Genome-Based Metabolic Reconstruction Unravels the Key Role of B12 in Methionine Auxotrophy of an Ortho-Phenylphenol-Degrading Sphingomonas ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
ORIGINAL RESEARCH
published: 10 January 2020
doi: 10.3389/fmicb.2019.03009
Genome-Based Metabolic
Reconstruction Unravels the Key
Role of B12 in Methionine
Auxotrophy of an
Ortho-Phenylphenol-Degrading
Sphingomonas haloaromaticamans
Chiara Perruchon † , Sotirios Vasileiadis † , Evangelia S. Papadopoulou and
Dimitrios G. Karpouzas*
Edited by: Laboratory of Plant and Environmental Biotechnology, Department of Biochemistry and Biotechnology, University
Dimitris G. Hatzinikolaou, of Thessaly, Larissa, Greece
National and Kapodistrian University
of Athens, Greece
Reviewed by:
Auxotrophy to amino acids and vitamins is a common feature in the bacterial
Dmitry A. Rodionov, world shaping microbial communities through cross-feeding relations. The amino acid
Sanford Burnham Prebys Medical auxotrophy of pollutant-degrading bacteria could hamper their bioremediation potential,
Discovery Institute, United States
Yujie Men, however, the underlying mechanisms of auxotrophy remain unexplored. We employed
University of California, Riverside, genome sequence-based metabolic reconstruction to identify potential mechanisms
United States
driving the amino acid auxotrophy of a Sphingomonas haloaromaticamans strain
*Correspondence:
Dimitrios G. Karpouzas
degrading the fungicide ortho-phenylphenol (OPP) and provided further verification
dkarpouzas@bio.uth.gr for the identified mechanisms via in vitro bacterial assays. The analysis identified
† These authors have contributed potential gaps in the biosynthesis of isoleucine, phenylalanine and tyrosine, while
equally to this work
methionine biosynthesis was potentially effective, relying though in the presence of
Specialty section:
B12. Supplementation of the bacterium with the four amino acids in all possible
This article was submitted to combinations rescued its degrading capacity only with methionine. Genome sequence-
Systems Microbiology,
based metabolic reconstruction and analysis suggested that the bacterium was
a section of the journal
Frontiers in Microbiology incapable of de novo biosynthesis of B12 (missing genes for the construction of
Received: 14 September 2019 the corrin ring) but carried a complete salvage pathway for corrinoids uptake from
Accepted: 16 December 2019 the environment, transmembrane transportation and biosynthesis of B12. In line with
Published: 10 January 2020
this the bacterium maintained its degrading capacity and growth when supplied with
Citation:
Perruchon C, Vasileiadis S,
environmentally relevant B12 concentrations (i.e., 0.1 ng ml−1 ). Using genome-based
Papadopoulou ES and Karpouzas DG metabolic reconstruction and in vitro testing we unraveled the mechanism driving
(2020) Genome-Based Metabolic
the auxotrophy of a pesticide-degrading S. haloaromaticamans. Further studies will
Reconstruction Unravels the Key Role
of B12 in Methionine Auxotrophy investigate the corrinoids preferences of S. haloaromaticamans for optimum growth and
of an Ortho-Phenylphenol-Degrading OPP degradation.
Sphingomonas haloaromaticamans.
Front. Microbiol. 10:3009. Keywords: Sphingomonas haloaromaticamans, methionine, ortho-phenylphenol, genome-based metabolic
doi: 10.3389/fmicb.2019.03009 reconstruction, B12 auxotrophy
Frontiers in Microbiology | www.frontiersin.org 1 January 2020 | Volume 10 | Article 3009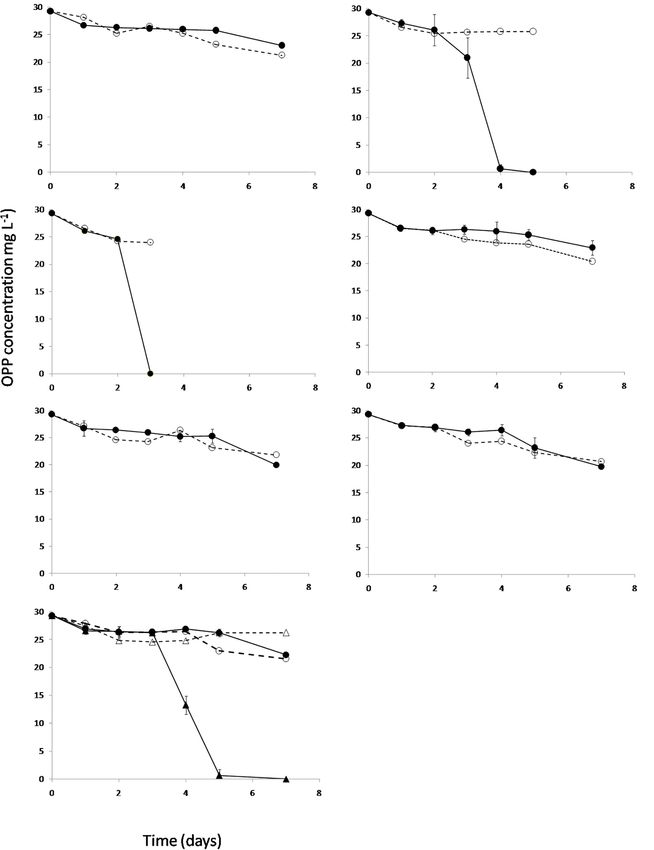
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
INTRODUCTION biosynthesis is the most energetically costly pathway and is
restricted to a number of bacteria and archaea (Roth et al., 1996).
Auxotrophy is the inability of an organism to synthesize A comparative genomic analysis showed that nearly 90% of the
a particular biomolecule which is necessary for its growth. 11,000 bacterial genomes examined had at least one and max 15
Auxotrophic bacterial mutants are known to evolve B12-dependent enzymatic families, yet only 37% carried the full
spontaneously upon long-term growth in an environment B12 biosynthetic pathway (Shelton et al., 2019), reinforcing the
rich in biomolecules leading bacteria to genetic drift and loss key role of B12 in the synergism among microbial populations.
of the corresponding biosynthetic pathways (Van der Kaaij Such B12-based symbiotic relationships have been described
et al., 2014). Amino acid auxotrophy is a common feature of between algae and bacteria (Croft et al., 2005; Romine et al.,
bacterial genomes which have been evolutionary optimized to 2017) or between bacteria (Men et al., 2012) shaping microbial
reduce the metabolic burden stemming from the production of communities in marine water systems (Heal et al., 2017) and the
energetically costly amino acids (i.e., phenylalanine, tyrosine, human gut (Degnan et al., 2014).
and methionine) (Mee et al., 2014). Instead their biosynthesis We recently isolated a Sphingomonas haloaromaticamans
is assigned to a few bacteria leading to the establishment of a strain which was able to degrade the fungicide ortho-
mutualistic trade among bacteria to ensure access to essential phenylphenol (OPP) only when supplemented with CA or
biomolecules (D’Souza et al., 2014; Pande et al., 2015). Vitamin co-cultivated with other bacteria (Perruchon et al., 2016).
auxotrophy constitutes another frequent bacterial phenotype in We aimed to further explore the mechanism driving this
aquatic ecosystems, terrestrial ecosystems, and the human gut. auxotrophy of the S. haloaromaticamans strain to amino acids.
