A Computational Systems Biology Approach Identifies SLUG as a Mediator of Partial Epithelial-Mesenchymal Transition (EMT)
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Research Article
Cells Tissues Organs Received: September 7, 2020
Accepted: October 19, 2020
DOI: 10.1159/000512520 Published online: February 10, 2021
A Computational Systems Biology Approach
Identifies SLUG as a Mediator of Partial
Epithelial-Mesenchymal Transition (EMT)
Ayalur R. Subbalakshmi a Sarthak Sahoo a Kuheli Biswas b Mohit Kumar Jolly a
aCentre
for BioSystems Science and Engineering, Indian Institute of Science, Bangalore, India; bDepartment of
Physical Sciences, Indian Institute of Science Education and Research, Kolkata, India
Keywords behaving as a phenotypic stability factor. Our simulations
Hybrid epithelial/mesenchymal phenotypes · SLUG · Cancer suggest that this specific property of SLUG emerges from the
systems biology · Epithelial-mesenchymal transition · topology of the regulatory network it forms with other key
Mathematical modeling · Phenotypic stability factor regulators of epithelial-mesenchymal plasticity. Clinical data
suggest that SLUG associates with worse patient prognosis
across multiple carcinomas. Together, our results indicate
Abstract that SLUG can stabilize hybrid E/M phenotype(s).
Epithelial-mesenchymal plasticity comprises reversible tran- © 2021 S. Karger AG, Basel
sitions among epithelial, hybrid epithelial/mesenchymal
(E/M) and mesenchymal phenotypes, and underlies various
aspects of aggressive tumor progression such as metastasis, Introduction
therapy resistance, and immune evasion. The process of cells
attaining one or more hybrid E/M phenotypes is termed as Cancer metastasis remains the major cause of cancer-
partial epithelial mesenchymal transition (EMT). Cells in hy- related deaths [Gupta and Massagué, 2006]. Metastasis is
brid E/M phenotype(s) can be more aggressive than those in a dynamic process with extremely high (>99.5%) attrition
either fully epithelial or mesenchymal state. Thus, identify- rates; the minority of cells successful in establishing me-
ing regulators of hybrid E/M phenotypes is essential to deci- tastases are highly plastic, i.e., capable of adapting to their
pher the rheostats of phenotypic plasticity and consequent environmental changes [Celià-Terrassa and Kang, 2016;
accelerators of metastasis. Here, using a computational sys- Jolly et al., 2017b]. An important axis of plasticity in-
tems biology approach, we demonstrate that SLUG (SNAIL2) volved in metastasis is epithelial-mesenchymal plasticity
– an EMT-inducing transcription factor – can inhibit cells (EMP), i.e., bidirectional transitions among epithelial,
from undergoing a complete EMT and thus stabilize them in mesenchymal, and one or more hybrid epithelial/mesen-
hybrid E/M phenotype(s). It expands the parametric range chymal (E/M) phenotypes, that is, a complete or partial
enabling the existence of a hybrid E/M phenotype, thereby epithelial mesenchymal transition (EMT) and its reverse
karger@karger.com © 2021 S. Karger AG, Basel Mohit Kumar Jolly
www.karger.com/cto Centre for BioSystems Science and Engineering
C-wing, 3rd floor, Biological Sciences Building
Indian Institute of Science, CV Raman Road, Bangalore 560012 (India)
mkjolly.15@gmail.com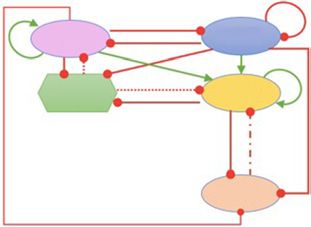
phenomena mesenchymal epithelial transition (MET) a detailed comparison between them, at least in terms of
[Jolly and Celia-Terrassa, 2019]. Recent in vitro, in vivo, their ability to induce a partial EMT, i.e., hybrid E/M
and in silico studies suggest that EMT or MET are not phenotype(s), remains to be done to acquire mechanistic
“all-or-none” processes; instead, cells can attain one or insights into their role in mediating the dynamics of EMP.
more hybrid E/M phenotypes which may be more plastic Here, we investigate the differences between the EMT-
as compared to “fully” epithelial or mesenchymal ones inducing potential of SLUG and SNAIL and investigate
[Font-Clos et al., 2018; Pastushenko et al., 2018; Tripathi the contribution of SLUG in being able to maintain a hy-
et al., 2020]. Consistently, cells acquiring hybrid E/M brid E/M phenotype. Previous mathematical models de-
phenotype(s) are crucial for tumorigenicity and/or me- veloped for EMP signaling have often not distinguished
tastasis-initiating properties across many cancer types between SNAIL and SLUG, and have taken them as a sin-
[Pastushenko et al., 2018; Kröger et al., 2019], and under- gle node [Lu et al., 2013; Tian et al., 2013; Hong et al.,
going a “full” EMT may lead to loss of such “stemness” 2015; Mooney et al., 2016]. With increasing knowledge
traits [Celià-Terrassa et al., 2012; Bierie et al., 2017]. Thus, about the different roles of SNAIL and SLUG in influenc-
hybrid E/M cells are likely to be the “fittest” for metastasis ing EMP in different contexts, including their effect on
[Jolly et al., 2018], a hypothesis which is strengthened by one another [Chen and Gridley, 2013; Gras et al., 2014;
increasing reports suggesting an association of hybrid Ganesan et al., 2015; Nakamura et al., 2018; Sundararajan
E/M phenotypes with worse clinicopathological features et al., 2019; Swain et al., 2019; Hamidi et al., 2020; Varan-
[Godin et al., 2020; Liao and Yang, 2020]. Therefore, it is kar et al., 2020], it becomes imperative to investigate their
critical to understand the molecular players regulating differential roles from a detailed mechanistic perspective.
hybrid E/M phenotypes to decode their effect on aggra- Therefore, we have adopted a mechanism-based compu-
vated metastasis and identify therapeutic approaches to tational systems biology approach to elucidate these dif-
restrict them. ferences. Our results suggest that SLUG is a weaker in-
Investigation of EMT/MET in developmental contexts ducer of EMT as compared to SNAIL; thus, SLUG, but
have helped identify the modulators of cancer cell plastic- not SNAIL, can often stabilize a hybrid E/M phenotype.
ity [Nieto et al., 2016]. One of the key molecules that has Clinical analysis indicates high SLUG expression to be as-
been associated with hybrid E/M phenotypes in various sociated with worse patient survival across carcinomas,
developmental contexts is SLUG (SNAIL2) [Jolly et al., endorsing the growing evidence for a more aggressive be-
2015]; resonating observations have also recently been havior of hybrid E/M phenotype(s).
seen in clinical settings [Steinbichler et al., 2020]. How-
ever, this association has been mostly at a phenomeno-
logical level. SLUG belongs to a family of zinc-finger tran- Materials and Methods
scription factors SNAI. In mammals, this family consists
Software and Datasets
of SNAIL (encoded by the SNAI1 gene; also SNAIL1), All computational analyses were performed using MATLAB
SLUG (encoded by the SNAI2 gene; also SNAIL2), and (version 2018b), and microarray datasets were downloaded using
SMUC (encoded by the SNAI3 gene; also SNAIL3) [Bar- the GEOquery R Bioconductor package [Davis and Meltzer, 2007].
rallo-Gimeno and Nieto, 2005]. While SMUC is much Pre-processing of the microarray datasets was performed for each
sample to obtain the gene-wise expression from probe-wise ex-
less studied, SNAIL and SLUG are observed across spe-
pression matrix using R (version 4.0.0).
cies from Drosophila melanogaster to humans with func-
tions varying from development to oncogenesis [Wu et EMT Score Calculation
al., 2005]. In Drosophila, they are involved in formation The EMT scores for each dataset were calculated using 76 GS,
of mesoderm and development of the central nervous sys- KS, and MLR methods as described earlier [Chakraborty et al.,
2020].
tem [Hemavathy et al., 2000]. In vertebrates, they are in-
76 GS: A weighted sum of 76 gene expression levels was used
volved in the formation of the neural crest [Hu et al., to calculate the score for each sample. The mean across all tumor
2014] and EMT of mesodermal cells, and they can also samples was subtracted to center the scores so that the grand mean
participate in EMT-independent functions such as pre- of the score was zero. Negative scores can be interpreted as M phe-
venting apoptosis from gamma-irradiation in hemato- notype, whereas the positive scores as E phenotype.
