Targeted therapy for adrenocortical tumors in transgenic mice through their LH receptor by Hecate-human chorionic gonadotropin b conjugate
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Endocrine-Related Cancer (2008) 15 635–648
Targeted therapy for adrenocortical tumors
in transgenic mice through their LH receptor
by Hecate-human chorionic gonadotropin b
conjugate
Susanna Vuorenoja1,2, Adolfo Rivero-Müller1, Adam J Ziecik3,
Ilpo Huhtaniemi1,4, Jorma Toppari1,2 and Nafis A Rahman1
1
Department of Physiology, University of Turku, Kiinamyllynkatu 10, 20520 Turku, Finland
2
Department of Pediatrics, University of Turku, 20520 Turku, Finland
3
Institute of Animal Reproduction and Food Research, Polish Academy of Sciences, 10-714 Olsztyn, Poland
4
Department of Reproductive Biology, Imperial College London, London W12 ONN, UK
(Correspondence should be addressed to N A Rahman; Email: nafis.rahman@utu.fi)
Abstract
Novel strategies are needed for the treatment of adrenocortical tumors that are usually resistant
to chemotherapy. Hecate, a 23-amino acid lytic peptide, was conjugated to the 15-amino acid
(81–95) fragment of the human chorionic gonadotropin b (CGb) chain, which would selectively
kill cancer cells expressing the LH receptor (LHR) sparing the normal ones with LHR. To prove
the principle that Hecate-CGb conjugate may eradicate tumors ectopically expressing plasma
membrane receptors, transgenic (TG) inhibin a-subunit promoter (inha)/Simian Virus 40 T-
antigen mice, expressing LHR in their adrenal gland tumors, were used as the experimental
model. Wild-type control littermates and TG mice with adrenal tumors were treated with either
Hecate or Hecate-CGb conjugate at the age of 6.5 months for 3 weeks and killed 7 days after
the last treatment. The Hecate-CGb conjugate reduced the adrenal tumor burden significantly in
TG male but not in female mice, in comparison with Hecate-treated mice. Hecate-CGb
conjugate treatment did not affect normal adrenocortical function as the serum corticosterone
level between Hecate and Hecate-CGb conjugate groups were similar. The mRNA and protein
expressions of GATA-4 and LHR colocalized only in tumor area, and a significant
downregulation of gene expression was found after the Hecate-CGb conjugate in comparison
with Hecate- and/or non-treated adrenal tumors by western blotting. This finding provides
evidence for a selective destruction of the tumor cells by the Hecate-CGb conjugate. Hereby,
our findings support the principle that Hecate-CGb conjugate is able to specifically destroy
tumor cells that ectopically express LHR.
Endocrine-Related Cancer (2008) 15 635–648
Introduction at this moment is complete adrenalectomy since
Adrenocortical carcinomas (ACCs) are rare and aggres- with partial adrenalectomies the survival rate remains
quite poor. However, even in patients undergoing
sive types of human malignancies with poor prognosis as
complete surgical resection, recurrent and metastatic
they often are diagnosed late and the survival rate
disease is common (Reincke et al. 1994, Bornstein et al.
remains quite low (Schulick & Brennan 1999b). ACCs 1999). Chemotherapy with mitotane (o,p 0 -DDD) or
tend to occur in the first and fifth decades of life and are cisplatin is indicated for advanced stages of disease,
relatively more common in women than men (Schulick & although the outcome remains often poor with plenty of
Brennan 1999a,b). There are still no efficient forms of side effects including adrenal insufficiency (Ahlman
therapy for adrenal tumors. The only effective treatment et al. 2001). Thus, there is a great need for new curative
Endocrine-Related Cancer (2008) 15 635–648 Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
DOI: 10.1677/ERC-08-0015
via free access
1351–0088/08/015–635 q 2008 Society for Endocrinology Printed in Great Britain Online version via http://www.endocrinology-journals.org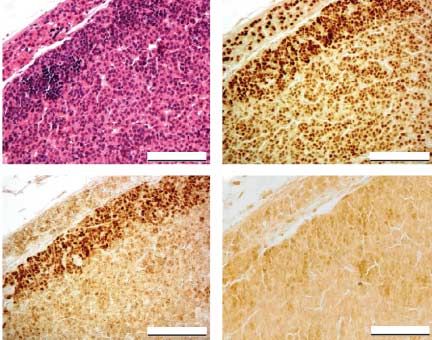
S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
methods as well as novel markers for early detection of destruction is necrosis without activation of
of these rare ACCs. Besides the normal gonadal apoptosis (Bodek et al. 2005b) and Hecate-CGb
expression of luteinizing hormone receptor (LHR), conjugate appears to selectively kill only cancer
there are reports about malignant androgen-producing cells with LHR and spare all other gonadal/non-
LH/human chorionic gonadotropin (hCG)-dependent gonadal healthy cells (even with LHR) due to
adrenal tumors (de Lange et al. 1980, Leinonen et al. negative/positive charge changes in their membrane
1991, Lacroix et al. 1999). The LHR is expressed at low potential (Leuschner & Hansel 2004, Bodek et al.
levels in normal human adrenal cortex (Pabon et al. 2005b). Hecate-CGb conjugate has previously been
1996), and hCG stimulates dehydroepiandrosterone studied in specific cancer cell lines and xenografts of
sulfate synthesis of human fetal adrenals (Jaffe et al. LHR-bearing carcinomas of prostate (Leuschner et al.
1981). LHR expression has been shown in different kinds 2001, 2003b, Bodek et al. 2005a), mammary gland
of human adrenal pathologies i.e., virilizing or Cushing’s (Bodek et al. 2003, Leuschner et al. 2003a), and ovary
syndrome (CS)-related adrenocorticotropin-independent (Gawronska et al. 2002). In vivo studies have so
macronodular adrenal hyperplasias (AIMAH; Lacroix far been carried out in transgenic (TG) mice bearing
et al. 1999, Bourdeau et al. 2001, Miyamura et al. 2002, ovarian and testicular tumors (Bodek et al. 2005b).
Feelders et al. 2003, Goodarzi et al. 2003, Mijnhout et al. TG mice expressing the inhibin a promoter/Simian
2004). Pregnancy-associated CS is also a known virus 40 (SV40) T-antigen (inha/Tag) were
condition (Sheeler 1994). LHR expression has also originally found to produce gonadal tumors with
been shown in aldosterone- (Saner-Amigh et al. 2006) or 100% penetrance at the age of 6 months, and if
androgen-producing LH-dependent adrenal adenomas gonadectomized prepubertally, they produced dis-
(Werk et al. 1973, Givens et al. 1975, Larson et al. 1976, cernible adrenal tumors at the same age, but never in
Smith et al. 1978, Takahashi et al. 1978, de Lange et al. intact mice (Kananen et al. 1996a, Rilianawati et al.
1980, Leinonen et al. 1991) and in ACCs (Wy et al. 2002,
1998, Rahman et al. 2004). The adrenal tumors and a
Barbosa et al. 2004). Thus, this abundant/upregulated
tumor-derived cell line (Ca1) express very high
LHR expression in the adrenal hyperplasia/adenomas
levels of LHR (Kananen et al. 1996a, Rilianawati
and adenocarcinomas makes them a susceptible target to
et al. 1998, Rahman et al. 2004), and the growth and
ligand as a strategy for tumor eradication through Hecate-
steroidogenesis of the adrenal tumors were shown to
human chorionic gonadotropin b (CGb) conjugate.
be dependent on LH stimulation (Mikola et al. 2003).
During the last few years, a novel targeted
The post-castration elevation of LH levels apparently
treatment approach for hormonally active, LHR-
induced the ectopic LHR expression, which together
bearing tumors has been developed using the
with the potent oncogene Tag expression triggered
Hecate-CGb conjugate (Leuschner et al. 2001).