Comparative genomics showed that each of the eight vitamins The hypothesis initially tested was that S. haloaromaticamans
was produced by 45–60% of the gut bacteria suggesting that has one or more incomplete amino acid biosynthesis pathways,
exchange of vitamins amongst human gut bacteria enables the hence the need for external supply of CA. To verify this
survival of members that could not synthesize any of these co- hypothesis, genome-based metabolic reconstruction of the 20
factors (Magnúsdóttir et al., 2015). Similarly, Gomez-Consarnau amino acids biosynthesis in S. haloaromaticamans identified
et al. (2018) showed that vitamins synthesis and exchange possible gaps in the biosynthesis of three amino acids, in addition
by bacteria leads to a mosaic of metabolic interdependencies to methionine whose biosynthetic pathway was complete but
which contribute to the shaping of microbial communities in relied on the de novo biosynthesis of B12. The latter was needed
marine ecosystems. For instance, Rhodobacterales dominated the as a co-factor of methionine synthase since the bacterium did
expression of vitamin-B12 synthesis, but relied on Flavobacteria not carry a B12-indepenedent isofunctional enzyme. In vitro
for the production and supply of vitamin B7. Price et al. assays identified methionine as the amino acid imposing the
(2018) challenged experimentally the value of comparative auxotrophy of S. haloaromaticamans and genome sequence-
genomic studies (i.e., their data showed that auxotrophy in based metabolic reconstruction suggested that the bacterium was
bacteria is not as common as claimed by comparative genomic unable to synthesize B12. Further in vitro studies with a range
studies) and suggested that their findings should be always of B12 concentrations verified its key role in the auxotrophy of
verified experimentally. S. haloaromaticamans, which could cover its B12 requirements
Amino acids auxotrophy has been reported in several probably by uptake and utilization of B12 through the corrinoids
pesticide-degrading bacteria. First, Sørensen et al. (2001) noted salvage pathway present in its genome.
that an isoproturon-degrading Sphingomonas strain SRS2 was
able to maintain its degrading capacity only under external
supplementation with casamino acids (CA); a mixture of all MATERIALS AND METHODS
20 essential amino acids. Follow up studies revealed that
strain SRS2 required external supply of methionine or the Bacterial Strain, Growth Conditions, and
presence of another bacterium in the culture (Sørensen et al., Chemicals
2002), which was probably able to provide to strain SRS2 The strain S. haloaromaticamans used in the current study was
the missing nutrient factors. The need for external supply of isolated from soil of a wastewater disposal site and it was able
amino acids to maintain their degrading activity and growth to degrade the fungicide OPP only when supplemented with CA
on organic pollutants is widespread among sphingomonads (Perruchon et al., 2016). The bacterium was routinely cultivated
with examples of a Sphingomonas xenophaga degrading 1- in minimal salts media supplemented with nitrogen (MSMN),
amino-4-bromoanthraquinone-2-sulfonic acid (Lu et al., 2015), OPP (30–50 mg L−1 ) and CA in a shaking incubator at 27◦ C
a Sphingobium sp. degrading MCPA (Onneby et al., 2014), and in the dark. MSMN preparation and OPP chromatographic
a Sphingomonas sp. degrading triclosan (Hay et al., 2001). This analysis was as described by Perruchon et al. (2016), while
has been assumed to be the result of incomplete amino acid bacterial growth was determined by measurement of the optical
biosynthetic pathways, however, the exact underlying mechanism density at 600 nm (OD600 ). Inoculation of flasks was performed
remains unknown. by fresh bacterial cells grown at the mid-log phase, pelleted
Vitamin B12 (cyanocobalamin) and its methyl and adenosyl by centrifugation, washed three times with sterile ddH2 O and
derivatives, are essential co-factors of several key enzymes like resuspended with MSMN to an OD600 of 0.1.
methionine synthase, ribonucleotide reductase, diol dehydratase, L-methionine, L-isoleucine, L-tyrosine, L-phenylalanine,
and ethanolamine ammonia lyase (Rodionov et al., 2003). Its L-homoserine, O-succinyl-L-homoserine, L-cystathionine,
Frontiers in Microbiology | www.frontiersin.org 2 January 2020 | Volume 10 | Article 3009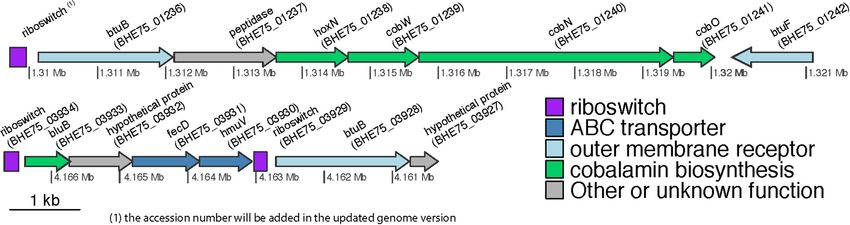
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
L-homocysteine and cyanocobalamin (synonym to vitamin Degradation of Ortho-Phenylphenol by
B12 in the manuscript), were purchased by Sigma-Aldrich Sphingomonas haloaromaticamans
(Taufkirchen, Germany) and they were used for the preparation
of aqueous solutions (0.05 mM). These were filter sterilized Externally Supplied With Amino Acids
and used for the preparation of MSMN containing amino Triplicate 40-ml cultures of MSMN + OPP (50 mg L−1 ) were
acids, B12 and intermediates of methionine biosynthesis at the supplemented with CA (0.15 g L−1 , CA) or methionine (3.6 mg
desired concentrations. L−1 , M), isoleucine (6.75 mg L−1 , I), tyrosine (2.7 mg L−1 ,
T), and phenylalanine (5.7 mg L−1 , P) added individually
and in all possible combinations. These concentrations were
Bioinformatic and Phylogenetic Analysis selected based on the concentrations of the given amino acids
Genome Sequence-Based Metabolic Reconstruction in the CA. Triplicate MSMN + OPP (50 mg L−1 ) samples
of Amino Acids and B12 Biosynthesis in not supplemented with any amino acids were also included for
Sphingomonas haloaromaticamans comparative purposes. All the above cultures were inoculated
Genomic analysis of S. haloaromaticamans has been described with S. haloaromaticamansas described above. Immediately after
by Perruchon et al. (2017) and the assembled and annotated inoculation and at regular intervals thereafter aliquots (0.5 ml)
genome is available at DDBJ/ENA/GenBank under the accession of each culture were removed and analyzed for residues of
number MIPT00000000. The genome annotation data were OPP. The degradation of OPP was also measured in triplicate
used for obtaining the Enzyme Commission (EC) numbers and flasks containing MSMN + OPP (50 mg L−1 ) which were not
reconstruct the biosynthetic pathways of all amino acids and inoculated to serve as abiotic controls.
B12 using the Pathway Tools software v19.0 (Karp et al., 2015)
bundled with the EcoCyc (Karp et al., 2014), BioCyc and MetaCyc Degradation of Ortho-Phenylphenol by
(Caspi et al., 2016) databases. Missing genes were searched Sphingomonas haloaromaticamans
against the genome translated open reading frames (ORFs) after Externally Supplied With B12 and
downloading the associated protein sequences of the RefSeq Methionine Precursors
database of the National Center for Biotechnology Information Triplicate 40-ml cultures of MSMN + OPP (30 mg L−1 ) were
(NCBI) using the basic local alignment search tool (BLAST) supplemented with appropriate amounts of aqueous solutions
v2.7.1+ (Camacho et al., 2009). of 0.05 mM of B12, methionine and its precursors homoserine,
O-succinyl-homoserine, cystathionine and homocysteine. In
B12 Transporters and Riboswitches Identification in the case of homocysteine, the ultimate precursor in the
the Genome of Sphingomonas haloaromaticamans biosynthesis of methionine, three further cultures were co-
The genome of S. haloaromaticamans was searched for supplemented with B12 to evaluate if its addition rescues
TonB-dependent transporters (TBDT) and associated energy the OPP degradation capacity of the bacterium. Triplicate
transducing ABC transporters involved in the uptake and cultures of MSMN + OPP (30 mg L−1 ) not supplemented with
translocation of corrinoids. Beyond the original annotation methionine, B12 or any methionine precursor were also included
described in Perruchon et al. (2017), a second NCBI RefSeq based for comparative purposes. All these cultures were inoculated
annotation was performed with BLAST. We further looked for with S. haloaromaticamans as described above. Immediately
cobalamin-associated riboswitches, acting as regulatory elements after inoculation and at regular intervals thereafter aliquots
of B12 uptake, using the Riboswitch scanner web-based tool (0.5 ml) of each culture were removed and analyzed for
(Mukherjee and Sengupta, 2016). OPP. The degradation of OPP was also measured in triplicate
MSMN + OPP (30 mg L−1 ) not inoculated with the bacterium
to serve as abiotic controls.