MLR: The EMT status by the MLR method is predicted based
poietic cells [Barrallo-Gimeno and Nieto, 2005]. The role on order of the categorical structures and the principle that the
of SLUG and SNAIL – the 2 more well-studied members hybrid E/M state falls in a region intermediary to E and M. The
of the SNAI family – has been found to be partially redun- sample scores range between 0 and 2 where 0 corresponds to pure
dant [Stemmler et al., 2019; Hemavathy et al., 2000]. But E and 2 corresponds to pure M, and a score of 1 indicates a maxi-
2 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/000512520
mally hybrid phenotype. These scores are calculated based on the from the RACIPE output data. The clusters were identified by per-
probability of a given sample being assigned to the E, E/M, and M forming hierarchical clustering on the RACIPE data by setting the
phenotypes. number of clusters to 4, and the number of steady-state solutions
KS: The scores were calculated by comparing cumulative dis- ending up in each of these clusters were quantified. We also per-
tribution functions of epithelial (E) and mesenchymal (M) signa- formed RACIPE on the same circuit but with either overexpressing
tures. If the score is positive, then the sample is said to be M, and SNAIL or SLUG by 10-fold. The same workflow as described above
if it is negative, it is considered to be E. was used to analyze the overexpression-based analysis.
Stemness Score Calculation Kaplan-Meier Analysis
The stemness score was calculated using CNCL as described ProgGene [Goswami and Nakshatri, 2014] was used for con-
[Akbar et al., 2020]. To identify the stem cell-like (CS/M) and non- ducting Kaplan-Meier analysis. The number of samples in SLUG-
stem cell like (NS/E) cells, the samples were clustered based on the high versus SLUG-low categories are given below:
expression of CSC/non-CSC gene list (CNCL). The quantitative − GSE31210 (pathological stage I-II lung adenocarcinomas): n
stemness score was calculated by the correlation of CNCL gene (high) = 113, n (low) = 113
expression values for each sample to the CS/M and NS/E matrices − GSE9891 (285 serous and endometrioid tumors of ovary, peri-
generated based on the median expression levels for each gene in toneum, and fallopian tube): n (high) = 138, n (low) = 138
the CNCL for CS/M and also for NS/E cells in the CCLE database. − GSE30161 (Ovarian carcinoma): n (high) = 29, n (low) = 29
The 2 Pearson’s correlation values generated for each matrix, CS/ − GSE14333 (Primary colorectal cancer sample): n (high) = 94, n
M(r) and NS/E(r), are plotted. (low) = 93
− TCGA-LUAD (Lung cancer sample): n (high) = 75, n (low) =
Mathematical Modeling 75
As per the schematic shown in Figure 2a, the dynamics of all 4 − GSE17536 (Colorectal cancer sample): n (high) = 87, n (low) =
molecular species (miR-200, SNAIL, ZEB, and SLUG) was de- 87
scribed by a system of coupled ordinary differential equations − GSE16125 (Colorectal cancer sample, stage [Duke’s I-IV]): n
(ODEs). The generic chemical rate equation given below describes (high) = 16, n (low) = 16
the level of a protein, mRNA, or micro-RNA (X): dX/dt = − GSE50827 (103 primary pancreatic ductal adenocarcinoma
gXHS(A,A0,n,λ) - kXX samples): n (high) = 16, n (low) = 16
Where the first term gX signifies the basal rate of production, − GSE26712 (Advanced stage ovarian cancer sample): n (high) =
the terms multiplied to gX represent the transcriptional/transla- 93, n (low) = 92
tional/post-translational regulations due to interactions among − GSE18520 (late-stage, high-grade papillary serous ovarian ad-
the species in the system, as defined by the Hill function enocarcinoma sample):
HS(A,A0,n,λ). The rate of degradation of the species (X) is defined − n (high) = 27, n (low) = 26
by the term kXX based on first order kinetics. The complete set of − GSE13876 (157 advanced stage serous ovarian cancer sample):
equations and parameters are presented in the supplementary ma- n (high) = 207, n (low) = 207
terial (for all online suppl. material, see www.karger.com/ − GSE3143 (Breast cancer sample): n (high) = 80, n (low) = 77
doi/10.1159/512520). − GSE2034 (Breast cancer sample): n (high) = 143, n (low) = 143
− GSE19615(Breast cancer sample): n (high) = 58, n (low) = 57
Mean Residence Time Analysis − GSE11121 (Node-negative breast cancer patient sample): n
As the degradation rate of ZEB mRNA and SLUG mRNA is (high) = 103, n (low) = 97
much greater than that of ZEB protein and miR-200, and also the − GSE2990 (189 invasive breast carcinoma sample): n (high) =
production rate of SLUG and SNAIL protein is much larger than 51, n (low) = 50
that of ZEB protein and miR-200, we assumed that ZEB mRNA,
SNAIL protein SLUG mRNA, and SLUG protein reach to the equi-
librium much faster relatively, i.e., (dm_Z)/dt = 0, dS/dt = 0, (dm_ Results
Sl)/dt = 0 and dSl/dt = 0.This assumption reduces the equations
given in the online supplementary material to 2 coupled ODEs of
ZEB and miR-200. Then, the dynamical system was simulated to SNAIL Is a Stronger Inducer of Complete EMT Than
obtain the time evolution of ZEB protein and miR-200 in presence SLUG
of external noise using Euler-Maruyama simulation. Dynamical First, using publicly available transcriptomics datasets
states of the system were coarse grained as an itinerary of basins (GSE80042. GSE40690, GSE43495), we quantified the ex-
visited from the time evolution of ZEB and miR-200. MRT was
calculated by multiplying the total number of successive states with tent of EMT induced by SNAIL and SLUG individually in
it [Biswas et al., 2019]. different cell lines. We used 3 different methods to quan-
tify EMT – 76 GS, MLR, and KS. MLR and KS score the
Random Circuit Perturbation samples on a spectrum of [0, 2] and [-1, 1]; the higher the
We performed random circuit perturbation (RACIPE) analysis score, the more mesenchymal the sample is. On the other
on the extended circuit (Fig. 4a) using the default settings for 10,000
parameter sets. We considered all RACIPE steady-state solutions hand, there is no predefined range of scores calculated by
up to tetra-stable parameter sets for principal component analysis 76 GS, and the higher the 76 GS score, the more epithe-
(PCA) plots. PCA was performed on all the steady-state solutions lial the sample is [Chakraborty et al., 2020].
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 3
DOI: 10.1159/000512520■ D0 *
■ EMT-D5 ns
20 ns ** ** ■ MET-3 ■ D0
*
0.2 ns 0.6
■ MET-5 ■ EMT-D5
0 ■ MET-20 ** ■ MET-3
76GS score
0
MLR score
KS score
■ D0 0.4 ■ MET-5
–20 ■ EMT-D5 ■ MET-20 **
–0.2 **
■ MET-3
**
0.2
–40 ■ MET-5 ns
■ MET-20 ** –0.4 ns ns
**
0
a GFP SNAI1 SNAI2 GFP SNAI1 SNAI2 GFP SNAI1 SNAI2
NS/E
40 * ns 0.6 * ns CS/M 1 Mock
2
DDR2
20 * 0.4
FN1
0.5
SNAI1
76GS score
VIM
1 SNAI2
DKK3
KS score
SLIT2
0 BNC2
SNAI3
CS/M
0.2 0 TMEM158
CLDN4 0
–20 –1
ST14
AP1M2
0 –0.5
CDH1
–40 –2 IRF6
ZNF165
–0.2 ** BSPRY
–1
O _C
O _C
O A
A _B
AI _B
SN IL2 A
SN IL2 B
AI _A
SN IL1 C
SN IL1 B
AI _A
_C
A _
A _
A _
M CK_
A _
SN CK
SN IL3
M IL3
M CK
SN L2
L1
SN L3
–1 –0.5 0 0.5 1
A
CK
1
2
3
CK
1
2
3
SN
b
AI
AI
AI
AI
AI
AI
O
O
NS/E
SN
SN
SN
SN
SN
SN
M
M
*
* NS/E 1
20 0.4 * CS/M Untreated
76GS score
2 Vector
SLIT2
KS score
*
ZNF165
IRF6 0.5 Twist
0 1 CDH1
SNAI1
CS/M
0.2
ST14
0 AP1M2
BNC2 0 SNAI2
–1
DDR2
CLDN4
–20 –2
DKK3
–0.5
** 0
TMEM158
VIM
FN1
–1
d
ct _a
ct b
tre to c
Un a r_a
at _b
Tw d_c
Tw st_c
Sl st_b
Sn _b
Sn il_b
Sn il_c
Sl l_a
Sl g_c
Tw _b
_a
Un ec or_
Ve or_
ed
or
SN t
ed
1
2
or
ist
1
2
–1 –0.5 0 0.5 1
Ve ed
ist
tre ted
ug
ug
ai
a
is
e
u
a
i
i
AI
AI
AI
AI
at
ct
ct
tre
Tw
Tw
V
at
at
SN
SN
SN
Un
Ve
Ve
c
tre
tre
NS/E
Un
Un
Fig. 1. SNAIL elicits a stronger EMT than SLUG. a EMT scores for GSE80042 using 76 GS, KS, and MLR meth-
ods b EMT scores for GSE40690 using 76 GS and KS methods. c Same as b but for GSE43495. d Left, top and
bottom: Clustering of samples using CNCL gene list in GSE40690 and GSE43495, respectively. Right, top and
bottom: Stemness score based analysis for GSE40690 and GSE43495 respectively. * Indicates a p value < 0.05 and
** indicates p 0.05).
In LNCaP prostate cancer cells with inducible and re- calculated across these methods correlated well with one
versible expression of SNAI1 or SNAI2 (GSE80042), another – while KS and MLR scores were positively cor-
those induced with SNAI1 showed a stronger extent of related, 76 GS scores correlated negatively with KS and
EMT as compared to those induced with SNAI2. This with MLR scores, as expected (online suppl. Fig. 1a–c).
