abundance of ectomycorrhizas (Figures
6B, 6C). There was no signi cant relationship between the fungal Shannon index and leaf δ13C (Figure
7A). Leaf δ13C was signi cantly and unimodally correlated with the relative abundance of plant
pathogens and valleyed at approximately 8.5% (Figure 7B). Leaf δ13C was signi cantly positively
correlated with the relative abundance of ectomycorrhizas (Figure 7C). Ectomycorrhizal relative
abundance was signi cantly negatively correlated with plant pathogen relative abundance (Figure S1).
Discussion
Biogeographic variations in soil fungal communities
Climate factors are the main factors affecting fungal biodiversity compared with other abiotic and biotic
factors (He et al. 2017; Thiergart et al. 2020; Fiona et al. 2020). China's temperature gradually rises from
North to South, thus causing similar parabolic variations in soil fungal diversity (Shannon index) across
latitude and MAT (Wang et al. 2019; Hu et al. 2019). The unimodal trend with fungal diversity peaking at
approximately 40 oN in the present study was similar to that found in other global patterns (Tedersoo et
al. 2014; Bahram et al. 2018). This peak in fungal diversity, which was at approximately 40 oN and at a
MAT of 7 oC, was similar to that found in EMF diversity in the temperate biome. This indicated that the
relationship between fungal biodiversity and latitude was mainly caused by the variation in EMF
biodiversity, which was mostly affected by temperature and precipitation (Tedersoo et al. 2012; 2014).
The relative abundance of ectomycorrhizas increased signi cantly from the warm south to the cold north,
which suggests that trees in colder areas are more dependent on ectomycorrhizal fungi (Miyamoto et al.
2018). High ectomycorrhizal abundance may enhance nutrient trading (Wyatt et al. 2014), improve
overall nutrient acquisition and the function of common mycorrhizal networks (Tedersoo et al. 2020), and
subsequently increase fungal biodiversity (Tedersoo, 2015; Hiiesalu et al. 2014). However, in the present
study, the adverse effects of EMF systems on fungal diversity emerged in Southeast China, north of 40
ºN. EMF can reduce mycorrhizal root colonization of neighboring AMF and enhance the competitive
dominance of EMF over AMF by promoting litter accumulation and limiting access to nutrients (Booth et
al. 2010; Becklin et al. 2012). In addition, EMF can suppress AMF colonization via allelopathic litter or
nutrient competition (Javaid, 2007; Becklin et al. 2012). Thus, fungal diversity decreases with an increase
in latitude in the forests of Southeast China, where EMF dominates soil-fungal systems (Gerz et al. 2016;
Tedersoo et al. 2020).
Precipitation in China gradually decreases from Southeast to Northwest. In the wet south, moisture is not
the main limiting factor; hence, we found no relationship between the fungal Shannon index and
longitude. However, precipitation is an important factor in the dry north. High soil moisture can inhibit
Page 6/22rhizomicrobial activity (Song et al. 2018), high fungal diversity can help plants to resist the adverse
effects of drought (Fahey et al. 2020), thus causing the fungal Shannon index to decrease signi cantly
as MAP increased (Preece et al. 2019). Furthermore, the fungal Shannon index decreased signi cantly
with longitude from the dry west to the wet east in North China in the present study. Similarly, the relative
abundance of ectomycorrhizas decreased as longitude increased, which may be due to the lack of soil
moisture for microbial activity and biodiversity in low MAP zones (Song et al. 2018; Preece et al. 2019;
Fahey et al. 2020). As the soil becomes more waterlogged in the east, trees rely more on mycorrhizal
fungi to protect their roots against the adverse effects of excessive soil moisture (Erlandson et al. 2016),
which explains why the relative abundance of EMF increased as longitude increased in high MAP zones
in the present study. For these reasons, the relative abundance of EMF showed a unimodal variation from
the dry west to the wet east of China.
It should be noted that the relative abundance of EMF samples from Changbai Mountain was much
higher than that from other forest reserves, which is probably due to the special geological history of
Changbai Mountain. Changbai Mountain is a dormant volcano; the last eruption was in 1702 (Yuan and
Sun 1990). After the eruption, substantial amounts of nutrients were left in the soil, which caused the
mycorrhizal fungi to proliferate. Furthermore, the combination of positive density dependence and
advantages of nutrition utilization may have led to a clustering of EMF seedlings around adult trees and
a lack of tree saplings with AMF around conspeci c adult AMF trees (Johnson et al. 2017; Sasaki et al.