Phylogenetic Analysis of MetB/Z
Closely related sequences to MetB/Z found to be encoded Degradation of Ortho-Phenylphenol by
in the genome of S. haloaromaticamans were retrieved from
NCBI with BLASTv2.2.27+ and were clustered with Cdhit
Sphingomonas haloaromaticamans
v4.6 (Li et al., 2001) to reduce sequence redundancy. The Supplied With a Range of B12
sequences were then aligned with Muscle v3.8.31 (Edgar, 2004). Concentrations
Improperly aligned and uninformative alignment blocks were Triplicate 40-ml cultures of MSMN + OPP (25 mg L−1 )
removed using Gblocks v0.91b (Talavera and Castresana, 2007). were supplemented with various amounts of different aqueous
The remaining concatenated alignment blocks were subjected solutions of B12 (1, 100 and 10000 mg L−1 ) aiming to final
to maximum likelihood phylogenies with the RAxML software concentrations of 0.1, 0.5, 1, 5, 10, 50, 100, 1000, and 10000 ng
v8.1.24 (Stamatakis, 2014) and 1000 bootstrap replicates using ml−1 in the medium. Triplicate cultures of MSMN + OPP
the best model according to ProtTest v3.4 (Abascal et al., 2005) (30 mg L−1 ) not supplemented with B12 were also included
and the associated Akaike information criterion values. Tree for comparative purposes. All cultures were inoculated with
visualization was performed using the APE v3.5 (Paradis et al., S. haloaromaticamans as described above. Immediately after
2004) and Phangorn v2.0.4 (Schliep, 2011) R v3.3.118 software inoculation and at regular intervals thereafter aliquots (0.5 ml)
packages (R Core Team, 2015). of each culture were removed and analyzed for OPP. The
Frontiers in Microbiology | www.frontiersin.org 3 January 2020 | Volume 10 | Article 3009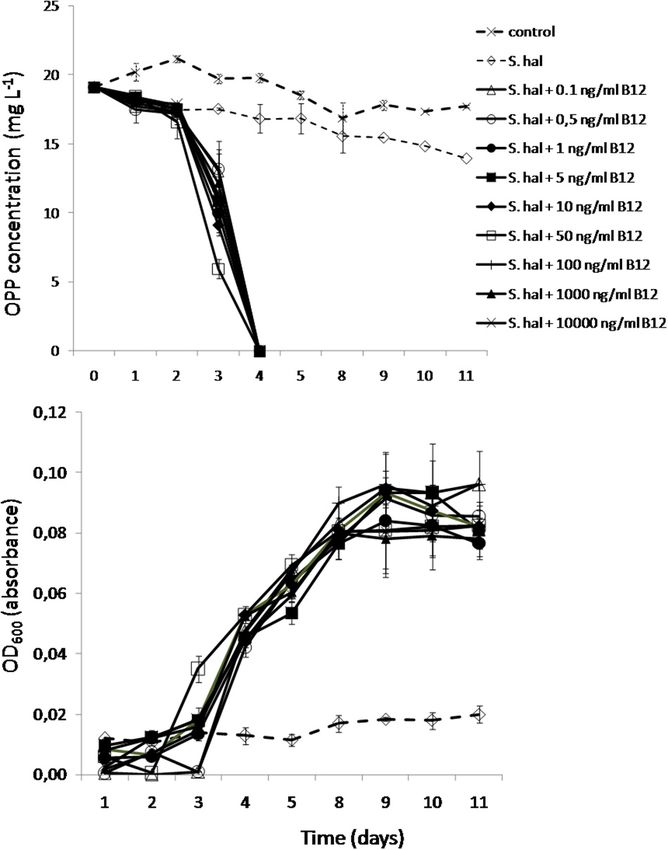
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
degradation of the fungicide was also measured in triplicate for the biosynthesis of tyrosine or phenylalanine (Graindorge
MSMN + OPP which were not inoculated to serve as abiotic et al., 2014). Alternatively, the biosynthesis of tyrosine and
controls. In parallel, we determined the growth of the bacterium phenylalanine proceeds via the transformation of prephenate to
along the degradation of OPP as described above. 4-hydroxyphenyl pyruvate or phenyl pyruvate, which are then
transaminated, with glutamate as amino group donor, to tyrosine
or phenylalanine, respectively (Ikeda, 2006). Genes for these
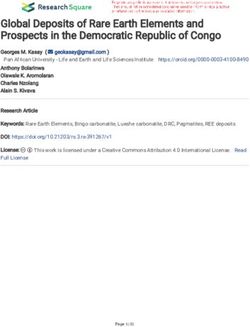
RESULTS AND DISCUSSION pathways were not found in the genome of S. haloaromaticamans.
Instead we identified two copies of the hisC gene encoding
Genome Sequence-Based Metabolic an imidazole acetole phosphate aminotransferase, which has
been reported before in another α-proteobacterium (Zymomonas
Reconstruction of the Biosynthesis of mobilis) to act also as tyrosine transaminase and phenylalanine
Amino Acids in Sphingomonas transaminase (Gu et al., 1995). This might indicate that the
haloaromaticamans tyrosine, phenylalanine biosynthetic pathway is not incomplete in
The first objective of the study was to identify, based on the S. haloaromaticamans, however, both amino acids were included
draft genome of S. haloaromaticamans, biosynthetic pathways in our in vitro tests for completeness.
of amino acids which were potentially incomplete. We noted No gene encoding 2-iminobutanoate deaminase
possible gaps in the biosynthetic pathways of phenylalanine, (EC.3.5.99.10), an enzyme with potential role in the
tyrosine and isoleucine (Figure 1 and Supplementary Table S1). biosynthesis of isoleucine was detected in the draft genome
The biosynthetic pathway of phenylalanine and tyrosine was of S. haloaromaticamans (Figure 1). The transformation of
lacking the enzyme prephenate transaminase (EC 2.6.1.79) threonine to 2-oxobutanoate was originally considered an
(Figure 1). This is commonly found in arogenate-competent one-step process controlled by threonine ammonia lyase (tdcB)
bacteria, like α-proteobacteria, and is responsible for the (Risso et al., 2008). It is now suggested that this metabolic step is
transamination of prephenate to arogenate which is then used a two-step process involving the formation of two intermediates
FIGURE 1 | Reconstruction of the biosynthetic pathways of methionine, isoleucine, tyrosine, and phenylalanine of Sphingomonas haloaromaticamans based on its
genome analysis and assemblage with the Pathway Tools software suit v18.5 (Karp et al., 2015) and the associated database. Gray color arrows indicate the
absence of the enzyme from the draft genome of S. haloaromaticamans. Gene annotation evidence according to BioCyc and RefSeq database hits is provided in
Supplementary Table S1.