trend was consistently seen across the 3 scoring methods Next, for GSE43495 and GSE40690 datasets, we also cal-
(Fig. 1a). Similarly, in immortalized human mammary culated the stemness scores using a gene list corresponding
epithelial cells transfected with retroviral constructs con- to EMT and stemness for breast cancer cells (CNCL) [Ak-
taining SNAI1, SNAI2 and SNAI3 (HMEC-hTERT), bar et al., 2020]. Cells overexpressing SNAI1 were found to
those induced with SNAI1 showed a significantly stron- show a stronger enrichment of this gene signature as com-
ger EMT induction as compared to those induced with pared to those overexpressing SNAI2 and/or TWIST
SNAI2 or SNAI3 (GSE40690); the degree of induction (Fig. 1d; left, top, bottom). When projected on the 2-di-
was the weakest for SNAI3. Again, this trend was consis- mensional representation of NS/E (non-cancer stem-like/
tent across scoring methods (Fig. 1b). Consistently, in hu- epithelial) and CS/M (cancer stem cell-like/mesenchymal)
man mammary epithelial cells, transduction of SNAI1 in- axes, the cells induced with SNAI1 were found to be more
duced a stronger EMT than that of SNAI2 or TWIST of CS/MHi-NS/ELow as compared to those induced with
(Fig. 1c) (GSE43495). In all these scenarios, the scores SNAI3 (CS/MLow-NS/EHi) (Fig. 1d; right, top); the SNAI2
4 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/0005125200.8
{E,H} {E,H,M}
I (Ext) 1.4
0.7
{H,M}
μ0200 mSI, × 104
1.2
SLUG SNAIL 0.6 {E}
λSIμ200
1 {H}
0.5 {M} {E,H}
{E}
0.4 0.8
µ200 ZEB {H,M} {M}
{H}
a 0.3 0.6
0 100 200 300 50 60 70 80 90
c I ext (103 molecules) I ext (103 molecules)
1,000
Core network
0.7 1
500
ZEB mRNA, molecules
{M} 0.8
0.6
0 {M}
{H}
λSmSI
1,000 {E} {H}
λSIS
SLUG network 0.6
{H,M} {E,H}
0.5
{E} {H,M}
500 {E,H}
0.4
0.4
0 0 50 100 150 40 50 60 70 80 90
0 50 100 d I ext (103 molecules) I ext (103 molecules)
b I ext (103 molecules)
Fig. 2 SLUG inhibits the progression to a complete EMT. a Sche- diagrams for the SLUG network driven by an external signal (l) for
matic representation of the EMT network coupled with SLUG. Red varying strength of interactions along the SLUG-miR200 axis: Left:
bars denote inhibition, green arrows indicate activation. Solid ar- Phase diagram for varying strength of repression of miR200 by
rows represent transcriptional regulation, dashed-line represents SLUG. Right: Phase diagram for varying threshold level of miR200
indirect regulation and dotted line represent micro-RNA mediated needed for SLUG repression (i.e., reciprocal link: miR-200 inhibit-
regulation. The circuit shown within a dotted rectangle is the con- ing SLUG). d Phase diagram for SLUG network driven by an exter-
trol case/core network (i.e., network without SLUG). b Bifurcation nal signal (l) for varying strength of interactions along the SLUG-
diagrams indicating the ZEB mRNA levels for increasing external SNAIL axis: Left: Phase diagram for varying strength of repression
signal (l) levels for the core network (top panel) and the network of SNAIL by SLUG. Right: Phase diagram for varying strength of
including SLUG, that is, SLUG network (bottom panel). c Phase the reciprocal link (i.e., repression of SLUG by SNAIL).
and/or TWIST-induced ones had a weaker enrichment of line suppl. Table 2). These interconnections were repre-
CS/MHi-NS/ELow signatures (Fig. 1d; right, bottom). Put sented by a set of coupled ODEs (online suppl. Section 1
together, these results suggest that SNAIL is a stronger in- and Table 1).
ducer of EMT than SLUG, and indicates the possibility of We started with first calculating the bifurcation dia-
SLUG being associated with a partial EMT phenotype in- gram of ZEB mRNA levels for varying levels of an exter-
stead of a completely mesenchymal one. nal EMT-inducing signal (Fig. 2b). As the value of EMT-
inducing signal increases, the cells can switch from an
SLUG Can Act as a “Phenotypic Stability Factor” for a epithelial state (ZEB mRNA < 100 molecules) to a hybrid
Hybrid E/M Phenotype E/M phenotype (100 < ZEB mRNA molecules 600 mole-
SLUG on EMT, we expanded the previous EMP circuitry cules); this trend is seen for the circuit that does not in-
we simulated earlier [Lu et al., 2013] by incorporating the clude SLUG (Fig. 2b; top) as well as the one that includes
interconnections of SLUG with miR-200 and SNAIL SLUG (Fig. 2b; bottom). However, in presence of SLUG,
(Fig. 2a). SLUG and SNAIL can inhibit each other, al- an interesting feature was seen which was not present in
beit indirectly; also, SLUG and miR-200 have been the control case – a parameter region in which the only
known to form a mutually inhibitory feedback loop (on- phenotype available to cells was the hybrid E/M one
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 5
DOI: 10.1159/000512520(Fig. 2b; bottom, gray region); thus, SLUG enabled the to repress SLUG. As this threshold value is increased (i.e.,
existence of a monostable hybrid E/M phenotype. Also, as the inhibition of SLUG by miR-200 is weakened), a larg-
in the presence of SLUG, a higher level of EMT-inducing er {H} region is observed at the expense of shrinking {E}
external signal I is required to push the cells to a com- and {M} regions (Fig. 2c; right). Put together, these results
pletely mesenchymal state, representing the effect of suggests that as the influence of SLUG is reduced (i.e.,
SLUG in preventing cells from undergoing a full EMT. weaker repression of miR-200 by SLUG, or stronger inhi-
To ascertain the robustness of this prediction, we per- bition of SLUG by miR-200), and the existence of a mo-
formed sensitivity analysis where each kinetic parameter nostable hybrid state ({H}) is disfavored. Conversely,
of the model was varied – one at a time – by ± 10%, and when the effective impact of SLUG in modulating the
the corresponding change in the range of values of the EMP dynamics is increased (i.e., a stronger repression of
external EMT-driving signal (I) enabling a hybrid E/M miR-200 by SLUG, or weaker inhibition of SLUG by miR-
phenotype (i.e., the interval of x-axis between double-sid- 200), the existence of a monostable {H} phase is facilitated.
ed arrows shown at the bottom) was measured (online Next, we quantified the effect of interactions between
suppl. Fig. 2a). Except for a few parameter cases, the SLUG and SNAIL in altering the EMP dynamics (Fig. 2d).
change in this interval of values of I was found to be less As the strength of repression of SNAIL by SLUG is de-
than 10% for a corresponding 10% change in the param- creased, the {E} phase shrinks and the {M} phase expands,
eter values, indicating the robustness of the system. A i.e., when SNAIL is inhibited weakly by SLUG, a milder
change in few selected parameters such as those control- activation of SNAIL by the external signal I is sufficient
ling ZEB self-activation or the effect of SLUG inhibiting to induce an EMT (Fig. 2d, left). However, no changes
SNAIL exhibited stronger sensitivity; but even in these were observed when the threshold value of SLUG needed
cases, the decrease in the range of values of I for which a to repress SNAIL was altered (online suppl. Fig. 2c). To
hybrid E/M existed was smaller as compared to the case determine the effects of the reciprocal link (i.e., transcrip-
in the absence of SLUG (dotted line in online suppl. tional inhibition of SLUG by SNAIL), we varied the
Fig. 2a). Thus, SLUG can be thought of as a potential strength of repression of the corresponding link, but did
“phenotypic stability factor” for hybrid E/M phenotype. not observe any appreciable change in the phase bound-
We next sought to examine how this trait of enabling aries (Fig 2d, right; online suppl. Fig. 2d).