2019). By promoting litter accumulation, EMF can limit nutrient acquisition to their own advantage (Booth
et al. 2010; Becklin et al. 2012; Yang et al. 2018). Over time, the aggregation of EMF can lead to
monodominance, resulting in an extremely high relative abundance of EMF on Changbai Mountain (Peh
et al. 2011).
Biogeographic variations in leaf δ15N and δ13C
Leaf δ15N increased signi cantly with latitude, which corresponded to the signi cant decrease in leaf
δ15N with increasing MAT. Similar relationships were found with the relative abundance of EMF, whcih
indicates that the ecogeographical pattern of leaf δ15N is mainly related to the mycorrhizal symbiotic
system. EMF can supply relatively 15N-enriched N to their hosts in the rhizosphere causing plant δ15N to
increase with EMF relative abundance (He et al. 2009; Angst et al. 2018; Song et al. 2020). Noticeably,
that leaf δ15N varied with latitude in the present study was similar to global patterns at 20º∼50 ºN
(Mayor et al. 2015), but this differed from Kang’s et al. (2011) study on a single species. This may
indicate that the changes in leaf δ15N with latitude are accompanied by adaptive changes in vegetation
rather than the adaptive changes of a single species. High moisture levels leads to a decrease in the
rhizomicrobial activity and the ability of mycorrhizal symbionts to obtain nutrition from decomposing soil
organic matter, thus causing leaf δ15N to decrease with MAP (Amundson et al. 2003; Song et al. 2017;
2018).
Page 7/22Leaf δ13C re ects the response and adaptation mechanisms of plants to speci c environments, and may
be used as an alternative indicator of the long-term water use e ciency of plants (Cernusak et al. 2013;
Gautam and Lee 2016; Acosta-Rangel et al. 2018). In the present study, leaf δ13C decreased signi cantly
with latitude, except at 30º∼37 ºN. This decreasing trend is similar to most studies, which is explained as
the phenotypic acclimation of plants to climate (Diefendorf et al. 2010; Sun et al. 2016; Li et al. 2017).
The decreasing trend may due to the special ability of plants to adapt to cool and dry environments that
occur from 30º to 37 ºN (Du et al. 2015). MAP is the strongest predictor of leaf δ13C among global
climate variables (Diefendorf et al. 2010). Water de cits might reduce either stomatal conductance or
stomatal density, leading to improved water use e ciency and positive leaf δ13C in plants (Cernusak et
al. 2013; Du et al. 2015; Li et al. 2017). This may explain the decrease in leaf δ13C from the dry west to
wet east in South China in the present study. Compared with the humid south, plants in the arid north
tend to reduce stomatal conductance, and have a relatively high photosynthetic rate for adapting to the
short growth season at high latitudes (Soolanayakanahally et al. 2009; Du et al. 2015; Li et al. 2017). This
may explain the higher δ13C found in the leaves from North China compared to South China in the
present study.
Associations between leaf δ15N, δ13C and soil fungal community
Rhizomicrobes, such as the common mycelial network of EMF, can supply relatively 15N-enriched N to
their hosts in the rhizosphere (He et al. 2009; Angst et al. 2018; Song et al. 2020), which explains why
plant δ15N increased as the relative abundance of EMF increased in the present study. EMF can inhibit
plant pathogens and protect plant root systems relatively more effectively than other mycorrhizal guilds
(Bennett et al. 2017; Teste et al. 2017; Kadowaki et al. 2018). Therefore, in our study, the negatively
correlation between leaf δ15N and plant pathogen relative abundance may be explained by the higher
relative abundance of EMF, which enables the plant-microbial system to be more e cient at transferring
15
N-enriched N (Kranabetter et al. 2015). Further, as the transfer e ciency of the root-microbial system is
enhanced, niche availability increases, thereby increasing biodiversity in the belowground ecosystem
(Bardgett and van der Putten 2014). This may explain why leaf δ15N was signi cantly positively
correlated with soil fungal Shannon index in the present study.
