Sensing the environment: Response ofCandidaalbicansto the X factor
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
MINIREVIEW
Sensing the environment: Response of Candida albicans to the
X factor
Fabien Cottier & Fritz A. Mühlschlegel
Department of Biosciences, University of Kent, Canterbury, UK
Correspondence: Fritz A. Mühlschlegel, Abstract
Department of Biosciences, University of
Kent, Canterbury CT2 7NJ, UK. Tel.: 144
Virulence of the fungal human pathogen Candida albicans is also attributed to its
1227 823 988; fax: 144 1227 763 912; ability to switch reversibly between yeast and hyphal growth forms. Morphogenesis
e-mail: f.a.muhlschlegel@kent.ac.uk in this yeast is influenced by the composition of the environment, activating
sensors, which consequently play an important role in fungal pathogenicity. This
Received 22 January 2009; accepted 24 review summarizes some of the main environmental sensors, their ligands and
February 2009. downstream signaling pathways in C. albicans. We will focus on proteins localized
First published online 30 March 2009. in the plasma membrane and on the interaction between cells and their environ-
ment. This will underline the convergence of several environmental signals onto
DOI:10.1111/j.1574-6968.2009.01564.x
the mitogen-activated protein kinase and protein kinase A pathways.
Editor: Derek Sullivan
Keywords
Candida albicans ; sensor; signal pathway;
environment.
of this yeast. In this review, we will describe in greater detail
Introduction selected fungal sensors, their respective ligands and also the
Every environment is a complex assembly of physical and pathways activated subsequent to ligand–sensor interaction.
biological factors that can affect an organism. Gases, water, Although we will focus on C. albicans, the model yeast
ion concentration and carbon sources, in addition to Saccharomyces cerevisiae will be used to compare sensing
metabolites produced by other living cells, are just a few processes and explain some pathway activations, which are
examples of substances that profoundly impact on a micro- not yet fully described in C. albicans.
organism. The diploid opportunistic fungal pathogen Can-
dida albicans is able to colonize a large range of
environments, where conditions are highly variable. Indeed, Glucose
it has been isolated from sand (Ghinsberg et al., 1994), Sugars, in particular glucose, are common carbon sources
plastic, skin surfaces (Mok & Barreto da Silva, 1984), the used by microorganisms. To increase the efficiency of
oral cavity, gastrointestinal tract, blood, liver and other glucose uptake, cells have developed several strategies. For
organs where environments are unique. This flexibility of example, Gpr1p, a six-transmembrane domain G protein-
adaptation implies a strong capacity of sensing and the coupled receptor (GPCR) in S. cerevisiae, activates the
ability to respond swiftly to environmental changes. Clearly, cAMP pathway in response to glucose (Kraakman et al.,
a large number of sensors are required to integrate and 1999). Here the signal is transduced by means of a direct
transmit information about the external environment to the interaction between Gpr1p and the Ga protein, Gpa2p (Xue
appropriate response pathways. Over the past few years, et al., 1998). Inactivation of the orthologs in C. albicans
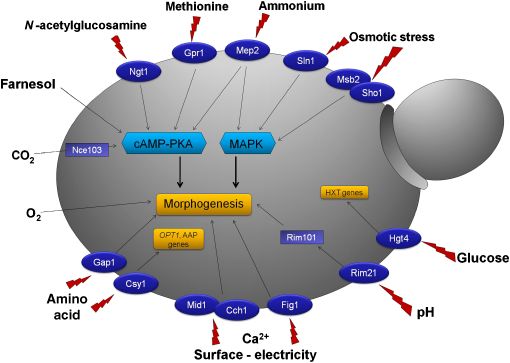
sensors for glucose, amino acids, ammonium, N-acetylglu- (GPR1 and GPA2) leads to a deficiency in hyphae formation
cosamine (GlcNAc) and farnesol, as well as gases, surfaces, on solid media (Miwa et al., 2004). The latter can be
electric fields, pH and osmotic stress have been identified rescued by the addition of exogenous cAMP. Furthermore,
(Fig. 1). Understanding how C. albicans senses its environ- the carboxy-terminus of Gpr1p was shown to interact with
ment is critical in light of the fact that most of these signals Gpa2p in a two-hybrid experiment (Miwa et al., 2004).
are known to be involved in controlling the morphogenesis Interestingly, the Gpr1p ortholog in C. albicans is a
FEMS Microbiol Lett 295 (2009) 1–9 c2009 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved2 F. Cottier & F.A. Mühlschlegel
Fig. 1. Schematic representation of identified
signals and sensors involved in morphogenesis
and gene regulation in Candida albicans. Putative
plasma membrane proteins (dark blue) are
represented with their respective stimulus (red
lightning bolt). Downstream activation of these
sensors is, if known, linked to the main pathways
involved (light blue).
seven-transmembrane domain protein that is not responsive In S. cerevisiae, Hxk2p is thought to function as an
to glucose but is responsive to methionine (Maidan et al., intracellular glucose sensor. This protein is able to bind and
2005). subsequently catalyze the phosphorylation of glucose in the
Saccharomyces cerevisiae also expresses 20 proteins from cytoplasm, but also localizes into the nucleus in the presence
the 12-transmembrane domain hexose transporter family. of this sugar molecule (Rández-Gil et al., 1998). Under the
Eighteen are transporters (Hxt1p–Hxt17p, Gal2p), and latter conditions, Hxk2p binds to the transcriptional repres-
Snf3p and Rgt2p are two proteins displaying features of sor Mig1p, known to regulate glucose-repressible genes
transporters but that also function as sensors; consequently, (Treitel & Carlson, 1995). The ortholog of Mig1p in
they are called ‘transceptors’ (Holsbeeks et al., 2004). C. albicans is a transcriptional repressor regulating gene
The Snf3p and Rgt2p glucose receptors impact on the involved in energy metabolism (Murad et al., 2001).
expression of the HXT genes via the transcription factor Furthermore, C. albicans Mig1p is able to functionally
Rgt1p, which operates as a repressor in the absence of complement the S. cerevisiae mutant (Zaragoza et al., 2000).