Hecate is a synthetic lytic peptide based on the adrenocortical tumorigenesis (Mikola et al. 2003).
structure of melittin, the principal toxin of honeybee LH dependence of the tumors was proven by findings
venom. It rapidly destroys the outer membrane shell that they failed to appear if the post-castration
of only negatively charged cells such as bacteria and increase in gonadotropins was blocked by either
cancer cells by making small pores to the membrane treating the mice with a gonadotropin-releasing
(Leuschner & Hansel 2004, Rivero-Muller et al. hormone antagonist or crossbreeding them to the
2007). Cancer cells have been shown to be negatively gonadotropin-deficient hpg (hypergonadotropic)
charged due to the changed distribution of phospha- genetic background (Cattanach et al. 1977, Kananen
tidylserines in the outer cell membrane and thus, in et al. 1997). The adrenocortical tumors tend to grow
comparison with the healthy cells, only negatively in prepubertally gonadectomized mice by hyperpla-
charged cells are destroyed by this lytic peptide sia–adenoma–carcinoma sequence, as hyperplasia of
(Utsugi et al. 1991). To target the action of this lytic the adrenal cortex/adrenal adenoma could be
peptide directly to the cancer cells through a observed at the age of 4 months, whereas discernible
membrane receptor, Hecate can be conjugated to a gonadal tumors were seen only at 6 months with low
ligand or its receptor-binding domain, such as, in our metastatic incidence (Rahman et al. 2004). In this
case, to the 15-amino acid fragment (81–95) of the study, we took the advantage of this inha/Tag TG
human CGb chain (Hecate-CGb conjugate). This adrenocortical tumor model in order to prove the
allows the Hecate-CGb conjugate to bind specifically principle that adrenocortical tumor cells expressing
and disrupt cells bearing the LHR. The same principle ectopically a hormone receptor are sensitive to the
should apply to the plasma membrane receptors Hecate-CGb conjugate treatment in vivo, sparing
ectopically expressing in cancer cells. The mechanism simultaneously healthy cells.
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
636 www.endocrinology-journals.org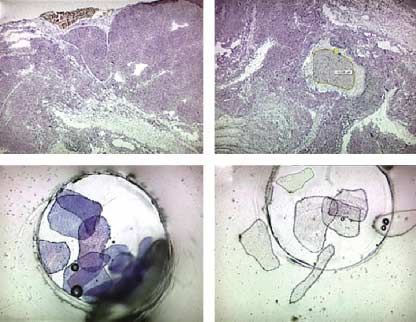
Endocrine-Related Cancer (2008) 15 635–648
Materials and methods xenografts (Leuschner et al. 2001) and TG mice with
gonadal tumors (Bodek et al. 2005b). Seven days
Experimental animals
following the last injection, mice were killed by
In order to induce adrenal tumors, we gonadectomized cervical dislocation and blood was collected by cardiac
inha/Tag TG mice prepubertally as described pre- puncture. Weights of body, tumor, and different organs
viously (Kananen et al. 1996a). As these mice have were recorded. Tissues were either snap-frozen in
earlier been extensively characterized (Kananen et al. liquid nitrogen, or fixed in Bouin’s solution or 4%
1996a, Rilianawati et al. 1998, 2000, Kero et al. 2000, paraformaldehyde and embedded in paraffin. Paraffin
Rahman et al. 2001, 2004), the discernible adrenocor- sections of 5 mm thickness were stained for further
tical tumors appear at the age of 6 months with 100% histological analysis with hematoxylin–eosin or used
penetrance. We started all the treatments at the age of for immunohistological analysis. For each tissue, at
6.5 months, in order to make sure that any treatment least five independent specimens were examined from
effect will be only due to the anti-tumoral effect, but each group.
not by prevention of the tumor development. Gona-
dectomy was performed under Avertin anesthesia and
postoperative buprenorphine analgesia was adminis- Northern hybridization analysis
tered on a routine basis. Seven to ten mice per Total RNA was isolated from cells using the single-step
treatment group were selected for the experiments. guanidinium thiocyanate–phenol–chloroform extrac-
Wild-type (WT) control littermate mice (C57Bl/6N) tion method (Chomczynski & Sacchi 1987). Twenty
were used as controls. For routine genotyping, PCR micrograms of RNA per lane were resolved on 1.2%
analysis was carried out using DNA extracts from ear denaturing agarose gel and transferred onto Hybond-XL
biopsies, as previously described (Kananen et al. nylon membranes (Amersham, Amersham Inter-
1995). After weaning at the age of 21 days, the mice national). Membranes were prehybridized overnight at
were housed two to four per cage, females and males in 65 8C in a solution containing 5! sodium chloride/
separate cages, in a room of controlled light (12 h sodium citrate (SSC), 5! Denhardt’s solution, 0.5%
light:12 h darkness) and temperature (21G1 8C). They SDS, 50% formamide, and 5 g/l of denatured calf
were fed with mouse chow SDS RM-3 (Whitham, thymus DNA. A complementary RNA probe for the rat
Essex, UK) and tap water ad libitum. The mice were LHR generated from a fragment of the LHR cDNA,
kept in a specific pathogen-free surrounding and were spanning nucleotides 441–849 of its extracellular
routinely screened for common mouse pathogens. The domain, subcloned into the pGEM-4Z plasmid was
University of Turku Ethics Committee on Use and used for hybridization (LaPolt et al. 1990). The
Care of Animals approved the animal experiments. [32P]-dUTP (800 Ci/mmol, Amersham) labeled probe
was generated using a Riboprobe system II kit
Preparation of drugs (Promega). The probes were purified with NickCol-
umns (Pharmacia Biotech). Hybridization was carried
Hecate and the Hecate-CGb conjugate were syn-
out at 65 8C for 20 h in the same prehybridization
thesized and purified in the Peptide and Protein
solution after an addition of labeled probe. After
Laboratory, Department of Virology, Hartman Institute,
hybridization, the membranes were washed twice in
University of Helsinki, as described previously
2! SSC with 0.1% SDS at room temperature for
(Bodek et al. 2003).
10 min, followed by two washes in 0.1! SSC with
0.1% SDS at 65 8C to remove most of the background.
Hecate and Hecate-CGb conjugate treatments
For GATA-4, the membranes were prehybridized
Male and female mice (inha/Tag or WT C57Bl control overnight at 42 8C in a solution containing 5! SSC,
littermates; nZ7–10 per group) were treated at the age 5! Denhardt’s, 0.5% SDS, 50% formamide, and 5 g/l
of 6.5 months with either Hecate-CGb conjugate denatured calf thymus DNA. The GATA-4 cDNA
(12 mg/kg b.w.) or Hecate (12 mg/kg b.w.) by i.p. probes were cut out from pMT2-GATA4 (Arceci et al.
injections. Since no substantial weight changes could 1993, Ketola et al. 1999), using EcoRI/PstI and SmaI
be observed between Hecate and Hecate-CGb restriction enzymes respectively labeled with Prime-
conjugate by adding hCG to Hecate, both Hecate and a-Gene kit (Pharmacia) using [a-P32]-dCTP for 4 h at
Hecate-CGb conjugate were given in the same dose 37 8C and purified with NickColumns. Hybridization
(Leuschner et al. 2001). The mice were injected once was carried out at 42 8C for 20 h in the same
per week for three consecutive weeks, according to an prehybridization solution after an addition of the
earlier protocol for in vivo treatment for nude mice denatured labeled cDNA probe. After hybridization,
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 637S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
the membranes were washed twice in 2! SSC and 0.1% antibody, dilution 1:4000; Acris Antibodies, Hidden-
SDS at room temperature for 10 min, followed by two hausen, Germany) were used separately. As secondary
washes in 0.1! SSC and 0.1% SDS at 42 8C to remove antibody, either bovine anti-goat IgG-HRP (sc-2350,
most of the background. Finally, the membranes were Santa Cruz Biotechnology) or ECL Anti-rabbit IgG,
exposed to Kodak X-ray films (Kodak XAR-5; Eastman HRP-linked whole AB (Amersham Biosciences) was
Kodak) at K70 8C for 4–7 days or to phosphor-imager used in dilution 1:10 000. Signals were visualized
(Fujifilm BAS-5000, Fujifilm IaI, Tokyo, Japan) for using ECL Plus Western blotting detection reagents
4–24 h. The intensities of specific bands were quantified (Amersham Pharmacia Biotech) and finally exposed to
using the Tina software (Raytest, Stranbenhardt, Fuji X-ray film (Super RX, Fuji Photo Film Ltd,
Germany) which came as an integrated program with Bedford, UK). The intensities of specific bands were
the phosphor-imager (Fujifilm BAS-500; El-Hefnawy quantified using the Tina Software (Raytest).