Leaf δ13C mainly re ects the long-term water use e ciency of plants (Cernusak et al. 2013; Gautam and
Lee 2016; Acosta-Rangel et al. 2018). EMF are bene cial to plants as they assist the plants with
environmental adaptation and water use e ciency (Hobbie and Colpaert 2004; Kranabetter et al. 2015;
Uroz et al. 2016), therefore leaf δ13C was signi cantly positively correlated with the relative abundance of
EMF. Similarly, plant pathogens are harmful to plants; hence, leaf δ13C decreased as the relative
abundance of plant pathogens increased. Notably, leaf δ13C increased with the relative abundance of
plant pathogens when both the relative abundance of plant pathogens and EMF were more than 10%.
This is probably due to enhanced adaptability and immunity of the plants and the ability of EMF to
protect the plants against pathogens, which may allow the ecosystem to bear more plant pathogens.
Page 8/22Conclusions
In summary, we reported large-scale biogeographic patterns in soil fungal diversity, ectomycorrhizal, and
plant pathogen abundance, and subsequently linked their diversity distributions to large-scale leaf δ15N
and δ13C patterns. The large-scale biogeographic patterns of EMF relative abundance suggest that the
soil-plant-microbial interactions contribute to the variations in soil fungal diversity, leaf δ15N, and δ13C
across different forest ecosystems in China. Climatic factors, i.e., MAT and MAP, may be affecting these
large-scale biogeographic variations. Leaf δ15N was mainly affected by the relative abundance of EMF
while leaf δ13C was mainly affected by the relative abundance of plant pathogens. Leaf δ15N and δ13C
may be indicators for re ecting the soil fungal communities in forest ecosystems; thus, they could be
used to predict the component and function of belowground fungal communities.
Declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Representative fungal OTU sequences have been published in Wang et al. (2019) (doi:
10.3389/fevo.2019.00216) and the European Nucleotide Archive (ENA) with NO. LT986405–LT998319.
Con icts of Interest
The authors declare no con icts of interest.
Funding
This study was sponsored by the National Key Research and Development Plan (2017YFC0504004).
Author Contributions
W. Song developed the ideas and designed the experimental plans, performed the experiments, analyzed
the data and wrote the manuscript.
Acknowledgements
We thank Dr. Gao Jie help collected data.
Page 9/22References
1. Acosta-Rangel, A., Avila-Lovera, E., Guzman, M.E.D., Torres, L., Haro, R., Arpaia, M.L., Focht, E.,
Santiago, L.S. (2018). Evaluation of leaf carbon isotopes and functional traits in avocado reveals
water-use e cient cultivars. Ecosyst. Environ. 263, 60–66. doi: 10.1016/j.agee.2018.04.021.
2. Aguilar-Romero, R.; Pineda-Garcia, F.; Paz, H.; Gonzalez-Rodriguez, A.; Oyama, K. (2017).
Differentiation in the water-use strategies among oak species from central Mexico. Tree Physiol.37,
915–925. doi: 10.1093/treephys/tpx033
3. Amundson, R., Austin, A. T., Schuur, E. A., Yoo, K., Matzek, V., Kendall, C., Uebersax, A., Brenner, D. L.,
Baisden, W. T. (2003). Global patterns of the isotopic composition of soil and plant nitrogen. Global
Biogeochem. Cy.17(1), 1031–1038. doi: 10.1029/2002gb001903
4. Angst, G., Mueller, K.E., Eissensta, D.M., Trumbore, S., Freeman, K.H., et al. (2018) Soil organic carbon
stability in forests: distinct effects of tree species identity and traits. Global Change Biol.25, 1529-
1546. doi: 10.1111/gcb.14548
5. Bahram, M., Hildebrand, F., Forslund, S. K., Anderson, J. L., Soudzilovskaia, N. A., Bodegom, P. M.,
Bengtsson-Palme, J., Anslan, S., Coelho, L.P., Harend, H. (2018). Structure and function of the global
topsoil microbiome. Nature560, 233–237. doi: 10.1038/s41586-018-0386-6
6. Bardgett, R. D., van der Putten, W. H. (2014). Belowground biodiversity and ecosystem functioning.
Nature515(7528), 505–511. doi:10.1038/nature13855
7. Becklin M., Pallo M. L., Galen C. (2012) Willows indirectly reduce arbuscular mycorrhizal fungal
colonization in understorey communities. J. Ecol.100, 343–351 . doi: 10.1111/.1365-
2745.2011.01903.x
8. Bennett, J. A., Maherali, H., Reinhart, K. O., Lekberg, Y., Hart, M. M., Klironomos, J. (2017). Plant-soil
feedbacks and mycorrhizal type in uence temperate forest population dynamics. Science,
355(6321), 181-184. doi: 1126/science.aai8212.