glucose, but as an activator in high concentrations of glucose
(Ozcan et al., 1996). In the C. albicans genome, a total of 19
hexose transporter orthologs have been identified (Sexton
Amino acids
et al., 2007). However, only a single ortholog for Snf3p and The SPS (Ssy1p–Ptr3p–Ssy5p) complex is a well-described
Rgt2p, Hgt4p, was found (Brown et al., 2006). Phenotypic amino acid-sensor system in S. cerevisiae (Forsberg &
analysis of a C. albicans hgt4 mutant showed defects in Ljungdahl, 2001). An ortholog for each protein of this
filamentation and virulence when compared with a wild- complex was identified in the genome of C. albicans. Csy1p,
type strain. A microarray study found that glucose induc- the C. albicans ortholog of the transceptor Ssy1p of the SPS
tion via Hgt4p induces the hexose transporter encoding complex is a 12-transmembrane domain protein predicted
genes HGT7, HXT10 and HGT12 (Brown et al., 2006). to be localized in the plasma membrane. Inactivation of CSY1
Furthermore, the hgt4 mutant is compromised in growth leads to the failure to induce the amino acid permease-
on solid media containing fructose, low concentrations of encoding (AAP) genes CAN1, GAP2 and GAP1 in the presence
glucose or mannose, and in the presence of antimycin A of histidine (Brega et al., 2004). In addition, on solid hyphal
(a respiration inhibitor). Finally, high concentrations of induction media (10% serum and Lee’s medium) mutants
glucose (4 0.1%) repress the expression levels of HGT4, grew principally as yeast, although this phenotype was not
which provides another argument to identify Hgt4p as a observed in liquid media or on Spider medium, or in response
glucose sensor in C. albicans (Brown et al., 2006). Finally, the to GlcNAc (Brega et al., 2004). As in S. cerevisiae, Csy1p
ortholog of Rgt1p in C. albicans acts downstream of Hgt4p activation induces the proteolytic cleavage of the two tran-
to repress several hexose transporter genes (Sexton et al., scription factors Stp1p and Stp2p, which remove the negative
2007). regulatory motif present in their N-terminal tail (Martı́nez &
c 2009 Federation of European Microbiological Societies FEMS Microbiol Lett 295 (2009) 1–9
Published by Blackwell Publishing Ltd. All rights reservedCandida albicans responses to the X factor 3
Ljungdahl, 2005). In S. cerevisiae, this reaction is mediated by on the same media (Biswas & Morschhäuser, 2005). Taken
Ptr3p and Ssy5p. Once Stp1p is activated, the protein migrates together, these results suggest that Mep2p is involved in the
to the nucleus and upregulates genes involved in protein activation of the MAPK and cAMP-PKA pathway through
degradation (SAP2) and peptide uptake (OPT1), which Ras1p. MEP2 expression is regulated by the two GATA
explains why an stp1 mutant is not able to grow on medium transcription factors Gat1p and Gln3p (Dabas &
with protein as the sole nitrogen source (Martı́nez & Ljung- Morschhäuser, 2007). Gln3p functions as an activator of
dahl, 2005). Stp2p activation is responsible for the positive MEP2 transcription under nitrogen-limiting conditions
regulation of AAP genes (CAN2, GAP2 and GAP1) (Martı́nez (Dabas & Morschhäuser, 2007). These results are in agree-
& Ljungdahl, 2005). ment with the findings in S. cerevisiae where Gln3p regulates
In S. cerevisiae, the 12-transmembrane domain protein MEP2 expression under nitrogen-limiting conditions. Here
Gap1p is thought to function as a ‘transceptor’. In addition Gln3p is phosphorylated through TOR kinases (Beck &
to its role as amino-acid transporter (Jauniaux & Grenson, Hall, 1999).
1990), this protein is involved in pathway activation of PKA
targets (Donaton et al., 2003). In C. albicans, the gap1
mutant is deficient in hyphae formation in response to
Gas sensing (CO2 and O2)
GlcNAc and nitrogen starvation but not to serum activation Candida albicans has been isolated from environments that
(Biswas et al., 2003). GlcNAc is also responsible for tran- differ significantly in the concentration of CO2, from
script regulation of GAP1 in a Cph1p-, Ras1p-dependent 0.033% (skin surface) (Ayliffe et al., 1988) to 5–6% (internal
manner. Thus, the MAPK pathway is involved in GAP1 organs) (Guyton & Hall, 2000; Monnin et al., 2001). In these
regulation and consequently could also be involved in niches, the concentration of O2 is also variable (Kessler et al.,
Gap1p-mediated filamentation (Biswas et al., 2003). Finally, 1974). CO2 was demonstrated to be a strong inducer of
the amino-acid sensor Gpr1p, an ortholog of a glucose filamentation; a wild-type strain produces hyphae in a 5%
sensor in S. cerevisiae, is responsive to methionine. The CO2-enriched environment, whereas under the same condi-
latter promotes the yeast-to-hyphae transition in a Gpr1p- tions, but in air, (normo-capneic conditions) this strain
dependent manner (Maidan et al., 2005). grows in a yeast form (Klengel et al., 2005). This phenotype
is lost in an adenylate cyclase mutant (cyr1). Hyphae
induction by CO2 required the C. albicans adenylyl cyclase
Ammonium Cyr1p, and thereby cAMP, to activate the PKA pathway.