Frontiers in Microbiology | www.frontiersin.org 4 January 2020 | Volume 10 | Article 3009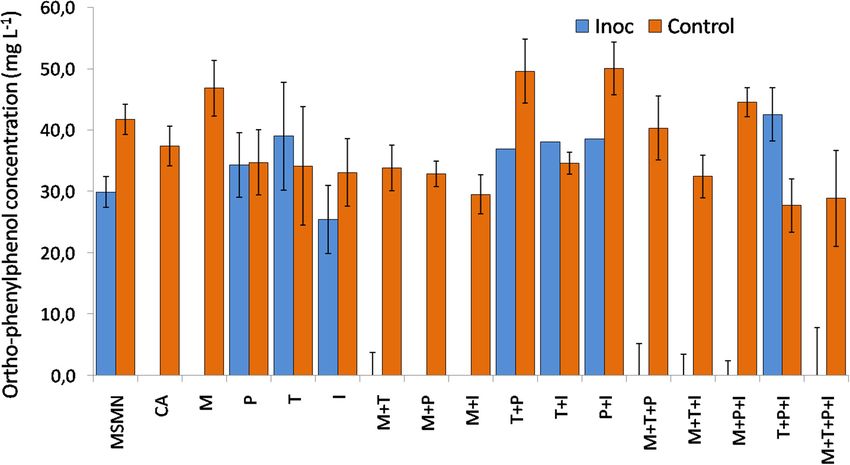
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
[(2Z)-2-aminobut-2-enoate and 2-iminobutanoate] before (Supplementary Figure S1) indicating a probable function
final formation of 2-oxobutanoate. Lambreht et al. (2012) as cystathionine-γ-synthase. The final step of methionine
proposed that 2-iminobutanoate deaminase accelerates the biosynthesis, the transformation of homocysteine to methionine,
transformation of 2-iminobutanoate to 2-oxobutanoate, a is controlled by two isoforms of methionine synthase; a
reaction which could occur spontaneously but its acceleration cobalamin-independent (MetE; 5-methyltetrahydropteroyltri-L-
could protect cells from the reactive intermediates. This indicates glutamate:L-homocysteine S-methyltransferase, EC.2.1.1.14) and
that isoleucine biosynthesis in S. haloaromaticamans might not its cobalamin-dependent isoform (MetH, EC.2.1.1.13) (Gonzalez
be incomplete but it was included in the following in vitro assays et al., 1996), only the latter detected in the genome of S.
for completeness. haloaromaticamans. In most bacteria, metE and metH coexists,
In contrast to the above three amino acids, the genome however, several bacteria possess only one of these two genes
of S. haloaromaticamans carries the necessary genes for the (Shelton et al., 2019). The widespread B12 auxotrophy of algae
biosynthesis of methionine (Figure 1 and Supplementary has been attributed to the evolutionary loss of MetE (Helliwell
Table S1). However, functional annotation of genes in the et al., 2011). Hence, the presence of only metH in the genome
pathway required clarification. We detected metA encoding of S. haloaromaticamans might be a limiting factor in the
homoserine-O-succinyl transferase (EC 2.3.1.46), which drives biosynthesis of methionine, for this reason it was included in our
the transformation of homoserine to O-succinyl-homoserine. in vitro studies.
This is transformed to homocysteine via transsulfuration or
sulfhydrylation, depending on the S source, cysteine, or sulfide,
respectively (Hwang et al., 2002). The former is a two-step process
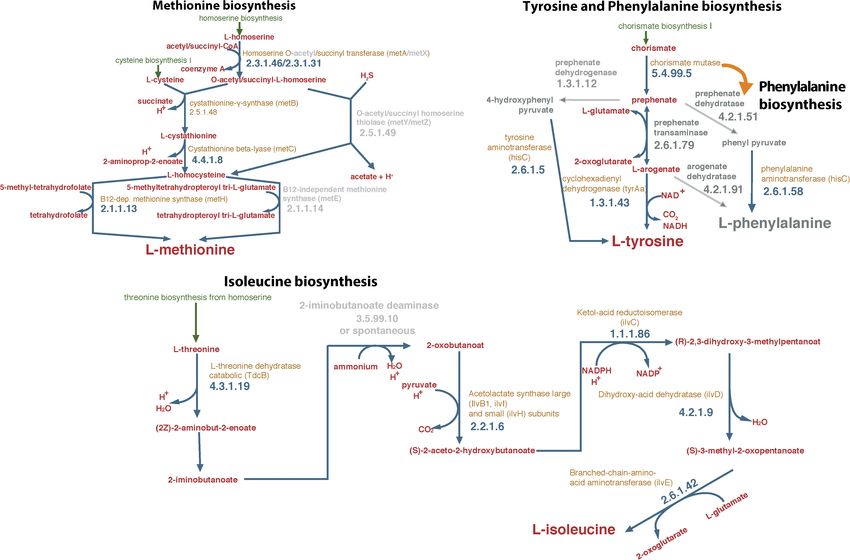
Methionine External Supply Rescues the
involving metB and metC encoding cystathionine-γ-synthase (EC Degradation Phenotype of
2.5.1.48) and cystathionine-β-lyase (EC 4.4.1.8), respectively. The Sphingomonas haloaromaticamans
latter is controlled by O-succinyl homoserine sulfhydrylase (EC Sphingomonas haloaromaticamans was able to effectively degrade
4.2.99.9) encoded by metZ (Ferla and Patrick, 2014). A gene OPP in 7 days in the presence of CA and in all treatments
showing high homology both to metB and metZ was detected where methionine was provided either alone or in combination
in the genome of S. haloaromaticamans. The enzymes encoded with other amino acids (Figure 2). In contrast, limited or no
by these two genes belong to the same cystathionine gamma- degradation of OPP was observed in the non-inoculated cultures
synthase (CGS) evolutionary family (Gophna et al., 2005) and and in the S. haloaromaticamans cultures supplemented with the
could function either as cystathione-γ-synthase or as O-succinyl other three amino acids and their combinations. Methionine is
homoserine sulfhydrylase depending on the source of S (Hacham the less abundant amino acid in bacterial proteins (Pasamontes
et al., 2003). Phylogenetic analysis showed that the relevant and Garcia-Vallve, 2006) but plays an important role in the
gene clustered with metB genes of various Sphingomonas strains initiation of translation, has a structural role in hydrophobic
FIGURE 2 | Degradation of OPP 48 h after inoculation (Inoc) with S. haloaromaticamans in duplicate cultures of a selective minimal medium (MSMN) supplemented
with CA (0.15 g L−1 , CA) or methionine (3.6 mg L−1 , M), isoleucine (6.75 mg L−1 , I), tyrosine (2.7 mg L−1 , T), and phenylalanine (5.7 mg L−1 , P) added individually
and in all possible combinations. The degradation of OPP was also determined in duplicate inoculated cultures of MSMN without any amino acid supplementation
(MSMN) and in corresponding non-inoculated controls (Control).