& Huhtaniemi 1998, El-Hefnawy et al. 2000, Manna
et al. 2001, Strassburg et al. 2002, Moran et al. 2006), Laser pressure catapulting (LPC)
and related to those of the 28S rRNA in the gel stained
LPC was performed by Laser Microbeam Microdissec-
with ethidium bromide. The molecular sizes of the
tion (PALM Microlaser Technologies, Bernried,
mRNA species were estimated by comparison with the
Germany) using a protocol described previously
mobility of the 18S and 28S rRNAs.
(Westphal et al. 2002). Briefly, for laser microdissec-
tion, cryosections (5 mm) were prepared from three
Hormone measurements
independent specimens. Subsequent dehydration was
Serum levels of LH were measured by immunofluoro- achieved using graded alcohol and xylene treatment as
metric assay for rat (Delfia; Wallac, Turku, Finland) as follows: 95% ethanol for 30 s (twice), 100% ethanol
described previously (Haavisto et al. 1993). Cortico- for 30 s (twice), and xylene for 5 min (twice). The
sterone levels were measured from diethyl ether extracts slides were dried under laminary flow for 10 min and
of the sera by RIA using a kit for rats and mice (MP then stained with hematoxylin using RNase-free
Biomedicals, Orangeburg, NY, USA) and progesterone conditions and kept on dry ice until LPC. LPC was
was measured by Delfia Progesterone Kit (Wallac). The performed using a Zeiss inverted microscope PALM
approximate assay sensitivities for LH, progesterone, Laser Micro-Beam System u.v. laser at 337 nm, linked
and corticosterone are 0.75 pg/tube, 50 fmol/tube, and to a PC with the required software programs. The
30 fmol/tube respectively. The intra- and inter-assay tumor and non-tumor areas to be dissected were
coefficients of variations for these assays were below selected by means of high-precision microcuts (the
10–15% respectively. specimens may range from as little as 1 to 1000 mm in
diameter). LPC dissection was performed using a few
Immunoblotting analysis for GATA-4 and LHR shots each of 100 mm in diameter from both tumor and
non-tumor areas and from each slide. After catapulting
Western blotting was used to determine the concen-
similar amounts of tumor and non-tumor area, the
tration changes in GATA-4 and LHR due to the
material was removed from the caps for further
different treatments (Hecate or Hecate-CGb conju-
analysis. Total RNA was extracted from tumor tissues
gate). Tissue samples were subjected to homogeni-
using RNeasy kit (Qiagen).
zation and protein concentration was measured using
the bicinchoninic acid (BCA) protein assay kit (Pierce,
RT-PCR and Southern hybridization
Rockford, IL, USA). Electrophoresis was carried out
through a 12.5% SDS-PAGE gel (1 mg protein/well). One microgram of total RNA was reverse transcribed
Dual Color Precision Plus Protein Standards (Bio-Rad) using 20 IU of avian myeloma virus reverse tran-
were used as standards. After electrophoresis, proteins scriptase (RT), 50 IU RNase inhibitor, 25 pmol random
were electrotransferred to Hybond-P PVDF Membrane primers, and 10 mmol of each dNTP (all from
(Amersham Biosciences) using Trans-Blot SD cell Promega), in 25 ml final volume for 1 h at room
(Bio-Rad) by 20 V for 60 min. After blocking the temperature. For amplification of LHR and GATA-4
membrane with TBS containing 2% fat-free milk cDNA, specific primers were used as described
powder C0.05 M Tween, the incubation with the previously (Morrisey et al. 1997, Kero et al. 2000).
primary antibody was performed at C4 8C overnight. PCR was performed in a 25 ml final volume, 4 ml RT
Anti-GATA-4 (goat polyclonal antibody, dilution solutions were mixed with 2 IU of the DyNAzyme II
1:100; C-20, sc-1237; Santa Cruz Biotechnology, DNA polymerase (Finnzymes Oy, Espoo, Finland),
Santa Cruz, CA, USA) or anti-LHR (rabbit polyclonal 10 pmol of each primer, and 10 mmol of each dNTP
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
638 www.endocrinology-journals.orgEndocrine-Related Cancer (2008) 15 635–648
(Zhang et al. 1994). The RT and PCRs were carried out Fazleabbas) and/or commercial rabbit polyclonal
sequentially in the same assay tube. First, the RT anti-GATA-4 IgG (dilution 1:800, Santa Cruz Bio-
reaction was carried out (50 8C for 10 min), followed technology). IHC for anti-steroidogenic factor (SF)-1
by a denaturation period of 3 min at 97 8C. Thereafter, (goat polyclonal antibody, 1:400, Santa Cruz Bio-
a PCR with 40 cycles (96 8C for 1.5 min, 57 8C for technology) was also carried out. The slides were
1.5 min, and 72 8C for 3 min, with a final extension incubated with primary antibody at 4 8C overnight, and
period of 10 min at 72 8C) was performed. The after incubation and washings with TBS–Tween buffer
sense primer for LHR corresponded to nucleotides the slides were incubated with the anti-rabbit (Vector
176–195 (5 0 -CTCTCACCTATCTCCCTGTC-3 0 ) and Laboratories, Inc., Burlingame, CA, USA) or anti-goat
the antisense primer to nucleotides 878–858 (Santa Cruz Biotechnology) secondary antibody. The
(5 0 -TCTTTCTTCGGCAAATTCCTG-3 0 ) of the avidin–biotin immunoperoxidase system was used to
mouse 700 bp fragment of the LHR cDNA. The visualize bound antibody (Vectastain Elite ABC Kit,
sense primer for GATA-4 corresponded to sense Vector Laboratories) with 3.3 0 -diaminobenzidine
nucleotides 1527–1550 (5 0 -AAACGGAAGCCCAA- (Sigma) as a substrate. As a control for antibodies,
GAACCTGAAT-3 0 ) and the antisense primer to adjacent sections were incubated with either 1%
1935–1953 (5 0 -GGCCCCCACGTCCCAAGTC-3 0 ) normal goat serum in PBS or rabbit IgG as a primary
(expected size: 427 bp) of mouse GATA-4 cDNA antibody to differentiate unspecific staining from
(Morrisey et al. 1997). As a control for RNA quality, a specific staining.