In low concentrations of ammonium as the sole nitrogen However, this activation seems to occur independently of
source, the two 11-transmembrane domain ammonium Ras1p (Klengel et al., 2005). In mammalian as well as
permeases Mep1p and Mep2p are essential for the growth bacterial cells, adenylate cyclase is described as directly
of C. albicans (Biswas & Morschhäuser, 2005). While Mep1p responsive to the concentration of bicarbonate (the hy-
is considered to be a simple ammonium transporter, Mep2p drated form of CO2 inside the cell) (Chen et al., 2000). The
is a transceptor in light of its involvement in the yeast-to- presence of this molecule changes the conformation of the
hyphae switch of C. albicans on nitrogen starvation medium enzyme and increases cAMP production (Steegborn et al.,
(SLAD) (Biswas & Morschhäuser, 2005). Comparison of 2005), identifying adenylate cyclase as a CO2 sensor. Bicar-
ammonium uptake and experiments with radioactive bonate ions (HCO–3), spontaneously produced from CO2 in
methylammonium on different MEP mutants and wild-type aqueous solution, seem to represent the active form of CO2
strains demonstrate that Mep2p is a less-efficient ammo- in the cells, as shown in bacteria and mammalian cells.
nium transporter compared with Mep1p (Biswas & However, a recent report demonstrates that a subset of
Morschhäuser, 2005). Mep2p possesses a cytoplasmic adenylyl cyclases is activated directly by molecular CO2
C-terminal tail of 74 amino acid residues that is considered (Townsend et al., 2009). Inactivation of the carbonic anhy-
essential for its function. Notably, removal of the first 57 drase (NCE103), which enhances the natural hydration of
amino acid residues of this domain leads to the loss of CO2 to bicarbonate, abrogates C. albicans’ growth in air.
filamentation on SLAD medium, whereas its function in Supplementation of the atmosphere with CO2 rescues the
ammonium uptake is not altered. Thus, the C-terminal tail growth defect of the nce103 mutant (Klengel et al., 2005).
of Mep2p is critical to transduce the signal, but dispensable Furthermore, increase of NaHCO3 concentration to physio-
for the transport of ammonium (Biswas & Morschhäuser, logical levels stimulates recombinant Cyr1p, suggesting that
2005). The introduction of a hyperactive allele of MEP2 the adenylate cyclase is a cytoplasmic sensor for CO2. In
(MEP2DC440) in a double efg1 cph1 mutant, or single ras1 higher eukaryotes, CO2 has been reported to pass the plasma
mutant, is not able to complement the filamentation defi- membrane through aquaporin water pores (Endeward et al.,
ciency on SLAD medium. However, this allele can comple- 2006, 2008), although this is still controversial (Missner
ment the filamentation deficit of single efg1 or cph1 mutants et al., 2008). In C. albicans, only a single aquaporin ortholog
FEMS Microbiol Lett 295 (2009) 1–9 c2009 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved4 F. Cottier & F.A. Mühlschlegel
has been identified (Aqy1p), but its inactivation does not tion of C. albicans hyphae to the cathode in an electrical
lead to any deficit in filamentation when the strain is field, it appears that only Cch1p is involved in this process
exposed to elevated CO2 concentrations (Klengel et al., (Brand et al., 2007). Thigmotropism and galvanotropism
2005). can be explained by a local increase of the Ca21 concentra-
CO2 is not the only gas that impacts on C. albicans hyphae tion inside the cell. This happens when a membrane stretch
formation. Indeed, low concentration of oxygen is an activator occurs after contact with a surface, or by depolarization of
of the yeast-to-hyphae switch (Setiadi et al., 2006). In addition, the membrane, as in the case of galvanotropism. Under
microaerophilic conditions are required for C. albicans chla- these conditions, calcium transporters are activated and
mydospore formation (big thick-walled spores) in an Efglp- produce a local increase of Ca21 concentration, which
dependent manner (Sonneborn et al., 1999). induces hyphal reorientation (Brand et al., 2007). Recently,
The transcription factor Efg1p, involved in the PKA Rsr1p (a Ras-like GTPase) and Bud2p (its GTPase-activating
pathway, plays an important role in the regulation of protein) have been shown to exhibit important roles in both
morphogenesis under hypoxic (99.9% N2) and embedded thigmotropism and galvanotropism (Brand et al., 2008).
conditions (Doedt et al., 2004; Setiadi et al., 2006). Here, The function of these proteins has not been fully clarified,
Efg1p is considered to function as a repressor due to the fact but they could act in the positioning of other proteins
that an efg1 mutant is hyperfilamentous. However, in involved in hyphal orientation.
normoxia (air), Efg1p has a positive role in hyphae forma-
tion (Stoldt et al., 1997). Ace2p and Czf1p are two other
transcription factors that are involved in the C. albicans
Stress
response to hypoxia. Inactivation of ACE2 leads to an Fungi and in particular C. albicans are responsive not only to
increase in expression of respiratory genes and to a defect the chemical composition of their environment but also to
in hyphae production in response to hypoxia (Mulhern its physical condition. Physical parameters such as oxidative
et al., 2006). These observations suggest that reduced and osmotic stress and also temperature, UV and antifungal
respiration may be the trigger for hypoxia-induced filamen- drugs are sensed by C. albicans through the HOG pathway.
tation. Contrary to Efg1p, Czf1p is positively involved in Hog1p is the MAP kinase responsible for stress resistance in
hyphae formation under embedded conditions (Brown yeast; it is activated by two pathways, which have already
et al., 1999). Furthermore, Czf1p is involved in the white/ been fully characterized in S. cerevisiae (Saito & Tatebayashi,
opaque switch in C. albicans under hypoxic conditions 2004). The main and historically first discovered pathway
(Ramı́rez-Zavala et al., 2008). involves a histidine kinase localized in the plasma mem-
brane, Sln1p, which is well described in S. cerevisiae (Posas
et al., 1996). In C. albicans, all members of this pathway
Thigmotropism and galvanotropism possess an ortholog, but elucidation of the interactions
In C. albicans, morphological differentiation is not only between the individual members is not as comprehensive
activated by a molecule, but can also be induced through yet. In the absence of stresses, Sln1p continuously phosphor-
contact with a surface (thigmotropism) or exposure to an ylates its coupling protein, Ypd1p, which in turn phosphor-
electrical field (galvanotropism). Thigmotropism is a well- ylates Ssk1p, blocking the activation of the MAPK pathway
defined process in other fungi such as Magnaporthe grisea (Posas et al., 1996). When stress is sensed, Ssk1p activates
(Lee & Dean, 1993). To penetrate a leaf, M. grisea differ- Ssk2/22p, which in turn transduces the signal to Pbs2p, and
entiate a specialized cell called appresorium. However, this then Hog1p, which migrates to the nucleus to control the
phenomenon will occur only if the fungi recognize a certain expression of target genes (Ferrigno et al., 1998). The second
hydrophobicity of the contact surface (Lee & Dean, 1993). In pathway in Hog1p activation involves the two plasma
C. albicans, thigmotropism has been reported when growing membrane proteins Sho1p and Msb2p. Both proteins play
hyphae come into contact with a ridge (0.79 mm 40 nm). a role in the activation of the MAPK Cek1p, and function in
On a smooth surface, hyphae keep their orientation, but if the control of C. albicans filamentation (Roman E, Cottier F,
hyphae cross a ridge, they modify their growth axis in 60% Ernst JF & Pla J, unpublished data). Cross-talk between the
of cases as opposed to continuing through the ridge (Brand two pathways exists in S. cerevisiae through Ste11p and
et al., 2007). This response involves two plasma membrane Pbs2p (Posas & Saito, 1997). In contrast, this interaction
proteins, Mid1p and Cch1p, which are components of the does not exist in C. albicans (Cheetham et al., 2007).
