THE MOLECULAR BASIS OF THE ANTICANCER PROPERTIES OF QUERCETIN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
REVIEW
Volume 3, issue 3, 2021: 496-520
THE MOLECULAR BASIS OF THE ANTICANCER
PROPERTIES OF QUERCETIN
S. Adorisio1, M. P. Argentieri2, P. Avato2, G. Caderni3, S. Chioccioli3, S. Cirmi2,
D. V. Delfino1, G. Greco4, P. Hrelia5, M. Iriti6, M. Lenzi5, G. E. Lombardo7, C. Luceri3,
A. Maugeri7, M. Montopoli8, I. Muscari9, M. F. Nanì10, M. Navarra7, S. Gasperini5,
E. Turrini4, C. Fimognari4
1
Foligno Nursing School, Department of Medicine, University of Perugia, Foligno, Perugia, Italy
2
Department of Pharmacy-Drug Sciences, University of Bari Aldo Moro, Bari, Italy
3
Department of NEUROFARBA, Section of Pharmacology and Toxicology, University of Florence, Florence, Italy
4
Department for Life Quality Studies, University of Bologna, Bologna, Italy
5
Department of Pharmacy and Biotechnologies, University of Bologna, Bologna, Italy
6
Department of Agricultural and Environmental Sciences, Milan State University, Milan, Italy
7
Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina,
Messina, Italy
8
Department of Pharmaceutical and Pharmacological Sciences, University of Padova, Padua, Italy
9
Department of Medicine, University of Perugia, Santa Maria Hospital, Terni, Italy
10
Department of Pharmacy, University of Naples Federico II, Naples, Italy
All the authors belong to the Working Group “Pharmacognosy, Phytotherapy and Nutraceuticals” of the Italian
Pharmacological Society, Milan, Italy
E-mail: carmela.fimognari@unibo.it
Doi: 10.36118/pharmadvances.2021.10
SUMMARY
Quercetin is a major constituent of various dietary products, which is increasingly being investigated as a therapeutic
option in the oncological field. It has attracted extensive interest due to its ability of interacting with different molecular
targets and evoking a broad spectrum of chemopreventive and anticancer activities. In this review, we have tried
to present and critically discuss its potential against an extensive range of cancers including lung, ovarian, prostate,
breast, colorectal, bladder cancers. We also highlighted studies that combined quercetin with standard anticancer drugs
and delivered it via novel techniques and included a detailed description of its proposed mechanism(s) of action, and
pharmacokinetic and safety profile.
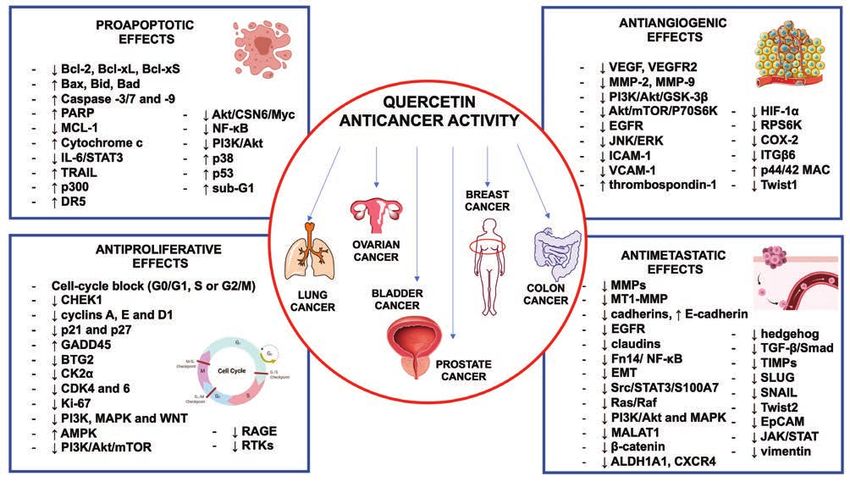
Key words Impact statement
Quercetin; cancer; bioavailability; mechanisms of action; Quercetin exhibits a broad spectrum of anticancer activi-
in vivo studies; in vitro studies. ties such as pro-apoptotic, antiproliferative, antiangiogenic,
and antimetastatic effects and blocks an extensive range of
cancers including lung, ovarian, prostate, breast, colorectal,
bladder cancers.
© 2021 The Italian Society of Pharmacology (SIF). Published by EDRA SPA. All rights reserved
496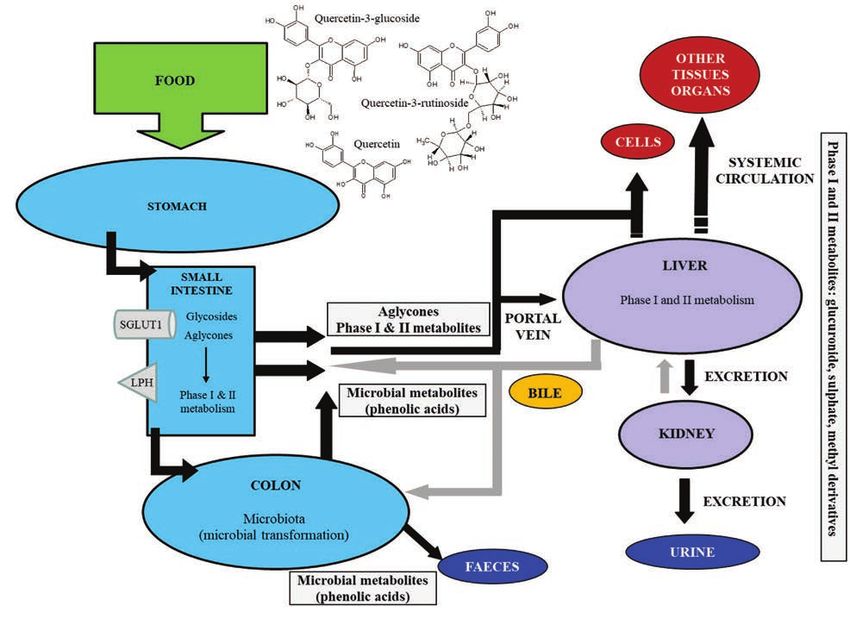
Anticancer activity of quercetin
INTRODUCTION
According to a 2021 report, cancer ranked as
the first or second most common contributor
to mortality across the world and a doubling
of its incidence is predicted by 2070 relative
to 2020 (1). Despite extensive knowledge on
the molecular aspects of this disease, differ-
ent modifications at molecular and genetic
levels in cancer cells result in some difficul-
ties to establish effective anticancer thera-
pies and overcome drug resistance caused by
cancer cells adapting to chemotherapy drugs.
Chemicals occurring in vegetables, fruits, Figure 1. Chemical structure of quercetin.
spices, grains, and other foods have been
found to effectively improve the anticancer
activity and protect against the side effects Quercetin bounded forms with sulfate or alkyl
of conventional anticancer treatments. Quer- residues are instead less present in plants (3).
cetin (3,3’,4’,5,7-pentahydroxyflavone) (figure Quercetin has been proven to play a broad
1) is a ubiquitous dietary bioflavonoid wide- spectrum of anticancer activities such as
ly synthesized in the leaves, flowers, fruits, pro-apoptotic, antiproliferative, antiangio-
and seeds of a variety of food plants (2). In genic, and antimetastatic effects and blocks
plants, the synthesis of this specialized me- an extensive range of cancers including lung,
tabolite proceeds via the combination of the ovarian, prostate, breast, colorectal, bladder
shikimate and acetate metabolic pathways. cancers (4). In this review, we summarize and
Quercetin may occur in plant cells as free analyze the anticancer properties of quercetin
aglycone, but most frequently in the conju- on several systemic tumors in vivo and in vitro
gated water-soluble glycosylated form giving and its main cellular and molecular mecha-
rise to a high number of quercetin derivatives nisms. Quercetin’s chemistry, pharmacokinet-
(3). Quercetin-3-O-glycosides with glucose, ics and safety are also briefly reviewed.