395 bp fragment of the L19 ribosomal protein gene was
co-amplified with each sample (sense primer, 5 0 -
GAAATCGCCAATGCCAACTC-3 0 ; antisense pri- Statistical analysis
mer, 5 0 -TCTTAGACCTGCGAGCCTCA-3 0 ). Kidney Statistical analyses were carried out by ANOVA and/or
RNA was used as a negative control. After RT-PCR, the non-parametrical Mann–Whitney Rank test using
20 ml aliquots of the reaction mixtures were loaded on SAS Enterprise Quide 3.0 program (SAS Institute Inc.,
1% agarose gel containing ethidium bromide (0.4 mg/ Cary, NC, USA). Non-parametrical tests were used
l), to identify the amplified DNA fragments. Southern when the variances were not equal. ANOVA was used
hybridization was used, according to standard tech- in other cases. P values !0.05 were regarded as
niques, to confirm the specificity of these PCR statistically significant. All the values are presented as
products. Hybridization was carried out with the 5 0 - meanGS.E.M.
end labeled oligonucleotide 5 0 -TGGAGAAGATGCA-
CAGTGGA-3 0 , corresponding to nucleotides 641–660
of LHR cDNA and for GATA-4 with the 5 0 - Results
AGTGGCACGTAGACGGGCGAGGAC-3 0 , corre-
sponding to nucleotides 1676–1699. The membranes Hecate-CGb conjugate treatment reduced
were washed according to the manufacturer’s instruc- adrenal tumor volume
tions and then exposed to Kodak X-ray film (XAR 5; Hecate-CGb conjugate had an antineoplastic effect on
Eastman Kodak). the adrenal tumors of inha/Tag TG mice. We have earlier
characterized the ontogeny of adrenocortical tumorigen-
esis in these mice and observed tumors weighing 20- to
Immunohistochemistry (IHC)
30-fold more than normal adrenals (290G69 mg) at
Paraformaldehyde-fixed paraffin sections (5 mm) of 6–6.5 months of age following pre-pubertal gonadect-
mouse adrenal tumors were deparaffinized and rehy- omy (Rahman et al. 2004). Following a 1-month
drated. For immunostaining, 3% H202 in water was treatment with Hecate-CGb conjugate, adrenal tumor
used to block endogenous peroxidases and all sections weight was reduced by an average of 59% in males
were boiled by microwave treatment for 15 min in compared with Hecate treatment (364G147 vs 150G
10 mM citric acid (pH 6.0) for antigen retrieval. Two 130 mg; Hecate versus Hecate-CGb conjugate; P!0.01;
washes were done after each step with 0.05 M Tris and Fig. 1A). In females, the difference was only 18% (292G
150 mM NaCl (TBS) with 0.1% Tween (TBS-T). 156 vs 240G225 mg; Hecate versus Hecate-CGb
Serial sections of adrenal gland and testes as a positive conjugate; PO0.05; Fig. 1B). As the age-related
control were subjected to immunohistochemistry with individual tumor progression rate was variable between
an anti-LHR antibody (dilution 1:1000, a rabbit inha/Tag TG mice and has been observed already earlier
polyclonal antiserum directed against human peptide in inha/Tag and Tag mice (Hanahan 1989, Kananen et al.
sequence, KKLPSRETFVNLLEA; a gift of Dr A 1995, 1996b, Rahman et al. 1998), we also analyzed the
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 639S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
differences in tumor weights/burden between the sexes
could be observed in the present study, which was in line
with the earlier observations (Rahman et al. 2004). The
adrenal tumors metastasized in 10% (nZ10 for both
male and/or female inha/Tag mice) cases in the Hecate-
treated group to liver (proven by histopathological
analysis, data not shown), but never in Hecate-CGb
conjugate-treated groups. According to earlier publi-
cations and observations, these tumors metastasized only
occasionally (around 10% cases) to liver, more
commonly in females than males (Kananen et al. 1996a).
LHR expression is higher in the adrenal tumors of
male mice
To find out why Hecate-CGb conjugate treatment was
less effective in female adrenal tumors, we quantified
LHR and GATA-4 mRNA expressions by northern
hybridization in male and female non-treated tumors
(Fig. 2). GATA-4 was included in the study due to its
close association with LHR expression and adreno-
cortical tumorigenesis (Kiiveri et al. 1999, Rahman et al.
2004). We also measured LHR and GATA-4 expressions
in the established Ca1 cell line derived from inha/Tag
adrenocortical tumor (Kananen et al. 1996a, Rilianawati
et al. 1998) and from non-treated adrenocortical tumors.
A significantly higher level of LHR mRNA expression
was found in the male than in the female in adrenal non-
treated tumors (P!0.01; Fig. 2). No significant
differences in GATA-4 mRNA levels between the female
and male adrenocortical tumors could be observed.
Endocrine consequences of the treatments
As sham-treated group (castrated TG mice treated with
vehicle) shows identical hormonal values with the
Hecate-treated group, which has been demonstrated
earlier for testicular and ovarian tumors (Bodek et al.
2005b), ovarian xenograft model (Gawronska et al.
Figure 1 Total adrenal weights of (A) male (nZ10) and 2002), and also rat mammary gland tumor model
(B) female (nZ10) inha/Tag transgenic (TG) and wild-type
(WT) (WT male, nZ7 and female, nZ8) mice after 3 weeks of
(Zaleska et al. 2004), in this study the sham-treated
Hecate-CGb conjugate or Hecate treatment. Tumor burden, group was not taken into account in order to concentrate
tumor weight/body weight in grams. The values are on the differences between the Hecate and Hecate-CGb
meanGS.E.M. *P!0.01; Hecate-CGb conjugate versus Hecate.
conjugate groups. In the WT groups, gonadectomy-
induced elevated levels of LH were clear due to the lack
tumor burden (tumor weight/body weight). This para- of negative feedback from the gonads. After 1 month of
meter also decreased after Hecate-CGb conjugate treatment with Hecate-CGb conjugate, no statistically
treatment significantly in males in comparison with significant differences could be seen in Hecate and
Hecate treatment. In females, the treatment effect Hecate-CGb conjugate treatment groups in the LH,
remained non-significant (Fig. 1A and B). We did not corticosterone, or progesterone levels in either sex
observe any statistically significant differences between (Fig. 3), although progesterone levels were twofold
the total body weights of the mice between the treatment lower, but not statistically significant in Hecate-CGb
groups (data not shown). No statistically significant conjugate-treated males in comparison with Hecate
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
640 www.endocrinology-journals.orgEndocrine-Related Cancer (2008) 15 635–648
group (in females no differences in hormonal values seen,
data not shown), due to the high individual variations
between the serum progesterone values (Fig. 3). Signi-
ficant change in LH levels was seen between gonadecto-
mized WT mice and the TG mice for both Hecate and
Hecate-CGb conjugate-treated groups (P!0.05; Fig. 3).
Serum progesterone levels were three- to four-fold
higher in the TG mice than in WT mice, which might
explain the lower level of LH in adrenal tumor-bearing
TG mice (P!0.05; Fig. 3). This finding is in agreement
with the earlier findings in adrenal tumors and adrenal
tumor-derived Ca1 cells (Kananen et al. 1996a)
indicating their steroidogenic activity, e.g., progesterone
secretion.
Hecate-CGb conjugate selectively destroyed the
LHR possessing adrenal tumor cells
To assess whether the Hecate-CGb conjugate treatment
can eradicate specifically the LHR-expressing adreno-
cortical tumor cells, we measured the LHR and GATA-4
protein expression levels in Hecate-CGb conjugate-
treated adrenals in comparison with the Hecate-treated
and non-treated adrenocortical tumor LHR protein
levels. Western blotting analysis revealed that there
was a concomitant significant decrease in LHR and
GATA-4 in the Hecate-CGb conjugate-treated adrenal
tumors (P!0.01 Hecate-CGb conjugate versus non-
treated or Hecate-treated adrenal tumors; Fig. 4),
indicating selective destruction of LHR and GATA-4
positive adrenocortical tumor cells. Higher GATA-4
Figure 2 Northern hybridization analysis of (A) LHR mRNA and expression compared with LHR after Hecate-CGb
(B) western blot analysis of the protein expression in inha/Tag conjugate might reflect the previously described
TG adrenal tumors of male and female mice and from tumor-
derived Ca1 cells. Here, in three independent experiments, phenomenon that these adrenocortical cells might
each time two different tumors for both sexes were used (nZ6 express GATA-4 without LHR, but not vice versa and
for both males and females). RNA was extracted from one-half their expression pattern was higher/broader (Kiiveri et al.
of the tumor and protein from the other half of the tumor.
(A) Each lane contains 20 mg total RNA. The migration of the 1999, 2002). The LHR or GATA-4 protein expression in
28S and 18S rRNAs is shown on the left side of the LHR panel. WT control mouse adrenals could not be detected.