high-affinity calcium uptake system, and Fig1p, a member Furthermore, how exactly Sln1p, Sho1p and Msb2p sense
of the low-affinity calcium system (Brand et al., 2007). stresses is unknown. One hypothesis, proposed by Tatebaya-
Strains lacking one of these proteins display a decrease in shi et al. (2007), is that highly glycosylated proteins like
growth axis modification subsequent to contacting the Msb2p could form a gel-like structure on the surface of the
ridge. In the case of galvanotropism, which involves orienta- cells that can sense high osmolarity, and subsequently
c 2009 Federation of European Microbiological Societies FEMS Microbiol Lett 295 (2009) 1–9
Published by Blackwell Publishing Ltd. All rights reservedCandida albicans responses to the X factor 5
modify the three-dimensional structure or the spatial or- a transporter of GlcNAc that could subsequently activate a
ientation of the protein to induce signal transduction. pathway involved in morphogenesis.
Chk1p and Nik1p are two other histidine kinase sensors
localized in the cytoplasm and involved in several processes Farnesol and homoserine lactone sensing
including morphogenesis, osmosensing and cell wall bio-
Quorum-sensing in C. albicans was established in 2001 by
genesis (Yamada-Okabe et al., 1999; Kruppa et al., 2003).
Hornby et al. (2001), who discovered that C. albicans
produce and secrete the sesquiterpene farnesol, which acts
pH as a hyphal repressor. Since then, farnesol sensing has been
Another important physical property of the fungal environ- described to be closely related to the Ras1p-cAMP-PKA
ment is the pH. Candida albicans modifies its morphology pathway. Indeed, addition of exogenous cAMP to the
depending of the pH value, ranging from yeast growth in culture medium containing farnesol restores filamentation
acidic conditions to hyphae formation in alkaline media (Davis-Hanna et al., 2008). However, the C. albicans sensor
(Davis, 2003). Such responses to pH are clearly relevant in for farnesol has not been identified. Farnesol does not
the pathogenicity of this fungus (De Bernardis et al., 1998). appear to be acting directly on Ras1p, because a strain
In S. cerevisiae (Su & Mitchell, 1993) pH sensing involves the constitutively expressing Ras1p does not bypass the farnesol
transcription factor Rim101p, activated at alkaline pH by repression of filamentation (Davis-Hanna et al., 2008). But
proteolysis of its C-terminal tail (Li & Mitchell, 1997). The Tup1p, a negative regulator of hyphal switch, acts in the
pathway also requires the proteins Rim101p, Rim8p, regulation of morphogenesis in response to farnesol (Ke-
Rim13p, Rim20p and Rim21p (Davis, 2003). The predicted baara et al., 2008). Indeed, a tup1 mutant is refractory to the
sensor of pH is the seven-transmembrane domain protein inhibitory effect of farnesol (Kebaara et al., 2008), which
Rim21p, localized in the plasma membrane (Davis, 2003). places the proteins involved in Tup1p activation as a possible
According to this theory, modification of the protonation of target for farnesol sensors. In a recent report, Xu et al. (2008)
charged amino acids in Rim21p induces a conformational show that the leucine-rich repeat domain of Cyr1p, which is
change of the protein and so activation of the pathway. important for hyphae induction, interacts with derivates of
Thus, at alkaline pH, Rim21p transduces a signal to Rim13p, bacterial molecules such as peptidoglycans. This interaction
which cleaves the C-terminal domain of Rim101p. Once leads to an increase of cAMP production and promotes the
Rim101p is truncated and thus activated, it then plays a yeast-to-hyphae transition (Xu et al., 2008). In analogy to
major role in the regulation of many effector genes (Bensen the above, a putative Ras1p-Cyr1p complex could interact
et al., 2004). with farnesol to repress adenylate cyclase activity and cAMP
production, leading to a repression of filamentation.
GlcNAc Inhibition of C. albicans filamentation by quorum-sen-
sing molecules was also observed with 3-oxo-C12 homo-
The amino sugar GlcNAc, present in several organisms, is
serine lactone, a quorum-sensing molecule produced by the
implied in cellular signaling processes. For example, it is
opportunistic gram-negative bacterium Pseudomonas aeru-
involved in Escherichia coli adhesin production (Sohanpal
ginosa coexisting with C. albicans in certain body niches.
et al., 2004), and was identified to induce hyphae formation
The effect of 3-oxo-C12 homoserine lactone can be mi-
in C. albicans (Mattia et al., 1982). The role of GlcNac in
micked with other 12-carbon backbone molecules such as
GAP1 expression has been described previously, but it is
dodecanol (Hogan et al., 2004). As with farnesol, no specific
unknown whether Gap1p is a sensor of GlcNAc (Biswas
sensor has yet been described for P. aeruginosa quorum-
et al., 2003). Ngt1p is a 12-transmembrane domain protein
sensing molecules.
acting as a GlcNAc transporter that is also involved in
C. albicans hyphal differentiation (Alvarez & Konopka,
2006). Using a green fluorescent protein construct, Ngt1p is
Pheromones
localized in the plasma membrane when cells are exposed to The pheromone-sensing pathway has been characterized in
GlcNAc. Saccharomyces cerevisiae is not able to metabolize great detail in S. cerevisiae (Dohlman & Slessareva, 2006).