galactose, xylose or rhamnose are the most
common products, as an example quercitrin The basis of chemical structure
(quercetin-3-O-rhamnoside) and rutin (quer- Despite its polyhydroxylated nature, querce-
cetin-3-O-rhamnosyl(1→6)glycoside); the hy- tin is a lipophilic compound scarcely soluble
droxyl group at C7 represents another com- in water; its derivatives can instead have a
mon O-glycosylation site in the molecule. lipophilic or hydrophilic character depend-
Both the hydroxylic functions at C3 and C7 ing on the type and number of functional
are often substituted at the same time, as an groups. Generally, glycosylation increases
example in 3-O-rhamnoside-7-O-glucoside. the hydrophilicity of the molecule, whereas
Quercetin C-glycosides are instead relative- quercetin methyl and alkyl derivatives are
ly rare in plants. Glycosides of quercetin can, lipophilic. Low solubility of the aglycone in
additionally, contain acyl substituents (3). Oth- water is a major challenge for its therapeutic
er frequently occurring derivatives of querce- applications (3).
tin include the formation of ethers (mostly Quercetin polyhydroxylated nature also de-
with methanol to give the corresponding me- termines its bioactivity. The catechol-contain-
thoxy derivatives) and may contain up to five ing B ring (figure 1), the 2,3-double bond
ether groups in various configurations; these in conjugation with the carbonyl function at
compounds can also have sugar substituents. C4 in the C ring and the hydroxyl substitu-
497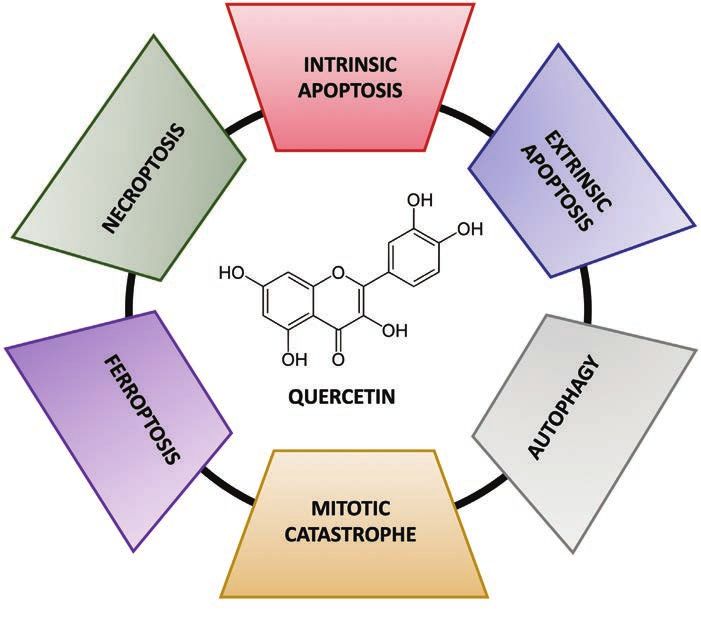
S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
ents at C3 and C5 positions are important can be transported into the enterocytes by
reactive centers in the molecule. Quercetin the sodium-glucose cotransporter (SGLUT1,
gets easily oxidized and, primarily through with a higher affinity for quercetin glucosides
the oxidation of its catechol group, can form than other glycosides), and then deglycosylat-
a quite stable reactive free o-semiquinone ed by cytosolic hydrolases (figure 2). More-
radical acting as a reactive oxygen species over, quercetin glycosides are substrate for
(ROS)-scavenger. It can further undergo au- lactase phlorizin hydrolase (LPH, a family of
to-oxidation and originate bioactive electro- β-glucosidases), a luminal brush border en-
philic tautomeric quinones that are able to zyme that catalyzes the deglycosylation and
bind to nucleophilic amino acid residues at enables the aglycone to pass through entero-
active site of target enzymes. Moreover, the cyte membranes via passive diffusion (figure
presence of the five hydroxyl groups enables 2). Again, the absorbed aglycone and its me-
quercetin to have chelating sites and form tabolites are transported passively or actively,
complexes with metal ions (especially cop- respectively, into the hepatic portal vein. The
per and iron) thus inhibiting the metal-medi- absorbed aglycone bound to serum albumin
ated generation of free oxidizing radicals (5). and the metabolites are transported to the
Quercetin is more active than its correspond- liver, where the aglycone is further metabo-
ing glycosides, while the flavonoid-metal lized (figure 2). Metabolism involves phase I
complexes with a lower redox potential have and phase II reactions in enterocytes and he-
a higher antioxidant potency than the free patocytes to produce water-soluble deriva-
aglycone (3, 5). tives (methyl, glucuronide, and sulfate conju-
gates), which are ultimately transported into
the systemic circulation for distribution to tar-
PHARMACOKINETICS get tissues (figure 2). The high levels of quer-
AND BIOAVAILABILITY cetin-conjugated metabolites detected in the
The chemical structure of the aglycone and bile indicate enterohepatic recirculation (fig-
the type and position of the sugar moiety are ure 2). Indeed, some metabolites are excret-
major determinants of absorption of querce- ed into the small intestine through this route
tin, as well as the composition of the diet, with and reabsorbed as aglycones after deconju-
important consequences on pharmacokinetic gation catalyzed by bacterial β-glucuronidase
parameters. For instance, dietary fat enhanc- and sulfatase. Noteworthy, after enterocyte
es the micellization of quercetin at the small absorption, quercetin, its glycosides and their
intestine (table I) and increases its absorption metabolites can be effluxed back into the in-
and bioavailability (6). testinal lumen by the multidrug resistance-as-
Quercetin occurs in crystalline form at body sociated pumps (7).
temperatures and is relatively lipophilic with Finally, quercetin glycosides not absorbed in
low water solubility, poor bioaccessibility, and the small intestine reach the large intestine
low oral bioavailability (table I). For this rea- where they are fermented by the colonic mi-
son, quercetin is often formulated as nanopar- crobiota (Bacteroides fragilis, Eubacterium
ticles to improve its poor solubility and bio- ramulus, Clostridium perfringens, Bacteroi-
availability. In the small intestine, quercetin des JY-6, Bifidobacterium B-9, Lactobacillus
aglycone passively diffuses from the intesti- L-2, and Streptococcus S-2), which degrades
nal lumen into the enterocytes where it is ei- quercetin aglycone into other phenolic com-
ther directly absorbed (via passive diffusion) pounds (mainly homoprocatechuic acid, pro-
or metabolized before absorption (via active tocatechuic acid and 4-hydroxybenzoic acid)
transport) into the hepatic portal vein (figure by ring cleavage at the heterocyclic C-ring
2). Differently from the aglycone, glycosides (figure 2) (8).
498
Anticancer activity of quercetin
Table I. Intervention studies on quercetin pharmacokinetics involving healthy subjects consuming food sources
of quercetin or treated with quercetin as pure compound.