The sizes of the different LHR mRNA splice variants (in kb) are
presented on the right. Two lanes for each type of sample are
depicted. The upper panel shows, on the top, the ethidium Histopathological analysis demonstrating anti-
bromide (EtBr) staining of the 28S rRNA for RNA loading
control. The middle and lower panels show northern hybrid- tumoral effects of Hecate-CGb conjugate
ization for GATA-4 and LHR mRNA respectively. The bottom treatment
panel shows densitometric quantification of the longest (7.0 kb)
LHR mRNA splice variant (filled bars) and GATA-4 (open bars) Histopathological analysis revealed that Hecate-CGb
in arbitrary densitometric units (the mean of Ca1 tumor cells for conjugate treatment of TG adrenocortical tumor mice
GATA-4 or LHR was designated as 100%) corrected for
intensity of the 28S rRNA band. Each bar represents the
meanGS.E.M. of three independent experiments, each time two (B) The upper panel shows GATA-4, middle panel LHR, and
different tumors for both sexes and from two different cell lower panel b-actin protein expressions in western blotting
extracts. Densitometric quantification of the six different bands analysis. Each lane represents three independent experiments,
is represented by one bar on the densitometric analysis. Each each time three different tumors for both sexes and from
band shows a different tumor/mouse. **P!0.01, *P!0.05 three different cell extracts. Each band shows a different
(P!0.01 and P!0.05; male versus female for LHR, male tumor/mouse. Ca1, adrenal tumor cell line derived from a
and/or female versus Ca1 cells for LHR). Ca1, adrenal transgenic mice expressing (inha)/Simian Virus 40 T-antigen
tumor cell line derived from a transgenic mice expressing (Tag) adrenal tumor; tu, tumor; on the right side, the molecular
(inha)/Simian Virus 40 T-antigen (Tag) adrenal tumor; tu, tumor. weights of the proteins in kilodaltons are indicated.
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 641S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
Figure 4 Western blot analysis for LHR and GATA-4. Protein
extracts from male inha/Tag TG mice treated with Hecate or
Hecate-CGb conjugate or from non-treated tumor samples
were analyzed by western blotting. The upper panel shows
LHR, GATA-4, and b-actin expressions and the lower panel
their densitometric analysis. The representative band data
shown are from one of the four different adrenal tumors/mouse
Figure 3 Serum (A) LH, (B) progesterone, and (C) corticoster- per group/independent experiments (nZ4). Each bar in
one concentrations in transgenic inha/Tag male mice after densitometric quantification shows the quantification of four
a 3-week treatment with Hecate-CGb conjugate or Hecate. separate independent tumor samples/experiments. Each bar
WT, wild-type or TG, inha/Tag mice treated with Hecate-CGb represents the meanGS.E.M. of four independent experiments,
conjugate or Hecate. The values are meanGS.E.M.; nZ5, each time different tumor sample. Hecate-CGb conjugate
*P!0.05; Hecate-CGb conjugate-treated TG mice versus downregulated significantly both LHR and GATA-4 expressions
Hecate or Hecate-CGb conjugate-treated WT mice. compared with non-treated control adrenal tumor or Hecate-
treated tumor (**P!0.001). No detectable LHR or GATA-4 was
detected in WT C57Bl mouse adrenals. On the right side, the
molecular weights of the proteins in kilodaltons are indicated.
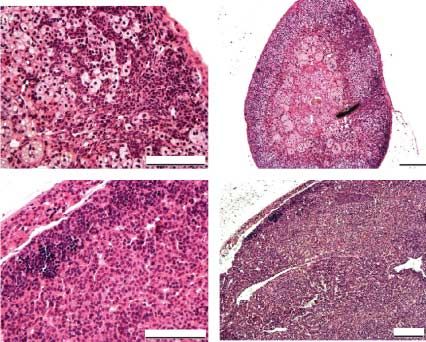
induced a definite reduction of the adrenocortical tumor
LHR and GATA-4 expressions are colocalized in
mass where residual tumor tissue could be observed in a
the tumors
reduced area of the zona glomerulosa in hematoxylin–
eosin-stained sections (Fig. 5B). This finding in the We next demonstrated that LHR and GATA-4 are
representative sample was reproducible as the same co-localized in the same adrenocortical tumor region.
phenomenon was observed in all stained samples of the The data obtained by LPC of the tumorous and non-
same treatment groups (nZ5 from each group). The tumorous tissues from the same slice demonstrated the
adrenal tumors metastasized in 10% (nZ10 for both co-localization of GATA-4 and LHR mRNA messages
males and/or female inha/Tag mice) cases in the in adrenal tumor tissue and their absence in normal
Hecate-treated group to liver (proven by histopatholo- adrenal cortex (Fig. 6). The finding in the representa-
gical analysis, data not shown), but never in Hecate- tive sample was reproducible as the same phenomenon
CGb conjugate-treated groups. According to earlier was observed in all other three samples.
publications and observations, these tumors metasta- We then investigated whether the expression
sized only occasionally (around 10% cases) to liver, patterns of these proteins were affected by the Hecate
more commonly in females than males (Kananen or Hecate-CGb conjugate treatment. GATA-4
et al. 1996a). expression was found to directly localize to the
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
642 www.endocrinology-journals.orgEndocrine-Related Cancer (2008) 15 635–648
Figure 5 Hematoxylin–eosin staining of adrenal tumors after
treatments with (A and C) Hecate or (B and D) Hecate-CGb
conjugate. (A and B, 20! magnification, C and D, 10!
magnification). The picture from (C) Hecate-treated tissue is
taken from the contralateral side of the big tumor to show the
expression of the tumorous tissue in the whole adrenal gland.
H–E-stained tumor area that was in line with our
previous findings (Rahman et al. 2004; Fig. 7A and C).
In Hecate-CGb conjugate-treated sections, no LHR
staining could be detected even in the same H–E-
stained, marginalized, and reduced tumorous area
(Fig. 7A and D). We also found SF-1 expression in
Figure 6 RT-PCR/Southern hybridization of the PALM micro-
the same tumor areas where GATA-4 expression could dissected samples. (A and B) Hematoxylin-stained specimens
be observed (Fig. 7B and C). of cryosections of the tumorous adrenal tissue. (A) The tumor
sections (dark-stained cells) and (B) non-tumor tissues (light-
stained area in the middle) were dissected out and catapulted.
(C) The catapulted tumor (WT, wild-type; adr tum, adrenal
Discussion tumor) and (D) normal non-tumorous tissues in the cap of an
Eppendorf tube (magnification 63!; adr non-tu, non-tumorus
The lack of a suitable animal model for adrenocortical adrenal tissue).
tumorigenesis has been a major obstacle for unraveling
the molecular mechanisms involved in their patho-
(Rao Ch et al. 2004, Alevizaki et al. 2006, Vuorenoja
genesis, as well as for improving their diagnostic and
et al. 2007) suggest that the adrenal cortex expresses low
therapeutic strategies. Thus, establishment of suitable
levels of LHR (Pabon et al. 1996) whose function
animal models is of extremely high importance in order
to study human adrenal tumorigenesis. Chronically becomes significant when LH levels are elevated, as after
elevated LH, ectopic/upregulated LHR expression, gonadectomy or postmenopausally. In light of the present
hyperplasia–adenoma–adenocarcinoma sequence tumor study, we could postulate that the upregulated LHR
formation, slow tumor growth, late discernible tumor expression in adrenal tumors is functionally relevant and
incidence (analogous to human postmenopausal/andro- can be used to target the tumors by the cytotoxic Hecate-
pausal age), low metastases frequency (5–10%): these CGb conjugate. This model serves as an example
characteristics make the inha/Tag TG murine model a demonstrating how ectopically expressed receptors in
good tumor relevance model for human ACCs studies. tumors can be used as therapeutic target.