GlcNAC and no ortholog of Ngt1p has been identified Sensing the pheromone from the opposite mating type (a or
(Alvarez & Konopka, 2006). However, introduction of an a) promotes remodeling of the cytoskeleton and nuclear
NGT1 allele into S. cerevisiae allows GlcNAc uptake in the fusion of the cells to complete the sexual cycle (Read et al.,
yeast (Alvarez & Konopka, 2006). A C. albicans ngt1 mutant 1992). Ste2p and Ste3p are the two GPCRs characterized as
is defective in hyphal formation in response to 2.5 mM of pheromone sensors in S. cerevisiae (Versele et al., 2001),
GlcNAc, but not to 100 mM or to the addition of serum sensing, respectively, a and a pheromone. Both are
(Alvarez & Konopka, 2006). Up until now it is still unknown 7-transmembrane domain proteins, with a cytoplasmic tail
whether Ngt1p has a real sensor function or whether it is just interacting with the GTPase-activating protein Sst2p. The
FEMS Microbiol Lett 295 (2009) 1–9 c2009 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved6 F. Cottier & F.A. Mühlschlegel
latter activates the GTP hydrolysis on Gpa1p (Ga protein) C. albicans sensing of surface reliefs (Brand et al., 2007).
subsequent to pheromone binding to the respective recep- However, thigmotropism can also involve sensing the hy-
tor. After this, Gpa1p separates from the bg dimer (named drophobicity of a surface (Lee & Dean, 1993). One example
Ste4p and Ste18p, respectively), and activates the Ste12p- is the plant pathogenic fungus M. grisea, where hydrophobic
MAPK pathway, leading to cell-cycle arrest and cell fusion. surfaces are required to induce appresorium formation (Lee
In C. albicans, the following genes involved in pheromone & Dean, 1993). A large range of other molecules encoun-
production were identified: MFA1 encoding for the a-factor tered by C. albicans currently have no clear sensors like
pheromone (Dignard et al., 2007) and MFALPHA for the mannitol, which is present in Lee and Spider media to
a-factor (Bennett et al., 2003). The Ste2p ortholog in enhance filamentation (Csank et al., 1998). In light of the
C. albicans (orf19.696) is needed for the morphological vast amount of information C. albicans receives from its
response of a cells to a-factor (Bennett et al., 2003). environment, it is clear that not all signals will lead to a
Recently, in addition to this sensor, two G protein subunits uniform response. Indeed, cells have to integrate the differ-
have been show to play a role in mating: Gpa1p and Ste4p ent signals to coordinate which response is best for survival.
(Dignard et al., 2008). Their interactions with the sensor are This process of information integration has only recently
not yet proven, but show that a similar complex to being addressed in C. albicans (Tuch et al., 2008). Finally,
S. cerevisiae is present in C. albicans. Mating in C. albicans even if a number of sensors have been identified, their
is possible after cells switch from white (round yeast) to method of interaction with the ligand and the structural
opaque cells (elongated yeast) (Slutsky et al., 1987). Opaque, modifications required to transduce the signal are largely
but not white, cells are able to release pheromone, to shmoo unknown. Clearly, this will have to be investigated for a
and mate (Miller & Johnson, 2002). Pheromones produced better understanding of the interaction of C. albicans with
by opaque cells can also promote biofilm formation in white its environment.
cells (Daniels et al., 2006).
Acknowledgements
Conclusion
In light of the space restrictions, important work from a
Several signals and their respective sensors have been
number of colleagues working in the field could not be cited.
discovered in C. albicans, allowing a better understanding
F.A.M. thanks Campbell W. Gourlay from the Kent Fungal
of how this fungus senses and responds to its environment.
Group for critically reading the manuscript. We would like
The vast majority of them seem to play a role in morpho-
to thank the two anonymous reviewers for their constructive
genesis, probably because it is one of the principal pheno-
comments. F.C. is a Postdoctoral Research Assistant funded
types studied in C. albicans due to its involvement in
by the BBSRC. Work in F.A.M.’s lab is funded by the MRC
pathogenicity. This highlights the possibility that other
and BBSRC.
sensors not involved in the yeast-to-hyphae switch are being
overlooked. Furthermore, although several sensors were
identified, they do not cover all signals from the environ-
References
ment, temperature being one example. In bacteria, tempera-
ture is described as directly influencing the structure of Aguilar PS, Hernandez-Arriga AM, Cybulsky LE, Erazo AC & de
DNA, RNA and protein and consequently their function Mendoza D (2001) Molecular basis of thermosensing: a two-
(Eriksson et al., 2002). In addition, the two-component component signal transduction thermometer in Bacillus
system DesKp/DesRp has been identified as a temperature subtilis. EMBO J 20: 1681–1691.
sensor. DesKp activity is dependent on plasma membrane Alvarez FJ & Konopka JB (2006) Identification of an
properties, which are altered by temperature. This modifica- N-acetylglucosamine transporter that mediates hyphal
induction in Candida albicans. Mol Biol Cell 18: 965–975.
tion in DesKp function is subsequently recognized by the
Ayliffe GA, Babb JR, Davies JG & Lilly HA (1988) Hand
DesRp gene regulator, which in turn induces the expression
disinfection: a comparison of various agents in laboratory and
of a lipid desaturase (Aguilar et al., 2001). Until now, an
ward studies. J Hosp Infect 11: 226–243.
analogous system has not been described in C. albicans. Beck T & Hall MN (1999) The TOR signalling pathway controls
Light and darkness are also strong signals. In C. albicans, the nuclear localization of nutrient-regulated transcription
production of chlamydospores is linked to darkness (Nobile factors. Nature 402: 689–692.
et al., 2003) and in other fungi like Neurospora crassa Bennett RJ, Uhl MA, Miller MG & Johnson AD (2003)
(Corrochano, 2007) the switch between light and darkness Identification and characterization of a Candida albicans
is critical for the circadian clock regulation and sporulation. mating pheromone. Mol Cell Biol 23: 8189–8201.