hybrid E/M phenotype changed when the strengths of in- SLUG is also known to self-activate in certain contexts
teractions between SLUG-miR200 and SLUG-SNAIL [Kumar et al., 2015; Zhou et al., 2019]. Thus, we examined
were varied. While in the presence of SLUG, the hybrid the changes in EMP dynamics upon including SLUG self-
state could exist in a monostable regime ({H}), it also co- activation. We observed that irrespective of the link
existed with epithelial and mesenchymal states, thus giv- whose strength was being varied, including self-activa-
ing rise to the multistable phases (i.e., combinations of tion of SLUG increases the parameter region for the exis-
coexisting phenotypes) {E,H}, {H,M} for varying strengths tence of monostable {H} region. For instance, the {H}
of external signal. On the other hand, neither the {E, H, phase was seen to persist even when SLUG repressed
M} phase nor the {E, M} one was seen in the circuit con- miR-200 weakly, and consequently, the emergence of the
taining SLUG, as opposed to the control case (compare tri-stable {E,H,M} phase was not noticed in presence of
the top vs. bottom in Fig. 2b). First, we analyzed the effect SLUG self-activation (compare online suppl. Fig 2e with
of interaction between SLUG and miR200 on the dynam- Fig. 2c, left). Similarly, transition into a completely mes-
ics (Fig. 2c). When the strength of repression of miR-200 enchymal state ({M}) required larger values of external
by SLUG is reduced, 2 key changes are seen: (a) the {M} signal I activating SNAIL (compare oline suppl. Fig 2f
region expands while the {E} region shrinks, i.e., it is pos- with Fig. 2d, left), leading to an expanded monostable hy-
sible for cells to exit an epithelial phenotype and/or un- brid ({H}) region. Therefore, SLUG self-activation may
dergo a complete EMT at lower values of external signal I, further enhance the likelihood of cells stably acquiring a
and (b) the monostable hybrid region ({H}) disappears, partial EMT state and prevent them from transitioning to
and the {E, H, M} phase emerges (Fig. 2c; left). Similar a fully mesenchymal state.
trends, albeit to a weaker extent, were also seen when the Finally, we replotted the abovementioned phase dia-
threshold level of SLUG needed to repress miR200 is al- grams for a few varied values of other parameters pertain-
tered (online suppl. Fig. 2b). Further, to determine the ef- ing to relative strengths of interaction of SLUG with miR-
fect of the reciprocal link (i.e., repression of SLUG by 200 and/or SNAIL. Altering the degree of inhibition of
miR200), we varied the threshold levels of miR200 needed SLUG by SNAIL did not significantly change the phase
6 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/0005125201,000
25
{M} state {M} 1,000 {M}
{E,M}
800
ZEB mRNA, molecules
ZEB mRNA, molecules
20 {E} state I = 50 × 103
800
{H} state
{E,H,M} 600 I = 55 × 103
15
{H,M}
MRT, h
600
400 {H}
10
400
{H}
5 200 I = 70 × 103
{E} 200
0 0 I = 78 × 103
50 60 70 0 200 400 600 800 1,000 0 200 400 600 800 1,000
a I ext (103 molecules) b Time, h Time, h
5 I = 53 × 103 700
{M} state 300
4 {E} state I = 51 × 103 600
250
ZEB mRNA, molecules
ZEB mRNA, molecules
{M}
{H} state
3 200 {H} 500
MRT, h
{H,M}
{H,E} {H} 150 400
2
100 300 {H}
1 I = 78 × 103
50 200
{E} I = 80 × 103
0 100
0
50 60 70 80 0 200 400 600 800 1,000 0 200 400 600 800 1,000
c I ext (103 molecules) d Time, h Time, h
Fig. 3. Mean residence time analysis for core network (i.e. absence lines indicate average ZEB mRNA values for a given state and ex-
of SLUG) versus the SLUG network. a Variation of mean residence ternal signal strength. c Same as a but for the SLUG network. The
time (MRT) with varying external signal (l) levels for the core net- blank profile for {H} indicates the parameter space in which the
work. b Stochastic simulation of core circuit (left) showing switch- only phenotype available is hybrid E/M (h) one. d Stochastic sim-
ing between E, M states at I = 50,000 molecules, switching among ulation for the SLUG network showing switch between E, H states
E, H, M states at I = 55,000 molecules, and switching between H, at I = 51,000 and I = 53,000 molecules, and switching between H,
M states at I = 70,000 and I = 78,000 molecules. The solid straight M states at I = 78,000 and I = 80,000 molecules.
diagram (online suppl. Fig. 3a, b) as compared to the con- ble hybrid E/M region ({H}) almost doubled, while the
trol case (Fig. 2d, left). However, increasing or decreasing latter case led to a 4-fold decrease in the same (compare
the strength of repression of SNAIL by SLUG modified online suppl. Fig 3e, f with Fig. 2d, right).
the phase diagram. When SLUG represses SNAIL strong- Overall, these simulations suggest that the stronger the
ly, a larger value of the external signal I (that activates influence of SLUG in inhibiting SNAIL and/or miR-200
SNAIL) is required to induce EMT (compare online sup- levels, the higher the likelihood of cells maintaining a hy-
pl. Fig 3c with Fig. 2d, right). In contrast, when the repres- brid E/M phenotype.
sion of SNAIL by SLUG was weakened, the monostable
epithelial and hybrid regions shrink, and a lower value of SLUG Can Alter the Mean Residence Time of Cells in
external signal (I) was sufficient to induce EMT (compare a Hybrid E/M Phenotype
online suppl. Fig 3d with Fig. 2d, right). Because relative- Previously identified “phenotypic stability factors” –
ly major changes in the phase diagram were observed GRHL2, OVOL1/2, and ΔNP63α – had another dynamical
when the inhibitory action of SLUG was altered rather trait: their presence increased the mean residence time
than the repression on SLUG being altered, we plotted (MRT) of cells in a hybrid E/M phenotype. MRT is the
these phase diagrams for 2 cases: (a) when SLUG inhibits mean (or average) time spent by the cells in a particular ba-
both miR-200 and SNAIL strongly, and (b) when SLUG sin of attraction – E, M, and hybrid E/M here – as calcu-
inhibits both miR-200 and SNAIL weakly. In the former lated via stochastic simulations [Biswas et al., 2019]. Thus,
case, the parameter region corresponding to a monosta- the larger the MRT of a phenotype, the higher the relative
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 7
DOI: 10.1159/000512520Cdh1 Control Slug Control
SLUG SNAIL
PC-2 (22.1% var.)
PC-2 (22.1% var.)
µ200 ZEB
E-CAD c PC-1 (65.9% var.) d PC-1 (65.9% var.)
a
Cdh1 Slug OE Snail OE
Cdh1
PC-2 (22.3% var.)
PC-2 (22.9% var.)
Control
PC-2 (22.1% var.)
H1 e PC-1 (65.1% var.) f PC-1 (62.8% var.)
Low High
M E
0.5 *
* ■ Control
ns ■ Snail OE
H2 Fraction of cases 0.4 ns ■ Slug OE
0.3 *
PC-1 (65.9% var.)
0.2 *
Cdh1 mir200 Slug Snail Zeb1 0.1
PC-1 0.47 0.49 –0.49 –0.47 –0.29
b PC-2 0.40 –0.32 0.22 0.32 –0.77 0
g E M H1
Fig. 4. RACIPE analysis reveals the existence of a SLUG+CDH1+ sion levels of SLUG. e PCA plot from RACIPE analysis in the case
hybrid E/M phenotype. a Schematic of the extended gene regula- of SLUG overexpression (colored by the gene expression levels of
tory network showing the interactions of SNAIL and SLUG with CDH1). f PCA plot from RACIPE analysis in the case of SNAIL
ZEB1, miR200 and CDH1. The green links represent activating over-expression (colored by the gene expression levels of CDH1).
links, and the red links represent inhibition links. Dotted line For e, f, the PC1 obtained is qualitatively similar to PC1 obtained
shows the microRNA-mediated regulation. Dashed-dotted line for the control (panels b–d) case; also, PC1 and PC2 explain simi-
shows indirect inhibition of ZEB by E-cadherin through β-catenin. lar variance, indicating no major changes in structure of the steady
b Principal Component Analysis (PCA) plot showing the different state solution set. g Quantification of the frequency of steady state
clusters that emerge from the RACIPE analysis, also identified via solutions for the clusters corresponding to E, M, H1 showing that
hierarchical clustering for the RACIPE output. E and M represent H1 phenotype is depleted by SNAIL overexpression while it is en-
the epithelial and the mesenchymal phenotypes respectively. H1 riched for by SLUG overexpression (* indicates a p value < 0.01 by
and H2 are 2 hybrid phenotypes. Contributions of the various using the Student t test with unequal variances; ns, denotes non-
genes to the 2 principal component axes PC1 and PC2 are listed (statistically) significant change). Error bars represent the mean ±
(PC1 represents the canonical EMT axis). c PCA plot colored by standard deviation for results obtained from n = 3 independent
the expression levels of CDH1. d PCA plot colored by the expres- RACIPE replicates.