Frontiers in Microbiology | www.frontiersin.org 5 January 2020 | Volume 10 | Article 3009Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans FIGURE 3 | Reconstruction of the biosynthetic pathway of B12 in S. haloaromaticamans based on its genome analysis and assemblage with the Pathway Tools software suit v18.5 (Karp et al., 2015). Enzyme names indicated with black or gray color were either detected or not detected in the genome of S. haloaromaticamans, respectively. Dashed arrows denote the steps of the cobalamin salvage pathway. Gene annotation evidence according to BioCyc and RefSeq database hits is provided in Supplementary Table S2. Frontiers in Microbiology | www.frontiersin.org 6 January 2020 | Volume 10 | Article 3009
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
cores of proteins, is involved in stabilizing interactions with the TonB/ExpB/ExpD complex components (inner membrane
aromatic amino acids in 1/3 of all known protein structures transportation system), 21 genes coded for genes with a potential
(Valley et al., 2012), and it is the key component of the cofactor role in the regulatory system of the TonB-dependent uptake
S-adenosyl-methionine which constitutes the main cellular (20 comprising the FecI/FecRand one being the iron uptake
carrier of methyl groups (Chiang et al., 1996). Our findings repressor “fur” gene), and 4 genes were screened due to either
are in agreement with previous studies which have reported their vicinity to TBDT or potential association (due to their
an auxotrophy of other pesticide-degrading sphingomonads to annotation or their content) with riboswitches (Supplementary
methionine (Sørensen et al., 2002; Onneby et al., 2014). However, Table S3). Sphingomonadales genomes are known to be rich
none of the above studies looked further into the mechanism in TBDT [i.e., Sphingomonas wittichii RW1 carried 134 TBDTs
driving the reported auxotrophy. In light of the genome-based (Tang et al., 2012)], in line with their high abundance in the
metabolic analysis of the methionine pathway the observed genome of S. haloaromaticamans. From the 43 TonB-dependent
auxotrophy of S. haloaromaticamans to methionine could be the receptors (i) 16 were annotated as putative B12 outer membrane
result of the absence of a complete B12 biosynthetic pathway in receptors (BtuB) (Chimento et al., 2003), showing best sequence
the studied bacterium. matches to the previously published S. haloaromaticamans strain
A175 DSM13477 genome (Wittich et al., 2007) (ii) seven were
Genome Sequence-Based Metabolic annotated as colicin receptors (CirA) (Ferguson et al., 2002),
and (iii) the rest as ferripyoverdine (FpvA), ferric pseudobactin
Reconstruction of the Biosynthesis of (PupA) (Adams et al., 2006), ferrichrome iron (FhuA) (Locher
B12 in Sphingomonas et al., 1998) and various siderophore receptors (Noinaj et al.,
haloaromaticamans 2010). Considering the very close structures of the several TBDT,
As a next step we reconstructed the biosynthetic pathway of B12 including B12/colicin/siderophore transporters, their sequence-
in S. haloaromaticamans to investigate its potency to synthesize based substrate determination should be considered with caution
the co-factor. The bacterial genome lacked all the genes for (Koster, 2001; Rodionov et al., 2003).
the biosynthesis of the corrin ring of B12 (cobIZGJMFKLH) From these TonB-dependent receptors only three fulfill the
(Figure 3 and Supplementary Table S2). Instead it carries nearly stringent criterium of Degnan et al. (2014) in order to be assigned
all genes of the lower pathway (cobABNSTWOCQDPUVCZ) a cobalamin transporter function, “to be located downstream of
from the conversion of hydrogenobyrinate to its a,c-diamide riboswitches.” These are RNA elements that change conformation
derivative through Co2+ chelation to adenosyl-cobalamin upon binding of a specific molecule affecting negatively or
(Raux et al., 2000). positively the transcription and translation of downstream
The absence of a de novo biosynthetic pathway of B12 in genes including B12 uptake and biosynthesis (Montange and
S. haloaromaticamans genome and its demonstrated absolute Batey, 2008). In silico screening of the genome for regulatory
requirement for B12-dependent methionine synthesis, led riboswitches identified three of them residing in the 50 region
us to speculate that the bacterium should possess a high of TBDT (Figure 4). The first (position of putative riboswitch:
affinity system for uptake and translocation of corrinoids. The 1309712-1309955) was located upstream of a region encoding
bacterial genome was searched for genes encoding for energy- BtuB, a protein of the HoxN family known to encode high
dependent uptake of B12. We detected 88 genes with potential affinity Co+2 /Ni+2 transporter genes (Komeda et al., 1997),
role in the energy-dependent uptake of various biomolecules cobW, cobN, and cobO, respectively, encoding proteins involved
including B12: 43 were annotated to code for TonB-dependent in Co chelation, cobalt chelatase subunit N and a corrinoid
receptors (outer membrane transportation system), 20 coded for adenosyltransferase (Rodionov et al., 2003), all involved in the
FIGURE 4 | Genetic organization of the genomic regions in S. haloaromaticamans downstream of the three riboswitches associated with B12-biosynthesis. Genes
annotation btuB: TonB-dependent corrinoid transporter; hoxN, high affinity Co+2 /Ni+2 transporter; cobW, cobalt-chelation associated protein; cobN, cobalt
chelatase subunit N; cobO, corrinoid adenosyltransferase; btuF, substrate-binding component of the ABC cobalamin transporter system; bluB,
5,6-dimethylbenzimidazole synthase; fecD, permease of a btuCD-like ABC cobalamin transporter system; hmuV, ATP-binding subunit of a butCD-like ABC
cobalamin transporter system.
Frontiers in Microbiology | www.frontiersin.org 7 January 2020 | Volume 10 | Article 3009Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans FIGURE 5 | The degradation of OPP by S. haloaromaticamans in MSMN (•, solid lines) without any supplementation (A) and with supplementation of methionine (B), B12 (C), and of various intermediates of the methionine biosynthetic pathway like homoserine (D), O-succinyl homoserine (E), cystathionine (F), and homocysteine [plus (N) or minus B12 (•)] (G). Each value is the mean of triplicates ± the standard deviation of the mean. In all treatments the degradation of OPP in non-inoculated cultures was also determined (◦, 4 dashed lines). Frontiers in Microbiology | www.frontiersin.org 8 January 2020 | Volume 10 | Article 3009
Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
biosynthesis of B12, and BtuF, the substrate-binding component
of the ABC transporter system for B12 uptake. Such a gene
organization has been reported in several proteobacteria where
hoxN and cobW are located immediately upstream of cobN
and the whole locus is flanked upstream with a riboswitch
(Rodionov et al., 2003). The second (position 4162841-4163031)
and the third riboswitches (position 4166483-4166691) were
closely localized. The former is located upstream of bluB, a
hypothetical protein and fecD (e-value1e-718, 78% identities and
86% positives with the Sphingopyxis granuli btuC, AMG76476.1),
hmuV (e-value8e-97, 66% identities and 79% positives with the
Sphingopyxis sp. strain C-1 btuD, GAO77867.1) genes encoding,
respectively, a permease and an ATP-binding protein forming
a btuCD-like ABC transporter component of the cobalamin
transportation system. BluB belongs to the flavin destructase
family proteins and controls the transformation of FMN2 to 5,6-
dimethylbenzimidazole (Taga et al., 2007), which is the lower
axial ligand of B12, and the preferred ligand in the cobamides
of most bacteria (Hazra et al., 2013). Considering the key role of
bluB in the biosynthesis or remodeling of salvaged corrinoids by
bacteria it is not surprising that its translation might be regulated
by B12-associated riboswitches. The third riboswitch is located in
the 50 region of btuB. In all other cases no riboswitches or other
regulatory systems were detected. This could be due to missing
alignment profiles from the databases which are necessary
for identifying riboswitches in sphingomonads, or, given that
cobalamin is a well-defined structure, maybe riboswitches do not
exist as a regulatory mechanism. In the latter case, a different FIGURE 6 | The degradation of OPP (A) and the growth of
regulatory system or even a housekeeping role of the associated S. haloaromaticamans (B) in MSMN supplemented with 0, 0.1, 0.5, 1, 5, 10,
50, 100, 1000, and 10000 ng ml−1 of B12. The degradation of OPP in
transporters could be possible.
non-inoculated controls was also determined (control). Each value is the mean
Overall, our in silico analysis of the biosynthetic pathway of triplicates ± the standard deviation.
of B12 suggests that S. haloaromaticamans is not able to
biosynthesize B12 and meet its cellular needs for the methionine
biosynthesis. However, its genome is enriched in receptors
for transportation of corrinoids across its outer membrane,
The Degradation and Growth of
operating the energetically inexpensive salvage pathway to serve Sphingomonas haloaromaticamans
its needs in the primary cellular metabolism. Supplied With a Range of B12
Concentrations
B12 and Not Methionine Precursors We finally tested the minimum concentrations of B12 provided
to S. haloaromaticamans to avert its auxotrophy to methionine
Rescues the Degradation Capacity of and degrade OPP. The bacterium was able to degrade OPP even
Sphingomonas haloaromaticamans when supplied with 0.1 ng ml−1 of B12 (Figure 6A), which is
Based on the genome sequence-based metabolic reconstruction close to the concentration levels of B12 and corrinoids found
evidence which suggested that S. haloaromaticamans was not in fresh water (2–6 ng L−1 ) (Kurata, 1986), soil solution (5 µg
capable of de novo biosynthesis of B12, we evaluated in vitro L−1 ) (Mozafar and Oertli, 1992), and wastewaters (0.05–5.2 mg
whether the supplementation of methionine precursors L−1 ) (Selimoglu et al., 2015). In addition, all concentrations
and/or B12 will rescue the degradation capacity of S. of B12 supported similar growth rates of S. haloaromaticamans
haloaromaticamans. The bacterium maintained its degrading (Figure 6B). As expected, no growth and degradation of OPP was
capacity against OPP when supplemented with methionine observed when the bacterium was not supplemented with B12.