Food source/aglycone/ Quercetin dose Plasma Urinary Reference
glycoside pharmacokinetics excretion/
concentration
Black tea (1600 mL/day) 49 mg (glycosides) NM 0.5% (123)
Onions (129 g/day) 13 mg (glycosides) NM 1.1%
Black tea (375 mL/day) 13.7 mg Cmax = 0.026 µM 0.252 µM (124)
Onions (50 g/day) 15.9 mg Cmax = 0.053 µM 0.509 µM
Red wine (750 mL/day) 14.2 mg Cmax = 0.026 µM 0.371 µM
Onions (NR) 68 mg Cmax = 0.74 µM 1.39% (125)
Tmax = 0.7 ± 1.1 h
T1/2 = 28 ± 92 h
AUC(0-36h) = 7.71 µM h
Apple sauce + peel (NR) 98 mg Cmax = 0.30 µM 0.44%
Tmax = 2.5 ± 0.7 h
T1/2 = 23 ± 32 h
AUC(0-36h) = 3.5 µM h
Rutin 100 mg Cmax = 0.30 µM 0.35%
Tmax = 9.3 ± 1.8 h
T1/2 = ND
AUC(0-36h) = 3.3 µM h
Onions (160 g) 100 mg Cmax = 2.31 µM 6.4% (126)
Tmax = 0.68 ± 0.22 h
T1/2 = 10.9 ± 4.1 h
AUC(0-24h) = 32.1 µM h
Quercetin‐4′‐O‐glucoside 100 mg Cmax = 2.12 µM 4.5%
Tmax = 0.70 ± 0.31 h
T1/2 = 11.9 ± 4.0 h
AUC(0-24h) = 27.8 µM h
Buckwheat tea (NR) 200 mg Cmax = 0.64 µM 1.0%
Tmax = 4.32 ± 1.83 h
T1/2 = 10.3 ± 3.5 h
AUC(0-36h) = 12.6 µM h
Rutin 200 mg Cmax = 0.32 µM 0.90%
Tmax = 6.98 ± 2.94 h
T1/2 = 11.8 ± 3.1 h
AUC(0-36h) = 8.3 µM h
Onions (100 g) 47 mg NM 1.17 µM (127)
Aglycone 544 mg NM 1.69 µM
Quercetin‐3‐O‐glucoside 151 mg Cmax = 5.0 µM 3.0% (128)
Tmax = 0.62 ± 0.2 h
T1/2 = 18.5 ± 0.8 h
AUC(0-72h) = 19.1 µM h
Quercetin‐4′‐O‐glucoside 154 mg Cmax = 4.5 µM 2.6%
Tmax = 0.45 ± 0.08 h
T1/2 = 17.7 ± 0.9 h
AUC(0-72h) = 18.9 µM h
Rutin 94 mg Cmax = 0.18 µM h NM (129)
Tmax = 6.0 ± 1.2 h
T1/2 = 28.1 ± 6.4 h
AUC(0-∞) = 3.7 µM h
499S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
Food source/aglycone/ Quercetin dose Plasma Urinary Reference
glycoside pharmacokinetics excretion/
concentration
Quercetin‐4′‐O‐glucoside 94 mg Cmax = 3.5 µM h NM
Tmax = less than 0.5 h
T1/2 = 21.6 ± 1.9 h
AUC(0-∞) = 18.8 µM h
Quercetin aglycone 1500 mg Cmax = 5.1 10-5 µM h 1.18% (130)
(daily per 1 week) Tmax = 3 h
T1/2 = 3.47 h
AUC(0-∞) = 0.21 µM h
Quercetin aglycone with 1095 mg Cmax = 1.1 µM NM (6)
fat-free muffin (< 0.5 g fat) Tmax = 5.7 h
Quercetin aglycone with low- Cmax = 1.24 µM
fat muffin (4.0 g fat) Tmax = 5.4 h
Quercetin aglycone with fat- Cmax = 1.6 µM
free muffin (15.4 g fat) Tmax = 6.7 h
Cmax: maximal plasma concentration; Tmax: time to reach Cmax; T1/2: elimination half-time; AUC: area under plasma concentration-time curve;
NR: not reported; NM: not measured.
Figure 2. Major processes involved in oral bioavailability of quercetin in humans (see the text for details);
SGLUT1: sodium-glucose cotransporter; LPH: lactase phlorizin hydrolase (adapted from Varoni et al., 2016
(122)).
500Anticancer activity of quercetin
In conclusion, quercetin bioavailability is cell-cycle blockage (11). Moreover, an in vivo
poor after a single oral intake and is char- study revealed that the anticancer activity of
acterized by high interindividual variability quercetin against thioacetamide-induced HCC
in its metabolism. According to pharmacoki- in rats was due to the inhibition of casein ki-
netics data (table I), it seems that glucosides nase-2α (CK2α), which in turn induced the sup-
from onions possess a higher absorption rate pression of cyclin D1 and Ki-67, pivotal mark-
compared to the glycosides from apples, tea, ers of proliferation (12). A study performed in
red wine or aglycones. Many factors may af- cervical cancer cell line (HeLa) evidenced that
fect quercetin absorption including food ma- quercetin anti-proliferative activity may be due
trix, dietary fat, vitamin C status, sugar moi- to the downregulation of phosphatidylinositol
eties, genetic polymorphisms, composition 3-kinase (PI3K), mitogen-activated protein ki-
of gut microbiota, taking drugs, body mass nase (MAPK), and WNT pathways, leading to
index, lifestyle, and health status. Howev- cell-cycle arrest in G2/M phase (13). Further-
er, there is no clear evidence demonstrating more, quercetin blocked the cell cycle at the
that gender and age affect quercetin bio- sub-G1 phase in gemcitabine-resistant pancre-
availability, although the existing studies in- atic cancer cells (MIA Paca-2 GEMR), via a mech-
volved a small number of volunteers. In any anism involving the reduction of the receptor
case, larger studies (N ≥ 20) are warranted for advanced glycation end products (RAGE),
to accurately evaluate bioavailability of quer- highly expressed by cancer cells, and the inhi-
cetin, as well as more research is needed to bition of the PI3K/Akt/mTOR axis (14). In ad-
develop (nano)formulations for improving its dition, quercetin enhanced the effect of pacl-
absorption and efficacy. itaxel in prostate cancer cells (PC-3), where it
significantly inhibited cell proliferation and ar-
rested cell cycle at the G2/M phase (15). Final-
GENERAL MECHANISMS ly, Soll and co-workers showed that quercetin
hindered melanoma cells (B16) proliferation
Induction of anti-proliferative effects equal to etoposide, by increasing sub-G1 pop-
Cancer progression is due to the uncontrolled ulation, suggesting apoptotic cell death (16).
growth of malignant cells, unable of either un- The reported evidence summarized in table
dergoing apoptosis or senescence. Therefore, II suggests the anti-proliferative and cell-cy-
the arrest of both proliferation and cell-cycle cle blocking properties of quercetin in a wide
progression represents a relevant target of an- plethora of cancers both in vitro and in vivo.
ticancer drugs. Quercetin elicits antitumor ef-
fects in different in vitro and in vivo models (9). Induction of cell death
In this regard, it has been demonstrated that Quercetin exerts its anticancer activity by pro-
quercetin suppressed hepatocellular carcinoma moting multiple cancer cell death mechanisms
(HCC) cell proliferation in a concentration- and (figure 3). Cell death is classified into two main
time-dependent manner as well as induced categories: programmed and unscheduled, as
cell-cycle arrest at different phases, in relation necrosis. Apoptosis is the most known and
to the cell line employed. Among the 13 HCC characterized programmed cell death (PCD)
cell lines tested, quercetin blocked cell cycle in mechanism. It is caspase-dependent and re-
G0/G1, S or G2/M phases in 4, 2 and 6 lines, lies upon two main pathways: the intrinsic (or
respectively (10). In HepG2 cell lines, querce- mitochondrial) and the extrinsic (or death re-
tin reduced cell proliferation along with lower- ceptor) pathway (17). A plethora of studies re-
ing the expression of the checkpoint kinase-1 ported quercetin’s ability to promote intrinsic,
(CHEK1), leading to the downstream regula- extrinsic or both apoptotic pathways in multi-
tion of both cyclins A and E and hence the ple cancer cell models including breast, colon,
501S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
Table II. Effects of quercetin on cell-cycle progression.
Time of Experimental
Effects on Cell Cycle Concentration/dose Reference
exposure model
Blockage of G0/G1, S or G2/M
25-100 µM 48 h 13 HCCa cell lines (10)
phases
Reduction of CHK1 and
20-100 µM 48 h HepG2 cells (11)
cyclins A/E
Block in G2/M phase via
downregulation of PI3K, 25-50 µM 24-48 h HeLa cells (13)
MAPK and WNT pathways
Sub-G1 population increase,
via RAGE involvement and MIA Paca-2 GEMR
25-200 µM 48 h (14)
modulation of PI3K/AKT/ cells
mTOR axis
Reduction of BTG2, p21 and 11.39 µM (+ 2.85 µM of
48 h K562 cells (131)
p27 expression curcumin)
5 days/week Thioacetamide-
Fall of CK2α, cyclin D1 and
100 mg/kg (8 weeks in induced HCCa in (12)
Ki-67 expression
total) rats
Increase of cells in sub-G1 50 mg/mL 24-48 h B16 cells (16)
Arrest of cell cycle at the
20 µM 24 h PC-3 cells (15)
G2/M phase
a
Hepatocellular carcinoma.
Figure 3. Schematic representation of quercetin-induced cell death mechanisms.