stability of the same. Hence, beyond enabling the parameter For the control case (i.e., circuit without SLUG), the
range of values of an external EMT-inducing signal for the epithelial state was found to be more stable in the {E,M}
existence of a hybrid E/M phenotype (as shown via bifurca- phase and the mesenchymal state is more stable in the
tion diagrams), an increase in MRT is usually thought of as {H,M} phase. All 3 states were found to be almost equally
a hallmark trait of a PSF. We investigated whether SLUG stable in the {E,H,M} phase (Fig. 3a). Cells were seen to
increases the MRT of cells in a hybrid E/M phenotype. be capable of transitioning among these different states
8 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/000512520High expression High expression High expression High expression
Low expression Low expression Low expression Low expression
1 1 1 1
HR 1.47(1.11–1.93)
HR 1.39(1.08–1.78) HR 1.18(1.04–1.35)
PVAL: 0.0065579
0.8 PVAL: 0.00952 0.8 PVAL: 0.0132961 0.8 0.8
Relapse free survival
Relapse free survival
Relapse free survival
Overall survival
0.6 0.6 5 years 0.6 5 years 0.6
0.4 0.4 0.4 0.4
3 years 5 years
3 years
0.2 5 years 0.2 0.2 0.2
3 years HR 2.32(1.55–3.48)
3 years PVAL: 4.33*10–5
0 0 0 0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
50
00
50
00
50
00
50
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
1,
1,
2,
2,
3,
3,
1,
2,
3,
4,
5,
6,
1,
2,
3,
4,
1,
2,
3,
4,
5,
a Days b Days c Days d Days
High expression High expression High expression High expression
Low expression Low expression Low expression Low expression
1 1 1 1
HR 1.45(1.08–1.93) HR 1.5(1.08–2.09) HR 1.68(1.2–2.36)
HR 3.3(1.52–7.17) PVAL: 0.0127057
0.8 0.8 0.8 PVAL: 0.0164884 0.8 PVAL: 0.0024593
PVAL: 0.0025606
Overall survival
Overall survival
Overall survival
Overall survival
0.6 0.6 0.6 0.6
0.4 0.4 0.4 0.4
3 years 5 years
3 years 5 years
0.2 5 years 0.2 0.2 0.2
3 years
3 years
0 0 0 0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
0
50
00
50
00
50
00
00
00
00
00
00
00
50
00
50
1,
1,
2,
2,
1,
2,
3,
4,
1,
2,
3,
1,
1,
e Days f Days g Days h Days
Fig. 5. SLUG correlates with poor patient survival. a–d Relapse-free survival of GSE31210 (lung cancer samples),
GSE9891 (ovarian cancer samples), GSE30161 (ovarian cancer sample), and GSE14333 (colorectal cancer sam-
ple), respectively. e–h Overall survival of TCGA-LUAD (lung cancer sample), GSE17536 (colorectal cancer sam-
ple), GSE16125 (colorectal cancer sample), and GSE50827 (pancreatic cancer sample). HR, denotes hazard ratio;
PVAL, denotes p value.
under the influence of noise, depending on the level of the At a transcriptional level, ZEB1, SNAIL, and SLUG can
EMT-inducing external signal (I) (Fig. 3b). In the pres- all repress E-cadherin [Batlle et al., 2000; Mooney et al.,
ence of SLUG, the hybrid state was found to be more sta- 2016; Sterneck et al., 2020], and SNAIL and SLUG can
ble than the epithelial state in the {E,H} phase and also in activate ZEB1 [Wels et al., 2011; Wu et al., 2017]. Further,
the {H,M} phase (Fig. 3c), and cells switched back and SLUG is known to self-activate [Kumar et al., 2015], but
forth from hybrid E/M state to epithelial or mesenchymal SNAIL self-inhibits [Peiró et al., 2006]. E-cadherin can
state depending on the value of the external signal (I) also indirectly repress ZEB1 by regulating the localization
(Fig. 3d). Thus, the presence of SLUG was capable of en- of β-catenin [Mooney et al., 2016].
abling a higher MRT for the hybrid E/M state. We used a computational framework RACIPE that en-
However, in the {H, M} phase, as the levels of I in- ables mapping the general dynamic properties of gene reg-
crease, the MRT of mesenchymal state can be larger than ulatory networks by solving a set of coupled ODEs to ob-
that of a hybrid E/M state, as the system approaches a tain the possible steady-state solutions for a given network
monostable mesenchymal state ({M}). These trends are and corresponding ensemble of parameter sets. All the pa-
also seen in potential landscape diagrams for correspond- rameters and initial conditions required for solving the set
ing cases (online suppl. Fig. 4). of ODEs are randomly sampled from biologically relevant
ranges [Huang et al., 2018]. We performed RACIPE anal-
The Role of SLUG in Stabilizing Hybrid E/M State Is a ysis on this gene regulatory network (Fig. 4a) to find the
Function of the Network Topology robust steady-state solutions emerging from it, for an en-
Next, we deciphered the generic emergent properties semble of parameter sets. We performed PCA on all the
of an extended gene regulatory network involving SNAIL, steady-state solutions produced by RACIPE (Fig. 4b). The
SLUG, ZEB1, miR-200, and E-cadherin (CDH1) (Fig. 4a). PCA plot reveals the existence of 4 distinct clusters: epi-
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 9
DOI: 10.1159/000512520thelial (E), mesenchymal(M) and 2 hybrid clusters (H1 relapse-free survival (online suppl. Fig. 5g, h). These con-
and H2), as confirmed by hierarchical clustering on all the text-specific differences of the association of SLUG with
steady-state solutions obtained via RACIPE (Fig. 4b). PC1 breast cancer prognosis traits remains to be better under-
explains more than 65% of the variance in the data, and stood.
can be considered as the “canonical” EMT axis, given its
composition that places CDH1 and miR-200 (epithelial
markers) in one group and ZEB1, SLUG, and SNAIL Discussion
(mesenchymal markers) in the other opposing one. Thus,
E and M clusters are well-segregated along the PC1 axis, Cancer cell plasticity and metastasis are complex and
and 2 clusters potentially representing hybrid E/M states emergent phenomena whose molecular underpinnings
are sandwiched between the E and M ends. These 2 clus- are yet to be comprehensively identified. An important
ters (H1, H2) seem to be segregated along PC2 axis. Bio- axis of cellular plasticity is EMP. Various families of tran-
logically, the hybrid cluster H1 can denote a SLUG + scription factors have been known to affect EMP –
CDH1+ cell phenotype which has been documented re- TWIST1/2, ZEB1/2, and SNAIL1/2 can induce EMT [La-
cently experimentally as well (Fig. 4c, d) [Sterneck et al., mouille et al., 2014], while OVOL1/2 and GRHL1/2/3 can
2020]. induce MET [Saxena et al., 2020; Sundararajan et al.,
Next, we compared the effect of overexpression of 2020]. Recent investigations have highlighted different
SLUG versus SNAIL. Interestingly, these both scenarios and possibly even a contradictory role of different mem-
showed opposite results: while SLUG overexpression en- bers of the same transcription factor family, for instance,
riched the SLUG+ CDH1+ phenotype, SNAIL overexpres- the opposing roles of ZEB1 and ZEB2 in melanoma [Car-
sion depleted it (Fig. 4e–g), suggesting that SNAIL and amel et al., 2013]. Here, we examined the differences in
SLUG may play different or even opposing roles in driv- the roles of SNAIL1 (encoded by the SNAI1 gene; also
ing EMP, owing to the differences in underlying network called SNAIL) and SNAIL2 (encoded by the SNAI2 gene;
topology. Furthermore, this enrichment of the SLUG+ also called SLUG) in inducing EMT.