9. Bever, J. D., Mangan, S. A., Alexander, H. M. (2015). Maintenance of plant species diversity by
pathogens. Rev. Ecol. Evol. Syst.46, 305–325. doi: 10.1146/annurev-ecolsys-112414-054306
10. Booth, M. G. , Hoeksema, J. D. (2010). Mycorrhizal networks counteract competitive effects of
canopy trees on seedling survival. Ecology91, 2294–2302. doi: 10.1890/09-1139.1
11. Branoff, B. L. (2017). Quantifying the in uence of urban land use on mangrove biology and ecology:
a meta-analysis. Global Ecol. Biogeogr.26, 1339-1356. doi: 1111/geb.12638
12. Cernusak, L. A. , Ubierna, N. , Winter, K. , Holtum, J. A. M. , Marshall, J. D. , Farquhar, G. D. (2013).
Environmental and physiological determinants of carbon isotope discrimination in terrestrial
plants. New Phytol.,200(4), 950-965. doi: 1111/nph.12423
13. Chen L., Swenson N.G., Ji N.N., Mi X.C., Ren H.B., Guo L.D., Ma K.P. (2019). Differential soil fungus
accumulation and density dependence of trees in a subtropical forest. Science333, 1755–1758. doi:
1126/science.aau1361
Page 10/2214. Davison, J., Moora, M., Öpik, M., Adholeya, A., Ainsaar, L., Bâ, A., Burla, S., Diedhiou, A. G., Hiiesalu, I.,
Jairus, T., Johnson, N. C., Kane, A., Koorem, K., Kochar, M., Ndiaye, C., Partel, M., Reier, U., Saks, U.,
Singh, R., Vasar, M., Zobel, M. (2015). Global assessment of arbuscular mycorrhizal fungus diversity
reveals very low endemism. Science 349, 970–973. doi: 10.1126/science.aab1161
15. Dawson, T.E., Mambelli, S., Plamboeck, A.H., Templer, P.H., Tu, K.P. (2002). Stable isotopes in plant
ecology. Rev. Ecol. Syst.33, 507–559. doi: 10.1146/annurev.ecolsys.33.020602.095451
16. Diefendorf, A. F., Mueller, K. E., Wing, S. L., Koch, P. L., Freeman, K. H. (2010). Global patterns in leaf
13
C discrimination and implications for studies of past and future climate. P Natl. Acad. Sci.
USA107(13), 5738-5743. doi: 10.1073/pnas.0910513107
17. Du, B., Kang, H., Zhu, Y., Zhou, X., Yin, S., Burgess, P. J., Liu, C. (2015). Variation of Oriental Oak
(Quercus variabilis) Leaf δ13C across Temperate and Subtropical China: Spatial Patterns and
Sensitivity to Precipitation. Forests6(7), 2296-2306. doi: 10.3390/f6072296
18. Erlandson, S., Savage, J. A., Cavenderbares, J., Peay, K. G. (2016). Soil moisture and chemistry
in uence diversity of ectomycorrhizal fungal communities associating with willow along an
hydrologic gradient. FEMS Microbiol. Ecol.92(1), v148. doi: 10.1093/femsec/ v148
19. Fahey, C., Koyama, A., Antunes, P. M., Dun eld, K. E., & Flory, S. L. (2020). Plant communities mediate
the interactive effects of invasion and drought on soil microbial communities. ISME J. 1-14. doi:
10.1038/s41396-020-0614-6
20. Farquhar GD, Ehleringer JR, Hubick KT. (1989). Carbon isotope discrimination and photosynthesis.
Rev. Plant Physiol. Plant Mol. Bio.40, 503–537. doi: 1040-251 9/89/0601 -503.