502Anticancer activity of quercetin
liver, lung and gastric cancer, melanoma, gli- with quercetin (10-60 μM) displayed monop-
oma, and leukemia, both in vitro (20-350 µM) olar and multipolar spindles, misaligned and
and in vivo (10-500 mg/kg) (9, 18, 19). mis-segregated chromosomes (26), all fea-
Apart from apoptosis, also autophagy rep- tures related to MC (25). In addition, polyploi-
resents a PCD mechanism. Although in most dy, cell enlargement, and multinucleation, ob-
cases autophagy acts as a protective cellu- served in A549-treated cells, further confirmed
lar process, thus eventually promoting tumor the induction of mitotic catastrophe by quer-
progression, in other circumstances autopha- cetin (26).
gy can lead to cell death (17). As recently re-
viewed, quercetin promoted in vitro (12.5-160 Inhibition of cancer cells’ migration
µM) and in vivo (50-120 mg/kg) autophagic and invasion
cell death and/or cytoprotective autophagy in The progression of cancer development may
leukemia, glioma, gastric, breast, ovarian (20), lead to the acquisition by tumor cells of the
and lung cancer (21). ability to invade nearby tissues and form me-
Accumulating evidence points out that mul- tastases in other organs, representing so far
tiple non-apoptotic forms of PCD also called one of the greatest challenges in oncology
non-canonical, can be triggered through in- (27). Quercetin was reported to block both
dependent apoptosis when the apoptot- migration and invasion of non-small cell lung
ic process is altered or inhibited. Ferropto- cancer cells, in vitro and in a xenograft mice
sis is an iron-dependent non-canonical PCD model, through the inhibition of the Src-medi-
mechanism, driven by the accumulation of ated Fn14/NF-κB pathway, known to promote
lipid peroxides. In hepatocellular carcinoma cell survival and metastatic capabilities of can-
HepG2 cells, quercetin (50 μM) promoted cer cells (28). Moreover, Guo and co-workers
both apoptosis and ferroptosis (22). Indeed, showed that quercetin hindered metastasis of
lysosome-dependent cell death induced by pancreatic ductal adenocarcinoma cells, both
quercetin was associated with the promotion in vitro and in vivo, via the inhibition of son-
of ferritin degradation (22), a process known ic hedgehog and tumor grow factor (TGF)-β/
as ferritinophagy, which enhances the cellular Smad signaling pathways, that in turn brought
free iron content, thus increasing lipid perox- to a suppression of the epithelial-mesenchy-
idation and promoting ferroptotic cell death mal transition (EMT) of these cells. This process
(23). Necroptosis, instead, relies on the acti- involves the acquisition of the mesenchymal
vation of RIPK1 (receptor interacting serine/ stem cells phenotype by epithelial ones and, in
threonine kinase 1) that leads to the formation pathological conditions, it leads to resistance
of the so-called necrosome complex. Notably, and migration of cancer cells (29). The effect
in MCF-7 breast cancer cells, quercetin (50 on EMT was observed also both in skin squa-
μM) suppressed cell proliferation by promot- mous carcinoma, where quercetin inhibited
ing both apoptosis and necroptosis. Indeed, migration and invasion through the reduction
the inhibition of RIP1K by necrostatin-1 as well of Src/STAT3/S100A7 signaling pathway (30),
as the inhibition of apoptosis by Z-VAD-fmk (a and in different oral squamous carcinoma cells,
pan-caspase inhibitor) restored MCF-7 cell via- where its treatment negatively affected both
bility, pointing out quercetin’s ability to trigger matrix metalloproteinase (MMP) and TGF-β1
these two cell death pathways (24). expression (31). In this last type of cancer,
Lastly, mitotic catastrophe (MC) is a type of quercetin also hampered migration and inva-
PCD occurring during mitosis because of se- sion by regulating micro-RNA-16 and homeo-
vere DNA damage, impaired mitotic machin- box A10, relevant in the correct regulation of
ery, and/or failure of mitotic control points (25). cell proliferation (32). The anti-metastatic prop-
A549 non-small cell lung cancer cells treated erties of quercetin have been also evaluated
503S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
in osteosarcoma cell lines, in which it reduced the PI3K/Akt pathway together with MAPK one
the expression of MMPs and increased that of (36). Another relevant pathway in metastasis is
their tissue inhibitors (TIMPs), along with affect- JAK/STAT one, which was demonstrated to be
ing the parathyroid hormone receptor-1, a typ- affected by quercetin in hepatocarcinoma cells
ical biomarker of metastatic osteosarcoma cells both in vitro and in vivo (37). Additionally, Lu
(33). MMPs and cadherins, markers of mesen- and collaborators showed that quercetin treat-
chymal and epithelial cells, respectively, have ment decreased the expression of Twist2 and
been shown to be modulated by quercetin in EpCAM and increased that of E-cadherin, oth-
estrogen-receptor (ER) positive breast carcino- er pivotal factors involved in EMT, in prostate
ma (BRC) cells, along with potentiating antitu- cancer cells resistant to docetaxel (38). Over-
mor activity of tamoxifen (34), as well in ovarian all, these reports, summarized in table III, sup-
metastatic cancer cells, where it inhibited also port the remarkable value of quercetin as an
PI3K/Akt, Ras/Raf pathways, EGFR, and clau- anti-metastatic agent, given its capability to
dins expression, therefore polarizing cells to- hamper EMT transition by simultaneously tar-
wards the epithelial state (35). Triple-negative geting both membrane and intracellular signal-
BRC cells have been employed to study the ing pathways leading to the up-regulation of
anti-migration and anti-invasion properties of multiple factors involved in invasion and migra-
quercetin, demonstrating to be an inhibitor of tion of cancer cells.
Table III. Main migration- and invasion-related pathways affected by quercetin in different cell lines.
Migration-/invasion-related Time of
Concentration Type of cancer Reference
pathways exposure
Decrease of MMPs, cadherins,
EGFR and claudins expression; Ovarian metastatic
50-75 µM 24 h (35)
blockage of PI3k/Akt and Ras/Raf cancer
pathways
Inhibition of the Src-mediated Non-small cell lung
100 µM 48 h (28)
Fn14/NF-κB cancer
Inhibition of Src/STAT3/S100A7; Skin squamous
20-40 µM 24 h (30)
blockage of EMT carcinoma
Inhibition of hedgehog and TGF-β/ Pancreatic ductal
10-100 µM 24-48 h (29)
Smad; blockage of EMT adenocarcinoma
Decrease of MMP and TGF- β1 Oral squamous
40 µM 24 h (31)
expression carcinoma
Suppression of MMPs, TIMPs and
20-100 µM 48 h Osteosarcoma (33)
parathyroid hormone receptor-1
Decrease of Twist2 and EpCAM
Docetaxel-resistant
and increase of E-cadherin 10 µM 48 h (38)
prostate cancer
expressions
Inhibition of PI3K/AKT and MAPK Triple-negative
25-50 µM 24-48 h (36)
pathways breast cancer
Blockage of JAK/STAT pathway 80-120 µM 12-36 h Hepatocarcinoma (37)
Estrogen-receptor
Inhibition of MMPs and cadherins
5-100 µM 48 h positive breast (34)
expression
carcinoma
Regulation of micro-RNA-16 and Oral squamous
25-100 µM 24-48 h (32)
homeobox A10 carcinoma
504Anticancer activity of quercetin
Epigenetic mechanisms quercetin (0.5-100 μM) in lung adenocarcino-
Recent research has shown that post-tran- ma, osteosarcoma, and ovarian cancer cells,
scriptional histone modifications, changes in respectively. In particular, miR-145 was shown
DNA methylation status, and regulation of to inhibit and control target genes involved in
non-coding RNAs can alter gene expression apoptosis, thus demonstrating the ability of
and modify the development of several types quercetin to influence micro-RNA expression
of cancer (39). patterns related to cancer (45).