CDH1+ population in the case of SLUG overexpression Our results suggest that SLUG, but not necessarily
happens at the expense of the M cell population (Fig. 4g), SNAIL, associates with a partial EMT or hybrid E/M phe-
lending credence to our previous results that SLUG is as- notype. These observations are reminiscent of experi-
sociated specifically with a partial EMT instead of a full ments suggesting that SNAIL is a stronger inducer of mo-
EMT phenotype. Besides, this analysis endorses that net- lecular and/or morphological features associated with
work topology can be used to pinpoint potential PSFs for EMT, as compared to SLUG and SNAIL3 [Gras et al.,
the hybrid E/M phenotypes [Jolly et al., 2016]. 2014]. Similar to other EMT-TFs, SLUG initiates EMT by
repressing CDH1 by binding to the E-boxes [Hajra et al.,
High SLUG Expression Correlates with Poor Prognosis 2002; Slabáková et al., 2011]; however, its binding affinity
Hybrid E/M phenotype is thought of as more aggressive to the corresponding E-box DNA element was found to
[Jolly et al., 2018; Pastushenko and Blanpain, 2019]. This be two orders of magnitude less than that of SNAIL [Bolós
hypothesis is further corroborated by the data showing et al., 2003]. Intriguingly, in TGFβ1-induced EMT in
poor patient survival for higher expression of PSFs such as BPH-1 cells, SLUG expression was found to be decreased
GRHL2, OVOL1/2, and NRF2 [Jolly et al., 2016; Bocci et al., at later time points (72–96 h), but SNAIL and ZEB1 levels
2019]. To ascertain the effects of SLUG on patient progno- were consistently high [Slabáková et al., 2011], indicating
sis, we investigated its role in patient outcomes. We ob- a possible role of SLUG initiating rather than maintaining
served that high SLUG expression associated consistently EMT [Savagner et al., 1997]. Consistently, SLUG levels
with poor relapse-free survival in lung, ovarian, and colorec- were observed to peak in intermediate states of EMT in
tal cancer (Fig. 5a–d), and with worse overall survival in human mammary epithelial cells treated with TGFβ [van
lung, ovarian, colorectal, and pancreatic cancer (Fig. 5e–h; Dijk et al., 2018], endorsing the specific association of
online suppl. Fig 5a-c), indicating that high SLUG levels can SLUG with a partially differentiated phenotype as wit-
correlate with poor prognosis in many carcinomas. nessed in multiple scenarios involving EMT – embryonic
In the case of breast cancer, the results were equivocal. development, wound healing, and carcinoma progres-
High SLUG expression correlated with better overall pa- sion [Savagner et al., 2005; Côme et al., 2006; Leroy and
tient survival (online suppl. Fig. 5d) and better metasta- Mostov, 2007; Hudson et al., 2009]. SLUG is also known
sis-free survival (online suppl. Fig. 5e, f), but with poor to activate P-cadherin [Idoux-Gillet et al., 2018], another
10 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/000512520proposed marker for partial EMT [Ribeiro and Paredes, miR-151, CBP, RCP, HDAC1, HDAC3, BRD4, and STAT3
2015]. [Daugaard et al., 2017; Hwang et al., 2017; Chesnelong et
The association of SLUG with partial EMT phenotype(s) al., 2019; Cheng et al., 2020; Dai et al., 2020; Hu et al., 2020;
also helps contextualize the growing literature of hybrid Jury et al., 2020; Kuang et al., 2020; Shafran et al., 2020;
E/M phenotype being more stem-like as compared to a Wang et al., 2020]. Moreover, SLUG may be involved in
complete EMT [Jolly et al., 2019], and the functional role feedback loops with one or more p63 isoforms [Herfs et al.,
of SLUG in enabling stemness [Sterneck et al., 2020]. Mam- 2010; Dang et al., 2016; Srivastava et al., 2018] that can also
mary stem cells exhibit high SLUG levels, and SLUG-ex- enable a partial EMT [Jolly et al., 2017a; Westcott et al.,
pressing normal mammary epithelial cells were seen to ex- 2020]. Thus, while the proposed role of SLUG in stabilizing
hibit a partial EMT phenotype and generated 17 times hybrid E/M phenotype(s) is largely robust to parametric
more organoids than the control case [Guo et al., 2012; Ye variations, including other links and/or nodes, it can alter
et al., 2015]. Upon knockdown of SLUG, mammary stem the landscape of EMP dynamics. A better understanding
cells are unable to transition to basal progenitor cells, lead- of similarities and differences in terms of networks acti-
ing to abnormal mammary architecture [Phillips et al., vated by SNAIL versus SLUG to influence the different
2014]. The mammosphere-forming potential of basal pro- states of EMT and associated traits such as drug resistance
genitor cells also decreased upon SLUG knockdown, thus [Kurrey et al., 2009; Li et al., 2020], immune evasion [Tripa-
the primary mammospheres generated from SLUG-KO thi et al., 2016], anoikis resistance [Huang et al., 2013], and
cells were unable to give rise to secondary mammospheres, autophagy [Xu et al., 2019] across different cancer types is
highlighting the role of SLUG on self-renewal processes in therefore required.
mammary gland morphogenesis [Nassour et al., 2012].
Consistent observations have been made in breast cancer
and prostate cancer, where SLUG degradation promotes Acknowledgement
the differentiation of breast cancer stem cells [Fraile et al.,
2020], and the basal prostate cells that expressed SLUG ex- This article has been uploaded on bioRxiv as a preprint: https://
hibited a partial EMT phenotype and displayed increased www.biorxiv.org/content/10.1101/2020.09.03.278085v1.
stemness ability [Kahonouva et al., 2020]. The functional
contribution of SLUG in vitro and/or in vivo enabling
stemness is also observed in human epidermal progenitor Statement of Ethics
cells [Mistry et al., 2014] as well as other cancers such as Not required for this type of study, as it does not involve any
glioblastoma [Chesnelong et al., 2019], hepatocellular car- human subjects/animal models/identifiable human material and
cinoma [Sun et al., 2014; Tang et al., 2016], colorectal can- data. The study is purely computational in nature and any clinical
cer [Kato et al., 2020], lung cancer [Kim et al., 2020], and data analyzed is publicly available in NCBI GEO.
squamous cell carcinomas [Yu et al., 2016; Moon et al.,
2020]. Whether higher stemness is connected to a hybrid
E/M phenotype in these cancers remains to be identified. Conflict of Interest
Our results indicate that the network topology that
The authors declare no conflicts of interest.
SLUG forms with SNAIL and miR-200 enables its role as a
“phenotypic stability factor” for a hybrid E/M phenotype.
It should be noted that the strengths of interactions of
Funding
SLUG with SNAIL and/or miR-200 can be context depen-
dent, thus altering its ability to be able to stabilize hybrid This work was supported by a Ramanujan Fellowship (SB/S2/
E/M phenotype(s). miR-200 family members have been RJN-049/2018) awarded to M.K.J. by the Science and Engineering
shown to inhibit SLUG in esophageal cancer and prostate Research Board (SERB), Department of Science and Technology
cancer [Liu et al., 2013; Zong et al., 2019; Basu et al., 2020], (DST), Government of India.
while SLUG can inhibit miR-200 in prostate cancer [Liu et
al., 2013]. On the other hand, SLUG and SNAIL have been
Author Contributions
reported to inhibit each other in oral cancer [Nakamura et
al., 2018] and breast cancer [Ganesan et al., 2015]. The ef- M.K.J. designed and supervised research; A.R.S., S.S., and K.B.
fect of SLUG on EMP dynamics can also be influenced by performed research and analyzed data. All authors contributed to
upstream regulators of SLUG such as RNF8, YAP, C/EBPδ, writing and editing of the manuscript.
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 11
DOI: 10.1159/000512520References
Akbar MW, Isbilen M, Belder N, Canli SD, Cheng D, Jin L, Chen Y, Xi X, Guo Y. YAP pro- Gupta GP, Massagué J. Cancer metastasis: build-
Kucukkaraduman B, Turk C, et al. A stemness motes epithelial mesenchymal transition by ing a framework. Cell. 2006;127(4):679–95.
and EMT based gene expression signature upregulating Slug expression in human Hajra KM, Chen DY, Fearon ER. The SLUG zinc-
identifies phenotypic plasticity and is a pre- colorectal cancer cells. Int J Clin Exp Pathol. finger protein represses E-cadherin in breast
dictive but not prognostic biomarker for 2020;13(4):701–10. cancer. Cancer Res. 2002;62(6):1613–8.
breast cancer. J Cancer. 2020;11(4):949–61. Chesnelong C, Hao X, Cseh O, Wang AY, Luch- Hamidi S, Nakaya Y, Nagai H, Alev C, Kasukawa
Barrallo-Gimeno A, Nieto MA. The Snail genes as man HA, Weiss S. SLUG directs the precursor T, Chhabra S, et al. Mesenchymal-epithelial
inducers of cell movement and survival: Im- state of human brain tumor stem cells. Can- transition regulates initiation of pluripotency
plications in development and cancer. Devel- cers. 2019;11(11):1635. exit before gastrulation. Development. 2020;
opment. 2005;132(14):3151–61. Côme C, Magnino F, Bibeau F, De Santa Barbara 147(3):dev184960.
Basu S, Chaudhary A, Chowdhury P, Karmakar P, Becker KF, Theillet C, et al. Snail and slug Hemavathy K, Ashraf SI, Ip YT. Snail/Slug family
D, Basu K, Karmakar D, et al. Evaluating the play distinct roles during breast carcinoma of repressors: Slowly going into the fast lane
role of hsa-miR-200c in reversing the epithe- progression. Clin Cancer Res. 2006; 12(18): of development and cancer. Gene. 2000;
lial to mesenchymal transition in prostate 5395–402. 257(1):1–12.
cancer. Gene. 2020;730:144264. Dai X, Xin Y, Xu W, Tian X, Wei X, Zhang H. Herfs M, Hubert P, Suarez-Carmona M, Reschner
Batlle E, Sancho E, Francí C, Domínguez D, Mon- CBP-mediated Slug acetylation stabilizes Slug A, Saussez S, Berx G, et al. Regulation of p63
far M, Baulida J, et al. The transcription factor and promotes EMT and migration of breast isoforms by snail and slug transcription fac-
snail is a repressor of E-cadherin gene expres- cancer cells. Sci China Life Sci. 2020. tors in human squamous cell carcinoma. Am
sion in epithelial tumour cells. Nat Cell Biol. Dang TT, Westcott JM, Maine EA, Kanchwala M, J Pathol. 2010;176(4):1941–9.