However, no photosensors have been described as yet in Bensen ES, Martin SJ, Li M, Berman J & Davis DA (2004)
C. albicans. Thigmotropism was shown to be involved in Transcriptional profiling in Candida albicans reveals new
c 2009 Federation of European Microbiological Societies FEMS Microbiol Lett 295 (2009) 1–9
Published by Blackwell Publishing Ltd. All rights reservedCandida albicans responses to the X factor 7
adaptive responses to extracellular pH and functions for cAMP signalling pathway and the regulation of
Rim101p. Mol Microbiol 54: 1335–1351. morphogenesis. Mol Microbiol 67: 47–62.
Biswas K & Morschhäuser J (2005) The Mep2p ammonium De Bernardis F, Mühlschlegel FA, Cassone A & Fonzi WA (1998)
permease controls nitrogen starvation-induced filamentous The pH of the host niche controls gene expression in and
growth in Candida albicans. Mol Microbiol 56: 649–669. virulence of Candida albicans. Infect Immun 66: 3317–3325.
Biswas S, Roy M & Datta A (2003) N-acetylglucosamine- Dignard D, El-Naggar AL, Logue ME, Butler G & Whiteway M
inducible CaGAP1 encodes a general amino acid permease (2007) Identification and characterization of MFA1, the gene
which co-ordinates external nitrogen source response and encoding Candida albicans a-factor pheromone. Eukaryot Cell
morphogenesis in Candida albicans. Microbiology 149: 6: 487–494.
2597–2608. Dignard D, André D & Whiteway M (2008) Heterotrimeric
Brand A, Shanks S, Duncan VM, Yang M, Mackenzie K & Gow G-protein subunit function in Candida albicans: both the
NA (2007) Hyphal orientation of Candida albicans is regulated alpha and beta subunits of the pheromone response G protein
by a calcium-dependent mechanism. Curr Biol 17: 347–352. are required for mating. Eukaryot Cell 7: 1591–1599.
Brand A, Vacharaksa A, Bendel C, Norton J, Haynes P, Henry- Doedt T, Krishnamurthy S, Bockmuhl DP, Tebarth B, Stempel C,
Stanley M, Wells C, Ross K, Gow NA & Gale CA (2008) An Russell CL, Brown AJ & Ernst JF (2004) APSES proteins
internal polarity landmark is important for externally induced regulate morphogenesis and metabolism in Candida albicans.
hyphal behaviors in Candida albicans. Eukaryot Cell 7: Mol Biol Cell 15: 3167–3180.
712–720. Dohlman HG & Slessareva JE (2006) Pheromone signaling
Brega E, Zufferey R & Mamoun CB (2004) Candida albicans pathways in yeast. Sci STKE 2006: cm6.
Csy1p is a nutrient sensor important for activation of amino Donaton MC, Holsbeeks I, Lagatie O, Van Zeebroeck G, Crauwels
acid uptake and hyphal morphogenesis. Eukaryot Cell 3: M, Winderickx J & Thevelein JM (2003) The Gap1 general
135–143. amino acid permease acts as an amino acid sensor for
Brown DH Jr, Giusani AD, Chen X & Kumamoto CA (1999) activation of protein kinase A targets in the yeast
Filamentous growth of Candida albicans in response to Saccharomyces cerevisiae. Mol Microbiol 50: 911–929.
physical environmental cues and its regulation by the unique Endeward V, Musa-Aziz R, Cooper GJ, Chen LM, Pelletier MF,
CZF1 gene. Mol Microbiol 34: 651–662. Virkki LV, Supuran CT, King LS, Boron WF & Gros G (2006)
Brown V, Sexton JA & Johnston M (2006) A glucose sensor in Evidence that aquaporin 1 is a major pathway for CO2 transport
Candida albicans. Eukaryot Cell 5: 1726–1737. across the human erythrocyte membrane. FASEB J 20: 1974–1981.
Cheetham J, Smith DA, da Silva Dantas A, Doris KS, Patterson Endeward V, Cartron JP, Ripoche P & Gros G (2008) RhAG
MJ, Bruce CR & Quinn J (2007) A Single MAPKKK regulates protein of the Rhesus complex is a CO2 channel in the human
the Hog1 MAPK pathway in the pathogenic fungus Candida red cell membrane. FASEB J 22: 64–73.
albicans. Mol Biol Cell 18: 4603–4614. Eriksson S, Hurme R & Rhen M (2002) Low-temperature sensors
Chen Y, Cann MJ, Litvin TN, Lourgenko V, Sinclair ML, Levin LR in bacteria. P Trans Roy Soc B 357: 887–893.
& Buck J (2000) Soluble adenylyl cyclase as an evolutionarily Ferrigno P, Posas F, Koepp D, Saito H & Silver PA (1998) Regulated
conserved bicarbonate sensor. Science 289: 625–628. nucleo/cytoplasmic exchange of HOG1 MAPK requires the
Corrochano LM (2007) Fungal photoreceptors: sensory importin b homologs NMD5 and XPO1. EMBO J 17: 5606–5614.
molecules for fungal development and behaviour. Photochem Forsberg H & Ljungdahl PO (2001) Genetic and biochemical
Photobio Sci 6: 725–736. analysis of the yeast plasma membrane Ssy1p–Ptr3p–Ssy5p
Csank C, Schröppel K, Leberer E, Harcus D, Mohamed O, sensor of extracellular amino acids. Mol Cell Biol 21: 814–826.
Meloche S, Thomas DY & Whiteway M (1998) Roles of the Ghinsberg RC, Bar Dov L, Rogol M, Sheinberg Y & Nitzan Y
Candida albicans mitogen-activated protein kinase homolog, (1994) Monitoring of selected bacteria and fungi in sand and
Cek1p, in hyphal development and systemic candidiasis. Infect sea water along the Tel Aviv coast. Microbios 77: 29–40.
Immun 66: 2713–2721. Guyton AC & Hall JE (2000) Textbook of Medical Physiology. W.B.