Studies using human xenografts and acute my- An interesting study reported that quercetin
eloid leukemia cell lines (HL60 and U937) have (20 µM) increases the efficacy of bromodomain
shown that treatment with quercetin (50 μM) and extraterminal domain inhibitors by sup-
downregulates histone deacetylase I (HDA- pressing the heterogeneous nuclear ribonu-
CI) protein levels, leading to DNA demethyl- cleoprotein A1, a nuclear protein, controlling
ation and accumulation of acetylated histones mRNA translation and transport, as well as by
3 and 4 in the promoter regions of genes in- decreasing survivin, an antiapoptotic protein
volved in apoptosis pathways, leading to their (46), in several tumor cell lines. An in-depth
transcriptional activation (40). Recently, a novel study was carried out to understand the mech-
chitosan-based quercetin nanohydrogel (ChiN- anism of action of quercetin by quantifying the
H/Q) has been reported to reduce the inhibi- biochemical activity of DNA methyltransferas-
tion of DNA methyltransferases (DNMT1/3A/3) es, HDACs, histone methyltransferases, and
and increase DNA methylation in HepG2 can- methylation of oncosuppressor gene selec-
cer cells with a half maximal inhibitory con- tions and global genomic DNA methylation on
centration (IC50) of 331 µM (41). Moreover, treated HeLa cells. Enzymatic assays showed
overexpression of the enzyme HDAC8 is an- that quercetin 25 and 50 μM modulates these
other important epigenetic alteration that has activities in a dose-dependent manner, while
been reported in colon cancer. Its inhibition molecular docking studies suggested that
in HCT116 cells by quercetin (IC50: 181.7 µM) quercetin could be a competitive inhibitor by
leads to increased acetylated H3K9 (histone H3, interacting with residues within the catalytic
lysine 9) and apoptosis through the activation cavity of several DNA methyltransferases and
of caspase-3/-7 (42). Regarding the control of HDACs (47).
post-transcriptional mechanisms, experimental Future research has the potential to expand
studies suggested that quercetin 50 μM mod- the use of dietary-based polyphenols, such as
ulates the expression of DBH-AS1, which is an quercetin, in treating cancer, especially in com-
important epigenetic reader in cancer therapy. bination with conventional drugs. Their use
Moreover, quercetin (0.87-7.79 μM) inhibition of could be an alternative and effective method
carbonic anhydrase isoforms (CA II, CA IX, and in cancer therapy leading to restoration of sev-
CA XII) was observed in previous cancer stud- eral aberrant epigenetic alterations.
ies (43). Recently, intravenous administration of
quercetin-modified metal-organic frameworks Angiogenesis
(Zr-MOF-QU) (50 mg/kg), a novel type of Zr- The ability to induce the formation of new
MOF nanoparticles, showed excellent efficiency blood vessels is one of the peculiar activities
for CA IX inhibition in tumor-bearing mice (44). of cancer cells, which allows them to better
Several in vitro studies showed that querce- access nutrients and oxygen and constitutes
tin was able to upregulate the miR-let 7 mi- an excellent way of tumor dissemination (48).
cro-RNA family in pancreatic ductal adeno- Quercetin modulates different cell signaling
carcinoma cells, thus interfering with K-Ras’s pathways involved in angiogenesis (figure
pathways. Furthermore, miR-16, miR-217, 4). The modulation of vascular endothelial
and miR-145 were found to be modulated by growth factor (VEGF) pathway seems to be
505S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
Figure 4. Major angiogenesis signaling pathways modulated by quercetin.
especially involved in this effect, consider- GSK-3β downstream signalling molecules on
ing its well-known key role in survival of en- human breast cancer cell lines (MCF-7 and
dothelial cells and related tumor angiogen- MDAMB-231) and the expression of VEGFR-2
esis (49). In HUVEC cells, quercetin (10-40 in HUVECs. Similar results were recorded on
µM) blocked the VEGF-mediated phosphor- chick embryos, where the formulation at 50
ylation of VEGF receptor 2 and its down- μM inhibited neovascularization.
stream protein kinases Akt, mTOR, and ribo- Analogously, Lupo et al. suggested that quer-
somal protein S6 kinase (50). Similar results cetin and its permethylated form at 25 μM in-
were recorded in PC-3 prostate cancer cells, hibited cell viability and migration, downregu-
where quercetin (10-40 µM) inhibited the se- lated VEGFR-2 and reduced Akt, ERK and JNK
cretion of VEGF (50). Moreover, intraperito- levels on human primary endothelial cells iso-
neal administration of quercetin (20 mg/kg/ lated from retinal microcapillaries (HREC). Sim-
day) inhibited the activation of Akt, mTOR ilar results were obtained on an ex vivo mod-
and P70S6K proteins and led to decreased el of rabbit aortic ring, where quercetin and
tumor’s weight and volume in a murine pros- its permethylated form disrupted microvessels
tate xenograft model (50). formation (52).
Balakrishnan et al. (51) studied the antiangio- A study performed on an abdominal aortic
genetic potential of a gold nanoparticle-based aneurysm (AAA) mouse model also demon-
delivery system. The formulation (50-100 μM) strated that quercetin (60 mg/kg) decreased
was able to inhibit MMP-2 and MMP-9 ac- neovascularization and the expression of pro-
tivity as well as to reduce the expression of angiogenic mediators, including VEGF-A, in-
p-EGFR/VEGFR-2 and p-PI3K/Akt/p-Akt/p- tercellular adhesion molecule 1 (ICAM-1), vas-
506Anticancer activity of quercetin
cular cell adhesion molecule 1 (VCAM-1) and HER2-overexpressing (BT-474) BRC cell line,
vascular endothelial cadherin, and inhibited quercetin (20-60 µM) activated the extrin-
the expression of cyclooxygenase-2 (COX-2) sic apoptotic pathway. In MDA-MB-231 cells,
and hypoxia-inducible factor 1α (HIF-1α), as- quercetin modulated the Akt/AMPK/mTOR
sociated with the upregulation of VEGF-A. The pathway (57) and, besides the effects on sig-
same study showed that quercetin-3-O-glu- naling proteins, at 20 µM it increased the activi-
curonide, a quercetin major circulating me- ty of several cell-cycle regulatory proteins such
tabolite, downregulated COX-2, HIF-1α and as p53, p21, and GADD45 (58). In the same
VEGF-A expression and matrix metallopro- cell line, quercetin (20-80 µM) inhibited aer-
teinases MMPs activities in vascular smooth obic glycolysis via impairing PFKP-LDHA axis
muscle cells isolated from AAA mice after 72h (59). Those results were confirmed in a xeno-
treatment at a concentration of 50 μM (53). graft BRC model, where quercetin (50 mg/kg
A similar decreased expression of proangio- twice daily intraperitoneally for a month) sup-
genic mediators and metastasis-associated pressed glycolysis and tumor metastasis (60).
factors, such as VEGF-A, VEGFR-2, COX-2, Quercetin 100 µM was also able to inhibit mi-
E-cadherin, Twist1 gene and integrin ITGβ6, gration and invasion of BRC stem cells via AL-
was observed in a human gastric cancer xeno- DH1A1, CXCR4, and EpCAM downregulation
graft mouse model after treatment with quer- (61). The exposure of BRC cell lines to querce-
cetin (20 mg/kg) or even more with its com- tin (50 and 100 µM) reduced the expression of
bination with the antitumoral drug irinotecan SLUG, SNAIL, and Twist transcription factors,
(54). It was also found that low concentration while up-regulated E-cadherin, thus suggest-
of quercetin (10 μg/mL) inhibited tube for- ing that quercetin can act as an EMT inhibitor
mation in HUVECs treated with conditioned (58). In a study by D’Arrigo et al. performed
medium obtained from U251 glioblastoma on MCF-7 (wild-type p53) and T47D (mutant
cells possibly by downregulating VEGF-A and p53) cells (62), quercetin, like other flavonoids,
MMP-2 and MMP-9 protein levels (55). showed a relevant degree of complementari-
Finally, the intravenous administration of poly- ty with estrogen and androgen receptors (62)
mer micelle-nanoencapsulated quercetin (60 and inhibited the survival of ER+ tumor-initiat-
mg/kg) significantly suppressed the growth ing cells (63). Moreover, an in silico and in vitro
of xenograft A2780S ovarian tumors in athy- screening indicated that quercetin had also a
mic nude mice through a significantly inhibi- high binding affinity for the cyclin-dependent
tion of microvessel density. Moreover, treating kinase 6 (CDK6) and inhibited 50% of its AT-
A2780S cells with quercetin (0-30 μg/mL, 48 h) Pase activity at 5.89 μM (64).
in vitro produced higher levels of phosphory- A novel nanoformulation of quercetin com-
lated p44/42 MAPK and phosphorylated Akt, posed of hyaluronic acid, copper ion, chelated
which are critical intracellular mediators of an- dextran-aldehyde was tested on MDA-MB-231
giogenesis (56). cells and BRCA-mutant TNBC HCC1395 cells.