(Figure 5B) and B12 (Figure 5C). However, it failed to
degrade OPP when it was not supplemented with any of
the above (Figure 5A) or supplemented with intermediates CONCLUSION
of methionine biosynthesis (Figures 5D–G) unless B12 was
also co-provided as in the case of homocysteine (Figure 5G). A combination of genome-based metabolic reconstruction and
These results verified genome-based metabolic reconstruction in vitro tests demonstrated that the auxotrophy of a fungicide-
findings that B12 is the main driver of the auxotrophy of degrading S. haloaromaticamans strain was driven by the lack of
S. haloaromaticamans to methionine. a MetE B12-independent cobalamine synthase and the absence
Frontiers in Microbiology | www.frontiersin.org 9 January 2020 | Volume 10 | Article 3009Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
of a de novo B12 biosynthetic pathway. The bacterium could Action of the “Operational Programme Education and Lifelong
maintain its degradation and growth capacity in the presence Learning” and co-funded by the European Social Fund
of environmentally relevant concentrations of B12 suggesting (ESF) and National Resources, Greece, and (ii) “OMIC-
that its application in natural and engineered systems would ENGINE-Synthetic Biology: From Omics Technologies to
not be hampered by its auxotrophy to B12, in line with the Genomic Engineering” (MIS 5002636) which is implemented
presence in its genome of several potential B12 outer membrane under the Action “Reinforcement of the Research and
receptors and a near complete cobalamine salvage pathway. Innovation Infrastructure,” funded by the Operational Program
Further studies will focus on (a) the corrinoids preferences of S. “Competitiveness, Entrepreneurship and Innovation” (NSRF
haloaromaticamans for optimum growth and OPP degradation 2014-2020) and co-financed by Greece and the European
using cross-feeding studies with corrinoid-producing bacteria Union (European Regional Development Fund). SV was
(Mesorhizobium loti) and external supply of selected corrinoids supported by the MSCA-IF-H2020 project EMIGRATE (Grant
(b) the role of the riboswitches detected on the regulation of Agreement No. 749463).
cobalamin biosynthesis and corrinoids uptake.
SUPPLEMENTARY MATERIAL
DATA AVAILABILITY STATEMENT
The Supplementary Material for this article can be found
The datasets generated for this study can be found in the online at: https://www.frontiersin.org/articles/10.3389/fmicb.
DDBJ/ENA/GenBank accession number MIPT00000000. 2019.03009/full#supplementary-material
FIGURE S1 | Phylogenetic analysis of the metB/Z gene encoding cystathione
AUTHOR CONTRIBUTIONS γ-synthase/O-succinyl homoserine sulfhydrylase detected in the genome of
Sphingomonas haloaromaticamans.
CP isolated the microorganism, performed the in vitro TABLE S1 | The functional annotation of the genes participating the biosynthetic
experiment, and drafted the manuscript. SV performed the pathways of methionine, isoleucine, tyrosine, and phenylalanine in the genome of
bioinformatic analysis and genome-based reconstruction, and the Sphingomonas haloaromaticamans strain. Annotation was performed using
helped in drafting of the manuscript. EP performed the UniProt and RefSeq BLAST searches and verified further through Pfam searches.
in vitro experiment and reviewed the manuscript. DK had the TABLE S2 | The functional annotation of the genes participating in cobalamin
experimental idea, supervised and planned the experiments, and biosynthetic pathways in the Sphingomonas haloaromaticamans strain.
revised the final manuscript. Annotation was performed using UniProt and RefSeq BLAST searches and
verified further through Pfam searches.
TABLE S3 | The functional annotation of the genes associated directly or indirectly
FUNDING with the energy-dependent uptake of various biomolecules (i.e., cobinamides,
siderophores, colicin etc.). Annotation was performed using UniProt and RefSeq
This work was funded by the projects (i) “Bioremediatomics – BLAST searches and verified further through Pfam searches. Purple color denotes
genes with TonB-dependent transporter function, orange color denotes genes
The Microbial Detoxification of Pesticides From the Fruit- with energy transducing role (ABC transporters), blue color denotes genes with
Packaging Industry: Using Omics in Bioremediation” (Grant regulatory function in B12 transportation and gray color denotes other genes with
No. 4781) which is implemented under the “ARISTEIA” indirect association with B12 biosynthesis or transportation.
REFERENCES Croft, M. T., Lawrence, A. D., Raux-Deery, E., Warren, M. J., and Smith, A. G.
(2005). Algae acquire vitamin B12 through a symbiotic relationship with
Abascal, F., Zardoya, R., and Posada, D. (2005). ProtTest: selection of best-fit bacteria. Nature 438, 90–93. doi: 10.1038/nature04056
models of protein evolution. Bioinformatics 21, 2104–2105. doi: 10.1093/ Degnan, P. H., Barry, N. A., Mok, K. C., Taga, M. E., and Goodman, A. L. (2014).
bioinformatics/bti263 Human gut microbes use multiple transporters to distinguish vitamin B12
Adams, H., Zeder-Lutz, G., Schalk, I., Pattus, F., and Celia, H. (2006). Interaction analogs and compete in the gut. Cell Host Microbe 15, 47–57. doi: 10.1016/j.
of TonB with the outer membrane receptor FpvA of Pseudomonas aeruginosa. chom.2013.12.007
J. Bacteriol. 188, 5752–5761. doi: 10.1128/jb.00435-06 D’Souza, G., Waschina, S., Pande, S., Bohl, K., Kaleta, C., and Kost, C. (2014). Less
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., is more: selective advantages can explain the prevalent loss of biosynthetic genes
et al. (2009). BLAST+: architecture and applications. BMC Bioinf. 10:421. doi: in bacteria. Evolution 68, 2559–2570. doi: 10.1111/evo.12468
10.1186/1471-2105-10-421 Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy
Caspi, R., Billington, R., Ferrer, L., Foerster, H., Fulcher, C. A., Keseler, I. M., and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gk
et al. (2016). The MetaCyc database of metabolic pathways and enzymes and h340
the BioCyc collection of pathway/genome databases. Nucleic Acids Res. 44, Ferguson, A. D., Chakraborty, R., Smith, B. S., Esser, L., van der Helm, D.,
D471–D480. doi: 10.1093/nar/gkv1164 and Deisenhofer, J. (2002). Structural basis of gating by the outer membrane
Chiang, P. K., Gordon, R. K., Tal, J., Zeng, G. C., Doctor, B. P., Pardhasaradhi, K., transporter FecA. Science 295, 1715–1719. doi: 10.1126/science.1067313
et al. (1996). S-Adenosylmethionine and methylation. FASEB J. 10, 471–480. Ferla, M. P., and Patrick, W. M. (2014). Bacterial methionine biosynthesis.