2000;2(2):84–9. Xing C, Pearson GW. ΔNp63α induces the ex- Hong T, Watanabe K, Ta CH, Villarreal-Ponce A,
Bierie B, Pierce SE, Kroeger C, Stover DG, Patta- pression of FAT2 and Slug to promote tumor Nie Q, Dai X. An Ovol2-Zeb1 Mutual Inhibi-
biraman DR, Thiru P, et al. Integrin-β4 iden- invasion. Oncotarget. 2016;7(19):28592–611. tory Circuit Governs Bidirectional and Multi-
tifies cancer stem cell-enriched populations of Daugaard I, Sanders KJ, Idica A, Vittayarukskul step Transition between Epithelial and Mes-
partially mesenchymal carcinoma cells. Proc K, Hamdorf M, Krog JD, et al. miR-151a in- enchymal States. PLOS Comput Biol. 2015;
Natl Acad Sci USA. 2017; 114(12): E2337– duces partial EMT by regulating E-cadherin 11(11):e1004569.
2346. in NSCLC cells. Oncogenesis. 2017;6(7):e366. Hu N, Strobl-Mazzulla PH, Bronner ME. Epigen-
Biswas K, Jolly MK, Ghosh A. Stability and mean Davis S, Meltzer PS, GEOquery . A bridge be- etic regulation in neural crest development.
residence times for hybrid epithelial/mesen- tween the Gene Expression Omnibus (GEO) Dev Biol. 2014;396(2):159–68.
chymal phenotype. Phys Biol. 2019; 16(2): and BioConductor. Bioinformatics. 2007; 23: Hu Y, Nie Q, Dai M, Chen F, Wu H. Histone
025003. 1846–7. Deacetylases Inhibit the Snail2-Mediated EMT
Bocci F, Tripathi SC, Vilchez Mercedes SA, Font-Clos F, Zapperi S, La Porta CAM. Topogra- During Metastasis of Hepatocellular Carcino-
George JT, Casabar JP, Wong PK, et al. NRF2 phy of epithelial-mesenchymal plasticity. ma Cells. Front Cell Dev Biol. 2020;8:752.
activates a partial Epithelial-Mesenchymal Proc Natl Acad Sci USA. 2018;115(23):59027. Huang B, Jia D, Feng J, Levine H, Onuchic JN, Lu
Transition and is maximally present in a hy- Fraile EA, Chavdoula E, Laliotis GI, Anastas V, M. RACIPE: a computational tool for model-
brid Epithelial/Mesenchymal phenotype. In- Serebrennikova O, Paraskevopoulou MD, et ing gene regulatory circuits using randomiza-
tegr Biol (Camb). 2019;11(6):251–63. al. The inhibition of KDM2B promotes the tion. BMC Syst Biol. 2018;12(1):74.
Bolós V, Peinado H, Pérez-Moreno MA, Fraga differentiation of basal-like breast cancer cells Huang RY, Wong MK, Tan TZ, Kuay KT, Ng AH,
MF, Esteller M, Cano A. The transcription via the posttranslational destabilization of Chung VY, et al. An EMT spectrum defines
factor Slug represses E-cadherin expression SLUG. bioRxiv. 2020;109819. an anoikis-resistant and spheroidogenic in-
and induces epithelial to mesenchymal transi- Ganesan R, Mallets E, Gomez-Cambronero J. The termediate mesenchymal state that is sensi-
tions: a comparison with Snail and E47 re- transcription factors Slug (SNAI2) and Snail tive to e-cadherin restoration by a src-kinase
pressors. J Cell Sci. 2003;116(Pt 3):499–511. (SNAI1) regulate phospholipase D (PLD) inhibitor, saracatinib (AZD0530). Cell Death
Caramel J, Papadogeorgakis E, Hill L, Browne GJ, promoter in opposite ways towards cancer Dis. 2013;4:e915.
Richard G, Wierinckx A, et al. A Switch in the cell invasion. Mol Oncol. 2016;10(5):663–76. Hudson LG, Newkirk KM, Chandler HL, Choi C,
Expression of Embryonic EMT-Inducers . Fossey SL, Parent AE, et al. Cutaneous wound
Drives the Development of Malignant Mela- Godin L, Balsat C, Eycke Yvan, Allard J, Royer C, reepithelialization is compromised in mice
noma. Cancer Cell. 2013;24(4):466–80. Remmelink M, et al. A Novel Approach for lacking functional Slug (Snai2). J Dermatol
Celià-Terrassa T, Kang Y. Distinctive properties Quantifying Cancer Cells Showing Hybrid Sci. 2009;56(1):19–26.
of metastasis-initiating cells. Genes Dev. Epithelial / Mesenchymal States in Large Se- Hwang MH, Cho KH, Jeong KJ, Park YY, Kim JM,
2016;30(8):892–908. ries of Tissue Samples : Towards a New Prog- Yu SL, et al. RCP induces Slug expression and
Celià-Terrassa T, Meca-Cortés O, Mateo F, Mar- nostic Marker. Cancers (Basel). 2020; 12(4): cancer cell invasion by stabilizing β1 integrin.
tínez de Paz A, Rubio N, Arnal-Estapé A, et al. 906. Oncogene. 2017;36(8):1102–11.
Epithelial-mesenchymal transition can sup- Goswami CP, Nakshatri H. PROGgeneV2 : en- Idoux-Gillet Y, Nassour M, Lakis E, Bonini F,
press major attributes of human epithelial tu- hancements on the existing database. BMC Theillet C, Du Manoir S, et al. Slug/Pcad path-
mor-initiating cells. J Clin Invest. 2012; Cancer. 2014;14::970. way controls epithelial cell dynamics in mam-
122(5):1849–68. Gras B, Jacqueroud L, Wierinckx A, Lamblot C, mary gland and breast carcinoma. Oncogene.
Chakraborty P, George JT, Tripathi S, Levine H, Fauvet F, Lachuer J, et al. Snail family mem- 2018;37(5):578–88.
Jolly MK. Comparative study of transcrip- bers unequally trigger EMT and thereby differ Jolly MK, Celià-Terrassa T. Dynamics of Pheno-
tomics-based scoring metrics for the epitheli- in their ability to promote the neoplastic typic Heterogeneity Associated with EMT
al-hybrid-mesenchymal spectrum. Front Bio- transformation of mammary epithelial cells. and Stemness during Cancer Progression. J
eng Biotechnol. 2020;8:220. PLoS One. 2014;9(3):e92254. Clin Med. 2019;8(10):1542.
Chen Y, Gridley T. Compensatory regulation of Guo W, Keckesova Z, Donaher JL, Shibue T, Jolly MK, Boareto M, Huang B, Jia D, Lu M, Ben-
the Snai1 and Snai2 genes during chondro- Tischler V, Reinhardt F, et al. Slug and Sox9 Jacob E, et al. Implications of the hybrid epi-
genesis. J Bone Miner Res. 2013; 28(6): 1412– cooperatively determine the mammary stem thelial/mesenchymal phenotype in metasta-
21. cell state. Cell. 2012;148(5):1015–28. sis. Front Oncol. 2015;5:155.
12 Cells Tissues Organs Subbalakshmi/Sahoo/Biswas/Jolly
DOI: 10.1159/000512520Jolly MK, Tripathi SC, Jia D, Mooney SM, Celik- Leroy P, Mostov KE. Slug Is Required for Cell Sur- Savagner P, Yamada KM, Thiery JP. The zinc-fin-
tas M, Hanash SM, et al. Stability of the hybrid vival during Partial Epithelial-Mesenchymal ger protein slug causes desmosome dissocia-
epithelial/mesenchymal phenotype. Oncotar- Transition of HGF-induced tubulogenesis. tion, an initial and necessary step for growth
get. 2016;7(19):27067–84. Mol Biol Cell. 2007;18(5):1943–52. factor-induced epithelial-mesenchymal tran-
Jolly MK, Boareto M, Debeb BG, Aceto N, Far- Li X, Strietz J, Bleilevens A, Stickeler E, Maurer J. sition. J Cell Biol. 1997;137(6):1403–19.
ach-Carson MC, Woodward WA, et al. In- Chemotherapeutic Stress Influences Epitheli- Savagner P, Kusewitt DF, Carver EA, Magnino F,
flammatory Breast Cancer: a model for inves- al-Mesenchymal Transition and Stemness in Choi C, Gridley T, et al. Developmental tran-
tigating cluster-based dissemination. NPJ Cancer Stem Cells of Triple-Negative Breast scription factor slug is required for effective
Breast Cancer. 2017a;3:21. Cancer. Int J Mol Sci. 2020;21(2):404. re-epithelialization by adult keratinocytes. J
Jolly MK, Tripathi SC, Somarelli JA, Hanash SM, Liao T-T, Yang M-H. Hybrid Epithelial/Mesen- Cell Physiol. 2005;202(3):858–66.