The formulation was highly cytotoxic for
HCC1395 cells, where it induced DNA dam-
EFFECT OF QUERCETIN age and apoptosis. In HCC1395-tumor-bear-
ON THE MOST COMMON CANCERS ing nude mice, treatment with the nanofor-
mulation induced a decrease in tumor volume
Breast cancer higher than that observed for quercetin (65).
The anticancer activity of quercetin on breast As a potent heat shock protein (HSP) 70 in-
cancer (BRC) is mediated via regulation of hibitor, quercetin (50 µM) was used as sensi-
various signaling pathways, but the exact tizer in a new modulated electro-hyperthermia
mechanism of its action remains elusive. In a treatment. On 4T1 murine BRC cells, querce-
507S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
tin synergistically decreased cell viability with quercetin at 50 and 75 µM decreased viabili-
respect to the two single pharmacological ty and induced apoptosis on human metastat-
strategies (66). As a P-gp inhibitor, nanofor- ic PA-1 cells, as indicated by Bcl-2 and Bcl-xL
mulated quercetin plus paclitaxel was tested decrease and caspase-3, caspase-9, Bid, Bad,
both in vitro and in vivo. In MCF-7 cells, quer- Bax, and cytochrome c increase (71). A recent
cetin 33 μM decreased the efflux of paclitaxel study tested the antitumor effects of querce-
and synergistically enhanced its cytotoxicity; tin encapsulated into monomethoxy poly(eth-
in xenograft MCF-7 BRC, intravenous adminis- ylene glycol, PEG)-poly(ε-caprolactone) mi-
tration of quercetin (5.1 mg/kg) decreased tu- celles (56). The nanoformulation (60 mg/kg)
mor weight without toxicity to normal tissues significantly inhibited tumor volume, induced
(67). Moreover, the inhibitory effects of quer- apoptosis and strongly inhibited angiogen-
cetin on CYP450 enzymes were exploited to esis of xenograft A2780S ovarian tumors. Of
improve mycophenolic acid’s anticancer activ- note, the nanoformulation was well tolerated,
ity on 7,12-dimethylbenz(a)anthracene-treated as indicated by no changes in animals’ body
rats (68). In conclusion, the pleiotropic activi- weight (56).
ty of quercetin against BRC and in particular Furthermore, the effect of a quercetin-PEGylat-
its ability to sensitize cancer cells and coun- ed liposomal formulation was investigated in
teract drug resistance makes it a promising sensitive and cisplatin-resistant A2780 ovarian
candidate for well-designed oncological clin- cancer cells. The formulation (50 µM) caused
ical trials. apoptosis and G0/G1 and G2/M arrest, as well
as inhibited cell proliferation of both clones. In
Ovarian and prostate cancers vivo studies performed on xenograft sensitive
Several studies have been conducted to elu- or cisplatin-resistant A2780 ovarian tumors,
cidate the molecular mechanisms of querce- found that the formulation (50 mg/kg) blocked
tin’s activity on ovarian cancer (OC). Quercetin tumor growth in both mice models (72).
inhibited cells’ growth and promoted apop- On prostate cancer (PC) LNCaP cells, querce-
tosis in a concentration-dependent manner tin 100 µM decreased Bcl-xL/Bcl-xS ratio and
in different OC cell lines. As an example, its amplified the efflux of Bax to the mitochondri-
proapoptotic activity was observed on A2780S al matrix leading to apoptosis (73). In addition,
cells, where quercetin (0.4-100 µM) activated quercetin (5-100 µM) downregulated HSP90
caspase-3 and -9, reduced the expression of expression and led to growth inhibition and
MCL-1 and Bcl-2, and increased the expression apoptosis of PC-3 cells (74). In the same cell
of Bax. Moreover, on the same cell line, it in- line, quercetin (25-125 µM) inhibited cell via-
hibited cell proliferation through the reduction bility by reducing the mRNA expression of dif-
of phosphorylated p44/42 MAPK and phos- ferent mitogenic factors including insulin-like
phorylated Akt (56). Similar results were record- growth factors (IGF)-I and II and increased that
ed in SKOV3 CDDP3 cisplatin-resistant cells, of IGFBP-3Rbeta, which led to a reduced se-
where quercetin (10-50 µM) interfered with the cretion of IGF-I and II (75).
G2/M phase, even if it did not affect cyclin B1 A recent study reported that quercetin (50-
levels (69). Besides, quercetin 20 µM induced 500 µM) reduced the expression of metasta-
endoplasmic reticulum stress in cisplatin-sen- sis-associated lung adenocarcinoma transcript
sitive OV2008 cells and their resistant variant 1 (MALAT1), which is overexpressed in PC, in-
resulting in mitochondria-mediated apoptosis duced apoptosis, and blocked EMT and inva-
via a p-STAT3/Bcl-2 and caspase-dependent sion and migration of PC-3 cells (38). Those
pathways (70), as demonstrated by Bcl-2 and results were confirmed on a mouse PC-3 xe-
Bcl-xL reduction and caspase-3, -9, Bid, Bax, nograft tumor, where intraperitoneal querce-
Bad, and cytochrome c increase. Similarly, tin (75 mg/kg) targeted MALAT1 and blocked
508Anticancer activity of quercetin
tumor growth (38) and angiogenesis through Due to the Janus face of autophagy, the effect
an increase in thrombospondin-1 protein and of quercetin-activated autophagy on apopto-
mRNA expression (76). sis is complicated, since it could be partner
Several in vivo studies recorded the querce- or opponent (82). A very recent publication
tin’s ability of enhancing the therapeutic ef- unraveled that quercetin (12.5-100 µM) in-
fect of different drugs. The combination of duced pro-apoptotic autophagy in two NS-
quercetin (50 mg/kg) and paclitaxel (5 mg/ CLC LC, A549 and H1299, involving the his-
kg) synergized the inhibition of tumor growth tone/protein deacetylase sirtulin 1 (SIRT1) and
induced by paclitaxel alone in a mouse PC-3 its downstream effector AMPK (AMP-activated
xenograft tumor (15). Moreover, quercetin (75 protein kinase), potent stimulators of cellular
mg/kg) synergized with 2-methoxyestradiol in autophagy (21).
inhibiting tumor growth, increasing Bax/Bcl-2 Cytoskeleton components, such as microtu-
ratio and caspase-3 activation, and reducing bules, microfilaments and vimentin, are key
microvessel density in both androgen-depen- targets for anticancer treatment, due to their
dent LNCaP and androgen-independent PC-3 importance in the regulation of mitosis, cell di-
xenograft tumors (77) and was able to reverse vision, cell migration, and cell death (83). Quer-
docetaxel resistance (78). cetin (10-60 µM) impacted on these essential
The cancer preventive effect of quercetin was cytoskeletal elements, disassembling vimentin,
also highlighted in a clinical trial where 433 microfilaments, and microtubules in A549 cells
men with confirmed primary PC and 538 con- and contributing to the failure of cytokinesis,
trols were evaluated in a case-control study: which leads to apoptosis and mitotic catastro-
24 mg intake of quercetin/day reduced PC risk phe (26). Besides, the inhibition by quercetin of
by 27% (79). vimentin and N-cadherin (26), both markers of
EMT, contrasted A549 migration, hence their
Lung cancer metastatic potential. A recent study highlight-
Lung cancer (LC) includes both small cell lung ed that the inhibition by quercetin of NSCLC
cancer and non-small cell LC (NSCLC), despite cells (HCC827) proliferation and migration was
the latter alone is responsible for 85% of tu- also mediated by the Src family kinases (28),
mors in this organ (80). Several in vitro studies high levels of which activate Fn14/NF-κB sig-
reported the antiproliferative and pro-apoptot- naling and promote the metastatic potential of
ic activity of quercetin on LC cells, mediated NSCLC cells (84). Other in vitro (100 µM) and
by both intrinsic and extrinsic pathways (18). in vivo (HCC827 xenografted BALB/c mice in-
For instance, in NSCLC cells quercetin prompt- traperitoneally treated with 100 mg/kg/day
ed mitochondrial depolarization triggering an quercetin for 3 weeks) studies confirmed that
imbalance in the Bax/Bcl2 ratio and downreg- the anti-NSCLC effects of quercetin clearly de-
ulating IL-6/STAT3 signaling pathway. Besides, pended on the inhibition of Src.