Our inha/Tag TG mouse is at the moment one of the We used in the present study an established inha/Tag
rare and unique murine models in which adrenocortical TG mouse model for post-gonadectomy adrenocortical
tumorigenesis and its molecular mechanisms or any tumorigenesis, where the pathophysiological and
treatment strategies can be investigated. Although the endocrine responses induced by tumorigenesis are
physiologic significances of extragonadal LHR well characterized. We found that Hecate-CGb con-
expression in reproductive function is questioned jugate treatment reduced the total adrenocortical tumor
(Pakarainen et al. 2005, 2007), some recent studies weights without detectable side effects, e.g., adrenal
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 643S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
tumors attracting larger amounts of Hecate-CGb
conjugate, exerting consequently a more severe lytic
effect. The reason for the higher LHR expression in
male adrenal tumors remains unknown. The putative
difference in tumor cell membrane charges between the
male and females could also be one of the potential
mechanistic reasons behind this differential treatment
effects, which needs to be further evaluated in future
studies.
As we did not find significant changes in the serum
endocrine profiles of the Hecate- or Hecate-CGb
conjugate-treated mice, the treatment outcome could
not be monitored by serum hormone measurements.
The progesterone levels are decreased in Hecate-CGb
conjugate male in comparison with Hecate group but the
Figure 7 Hematoxylin–eosin staining (A) and immunohisto- high individual variations made them non-significant.
chemistry for (B) SF-1, (C) GATA-4 and (D) LHR in Hecate-CGb
conjugate-treated adrenal tumor sections. LH levels were higher in gonadectomized WT than in
TG males, which indicated increased endocrine activity
insufficiency. Curiously, the treatment was, for of the tumors. The hormone with increased production
unknown reason, ineffective in female with adrenocor- by the tumors exerting a negative feedback action of LH
tical tumors. The adrenal response to elevated LH in secretion is probably progesterone. However, the
inha/Tag mice was probably more sensitive in females adrenal tumors can also produce sex steroids (androgens
and we believe it was also more aggressive, which could or estrogens; Leinonen et al. 1991, Kananen et al.
explain why immortalization of Ca1 cells from a female 1996a, Lacroix et al. 1999, Bielinska et al. 2005). No
effect of the treatment could be seen on corticosterone
inha/Tag founder mouse tumor was possible (Kananen
levels between Hecate and Hecate-CGb conjugate
et al. 1996a, Rilianawati et al. 1998). It has been shown
treatment groups, which indicates that the Hecate-
that the castrated male mice developed adrenocortical
CGb conjugate treatment does not destroy normal
tumors more slowly than gonadectomized females
adrenal tissue or induce adrenocortical hypofunction.
along with the ectopic LHR expression (Bielinska
This additionally supports the principle that this peptide
et al. 2003). Moreover, the elevated LH levels induced
attacks only the negatively charged tumor cells and
abundant ectopic LHR expression and acted as a tumor
spares the healthy ones.
promoter selectively only in the intact female double-
We have shown earlier a precise co-initiation of
positive TG mice, where bLHb-CTP mice producing GATA-4 along with the LHR expression during
constitutively elevated levels of LH (Risma et al. 1995) adrenocortical tumorigenesis ontogeny (Rahman et al.
were crossbred with inha/Tag TG mice (Mikola et al. 2004). GATA-4 has been proposed to be a potential
2003). This never happened in the male double-positive marker for adrenocortical tumorigenesis (Kiiveri et al.
mice (Mikola et al. 2003). In humans, ACCs are also 1999), along with LHR (Rahman et al. 2004). Further
more common in postmenopausal women than in aging proof is needed to determine whether GATA-4 works as
males (Schulick & Brennan 1999a,b). Therefore, it a specific co-activator for LHR (or even vice versa) in
could be an established phenomenon that female adrenocortical tumorigenesis. Thus, we wanted to see
adrenals respond more readily to high LH and whether GATA-4 expression would also diminish due
tumorigenesis might be more aggressive in them, to the Hecate-CGb conjugate treatment, which could
which might imply that the suppression of LH action lead us to further mechanistic studies in the future. As
needed to block tumorigenesis in females should be there are some difficulties with the availability of good
more complete/stronger, whereas male tumorigenesis LHR antibodies, GATA-4 immunohistochemistry
can be blocked with less-effective suppression. It is could also additionally serve as a marker for treatment
likely that this mechanism occurred in our Hecate-CGb efficacy. We found that both LHR and GATA-4 protein
conjugate-treated inha/Tag mice where, with the expressions decreased significantly after Hecate-
applied dose of the drug, we achieved much a better CGb conjugate treatment compared with Hecate. By
response in the males than in the females. Probably, this immunohistochemistry, we could detect GATA-4 in
phenomenon was additionally helped by the higher residual tumor area indicating that some tumor
concentration of LHR in the tumor mass in the male steroidogenic cells are still left, as GATA-4 is expressed
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
644 www.endocrinology-journals.orgEndocrine-Related Cancer (2008) 15 635–648
only in tumor adrenocortical steroidogenic cells LHR antibody and Dr Sonia Bourguiba for fruitful
(Kiiveri et al. 1999, 2002, Peterson et al. 2004), but discussions. The authors also thank Ms Johanna
LHR staining was no longer detectable in the Hecate- Lahtinen, Ms Tarja Laiho and Ms Heli Niittymäki for
CGb conjugate-treated samples. These results indicated their technical assistance and the personnel of Turku
that Hecate-CGb conjugate killed specifically the tumor University Animal Facility for taking good care of the
cells expressing LHR. This finding is well in line with mice. This project was supported by grants from the
the adrenal tumor weight results. Even after the Hecate- Finnish Cultural Foundation at Varsinais-Suomi (S V),
CGb conjugate treatment, the adrenal has not returned the Moikoinen Cancer Research Foundation (N R), the
to totally normal size. However, GATA-4 and SF-1 Academy of Finland (I H, J T, N R), and the Turku
were colocalized in the same tumor area. SF-1 is known University Hospital (J T). We declare that there is no
to be another factor activating adrenocortical steroido- conflict of interest that would prejudice this manuscript’s
genesis and it is shown to transactivate both GATA-4 impartiality.
and LHR activities (Tremblay & Viger 1999). The
expression pattern of SF-1 may implicate its role as a
co-activator of GATA-4 and LHR in adrenocortical
tumorigenesis of the inha/Tag TG mice.
References
Human adrenal LH/hCG responsive tumors have been Ahlman H, Khorram-Manesh A, Jansson S, Wangberg B,
well described (Leinonen et al. 1991, Lacroix et al. Nilsson O, Jacobsson CE & Lindstedt S 2001 Cytotoxic
1999). Higher basal levels of corticosteroids in post- treatment of adrenocortical carcinoma. World Journal of
menopausal women are also associated with higher Surgery 25 927–933.
circulating LH and LHR expressions in adrenals (Pabon Alevizaki M, Saltiki K, Mantzou E, Anastasiou E &
Huhtaniemi I 2006 The adrenal gland may be a target of
et al. 1996, Alevizaki et al. 2006). A recent publication
LH action in postmenopausal women. European
confers that ectopic LHR may induce adrenal hyper-
Journal of Endocrinology 154 875–881.
plasia- and Cushing’s syndrome-like symptoms as well Arceci RJ, King AA, Simon MC, Orkin SH & Wilson DB
as hCG-mediated steroidogenic activity (Mazzuco et al. 1993 Mouse GATA-4: a retinoic acid-inducible GATA-
2006). Ectopic expression of LHR in the adrenal gland binding transcription factor expressed in endodermally
has been shown to be associated with not only derived tissues and heart. Molecular and Cellular Biology
adrenocorticotropin-independent Cushing’s syndrome 13 2235–2246.
but also benign aldosterone-producing adenomas Barbosa AS, Giacaglia LR, Martin RM, Mendonca BB & Lin
(Saner-Amigh et al. 2006). There are also reports about CJ 2004 Assessment of the role of transcript for GATA-4
malignant androgen-producing LH-dependent adrenal as a marker of unfavorable outcome in human adreno-
tumors (de Lange et al. 1980, Leinonen et al. 1991). cortical neoplasms. BMC Endocrine Disorder 4 3.