21. Fiona, M. S., Paul, B.L. G., Inma L., Davey, L. J., Simon C., David A. R. (2020). Soil textural
heterogeneity impacts bacterial but not fungal diversity. Soil Biol. Biochem.144, 107766. doi:
1016/j.soilbio.2020.107766
22. Gautam, M.K., Lee, K.S. (2016). Application of stable isotopes in ecosystem research. Sci.110, 1288–
1306. doi: 10.18520/cs/v110/i7/1288-1306.
23. Gerz, M. , Bueno, C. G. , Zobel, M. , Moora, M. , Pugnaire, F. (2016). Plant community mycorrhization in
temperate forests and grasslands: relations with edaphic properties and plant diversity. Veg. Sci.27,
89–99. doi: 10.1111/jvs.12338
24. Hanson, C. A., Fuhrman, J. A., Horner-Devine, M. C., Martiny, J. B. H. (2012). Beyond biogeographic
patterns: processes shaping the microbial landscape. Rev. Genet.,10(7), 845. doi:
10.1038/nrmicro2795
25. He, J., Tedersoo, L., Hu, A. , Han, C. , He, D. , et al. (2017). Greater diversity of soil fungal communities
and distinguishable seasonal variation in temperate deciduous forests compared with subtropical
evergreen forests of eastern china. FEMS Microbiol. Ecol.93, x069. doi: 10.1093/femsec/ x069
26. He, X., Xu, M., Qiu, G. Y., Zhou, J. (2009). Use of 15N stable isotope to quantify nitrogen transfer
between mycorrhizal plants. Plant Ecol.2(3), 107-118. doi: 10.1093/jpe/rtp015
Page 11/2227. Hiiesalu, I. , Meelis Pärtel, Davison, J. , Gerhold, P. , Metsis, M. , Moora, M. , et al. (2014). Species
richness of arbuscular mycorrhizal fungi: associations with grassland plant richness and biomass.
New Phytol.203, 233–244. doi: 10.1111/nph.12765.
28. Hobbie, E. A., Colpaert J. V. (2004). Nitrogen availability and mycorrhizal colonization in uence water
use e ciency and carbon isotope patterns in pinus sylvestris. New Phytol.164(3), 515-525. doi:
10.1111/j.1469-8137.2004.01187.x
29. Hobbie, E. A., Hogberg, P. (2012). Nitrogen isotopes link mycorrhizal fungi and plants to nitrogen
dynamics. New Phytol.196(2), 367-382. doi: 1111/j.1469-8137.2012.04300.x
30. Hu, Y., Veresoglou, S.D., Tedersoo, L., Xu, T., Ge, T., Liu, L., Chen, Y., Hao, Z., Su, Y., Rillig, M.C., Chen,
B.D. (2019). Contrasting latitudinal diversity and co-occurrence patterns of soil fungi and plants in
forest ecosystems. Soil Biol. Biochem.131, 100–110. doi: 1016/j.soilbio.2019.01.001
31. Huang, Y., Zhang, X., Fu, S., Zhang, W. (2019). Environmental ltering drives local soil fungal beta
diversity more than dispersal limitation in six forest types along a latitudinal gradient in Eastern
China. Forests10, 863. doi: 10.3390/f10100863
32. Javaid, A. (2007). Allelopathic interactions in mycorrhizal associations. Allelopathy J.20(1), 29-42.
doi: 10.2134/agronj2007.0122L
33. Johnson, D. J. , Clay, K. , Phillips, R. P. (2017). Mycorrhizal associations and the spatial structure of
an old-growth forest community. Oecologia186, 195–204. doi: 10.1007/s00442-017-3987-0
34. Kadowaki, K. , Yamamoto, S. , Sato, H. , Tanabe, A. S. , Hidaka, A. , Toju, H. (2018). Mycorrhizal fungi
mediate the direction and strength of plant–soil feedbacks differently between arbuscular
mycorrhizal and ectomycorrhizal communities. Biol.1, 196. doi: 10.1038/s42003-018-0201-9.