quercetin triggered TRAIL-induced apoptosis Receptor tyrosine kinases (RTKs) represent one
by TNF-family death receptors binding (9, 18) of the most frequently deregulated family of
and by the regulation of epigenetic pathways proteins in LC, playing a key role in the con-
(81). In particular, in the p53-mutant H1299 trol of tumor cell proliferation (85). Through a
LC cells, quercetin 5 µM raised p300 expres- computational studies, Baby and colleagues
sion, which is responsible for the acetylation of (86) demonstrated that quercetin mimics the
the lysine residues of histones. Thanks to this interactions of ATP in the active site of RTKs
mechanism, quercetin enhanced the expres- (EGFR, FGFR1, IGF1R and c-Met) leading to
sion of the death receptor DR5 and the anti- inhibition of RTKs overexpression.
cancer effects of HDAC inhibitors (trichostatin A recent in silico screening and in vitro exper-
and vorinostat) (81). iments evidenced that the pro-apoptotic ef-
509S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
fects of quercetin (~ 50 µM) involve the inhi- ylase activity and polyamines biosynthesis ob-
bition of CDK6 (64), the nuclear expression of served in human DLD-1 colon cancer cell (98).
which is negatively associated with the overall Recent studies suggest that quercetin may in-
survival of lung cancer patients (87). crease the cytotoxic effect of standard anti-
Different quercetin-loaded nanoparticles syn- tumoral drugs also in CRC cells. In particular,
ergistically enhanced the anticancer effica- quercetin (33 µM) increased doxorubicin accu-
cy of paclitaxel (88, 89) and gefitinib (90) in mulation and enhanced the cytotoxic effect of
vitro and in vivo. As an example, quercetin doxorubicin on P-gp-overexpressing SW620/
and gefitinib encapsulated into PLGA-PEG Ad300 cells (99). Similar results were recorded
nanoparticles synergistically reduced the IC50 on HT-29 cells, where quercetin (50 µM) en-
values of the single drugs on PC-9 cells (IC50 hanced cisplatin-induced apoptosis. The effect
quercetin-nanoparticles: 2.12 µg/mL; IC50 gefi- was partially due to the inhibition of the acti-
tinib-nanoparticles: 2.57 µg/mL; IC50 quercetin/ vation of NF-κB expression (100).
gefitinib-nanoparticles: 0.67 µg/mL) (90). Quercetin also exerts antiangiogenic effects
in colorectal cancer cells. Since DLD-1 colon
Colorectal cancer cancer cells have been found to release an-
Quercetin has been shown to reduce prolif- giogenic factors, Xiao et al. co-cultured these
eration and to induce apoptosis and cell-cy- cells with HUVECs. At 100 and 200 µM, quer-
cle arrest in a number of colorectal cancer cetin significantly reduced endothelial tube
(CRC) cells, such as HCT116, HT-29, SW 480, formation (101).
SW 620, Caco-2, LoVo, Colo320 DM cells Several murine and rat models have been
(91). Specifically, van Erk et al. (2005) report- used to investigate quercetin effects in CRC.
ed that 5 µM quercetin downregulates key Shree et al. tested the chemopreventive effect
cell-cycle genes (e.g., CDK6, CDK4 and cyclin of quercetin (25 or 50 mg/kg bodyweight) on
D1) in Caco-2 cells (92). Moreover, at 20, 50, 1,2 dimethyl hydrazine (DMH)-induced rat co-
100 and 200 µM it induces apoptosis in HT29 lon cancer. Quercetin significantly improved
cells with a concentration-dependent mech- DMH-induced pathological modifications. In
anism involving the Akt/CSN6/Myc signaling particular, it reduced proliferation and colon
axis (93) and at 200 µM exhibits pro-apoptot- cancer early markers (mucin depletion and
ic effects on Caco-2 and SW620 cell lines via goblet cell disintegration), adenomatous pol-
nuclear factor kappa-B (NF-κB) signaling path- yposis coli and β-catenin, and tumor incidence
way inhibition and Bcl-2 and Bax modulation and multiplicity (102). Similar results were re-
(94). In the same cell line (Caco-2), quercetin corded on N-methyl nitrosourea-induced rat
(20- 100 µM) has been found to induce apop- colon cancer, where quercetin (50 mg/kg)
tosis through the modulation of the apoptotic blocked the overexpression of Wnt5a and
extrinsic pathway (95) and reduce topoisomer- up-regulated the expression of Axin-1, and de-
ase II-induced DNA cleavage (96). An intrigu- creased the serum level of TAG72 and GAL3
ing result is the demonstration that nutrition- in colon cancer bearing rats (103).
ally relevant concentrations of quercetin (0.1 Quercetin suppressed colon carcinogenesis
or 1 µM) mimicked the 17β-estradiol-induced also on mouse colon carcinogenesis induced
apoptotic effect in ERβ1-containing DLD- by azoxymethane/dextran sodium sulfate.
1 colon cancer cell line and activated p38, Quercetin (30 mg/kg) significantly decreased
which leads to caspase-3 activation and PARP multiplicity and size of colon tumors and ex-
cleavage (97). pression of oxidative stress and inflammation
Quercetin (0.1-100 µM) may also alter the me- markers (104).
tabolism of actively proliferating cells through A recent study suggested that 3,4-dihydroxy-
the significant decrease in ornithine decarbox- phenylacetic acid (0.05-200 µM), an antioxi-
510Anticancer activity of quercetin
dant microbiota-derived metabolite of querce- ited BC growth also in nude mice (111). Other
tin, protects against neoplastic mouse colonic studies explored the use of quercetin in asso-
transformation induced by hemin, a metabo- ciation with anticancer therapies with the aim
lite of myoglobin (105). In particular, the me- of overcoming multiple drug resistance (MDR)
tabolite prevented the reduction of apoptosis, phenotype (113). Quercetin (250 and 500 µM)
the increase in ROS levels and nucleic ac- plus gemcitabine 10 µM decreased the ex-
ids’ oxidation, and the decrease in the mito- pression of proteins involved in MDR including
chondrial membrane potential caused by he- the ABCC2 compared to the administration
min exposure. of the individual molecules alone (113). More-
Overall, quercetin exerts chemotherapeutic over, quercetin plus cisplatin (both at 50 μM)
and chemopreventive effects in colorectal can- synergistically reduced T24 and UMUC cell vi-
cer models, which have been demonstrated to ability (114). A recent study documented that
be mediated through various mechanisms, in- a quercetin-zinc complex ≥ 12.5 μM decreased
cluding cell-cycle arrest, increase in apoptosis, viability, cell migration and invasiveness and
antioxidant properties, regulation of signaling increased apoptosis in BC cells, with a mech-
pathways involved in CRC development, in- anism involving down-regulation of pAkt/Akt
hibition of angiogenesis. Taking into account and MT1-MMP protein expression (115). In
that quercetin is present in many commonly addition, quercetin incorporated in sodium or
food items, its beneficial effects on CRC are of zinc titanate nanotubes (both at 25-200 µg/
promise in the light of the well-established re- mL) decreased viability of BC cells and their
lationship between dietary habits and CRC risk. ability to form clones, thus suggesting that
these nanostructures can interfere with cancer
Bladder cancer cell proliferation (116).
The first study on quercetin and bladder can-
cer (BC) cell lines was carried out by Ma et
al. (2006): quercetin 150 and 200 μM inhibited CONCLUSIONS
cell growth, induced apoptosis, and arrested Many in vitro and in vivo studies were per-
cell cycle in G0/G1 phase (100 μM) (106). An formed on quercetin, which documented its
antiproliferative effect of quercetin was also ability of inducing anticancer effects on differ-
observed in human and murine BC cell lines ent tumors and through different mechanisms
(MB49, T24, UMUC3, 253J) via the activation (figure 5) and its great potential in the onco-
of AMPK signaling (IC50: 40-60 μM) (107), but logical field. However, quercetin undergoes a
also via other mechanisms such as alterations complex metabolism, transport, and distribu-
in the extracellular catabolism of nucleotides tion, which may not allow to reach adequate
(108) or activation of K channels (109). Besides, concentrations for pharmacological effects in
quercetin 100 μM downregulated MCT1 activi- target tissues. Thus, appropriate plasma con-
ty and promoted apoptosis in endothelial and centrations in a similar high range such as
T24 BC cells co-culture (110). Chen et al. re- those used on in vitro preclinical models could
ported that isoquercitrin (quercetin-3-O-gluco- be not achieved for quercetin and many of its
side) (ISO) 400 μM inhibited BC cells prolifer- anticancer activities recorded in vitro may not
ation and promoted apoptosis via suppression be attainable in vivo. Nanoformulation-based
of PI3K/Akt survival signaling pathway (111). approaches including liposomes, microemul-
Its antiproliferative effect was also demon- sion, nanoparticles, and solid lipid nanopar-
strated in T24 cells where 20-80 μM of ISO ticles have been developed with improved
caused ROS overproduction and activation of bioavailability and biologic features such as bi-
the AMPK signaling pathway (112). ISO orally phasic, inotropic and lusitropic characteristics.
administered (doses were not indicated) inhib- As an example, the encapsulation of querce-
511S. ADORISIO, M. P. ARGENTIERI, P. AVATO, ET AL.