Chimento, D. P., Mohanty, A. K., Kadner, R. J., and Wiener, M. C. (2003). Microbiology 160, 1571–1584. doi: 10.1099/mic.0.077826-0
Substrate-induced transmembrane signaling in the cobalamin transporter Gomez-Consarnau, L., Sachdeva, R., Gifford, S. M., Cutter, L. S., Fuhrman, J. A.,
BtuB. Nat. Struct. Biol. 10, 394–401. doi: 10.1038/nsb914 Sanudo-Wilhelmy, S. A., et al. (2018). Mosaic patterns of B-vitamin synthesis
Frontiers in Microbiology | www.frontiersin.org 10 January 2020 | Volume 10 | Article 3009Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
and utilization in a natural marine microbial community. Environ. Microbiol. Lu, H., Guan, X., Wang, J., Zhou, J., and Zhang, H. (2015). Enhanced
20, 2809–2823. doi: 10.1111/1462-2920.14133 bio-decolorization of 1-amino-4-bromoanthraquinone-2-sulfonic acid by
Gonzalez, J. C., Peariso, K., Penner-Hahn, J. E., and Matthews, R. G. (1996). Sphingomonas xenophaga with nutrient amendment. J. Environ. Sci. 27, 124–
Cobalamin-independent methionine synthase from Escherichia coli: a zinc 130. doi: 10.1016/j.jes.2014.05.041
metalloenzyme. Biochemistry 35, 12228–12234. doi: 10.1021/bi9615452 Magnúsdóttir, S., Ravcheev, D., de Crécy-Lagard, V., and Thiele, I. (2015).
Gophna, U., Bapteste, E., Doolittle, W. F., Biran, D., and Ron, E. Z. (2005). Systematic genome assessment of B-vitamin biosynthesis suggests co-operation
Evolutionary plasticity of methionine biosynthesis. Gene 355, 48–57. doi: 10. amonggut microbes. Front. Genet. 6:148. doi: 10.3389/fgene.2015.00148
1016/j.gene.2005.05.028 Mee, M. T., Collins, J. J., Church, G. M., and Wang, H. H. (2014). Syntrophic
Graindorge, M., Guistini, C., Kraut, A., Moyet, L., Curien, G., and Matringe, exchange in synthetic microbial communities. Proc. Natl. Acad. Sci. 111, E2149–
M. (2014). Three different classes of aminotransferases evolved prephenate E2156. doi: 10.1073/pnas.1405641111
aminotransferase functionality in arogenate-competent microorganisms. Men, Y., Feil, H., VerBerkmoes, N. C., Shah, M. B., Johnson, D. R., Lee, P. K. H.,
J. Biol. Chem. 289, 3198–3208. doi: 10.1074/jbc.M113.486480 et al. (2012). Sustainable syntrophic growth of Dehalococcoides ethenogenes
Gu, W., Zhao, G., Eddy, C., and Jensen, R. A. (1995). Imidazole acetol phosphate strain 195 with Desulfovibrio vulgaris Hildenborough and Methanobacterium
aminotransferase in Zymomonas mobilis: molecular genetic, biochemical, and congolense: global transcriptomic and proteomic analyses. ISME J. 6, 410–421.
evolutionary analyses. J. Bacteriol. 177, 1576–1584. doi: 10.1128/jb.177.6.1576- doi: 10.1038/ismej.2011.111
1584.1995 Montange, R. K., and Batey, R. T. (2008). Riboswitches: emerging themes in RNA
Hacham, Y., Gophna, U., and Amir, R. (2003). In vivo analysis of various substrates structure and function. Annu. Rev. Biophys. 37, 117–133. doi: 10.1146/annurev.
utilized by cystathionine γ-synthase and O-acetylhomoserine sulfhydrylase biophys.37.032807.130000
in methionine biosynthesis. Molec. Biol. Evol. 20, 1513–1520. doi: 10.1093/ Mozafar, A., and Oertli, J. J. (1992). Uptake of microbially -produced vitamin (B12)
molbev/msg169 by soybean roots. Plant Soil 139, 23–30. doi: 10.1007/bf00012838
Hay, A. G., Dees, P. M., and Sayler, G. M. (2001). Growth of a bacterial consortium Mukherjee, S., and Sengupta, S. (2016). Riboswitch Scanner: an efficient pHMM-
on triclosan. FEMS Microbiol. Ecol. 36, 105–112. doi: 10.1016/s0168-6496(01) based web-server to detect riboswitches in genomic sequences. Bioinformatics
00127-1 32, 776–778. doi: 10.1093/bioinformatics/btv640
Hazra, A. B., Tran, J. L. A., Crofts, T. S., and Taga, M. E. (2013). Analysis of substrate Noinaj, N., Guillier, M., Barnard, T. J., and Buchanan, S. K. (2010). TonB-
specificity in CobT homologs reveals widespread preference for DMB, the lower dependent transporters: regulation, structure, and function. Annu. Rev.
axial ligand of vitamin B12. Chem. Biol. 20, 1275–1285. doi: 10.1016/j.chembiol. Microbiol. 64, 43–60. doi: 10.1146/annurev.micro.112408.134247
2013.08.007 Onneby, K., Hakansson, S., Pizzul, L., and Stenstrom, J. (2014). Reduced
Heal, K. R., Qin, W., Ribalet, F., Bertagnolli, A. D., Coyote-Maestras, W., Hmelo, leaching of the herbicide MCPA after bioaugmentation with a formulated and
L. R., et al. (2017). Two distinct pools of B12 analogs reveal community stored Sphingobium sp. Biodegradation 25, 291–300. doi: 10.1007/s10532-013-
interdependencies in the ocean. PNAS 114, 364–369. doi: 10.1073/pnas. 9660-3
1608462114 Pande, S., Shitut, S., Freund, L., Westermann, M., Bertels, F., Colesie, C., et al.
Helliwell, K. E., Wheeler, G. L., Leptos, K. C., Goldstein, R. E., and Smith, A. G. (2015). Metabolic cross-feeding via intercellular nanotubes among bacteria.
(2011). Insights into the evolution of vitamin B12 auxotrophy from sequenced Nature Commun. 6, 6238–6251. doi: 10.1038/ncomms7238
algal genomes. Mol. Biol. Evol. 28, 2921–2933. doi: 10.1093/molbev/ms Paradis, E., Claude, J., and Strimmer, K. (2004). APE: analyses of phylogenetics
r124 and evolution in R language. Bioinformatics 20, 289–290. doi: 10.1093/
Hwang, B.-J., Yeom, H.-I., Kim, Y., and Lee, H.-S. (2002). Corynebacterium bioinformatics/btg412
glutamicum utilizes both trans-sulfuration and direct sulfhydrylation pathways Pasamontes, A., and Garcia-Vallve, S. (2006). Use of a multi-way method to analyze
for methionine biosynthesis. J. Bacteriol. 184, 1277–1286. doi: 10.1128/jb.184. the amino acid composition of a conserved group of orthologous proteins in
5.1277-1286.2002 prokaryotes. BMC Bioinf. 7:257. doi: 10.1186/1471-2105-7-257
Ikeda, M. (2006). Towards bacterial strains overproducing L-tryptophan and other Perruchon, C., Patsioura, V., Vasileiadis, S., and Karpouzas, D. G. (2016).
aromatics by metabolic engineering. Appl. Microbiol. Biotechn. 69, 615–626. Isolation and characterisation of a Sphingomonas strain able to degrade the
doi: 10.1007/s00253-005-0252-y fungicide ortho-phenylphenol. Pest Manag. Sci. 72, 113–124. doi: 10.1002/
Karp, P., Weaver, D., Paley, S., Fulcher, C., Kubo, A., Kothari, A., et al. (2014). The ps.3970
EcoCyc Database. EcoSal Plus. 41, D605–D612. doi: 10.1128/ecosalplus.ESP- Perruchon, C., Vasileiadis, S., Rousidou, C., Papadopoulou, E. S., Tanou,
0009-2013 G., Samiotaki, M., et al. (2017). Metabolic pathway and cell adaptation
Karp, P. D., Latendresse, M., Paley, S. M., Krummenacker, M., Ong, Q. D., mechanisms revealed through genomic, proteomic and transcription analysis
Billington, R., et al. (2015). Pathway Tools version 19.0 update: software for of a Sphingomonas haloaromaticamans strain degrading ortho-phenylphenol.
pathway/genome informatics and systems biology. Brief Bioinform. 17, 877– Sci. Rep. 7:6449. doi: 10.1038/s41598-017-06727-6
890. doi: 10.1093/bib/bbv079.2015 Price, M. N., Zane, G. M., Kuehl, J. V., Melnyk, R. A., Wall, J. D., Deutschbauer,
Komeda, H., Kobayashi, M., and Shimizu, S. (1997). A novel transporter involved A. M., et al. (2018). Filling gaps in bacterial amino acid biosynthesis pathways
in cobalt uptake. Proc. Natl. Acad. Sci. U.S.A. 94, 36–41. doi: 10.1073/pnas.94. with high-throughput genetics. PLoS Genet. 14:e1007147. doi: 10.1371/journal.