Dabas N & Morschhäuser J (2007) Control of ammonium Saunders, Philadelphia.
permease expression and filamentous growth by the GATA Hogan DA, Vik A & Kolter R (2004) A Pseudomonas aeruginosa
transcription factors GLN3 and GAT1 in Candida albicans. quorum-sensing molecule influences Candida albicans
Eukaryot Cell 6: 875–888. morphology. Mol Microbiol 54: 1212–1223.
Daniels KJ, Srikantha T, Lockhart SR, Pujol C & Soll DR (2006) Holsbeeks I, Lagatie O, Van Nuland A, Van de Velde S &
Opaque cells signal white cells to form biofilms in Candida Thevelein JM (2004) The eukaryotic plasma membrane as a
albicans. EMBO J 25: 2240–2252. nutrient-sensing device. Trends Biochem Sci 29: 556–564.
Davis D (2003) Adaptation to environmental pH in Candida Hornby JM, Jensen EC, Lisec AD, Tasto JJ, Jahnke B, Shoemaker
albicans and its relation to pathogenesis. Curr Genet 44: 1–7. R, Dussault P & Nickerson KW (2001) Quorum sensing in the
Davis-Hanna A, Piispanen AE, Stateva LI & Hogan DA (2008) dimorphic fungus Candida albicans is mediated by farnesol.
Farnesol and dodecanol effects on the Candida albicans Ras1- Appl Environ Microb 67: 2982–2992.
FEMS Microbiol Lett 295 (2009) 1–9 c2009 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reserved8 F. Cottier & F.A. Mühlschlegel
Jauniaux JC & Grenson M (1990) GAP1, the general amino acid formation in the pathogenic fungus Candida albicans.
permease gene of Saccharomyces cerevisiae. Nucleotide Eukaryot Cell 3: 919–931.
sequence, protein similarity with the other bakers yeast amino Mok WY & Barreto da Silva MS (1984) Mycoflora of the human
acid permeases, and nitrogen catabolite repression. Eur J dermal surfaces. Can J Microbiol 30: 1205–1209.
Biochem 190: 39–44. Monnin E, Indermuhle A, Dallenbach A, Fluckiger J, Stauffer B,
Kebaara BW, Langford ML, Navarathna DH, Dumitru R, Stocker TF, Raynaud D & Barnola JM (2001) Atmospheric
Nickerson KW & Atkin AL (2008) Candida albicans Tup1 is CO2 concentrations over the last glacial termination. Science
involved in farnesol-mediated inhibition of filamentous- 291: 112–114.
growth induction. Eukaryot Cell 7: 980–987. Mulhern SM, Logue ME & Butler G (2006) Candida albicans
Kessler M, Höper J, Schäfer D & Starlinger H (1974) transcription factor Ace2 regulates metabolism and is required
Sauerstofftransport im Gewebe. Mikrozirkulation (Ahnefeld for filamentation in hypoxic conditions. Eukaryot Cell 5:
FW, Burri C, Dick W & Halmágyi M, eds), pp. 36–52. 2001–2013.
Springer-Verlag, Berlin. Murad AM, d’Enfert C, Gaillardin C, Tournu H, Tekaia F, Talibi
Klengel T, Liang WJ, Chaloupka J et al. (2005) Fungal adenylyl D, Marechal D, Marchais V, Cottin J & Brown AJ (2001)
cyclase integrates CO2 sensing with cAMP signalling and Transcript profiling in Candida albicans reveals new cellular
virulence. Curr Biol 15: 2021–2026. functions for the transcriptional repressors CaTup1, CaMig1
Kraakman L, Lemaire K, Ma P, Teunissen AW, Donaton MC, Van and CaNrg1. Mol Microbiol 42: 981–993.
Dijck P, Winderickx J, de Winde JH & Thevelein JM (1999) A Nobile CJ, Bruno VM, Richard ML, Davis DA & Mitchell AP
Saccharomyces cerevisiae G-protein coupled receptor, Gpr1, is (2003) Genetic control of chlamydospore formation in
specifically required for glucose activation of the cAMP
Candida albicans. Microbiology 149: 3629–3637.
pathway during the transition to growth on glucose. Mol Ozcan S, Leong T & Johnston M (1996) Rgt1p of Saccharomyces
Microbiol 32: 1002–1012.
cerevisiae, a key regulator of glucose-induced genes, is both an
Kruppa M, Goins T, Cutler JE et al. (2003) The role of the
activator and a repressor of transcription. Mol Cell Biol 16:
Candida albicans histidine kinase (CHK1) gene in the
6419–6426.
regulation of cell wall mannan and glucan biosynthesis. FEMS
Posas F & Saito H (1997) Osmotic activation of the HOG MAPK
Yeast Res 3: 289–299.
pathway via Ste11p MAPKKK: scaffold role of Pbs2p MAPKK.
Lee YH & Dean RA (1993) cAMP regulates infection structure
Science 276: 1702–1705.
formation in the plant pathogenic fungus Magnaporthe grisea.
Posas F, Wurgler-Murphy SM, Maeda T, Witten EA, Thai TC &
Plant Cell 5: 693–700.
Saito H (1996) Yeast HOG1 MAP kinase cascade is regulated
Li W & Mitchell AP (1997) Proteolytic activation of Rim1p, a
by a multistep phosphorelay mechanism in the SLN1-YPD1-
positive regulator of yeast sporulation and invasive growth.
SSK1 ‘two-component’ osmosensor. Cell 86: 865–875.
Genetics 145: 63–73.