Figure 5. Schematic representation of the main molecular targets of quercetin.
↑: increase activity/expression; ↓: decrease activity/expression; ALDH1A1: aldehyde dehydrogenase 1 family member A1; Akt: protein kinase
B; AMPK: AMP-activated protein kinase; Bax: Bcl-2-associated X protein; Bcl-2: B-cell lymphoma 2; Bcl-xL: B-cell lymphoma-extralarge; Bcl-xS:
Bcl-x short; Bad: Bcl-2 associated agonist of cell death; Bid: BH3 interacting-domain death agonist; CDK2: cyclin-dependent kinase 2; CDK4:
cyclin-dependent kinase 4; CDK6: cyclin-dependent kinase 6; CHEK1: checkpoint kinase-1; CK2α: casein kinase-2α; COX-2: cyclooxygenase-2;
CSN6: constitutive photomorphogenesis 9 signalosome 6; CXCR4: C-X-C motif chemokine receptor 4; DR: death receptor; EGFR: epidermal
growth factor receptor; EMT: epithelial-mesenchymal transition; EpCAM: epithelial cell adhesion molecule; ERK: extracellular signal-regulated
protein kinase; Fn14: fibroblast growth factor-inducible protein 14; GADD45: growth arrest and DNA damage-inducible 45 protein; GSK-
3β: glycogen synthase kinase 3 beta; HIF-1α: hypoxia inducible factor-1α; ICAM-1: intracellular adhesion molecule-1; IL-6: interleukin-6;
ITGβ6: integrin β6; JAK: Janus kinase; JNK: c-Jun N-terminal kinases; MALAT1: metastasis-associated lung adenocarcinoma transcript 1;
MAPK: mitogen-activated protein kinase; MCL-1: Myeloid Cell Leukemia 1; MMP: metalloproteinase; MT1-MMP: membrane type-1 matrix
metalloproteinase; mTOR: mammalian target of rapamycin; NF-kB: nuclear factor kappa-light-chain-enhancer of activated B cells; p53: tumor
protein p53; p21: cyclin-dependent kinase inhibitor 1A; PARP: poly (ADP-ribose) polymerase; PI3K: phosphatidylinositol 3-kinase; RAGE:
receptor for advanced glycation end products; RPS6K: ribosomal protein S6 kinase; RTKs: receptor tyrosine kinases; P70S6K: phosphoprotein
70 ribosomal protein S6 kinase; S100 calcium binding protein A7; STAT3: signal transducer and activator of transcription 3; TGF: tumor
growth factor; TIMPs: tissue inhibitors of metalloproteinases; TRAIL: tumor necrosis factor (TNF)-related apoptosis-inducing ligand; Twist:
twist homolog; VCAM-1: vascular cell adhesion molecule 1; VEGF: vascular endothelial growth factor; VEGFR2: vascular endothelial growth
factor receptor 2.
tin into biodegradable monomethoxy poly ly exert their effects on tumor cells without
(ethylene glycol)-poly(ε-caprolactone) micelles damaging healthy cells. What emerges from
improved the dispersion in water of quercetin the few studies on normal human cells (lung
and its in vivo anticancer activity (56). Similar embryonic fibroblasts, umbilical vein endothe-
results have been observed with PEGylated lial cells, peripheral blood lymphocytes) is that
liposomal quercetin, which provided a sus- quercetin is able to block proliferation or in-
tained release of quercetin and resulted in an duce apoptosis on cancer cells at concentra-
efficient formulation for in vivo tumor growth tions (< 50 µM) exerting no or little effects on
inhibition (72). healthy cells (117).
A crucial aspect in the oncological field is the However, the safety profile of quercetin is not
identification of compounds able to selective- yet fully understood. Based on its polyphenol
512Anticancer activity of quercetin
structure with high number of hydroxyl groups tentially carcinogenic compound in the Ames
and pi orbitals, quercetin is associated in first test, but long-term animal toxicity studies did
instance with antioxidant properties. Taking not confirm its carcinogenic potential (121).
into account the presence of a hydroxyl group These controversial results could be due to the
in position 3 that is subject to tautomerism high non-physiological concentrations used in
and two hydroxyl groups on the C-ring that are the in vitro studies, which are often performed
subject to oxidation, quercetin can also lead to at high concentrations of quercetin, ranging
the formation of highly reactive quinones. Qui- from 25 μM to 200 μM, and do not take into
nones can react with thiols potentially causing account the complex pharmacokinetic profile
DNA and protein damage (118). Antioxidant of quercetin.
and pro-oxidant effects of quercetin depends All in all, quercetin is a well-studied com-
on its cellular concentrations and on the cellu- pound with a broad range of biologic advan-
lar levels of reduced GSH: low concentrations tages. A peculiar characteristic of quercetin
of quercetin increase the antioxidant capaci- is its ability to interact with multiple cellular
ty of cells; higher concentrations of quercetin targets and modulate the activity of several
reduce antioxidant capacity and GSH content signaling pathways. Taking into account that
leading to cellular damage (119). With particu- they include key proteins within the same
lar regard to dietary supplementation, human signaling network, quercetin may act with a
intervention studies did not report pro-oxida- pleiotropic, multilevel and synergistic mech-
tive effects of quercetin at doses of 500-1000 anism of action. Thus, quercetin’s broad tar-
mg/day (i.e., a high daily supplementation) ad- get profile may represent a useful therapeutic
ministered up to 12 weeks, but it is still unclear strategy to tackle one of the most complex
whether quercetin could evoke pro-oxidative dynamic human diseases like cancer. Further
effects in humans after a long-term use (120). in vivo studies and sound clinical trials are re-
Similarly, chronic toxicity animal studies evi- quired to definitely assess its safety and effi-
denced potential nephrotoxic effects for quer- cacy and grant a fully understanding of quer-
cetin, but human intervention studies rarely cetin’s therapeutic potential both alone and
recorded adverse effects following supple- in combination with standard anticancer che-
mental quercetin intake, but no safety data motherapy.
are available after long-term use (> 12 weeks)
of high quercetin doses (≥ 1000 mg) (120).
Based on in vitro studies, some additional CONFLICT OF INTERESTS
critical safety aspects emerged for quercetin. The authors declare that they have no conflict
Quercetin emerged as a mutagenic and po- of interests.
REFERENCES
1. Soerjomataram I, Bray F. Planning for to- 4. Vafadar A, Shabaninejad Z, Movahedpour
morrow: global cancer incidence and the A, et al. Quercetin and cancer: new in-
role of prevention 2020–2070. Nat Rev sights into its therapeutic effects on ovari-
Clin Oncol 2021. Epub ahead of print. an cancer cells. Cell Biosci 2020;10(1):32.
2. Wach A, Pyrzyńska K, Biesaga M. Querce- 5. Treml J, Šmejkal K. Flavonoids as po-
tin content in some food and herbal sam- tent scavengers of hydroxyl radi-
ples. Food Chem 2007;100(2):699–704. cals. Compr Rev Food Sci Food Saf
3. Materska M. Quercetin and its derivatives: 2016;15(4):720–38.
chemical structure and bioactivity- a re- 6. Guo Y, Mah E, Davis CG, et al. Dietary
view. Pol J Food Nutr Sci 58(4):407–13. fat increases quercetin bioavailability in
513You can also read

















