1.36 pgen.1007147
Koster, W. (2001). ABC transporter-mediated uptake of iron, siderophores, R Core Team. (2015). R: A Language and Environment for Statistical Computing,
heme and vitamin B12. Res. Microbiol. 152, 291–301. doi: 10.1016/s0923- Reference Index Version 3.2.2. 2015. Vienna: R Core Team.
2508(01)01200-1 Raux, E., Schubert, H. L., and Warren, M. J. (2000). Biosynthesis of cobalamin
Kurata, A. (1986). Chrysophytes: Aspects and Problems, eds J. Kristiansen, and R. A. (vitamin B12): a bacterial conundrum. Cell. Moll. Life Sci. 57, 1880–1893. doi:
Andersen, (Cambridge: Cambridge Univ. Press), 185–196. 10.1007/pl00000670
Lambreht, J. A., Flynn, J. M., and Downs, D. M. (2012). Conserved YjgF Risso, C., Van Dien, S. J., Orloff, A., Lovley, D. R., and Coppi, M. V. (2008).
protein family deaminates reactive enamine/imine intermediates of pyridoxal Elucidation of an alternative isoleucine biosynthesis pathway in Geobacter
5-phosphate (PLP)-dependent enzyme reactions. J. Biol. Chem. 287, 3454–3461. sulforreducens. J. Bacteriol. 190, 2266–2274. doi: 10.1128/jb.01841-07
doi: 10.1074/jbc.M111.304477 Rodionov, D., Vitreschak, A. G., Mironov, A. A., and Gelfand, M. S. (2003).
Li, W., Jaroszewski, L., and Godzik, A. (2001). Clustering of highly homologous Comparative genomics of the vitamin B12 metabolism and regulation in
sequences to reduce the size of large protein databases. Bioinformatics 17, prokaryotes. J. Biol. Chem. 278, 41148–41159. doi: 10.1074/jbc.m30583
282–283. doi: 10.1093/bioinformatics/17.3.282 7200
Locher, K. P., Rees, B., Koebnik, R., Mitschler, A., Moulinier, L., Rosenbusch, J. P., Romine, M. F., Rodionov, D. A., Maezato, Y., Osterman, A. L., and Nelson, W. C.
et al. (1998). Transmembrane signaling across the ligand-gated FhuA receptor: (2017). Underlying mechanisms for syntrophic metabolism of essential enzyme
crystal structures of free and ferrichrome-bound states reveal allosteric changes. cofactors in microbial communities. ISME J. 11, 1434–1446. doi: 10.1038/ismej.
Cell 95, 771–778. doi: 10.1016/s0092-8674(00)81700-6 2017.2
Frontiers in Microbiology | www.frontiersin.org 11 January 2020 | Volume 10 | Article 3009Perruchon et al. B12 Auxotrophy Mechanism of S. haloaromaticamans
Roth, J. R., Lawrence, J. G., and Bobik, T. A. (1996). Cobalamin (coenzyme B12): Tang, K., Jiao, N., Liu, K., Zhang, Y., and Li, S. (2012). Distribution and functions
synthesis and biological significance. Annu. Rev. Microbiol. 50, 137–181. doi: of TonB-dependent transporters in marine bacteria and environments:
10.1146/annurev.micro.50.1.137 implications for dissolved organic matter utilization. PLoS One 7:e41204. doi:
Schliep, K. P. (2011). Phangorn: phylogenetic analysis in R. Bioinformatics 27, 10.1371/journal.pone.0041204
592–593. doi: 10.1093/bioinformatics/btq706 Valley, C. C., Cembran, A., Perlmutter, J. D., Lewis, A. K., Labello, N. P., Gao, J.,
Selimoglu, F., Obek, E., Karatas, F., Arslan, E. I., and Tatar, S. Y. (2015). et al. (2012). The methionine-aromatic motif plays a unique role in stabilizing
Determination of amounts of some vitamin B groups indomestic wastewater protein structure. J. Biol. Chem. 287, 34979–34991. doi: 10.1074/jbc.M112.
treatment plants. Turk. J. Sci. Technol. 10, 1–5. 374504
Shelton, A., Seth, E. C., Mok, K. C., Han, A. W., Jackson, S. N., Haft, D. R., Van der Kaaij, H., Desiere, F., Mollet, B., and Germond, J.-E., (2014). L-Alanine
et al. (2019). Uneven distribution of cobamide biosynthesis and dependence in auxotrophy of Lactobacillus johnsonii as demonstrated by physiological,
bacteria predicted by comparative genomics. ISME J. 13, 789–804. doi: 10.1038/ genomic, and gene complementation approaches. Appl. Environ. Microbiol. 70,
s41396-018-0304-9 1869–1873. doi: 10.1128/aem.70.3.1869-1873.2004
Sørensen, S. R., Ronen, Z., and Aamand, J. (2001). Isolation from agricultural soil Wittich, R.-M., Busse, H.-J., Kämpfer, P., Macedo, A. J., Tiirola, M., Wieser,
and characterization of a Sphingomonas sp. able to mineralize the phenylurea M., et al. (2007). Sphingomonas fennica sp. nov. and Sphingomonas
herbicide isoproturon. Appl. Environ. Microbiol. 67, 5403–5409. doi: 10.1128/ haloaromaticamans sp. nov., outliers of the genus Sphingomonas.
aem.67.12.5403-5409.2001 Int. J. Syst. Evol. Microbiol. 57, 1740–1746. doi: 10.1099/ijs.0.64
Sørensen, S. R., Ronen, Z., and Aamand, J. (2002). Growth in co-culture stimulates 835-0
metabolism of the phenylurea herbicide isoproturon by Sphingomonas sp strain
SRS2. Appl. Environ. Microbiol. 68, 3478–3485. doi: 10.1128/aem.68.7.3478- Conflict of Interest: The authors declare that the research was conducted in the
3485.2002 absence of any commercial or financial relationships that could be construed as a
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and potential conflict of interest.
post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/
bioinformatics/btu033 Copyright © 2020 Perruchon, Vasileiadis, Papadopoulou and Karpouzas. This is an
Taga, M. E., Larsen, N. A., Howard-Jones, A. R., Walsh, C. T., and Walker, G. C. open-access article distributed under the terms of the Creative Commons Attribution
(2007). BluB canibalizes flavin to form the lower ligand of vitamin B12. Nature License (CC BY). The use, distribution or reproduction in other forums is permitted,
446, 449–453. doi: 10.1038/nature05611 provided the original author(s) and the copyright owner(s) are credited and that the
Talavera, G., and Castresana, J. (2007). Improvement of phylogenies after removing original publication in this journal is cited, in accordance with accepted academic
divergent and ambiguously aligned blocks from protein sequence alignments. practice. No use, distribution or reproduction is permitted which does not comply
Syst. Biol. 56, 564–577. doi: 10.1080/10635150701472164 with these terms.
Frontiers in Microbiology | www.frontiersin.org 12 January 2020 | Volume 10 | Article 3009You can also read