35. Kang, H., Liu, C., Yu, W., Wu, L., Chen, D., Sun, X., Ma, X., Hu, H., Zhu, X. (2011). Variation in foliar δ15N
among oriental oak (Quercus variabilis) stands over eastern China: Patterns and interactions.
Geochem. Explor.110(1), 8-14. doi: 10.1016/j.gexplo.2011.02.002
36. Kraft, N.J.B., Comita, L.S., Chase, J.M., Sanders, N.J., Swenson, N.G., Crist, T.O., Stegen, J.C., Vellend,
M., Boyle, B., Anderson, M.J. Cornell, H.V., Davies, K.F., Freestone, A.L., Inouye, B.D., Harrison, S.P.,
Myers, J.A. (2011). Disentangling the drivers of diversity along latitudinal and elevational gradients.
Science333, 1755–1758. doi: 1126/science.1208584
37. Kranabetter, J. M. , Stoehr, M. , O"Neill, G. A. (2015). Ectomycorrhizal fungal maladaptation and
growth reductions associated with assisted migration of douglas- r. New Phytol.206(3), 1135-1144.
doi: 10.1111/nph.13287
38. Laliberté, E., Lambers, H., Burgess, T. I., Wright, S. J. (2015). Phosphorus limitation, soil-borne
pathogens and the coexistence of plant species in hyperdiverse forests and shrublands. New Phytol.
206, 507–521. doi: 10.1111/nph.13203
39. Li, M., Peng, C., Wang, M., Yang, Y., Zhang, K., Li, P., Yang, Y., Ni, J., Zhu, Q. (2017). Spatial patterns of
leaf δ13C and its relationship with plant functional groups and environmental factors in China.
Geophys. Res. Biogeosci.122(7), 1564-1575. doi:10.1002/2016JG003529
Page 12/2240. Mayor, J. , Bahram, M. , Henkel, T. , Buegger, F. , Pritsch, K. , Tedersoo, L. (2015). Ectomycorrhizal
impacts on plant nitrogen nutrition: emerging isotopic patterns, latitudinal variation and hidden
mechanisms. Lett.18(1), 96-107. doi: 10.1111/ele.12377
41. Miyamoto, Y., Terashima, Y., Nara, K. (2018). Temperature niche position and breadth of
ectomycorrhizal fungi: reduced diversity under warming predicted by a nested community structure.
Global Change Biol.24, 5724-5737 doi:10.1111/gcb.14446
42. Morford, S. L. , Houlton, B. Z. , Dahlgren, R. A. (2011). Increased forest ecosystem carbon and
nitrogen storage from nitrogen rich bedrock. Nature477(7362), 78-81. doi: 10.1038/nature10415
43. Nguyen, N. H., Song, Z. W., Bates, S. T., Branco, S., Tedersoo, L., Schilling, M. J., Kennedy P. G. (2016).
FUNGuild: an open annotation tool for parsing fungal community datasets by ecological guild.
Fungal Ecol.20, 241–248. doi: 10.1016/j.funeco.2015.06.006
44. Peh, K. S., Lewis, S. L., Lloyd, J. (2011) Mechanisms of monodominance in diverse tropical tree‐
dominated systems. Ecol.99, 891–898. doi: 10.1111/j.1365-2745.2011.01827.x
45. Powell, J.R., Karunaratne, S., Campbell, C.D., Yao, H., Robinson, L., Singh, B.K. (2015). Deterministic
processes vary during community assembly for ecologically dissimilar taxa. Commun.6, 8444. doi:
10.1038/ncomms9444
46. Preece, C. , Verbruggen, E. , Liu, L. , Weedon, J. T. , Peñuelas, Josep. (2019). Effects of past and
current drought on the composition and diversity of soil microbial communities. Soil Biol. Biochem.4,
122–131. doi: 10.1016/j.soilbio.2018.12.022.