Hence, LH/hCG-dependent mechanisms of adrenal Bielinska M, Genova E, Boime I, Parviainen H, Kiiveri S,
pathogenesis do exist and they may be clinically Leppaluoto J, Rahman N, Heikinheimo M & Wilson DB
2005 Gonadotropin-induced adrenocortical neoplasia in
important in circumstances with elevated LH levels,
NU/J nude mice. Endocrinology 146 3975–3984.
e.g., in connection with polycystic ovarian syndrome and Bielinska M, Parviainen H, Porter-Tinge SB, Kiiveri S,
after menopause in women. Genova E, Rahman N, Huhtaniemi IT, Muglia LJ,
Taken together, the present data provide novel Heikinheimo M & Wilson DB 2003 Mouse strain
evidence that targeted destruction of adrenocortical susceptibility to gonadectomy-induced adrenocortical
tumors expressing ectopically LHR by the Hecate-CGb tumor formation correlates with the expression of GATA-
conjugate is possible. In addition to the currently used TG 4 and luteinizing hormone receptor. Endocrinology 144
mouse model, our findings prove the more general 4123–4133.
principle that receptors expressed ectopically in malig- Bodek G, Rahman NA, Zaleska M, Soliymani R, Lankinen H,
nant cells can be exploited in targeted cytotoxic therapies Hansel W, Huhtaniemi I & Ziecik AJ 2003 A novel
without affecting the normal healthy cells. approach of targeted ablation of mammary carcinoma cells
through luteinizing hormone receptors using Hecate-CGb
conjugate. Breast Cancer Research and Treatment 79
Acknowledgements 1–10.
Bodek G, Kowalczyk A, Waclawik A, Huhtaniemi I & Ziecik
This work was supported by grants from Finnish Cultural AJ 2005a Targeted ablation of prostate carcinoma cells
Foundation at Varsinais-Suomi, the Finnish Cancer through LH receptor using Hecate-CGb conjugate:
Society, the Moikoinen Cancer Research Foundation, functional characteristic and molecular mechanism of cell
the Academy of Finland, and Turku University Hospital. death pathway. Experimental Biology and Medicine 230
We thank Dr A Fazleabbas for the kind donation of the 421–428.
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 645S Vuorenoja et al.: Hecate-CGb conjugate and adrenocortical tumor
Bodek G, Vierre S, Rivero-Muller A, Huhtaniemi I, Ziecik Hanahan D 1989 Transgenic mice as probes into complex
AJ & Rahman NA 2005b A novel targeted therapy of systems. Science 246 1265–1275.
Leydig and granulosa cell tumors through the luteinizing Jaffe RB, Seron-Ferre M, Crickard K, Koritnik D, Mitchell
hormone receptor using a hecate-chorionic gonadotropin BF & Huhtaniemi IT 1981 Regulation and function of the
beta conjugate in transgenic mice. Neoplasia 7 497–508. primate fetal adrenal gland and gonad. Recent Progress in
Bornstein SR, Stratakis CA & Chrousos GP 1999 Adreno- Hormone Research 37 41–103.
cortical tumors: recent advances in basic concepts and Kananen K, Markkula M, Rainio E, Su JG, Hsueh AJ &
clinical management. Annals of Internal Medicine 130 Huhtaniemi IT 1995 Gonadal tumorigenesis in transgenic
759–771. mice bearing the mouse inhibin alpha-subunit promoter/-
Bourdeau I, D’Amour P, Hamet P, Boutin JM & Lacroix A simian virus T-antigen fusion gene: characterization of
2001 Aberrant membrane hormone receptors in inciden- ovarian tumors and establishment of gonadotropin-
tally discovered bilateral macronodular adrenal hyper- responsive granulosa cell lines. Molecular Endocrinology
plasia with subclinical Cushing’s syndrome. Journal of 9 616–627.
Clinical Endocrinology and Metabolism 86 5534–5540. Kananen K, Markkula M, Mikola M, Rainio EM, McNeilly A
Cattanach BM, Iddon CA, Charlton HM, Chiappa SA & Fink & Huhtaniemi I 1996a Gonadectomy permits adrenocor-
G 1977 Gonadotrophin-releasing hormone deficiency in a tical tumorigenesis in mice transgenic for the mouse
mutant mouse with hypogonadism. Nature 269 338–340. inhibin alpha-subunit promoter/simian virus 40 T-antigen
Chomczynski P & Sacchi N 1987 Single-step method of fusion gene: evidence for negative autoregulation of the
RNA isolation by acid guanidinium thiocyanate- phenol- inhibin alpha- subunit gene. Molecular Endocrinology 10
chloroform extraction. Analytical Biochemistry 162 1667–1677.
156–159. Kananen K, Markkula M, El-Hefnawy T, Zhang FP, Paukku T,
El-Hefnawy T & Huhtaniemi I 1998 Progesterone can Su JG, Hsueh AJ & Huhtaniemi I 1996b The mouse inhibin
participate in down-regulation of the luteinizing hormone alpha-subunit promoter directs SV40 T-antigen to Leydig
receptor gene expression and function in cultured murine cells in transgenic mice. Molecular and Cellular
Leydig cells. Molecular and Cellular Endocrinology 137 Endocrinology 119 135–146.
127–138. Kananen K, Rilianawati, Paukku T, Markkula M, Rainio
El-Hefnawy T, Manna PR, Luconi M, Baldi E, Slotte JP & EM & Huhtanemi I 1997 Suppression of gonadotropins
Huhtaniemi I 2000 Progesterone action in a murine inhibits gonadal tumorigenesis in mice transgenic
Leydig tumor cell line (mLTC-1), possibly through a for the mouse inhibin alpha-subunit promoter/simian
nonclassical receptor type. Endocrinology 141 247–255. virus 40 T-antigen fusion gene. Endocrinology 138
Feelders RA, Lamberts SW, Hofland LJ, van Koetsveld PM, 3521–3531.
Verhoef-Post M, Themmen AP, de Jong FH, Bonjer HJ, Kero J, Poutanen M, Zhang FP, Rahman N, McNicol AM,
Clark AJ, van der Lely AJ et al. 2003 Luteinizing Nilson JH, Keri RA & Huhtaniemi IT 2000 Elevated
hormone (LH)-responsive Cushing’s syndrome: the luteinizing hormone induces expression of its receptor
demonstration of LH receptor messenger ribonucleic acid and promotes steroidogenesis in the adrenal cortex.
in hyperplastic adrenal cells, which respond to chorionic Journal of Clinical Investigation 105 633–641.
gonadotropin and serotonin agonists in vitro. Journal of Ketola I, Rahman N, Toppari J, Bielinska M, Porter-Tinge
Clinical Endocrinology and Metabolism 88 230–237. SB, Tapanainen JS, Huhtaniemi IT, Wilson DB &
Gawronska B, Leuschner C, Enright FM & Hansel W 2002 Heikinheimo M 1999 Expression and regulation of
Effects of a lytic peptide conjugated to beta HCG on transcription factors GATA-4 and GATA-6 in developing
ovarian cancer: studies in vitro and in vivo. Gynecologic mouse testis. Endocrinology 140 1470–1480.
Oncology 85 45–52. Kiiveri S, Siltanen S, Rahman N, Bielinska M, Lehto VP,
Givens JR, Andersen RN, Wiser WL, Donelson AJ & Huhtaniemi IT, Muglia LJ, Wilson DB & Heikinheimo M
Coleman SA 1975 A testosterone-secreting, gonado- 1999 Reciprocal changes in the expression of transcrip-
tropin-responsive pure thecoma and polycystic ovarian tion factors GATA-4 and GATA-6 accompany adreno-
disease. Journal of Clinical Endocrinology and cortical tumorigenesis in mice and humans. Molecular
Metabolism 41 845–853. Medicine 5 490–501.