Levine H. Epithelial-mesenchymal plasticity : chymal State in Cancer Metastasis: Clinical Saxena K, Srikrishnan S, Celia-Terrassa T, Jolly
How have quantitative mathematical models Significance and Regulatory Mechanisms. MK. OVOL1/2: drivers of epithelial differen-
helped improve our understanding ?. Mol Cells. 2020;9(3):623. tiation in development, disease and repro-
Oncol. 2017b;11:739–54. Liu YN, Yin JJ, Abou-Kheir W, Hynes PG, Casey gramming. Cells Tissues Organs. 2020.
Jolly MK, Mani SA, Levine H. Hybrid epithelial/ OM, Fang L, et al. MiR-1 and miR-200 inhib- Shafran JS, Jafari N, Casey AN, Győrffy B, Denis
mesenchymal phenotype(s): The ‘fittest’ for it EMT via Slug-dependent and tumorigene- GV. BRD4 regulates key transcription factors
metastasis? Biochim Biophys Acta Rev Can- sis via Slug-independent mechanisms. Onco- that drive epithelial–mesenchymal transition
cer. 2018;1870:151–7. gene. 2013;32(3):296–306. in castration-resistant prostate cancer. Pros-
Jolly MK, Somarelli JA, Sheth M, Biddle A, Tripa- Lu M, Jolly MK, Levine H, Onuchic JN, Ben-Jacob tate Cancer Prostatic Dis. 2020. .
thi SC, Armstrong AJ, et al. Hybrid epithelial/ E. MicroRNA-based regulation of epithelial- Slabáková E, Pernicová Z, Slavíčková E,
mesenchymal phenotypes promote metasta- hybrid-mesenchymal fate determination. Staršíchová A, Kozubík A, Souček K. TGF-
sis and therapy resistance across carcinomas. Proc Natl Acad Sci USA. 2013; 110(45): β1-induced EMT of non-transformed pros-
Pharmacol Ther. 2019;194:161–84. 18144–9. tate hyperplasia cells is characterized by early
Jury D, Daugaard I, Sanders KJ, Hansen LL, Agal- Mistry DS, Chen Y, Wang Y, Zhang K, Sen GL induction of SNAI2/Slug. Prostate. 2011;
liu D, Pedersen IM. miR-151a enhances Slug SNAI2 Controls the Undifferentiated State of 71(12):1332–43.
dependent angiogenesis. Oncotarget. 2020; Human Epidermal Progenitor Cells. Stem Srivastava K, Pickard A, Craig SG, Quinn GP,
11(23):2160–71. Cells. 2014;32(12):3209–18. Lambe SM, James JA, et al. ΔNp63γ/SRC/Slug
Kahonouva Z, Remsik J, Fedr R, Bouchal J, Moon JH, Lee SH, Koo BS, Kim JM, Huang S, Cho Signaling Axis Promotes Epithelial-to-Mes-
Mickova A, Slabakova E, et al. Slug-express- JH, et al. Slug is a novel molecular target for enchymal Transition in Squamous Cancers.
ing mouse prostate epithelial cells have in- head and neck squamous cell carcinoma Clin Cancer Res. 2018;24(16):3917–27.
creased stem cell potential. Stem Cell Res. stem-like cells. Oral Oncol. 2020;111:104948. Steinbichler TB, Dudas J, Ingruber J, Glueckert R,
2020;46:101844doi. Mooney SM, Jolly MK, Levine H, Kulkarni P. Sprung S, Fleischer F, et al. Slug Is A Surrogate
Kato Y, Kondo S, Itakura T, Tokunaga M, Hata- Phenotypic plasticity in prostate cancer: Role Marker of Epithelial to Mesenchymal Transi-
yama S, Katayama K, et al. SNAIL- and of intrinsically disordered proteins. Asian J tion (EMT) in Head and Neck Cancer. J Clin
SLUG-induced side population phenotype of Androl. 2016;18(5):704–10. Med. 2020;9(7):2061.
HCT116 human colorectal cancer cells and its Nakamura R, Ishii H, Endo K, Hotta A, Fujii E, Stemmler MP, Eccles RL, Brabletz S, Brabletz T.
regulation by BET inhibitors. Biochem Bio- Miyazawa K, et al. Reciprocal expression of Non-redundant functions of EMT transcrip-
phys Res Commun. 2020;521(1):152–7. slug and snail in human oral cancer cells. tion factors. Nat Cell Biol. 2019;21(1):102–12.
Kim BN, Ahn DH, Kang N, Yeo CD, Kim YK, Lee PLoS One. 2018;13(7):e0199442. Sterneck E, Poria DK, Balamurugan K. Slug and
KY, et al. TGF-β induced EMT and stemness Nassour M, Idoux-Gillet Y, Selmi A, Côme C, E-Cadherin: Stealth Accomplices?. Front Mol
characteristics are associated with epigenetic Faraldo ML, Deugnier MA, et al. Slug controls Biosci. 2020;7:138.
regulation in lung cancer. Sci Rep. 2020;10(1): stem/progenitor cell growth dynamics during Sun Y, Song GD, Sun N, Chen JQ, Yang SS. Slug
10597. mammary gland morphogenesis. PLoS One. overexpression induces stemness and pro-
Kröger C, Afeyan A, Mraz J, Eaton EN, Reinhardt 2012;7(12):e53498. motes hepatocellular carcinoma cell invasion
F, Khodor YL, et al. Acquisition of a hybrid Nieto MA, Huang RY, Jackson RA, Thiery JP. and metastasis. Oncol Lett. 2014;7(6):1936–40.
E/M state is essential for tumorigenicity of EMT: 2016. Cell. 2016;166(1):21–45. Sundararajan V, Tan M, Tan TZ, Ye J, Thiery JP,
basal breast cancer cells. Proc Natl Acad Sci Pastushenko I, Blanpain C. EMT Transition Huang RY. SNAI1 recruits HDAC1 to suppress
USA. 2019;116(15):7353–62. States during Tumor Progression and Metas- SNAI2 transcription during epithelial to mes-
Kuang J, Min L, Liu C, Chen S, Gao C, Ma J, et al. tasis. Trends Cell Biol. 2019;29(3):212–26. enchymal transition. Sci Rep. 2019;9(1):8295.
RNF8 promotes epithelial-mesenchymal tran- Pastushenko I, Brisebarre A, Sifrim A, Fioramon- Sundararajan V, Pang QY, Choolani M, Huang
sition in lung cancer cells via stabilization of ti M, Revenco T, Boumahdi S, et al. Identifica- RY-J. Spotlight on the Granules (Grainyhead-
Slug. Mol Cancer Res. 2020;18(11):1638–49. tion of the tumour transition states occurring Like Proteins) – From an Evolutionary Con-
Kumar B, Uppuladinne MV, Jani V, Sonavane U, during EMT. Nature. 2018;556(7702):463–8. served Controller of Epithelial Trait to Pio-
Joshi RR, Bapat SA. Auto-regulation of Slug Peiró S, Escrivà M, Puig I, Barberà MJ, Dave N, neering the Chromatin Landscape. Front Mol
mediates its activity during epithelial to mes- Herranz N, et al. Snail1 transcriptional re- Biosci. 2020;7:213.
enchymal transition. Biochim Biophys Acta. pressor binds to its own promoter and con- Swain SD, Grifka-Walk HN, Gripentrog J, Lehm-
2015;1849(9):1209–18. trols its expression. Nucleic Acids Res. 2006; ann M, Deuling B, Jenkins B, et al. Slug and
Kurrey NK, Jalgaonkar SP, Joglekar AV, Ghanate 34(7):2077–84. Snail have differential effects in directing co-
AD, Chaskar PD, Doiphode RY, et al. Snail Phillips S, Prat A, Sedic M, Proia T, Wronski A, lonic epithelial wound healing and partially
and slug mediate radioresistance and chemo- Mazumdar S, et al. Cell-state transitions regu- mediate the restitutive effects of butyrate. Am
resistance by antagonizing p53-mediated lated by SLUG are critical for tissue regenera- J Physiol Gastrointest Liver Physiol. 2019;
apoptosis and acquiring a stem-like pheno- tion and tumor initiation. Stem Cell Reports. 317(4):G531–44.
type in ovarian cancer cells. Stem Cells 2009; 2014;2(5):633–47. Tang B, Qi G, Tang F, Yuan S, Wang Z, Liang X,
27(9):2059–68. Ribeiro AS, Paredes J. P-Cadherin Linking Breast et al. Aberrant JMJD3 expression upregulates
Lamouille S, Xu J, Derynck R. Molecular mecha- Cancer Stem Cells and Invasion: A Promising slug to promote migration, invasion, and
nisms of epithelial-mesenchymal transition. Marker to Identify an “Intermediate/Metasta- stem cell-like behaviors in hepatocellular car-
Nat Rev Mol Cell Biol. 2014;15(3):178–96. ble” EMT State. Front Oncol. 2015;4:371. cinoma. Cancer Res. 2016;76(22):6520–32.
SLUG Stabilizes a Hybrid E/M Phenotype Cells Tissues Organs 13
DOI: 10.1159/000512520You can also read

















