47. Ricklefs, R. E. A. (2004). comprehensive framework for global patterns in biodiversity. Lett.7, 1–15.
doi: 10.1046/j.1461-0248.2003.00554.x
48. Sah, S.P., Rita, H., Ilvesniemi, H. (2006). ¹ N natural abundance of foliage and soil across boreal
forests of Finland. Biogeochemistry80, 277–288. doi: 2307/20456403
49. Sasaki, T. , Konno, M. , Hasegawa, Y. , Imaji, A. , Terabaru, M. , Nakamura, R. , Ohira, N. , Matsukura, K.
, Seiwa, K. (2019). Role of mycorrhizal associations in tree spatial distribution patterns based on size
class in an old-growth forest. Oecologia189, 971–980. doi: 10.1007/s00442-019-04376-2
50. Sikes, B. A., Cottenie, K., Klironomos, J. N. (2009). Plant and fungal identitydetermines pathogen
protection of plant roots by arbuscular mycorrhizas. Ecol. 97, 1274–1280. doi: 10.1111/j.1365-
2745.2009.01557.x
51. Song, W., Tong, X., Liu, Y., Li, W. (2020). Microbial community, newly sequestered soil organic carbon,
and δ15N variations driven by tree roots, Microbiol.11, 314. doi: 10.3389/fmicb.2020.00314
52. Song, W., Tong, X., Zhang, J., Meng, P., Li, J. (2018) How a root-microbial system regulates the
response of soil respiration to temperature and moisture in a plantation. J. Environ. Stud.27, 2749–
2756. doi: 10.15244/pjoes/81271
53. Song, W.C., Tong, X.J., Zhang, J.S., Meng, P., Li, J. (2017). Autotrophic and heterotrophic components
of soil respiration caused by rhizosphere priming effects in a plantation. Plant Soil Environ.63, 295–
299.
Page 13/2254. Soolanayakanahally, R. Y. , Guy, R. D. , Silim, S. N. , Drewes, E. C. , Schroeder, W. R. (2009). Enhanced
assimilation rate and water use e ciency with latitude through increased photosynthetic capacity
and internal conductance in balsam poplar (populus balsamifera). Plant Cell Environ. 32(12), 1821-
1832. doi: 10.1111/j.1365-3040.2009.02042.x
55. Sun, X. , Kang, H. , Chen, H. Y. H. , Du, B. , Yin, S. , Zhou, X. , Searle E. B., Liu, C. (2016). Phenotypic
plasticity controls regional-scale variation inquercus variabilisleaf δ13Trees30(4), 1445-1453. doi:
10.1007/s00468-016-1380-y
56. Takizawa, Y. , Dharampal, P. S. , Steffan, S. A. , Takano, Y. , Ohkouchi, N. , Chikaraishi, Y. (2017). Intra-
trophic isotopic discrimination of 15N/14N for amino acids in autotrophs: implications for nitrogen
dynamics in ecological studies. Evol.7(9), 2916-2924. doi: 10.1002/ece3.2866
57. Tcherkez, G. (2011). Natural 15N/14N isotope composition in C3 leaves: are enzymatic isotope effects
informative for predicting the 15N-abundance in key metabolites? Plant Biol.38, 1–12. doi:
10.1071/FP10091
58. Tedersoo, L. (2015). Tree diversity and species identity effects on soil fungi, protists and animals are
context dependent. ISME J.10, 346–362. doi: 10.1038/ismej.2015.116.
59. Tedersoo, L., Bahram, M., Põlme, S., Kõljalg, U., Yorou, N. S., Wijesundera, R., et al. (2014). Global
diversity and geography of soil fungi. Science346, doi: 10.1126/science.1256688
60. Tedersoo, L., Bahram, M., Toots, M., DiÉDhiou, A. G., Henkel, T. W., KjØLler, R., Morris, M. H., Nara, K.,
Nouhra, E., Peay, K. G., Polme, S., Ryberg, M., Smith, M. E., Koljalg, U. (2012). Towards global patterns
in the diversity and community structure of ectomycorrhizal fungi. Ecol.21, 4160–4170. doi:
10.1111/j.1365-294X.2012.05602.x
61. Tedersoo, L., Bahram, M., Zobel, M. (2020). Plant-soil feedbacks and mycorrhizal type in uence
temperate forest population dynamics. Science, 367(6480), eaba1223. doi: 1126/science.aba1223.
62. Teste, F. P., Kardol, P., Turner, B. L.,Wardle, D. A., Zemunik, G., Renton,M., et al. (2017). Plant-soil
feedback and the maintenance of diversity in Mediterranean climate shrublands. Science355, 173–
176. doi: 10.1126/science.aai8291
63. Thiergart, T., Durán, P., Ellis, T. et al. (2020) Root microbiota assembly and adaptive differentiation
among European Arabidopsis populations. Ecol. Evol.4, 122–131. doi: 10.1038/s41559-019-1063-3.