Goodarzi MO, Dawson DW, Li X, Lei Z, Shintaku P, Rao CV Kiiveri S, Liu J, Westerholm-Ormio M, Narita N, Wilson
& Van Herle AJ 2003 Virilization in bilateral macro- DB, Voutilainen R & Heikinheimo M 2002 Tran-
nodular adrenal hyperplasia controlled by luteinizing scription factors GATA-4 and GATA-6 during mouse
hormone. Journal of Clinical Endocrinology and and human adrenocortical development. Endocrine
Metabolism 88 73–77. Research 28 647–650.
Haavisto AM, Pettersson K, Bergendahl M, Perheentupa A, Lacroix A, Hamet P & Boutin JM 1999 Leuprolide acetate
Roser JF & Huhtaniemi I 1993 A supersensitive therapy in luteinizing hormone–dependent Cushing’s
immunofluorometric assay for rat luteinizing hormone. syndrome. New England Journal of Medicine 341
Endocrinology 132 1687–1691. 1577–1581.
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
646 www.endocrinology-journals.orgEndocrine-Related Cancer (2008) 15 635–648
de Lange WE, Pratt JJ & Doorenbos H 1980 A gonadotrophin secretion by vasopressin and catecholamines: detection of
responsive testosterone producing adrenocortical ade- the aberrant hormone receptors on adrenal gland.
noma and high gonadotrophin levels in an elderly woman. Endocrine 19 319–326.
Clinical Endocrinology 12 21–28. Moran DS, Eli-Berchoer L, Heled Y, Mendel L, Schocina M
LaPolt PS, Oikawa M, Jia XC, Dargan C & Hsueh AJ 1990 & Horowitz M 2006 Heat intolerance: does gene
Gonadotropin-induced up- and down-regulation of rat transcription contribute? Journal of Applied Physiology
ovarian LH receptor message levels during follicular 100 1370–1376.
growth, ovulation and luteinization. Endocrinology 126 Morrisey EE, Ip HS, Tang Z & Parmacek MS 1997 GATA-4
3277–3279. activates transcription via two novel domains that are
Larson BA, Vanderlaan WP, Judd HL & McCullough DL conserved within the GATA-4/5/6 subfamily. Journal of
1976 A testosterone-producing adrenal cortical adenoma Biological Chemistry 272 8515–8524.
in an elderly woman. Journal of Clinical Endocrinology Pabon JE, Li X, Lei ZM, Sanfilippo JS, Yussman MA & Rao
and Metabolism 42 882–887. CV 1996 Novel presence of luteinizing hormone/chor-
Leinonen P, Ranta T, Siegberg R, Pelkonen R, Heikkila P & ionic gonadotropin receptors in human adrenal glands.
Kahri A 1991 Testosterone-secreting virilizing adrenal Journal of Clinical Endocrinology and Metabolism 81
adenoma with human chorionic gonadotrophin receptors 2397–2400.
and 21-hydroxylase deficiency. Clinical Endocrinology Pakarainen T, Zhang FP, Poutanen M & Huhtaniemi I 2005
34 31–35. Fertility in luteinizing hormone receptor-knockout mice
Leuschner C & Hansel W 2004 Membrane disrupting lytic after wild-type ovary transplantation demonstrates
peptides for cancer treatments. Current Pharmaceutical redundancy of extragonadal luteinizing hormone action.
Design 10 2299–2310. Journal of Clinical Investigation 115 1862–1868.
Leuschner C, Enright FM, Melrose PA & Hansel W 2001 Pakarainen T, Ahtiainen P, Zhang FP, Rulli S, Poutanen M &
Targeted destruction of androgen-sensitive and -insensi- Huhtaniemi I 2007 Extragonadal LH/hCG action-not yet
tive prostate cancer cells and xenografts through time to rewrite textbooks. Molecular and Cellular
luteinizing hormone receptors. Prostate 46 116–125.
Endocrinology 269 9–16.
Leuschner C, Enright FM, Gawronska B & Hansel W 2003a
Peterson RA II, Kiupel M, Bielinska M, Kiiveri S,
Membrane disrupting lytic peptide conjugates destroy
Heikinheimo M, Capen CC & Wilson DB 2004
hormone dependent and independent breast cancer cells
Transcription factor GATA-4 is a marker of anaplasia in
in vitro and in vivo. Breast Cancer Research and
adrenocortical neoplasms of the domestic ferret (Mustela
Treatment 78 17–27.
putorius furo). Veterinary Pathology 41 446–449.
Leuschner C, Enright FM, Gawronska-Kozak B & Hansel W
Rahman NA, Kananen Rilianawati K, Paukku T, Mikola M,
2003b Human prostate cancer cells and xenografts are
Markkula M, Hamalainen T & Huhtaniemi IT 1998
targeted and destroyed through luteinizing hormone
Transgenic mouse models for gonadal tumorigenesis.
releasing hormone receptors. Prostate 56 239–249.
Molecular and Cellular Endocrinology 145 167–174.
Manna PR, El-Hefnawy T, Kero J & Huhtaniemi IT 2001
Rahman NA, Kiiveri S, Siltanen S, Levallet J, Kero J, Lensu
Biphasic action of prolactin in the regulation of murine
T, Wilson DB, Heikinheimo MT & Huhtaniemi IT 2001
Leydig tumor cell functions. Endocrinology 142 308–318.
Mazzuco TL, Chabre O, Feige JJ & Thomas M 2006 Adrenocortical tumorigenesis in transgenic mice: the role
Aberrant expression of human luteinizing hormone of luteinizing hormone receptor and transcription factors
receptor by adrenocortical cells is sufficient to provoke GATA-4 and GATA-61. Reproductive Biology 1 5–9.
both hyperplasia and Cushing’s syndrome features. Rahman NA, Kiiveri S, Rivero-Muller A, Levallet J, Vierre S,
Journal of Clinical Endocrinology and Metabolism 91 Kero J, Wilson DB, Heikinheimo M & Huhtaniemi I 2004
196–203. Adrenocortical tumorigenesis in transgenic mice expres-
Mijnhout GS, Danner SA, van de Goot FR & van Dam EW sing the inhibin alpha-subunit promoter/simian virus
2004 Macronodular adrenocortical hyperplasia in a 40 T-antigen transgene: relationship between ectopic
postmenopausal woman. Netherlands Journal of expression of luteinizing hormone receptor and transcri-
Medicine 62 454–455. ption factor GATA-4. Molecular Endocrinology 18
Mikola M, Kero J, Nilson JH, Keri RA, Poutanen M & 2553–2569.
Huhtaniemi I 2003 High levels of luteinizing hormone Rao ChV, Zhou XL & Lei ZM 2004 Functional luteinizing
analog stimulate gonadal and adrenal tumorigenesis in hormone/chorionic gonadotropin receptors in human
mice transgenic for the mouse inhibin-alpha-subunit adrenal cortical H295R cells. Biology of Reproduction 71
promoter/Simian virus 40 T-antigen fusion gene. 579–587.
Oncogene 22 3269–3278. Reincke M, Karl M, Travis WH, Mastorakos G, Allolio B,
Miyamura N, Taguchi T, Murata Y, Taketa K, Iwashita S, Linehan HM & Chrousos GP 1994 p53 Mutations in
Matsumoto K, Nishikawa T, Toyonaga T, Sakakida M & human adrenocortical neoplasms: immunohistochemical
Araki E 2002 Inherited adrenocorticotropin-independent and molecular studies. Journal of Clinical Endocrinology
macronodular adrenal hyperplasia with abnormal cortisol and Metabolism 78 790–794.
Downloaded from Bioscientifica.com at 02/01/2021 07:18:49PM
via free access
www.endocrinology-journals.org 647You can also read

















