Ramı́rez-Zavala B, Reuss O, Park YN, Ohlsen K & Morschhäuser J
Maidan MM, De Rop L, Serneels J, Exler S, Rupp S, Tournu H,
Thevelein JM & Van Dijck P (2005) The G protein-coupled (2008) Environmental induction of white-opaque switching in
receptor Gpr1 and the Galpha protein Gpa2 act through the Candida albicans. PLoS Pathog 4: e1000089.
cAMP-protein kinase A pathway to induce morphogenesis in Rández-Gil F, Herrero P, Sanz P, Prieto JA & Moreno F (1998)
Candida albicans. Mol Biol Cel 16: 1971–1986. Hexokinase PII has a double cytosolic-nuclear localisation in
Martı́nez P & Ljungdahl PO (2005) Divergence of Stp1 and Stp2 Saccharomyces cerevisiae. FEBS Lett 425: 475–478.
transcription factors in Candida albicans places virulence Read EB, Okamura HH & Drubin DG (1992) Actin- and tubulin-
factors required for proper nutrient acquisition under amino dependent functions during Saccharomyces cerevisiae mating
acid control. Mol Cell Biol 25: 9435–9446. projection formation. Mol Biol Cell 3: 429–444.
Mattia E, Carruba G, Angiolella L & Cassone A (1982) Induction Saito H & Tatebayashi K (2004) Regulation of the osmoregulatory
of germ tube formation by N-acetyl-D-glucosamine in HOG MAPK cascade in yeast. J Biochem 136: 267–272.
Candida albicans: uptake of inducer and germinative response. Setiadi ER, Doedt T, Cottier F, Noffz C & Ernst JF (2006)
J Bacteriol 152: 555–562. Transcriptional response of Candida albicans to hypoxia:
Miller MG & Johnson AD (2002) White–opaque switching in linkage of oxygen sensing and Efg1p-regulatory networks.
Candida albicans is controlled by mating-type locus J Mol Biol 361: 399–411.
homeodomain proteins and allows efficient mating. Cell 110: Sexton JA, Brown V & Johnston M (2007) Regulation of sugar
293–302. transport and metabolism by the Candida albicans Rgt1
Missner A, Kügler P, Saparov SM, Sommer K, Mathai JC, Zeidel transcriptional repressor. Yeast 24: 847–860.
ML & Pohl P (2008) Carbon dioxide transport through Slutsky B, Staebell M, Anderson J, Risen L, Pfaller M & Soll DR
membranes. J Biol Chem 283: 25340–25347. (1987) ‘White–opaque transition’: a second high-frequency
Miwa T, Takagi Y, Shinozaki M, Yun CW, Schell WA, Perfect JR, switching system in Candida albicans. J Bacteriol 169: 189–197.
Kumagai H & Tamaki H (2004) Gpr1, a putative G-protein- Sohanpal BK, El-Labany S, Lahooti M, Plumbridge JA &
coupled receptor, regulates morphogenesis and hypha Blomfield IC (2004) Integrated regulatory responses of fimB to
c 2009 Federation of European Microbiological Societies FEMS Microbiol Lett 295 (2009) 1–9
Published by Blackwell Publishing Ltd. All rights reservedCandida albicans responses to the X factor 9
N-acetylneuraminic (sialic) acid and GlcNAc in Escherichia Treitel MA & Carlson M (1995) Repression by SSN6-TUP1 is
coli K-12. P Natl Acad Sci USA 101: 16322–16327. directed by MIG1, a repressor activator protein. P Natl Acad
Sonneborn A, Bockmuhl DP & Ernst JF (1999) Chlamydospore Sci USA 92: 3132–3136.
formation in Candida albicans requires the Efg1p Tuch BB, Galgoczy DJ, Hernday AD, Li H & Johnson AD (2008)
morphogenetic regulator. Infect Immun 67: 5514–5517. The evolution of combinatorial gene regulation in fungi. PLoS
Steegborn C, Litvin TN, Levin LR, Buck J & Wu H (2005) Biol 6: e38.
Bicarbonate activation of adenylyl cyclase via promotion of Versele M, Lemaire K & Thevelein JM (2001) Sex and sugar in
catalytic active site closure and metal recruitment. Nat Struct yeast: two distinct GPCR systems. EMBO Rep 2: 574–579.
Mol Biol 12: 32–37. Xu XL, Lee RT, Fang HM, Wang YM, Li R, Zou H, Zhu Y & Wang
Stoldt VR, Sonneborn A, Leuker CE & Ernst JF (1997) Efg1p, an Y (2008) Bacterial peptidoglycan triggers Candida albicans
essential regulator of morphogenesis of the human pathogen hyphal growth by directly activating the adenylyl cyclase
Candida albicans, is a member of a conserved class of bHLH Cyr1p. Cell Host Microbe 4: 28–39.
proteins regulating morphogenetic processes in fungi. EMBO J Xue Y, Batlle M & Hirsch J (1998) GPR1 encodes a putative G
16: 1982–1991. protein-coupled receptor that associates with the Gpa2p
Su SS & Mitchell AP (1993) Molecular characterization of the yeast Galpha subunit and functions in a Ras-independent pathway.
meiotic regulatory gene RIM1. Nucleic Acids Res 21: 3789–3797. EMBO J 17: 1996–2007.
Tatebayashi K, Tanaka K, Yang HY, Yamamoto K, Matsushita Y, Yamada-Okabe T, Mio T, Ono N, Kashima Y, Matsui M, Arisawa
Tomida T, Imai M & Saito H (2007) Transmembrane mucins M & Yamada-Okabe H (1999) Roles of three histidine
Hkr1 and Msb2 are putative osmosensors in the SHO1 branch kinase genes in hyphal development and virulence of the
of yeast HOG pathway. EMBO J 26: 3521–3533. pathogenic fungus Candida albicans. J Bacteriol 181:
Townsend PD, Holliday PM, Fenyk S, Hess KC, Gray MA, 7243–7247.
Hodgson DR & Cann MJ (2009) Stimulation of mammalian Zaragoza O, Rodrı́guez C & Gancedo C (2000) Isolation of the
G-protein-responsive adenylyl cyclases by carbon dioxide. J MIG1 gene from Candida albicans and effects of its disruption
Biol Chem 284: 784–791. on catabolite repression. J Bacteriol 182: 320–326.
FEMS Microbiol Lett 295 (2009) 1–9 c2009 Federation of European Microbiological Societies
Published by Blackwell Publishing Ltd. All rights reservedYou can also read

















