64. Uroz, S., Buée, M., Deveau, A., Mieszkin, S., Martin, F. (2016). Ecology of the forest microbiome:
Highlights of temperate and boreal ecosystems. Soil Biol. Biochem.103, 471–488.
doi:10.1016/j.soilbio.2016.09.006
65. Wang, B. , Wu, Y. , Chen, D. (2017). Stable isotopes of carbon and nitrogen help to predict the
belowground communities at a regional scale. Rep. 7, 7276. doi: 10.1038/s41598-017-07517-w
66. Wang, P., Chen, Y., Sun, Y., Tan, S., Zhang, S., Wang, Z., Zhou, J., Zhang, G., Shu, W., Luo, C., Kuang, J.
(2019) Distinct biogeography of different fungal guilds and their associations with plant species
richness in forest ecosystems. Ecol. Evol.7, 216. doi: 10.3389/fevo.2019.00216
Page 14/2267. Wyatt, G. A. K. , Kiers, E. T. , Gardner, A. , West, S. A. . (2014). A biological market analysis of the plant-
mycorrhizal symbiosis. Evolution68(9), 2603-2618. doi: 10.1111/evo.12466
68. Yang, H., Zhao, X., Liu, C. et al. (2018) Diversity and characteristics of colonization of root-associated
fungi of Vaccinium uliginosum. Rep.8, 15283. doi 10.1038/s41598-018-33634-1
69. Yuan, S. M, Sun, X. J. (1990). The vegetational and environmental history at the west foot of
changbai mountain, northeast china during the last 1000 years. Acta Bot. Sinica.32(7):558-567.
70. Zeilinger, S., Gupta, V.K., Dahms, T.E.S., Silva, R.N., Singh, H.B., Upadhyay, R.S., Gomes, E.V., Tsui,
C.K.M., Nayak, S.C. (2015). Friends or foes? Emerging insights from fungal interactions with plants.
FEMS Microbiol. Rev.40, 182–207. doi: 1093/femsre/fuv045
Figures
Figure 1
Map showing sampling sites within 33 forest reserves in China. Note: The designations employed and the
presentation of the material on this map do not imply the expression of any opinion whatsoever on the
part of Research Square concerning the legal status of any country, territory, city or area or of its
Page 15/22authorities, or concerning the delimitation of its frontiers or boundaries. This map has been provided by
the authors.
Figure 2
Relationships between latitude and (A) soil fungal Shannon index, (B) ectomycorrhizal relative
abundance, (C) leaf δ15N, and (D) leaf δ13C. CB refers to samples from Changbai Mountain.
Page 16/22Figure 3
Relationships between longitude and (A) soil fungal Shannon index, (B) ectomycorrhizal relative
abundance, (C) leaf δ15N, and (D) leaf δ13C.
Page 17/22Figure 4
Relationships between mean annual temperature and (A) soil fungal Shannon index, (B) ectomycorrhizal
relative abundance, (C) leaf δ15N, and (D) leaf δ13C. CB refers to samples from Changbai Mountain.
Page 18/22Figure 5
Relationships between mean annual precipitation and (A) soil fungal Shannon index, (B) ectomycorrhizal
relative abundance, (C) leaf δ15N, and (D) leaf δ13C. CB refers to samples from Changbai Mountain.
Page 19/22Figure 6
Relationships between leaf δ15N and soil fungal (A) Shannon index, (B) plant pathogen, and (C)
ectomycorrhizal relative abundance.
Page 20/22Figure 7
Relationships between leaf δ13C and soil fungal (A) Shannon index, (B) plant pathogen, and (C)
ectomycorrhizal relative abundance.
Supplementary Files
Page 21/22This is a list of supplementary les associated with this preprint. Click to download.
SUPPLEMENTARYMATERIALS.docx
table.docx
FigS1.pdf
Page 22/22You can also read

















































