Cutting Edge: Mast Cells Critically Augment Myeloid-Derived Suppressor Cell Activity
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Cutting Edge: Mast Cells Critically Augment
Myeloid-Derived Suppressor Cell Activity
Sheinei J. Saleem,*,1 Rebecca K. Martin,*,1 Johanna K. Morales,†
Jamie L. Sturgill,* David R. Gibb,* Laura Graham,‡,x Harry D. Bear,‡,x
Masoud H. Manjili,*,‡ John J. Ryan,† and Daniel H. Conrad*
Myeloid-derived suppressor cells (MDSCs) are primar- the release of small soluble oxidizers, the depletion of essential
ily recognized for their immunosuppressive properties amino acids from the local extracellular environment, and the
in malignant disease. However, their interaction with impairment of T cell/Ag recognition, all ultimately leading
other innate immune cells and their regulation of im- to T cell suppression (1). This MDSC-mediated immune
mune responses, such as in parasitic infection, necessi- suppression and impairment of cancer immunotherapy has
tate further characterization. We used our previously been extensively investigated (2). However, by limiting immune
published mouse model of MDSC accumulation to ex- activation, MDSCs can serve a beneficial role in transplanta-
amine the immunoregulatory role of MDSCs in B16 tion, autoimmunity, and sepsis (3). This dual role of MDSCs
melanoma metastasis and Nippostrongylus brasiliensis depends upon NO production, the cytokine milieu, and the
infection. In this study, we demonstrate that the ac- interaction among MDSCs, T cells, and NK cells (3–6).
tivity of MDSCs is dependent on the immune stimuli Another interaction that may be physiologically significant
and subset induced. Monocytic MDSCs predictably is MDSC/mast cell (MC) collaboration. Although MCs
suppressed antitumor immune responses but granulo- have been well documented to mediate allergic inflammation,
cytic MDSCs surprisingly enhanced the clearance of their demonstrated involvement in neoplastic disease is more
N. brasiliensis infection. Intriguingly, both results were recent. MCs were shown to accumulate in the tumor mi-
dependent on MDSC interaction with mast cells (MCs), croenvironment and correlate with poor prognosis (7). This
as demonstrated by adoptive-transfer studies in MC- is largely due to the secretion of various proinflammatory
deficient (KitWsh/Wsh) mice. These findings were further cytokines produced by the MCs (8). Additionally, MCs
supported by ex vivo cocultures of MCs and MDSCs, contribute to MDSC recruitment to the tumor site via the
indicating a synergistic increase in cytokine production. production of 5-lipoxygenase and IL-17 (9, 10). Given that
MC-deficient mice have weakened tumor growth and that
Thus, MCs can enhance both immunosuppressive and
MCs recruit MDSCs to the tumor microenvironment, it is
immunosupportive functions of MDSCs. The Journal
necessary to further examine the contribution of this inter-
of Immunology, 2012, 189: 511–515.
action to tumor metastasis. Furthermore, although several
studies indicate the expansion, recruitment, and activity of
MDSCs in parasitic infections, they do not clarify the extent
A
cute inflammatory reactions stimulate the develop-
ment and recruitment of myeloid lineage cells, in- to which MDSCs regulate this immune response (11).
cluding neutrophils and macrophages. Unresolved Therefore, we analyzed the effect of MDSCs in anti-
inflammation increases myelopoiesis, shifting the distribution helminth immunity and B16 metastasis, as well as the in-
of myeloid subpopulations. This results in the premature bone teraction with MCs during the immune response. We used
marrow release of a heterogeneous population of mononuclear our previously published mouse model of MDSC accumu-
(CD11b+Ly6Chigh) and polymorphonuclear (CD11b+Ly6- lation in a tumor-free environment (12). In these mice, the
Ghigh) cells, known as myeloid-derived suppressor cells overexpression of a distintegrin and metalloproteinase 10
(MDSCs). These cells are widely studied in the context of (ADAM10) promotes the expansion of MDSCs that are
neoplasia. MDSCs exert their proneoplastic effects through analogous to tumor-induced MDSCs. Adoptive-transfer (AT)
*Department of Microbiology and Immunology, Virginia Commonwealth University, Rich- The online version of this article contains supplemental material.
mond, VA 23298; †Department of Biology, Virginia Commonwealth University, Richmond,
Abbreviations used in this article: ADAM10, a distintegrin and metalloproteinase 10;
VA 23298; ‡Massey Cancer Center, Virginia Commonwealth University, Richmond, VA
AIT, adoptive immunotherapy; AT, adoptive transfer; A10Tg, a distintegrin and metal-
23298; and xDivision of Surgical Oncology, Department of Surgery, Virginia Commonwealth
loproteinase 10 Tg; BMMC, bone marrow-derived mast cell; CYP, cyclophosphamide;
University, Richmond, VA 23298
GEM, gemcitabine; L3, stage 3 larvae; MC, mast cell; MDSC, myeloid-derived sup-
1
S.J.S. and R.K.M. contributed equally to this work. pressor cell; VCU, Virginia Commonwealth University; WT, wild-type.
Received for publication March 1, 2012. Accepted for publication May 22, 2012.
Copyright Ó 2012 by The American Association of Immunologists, Inc. 0022-1767/12/$16.00
This work was supported by National Institutes of Health Grants RO1AI19697 and
U19AI077435, Project 2 (to D.H.C.) and F31CA159877 (to S.J.S.).
Address correspondence and reprint requests to Dr. Daniel H. Conrad, Virginia Com-
monwealth University, P.O. Box 980678, Richmond, VA 23298. E-mail address:
dconrad@vcu.edu
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1200647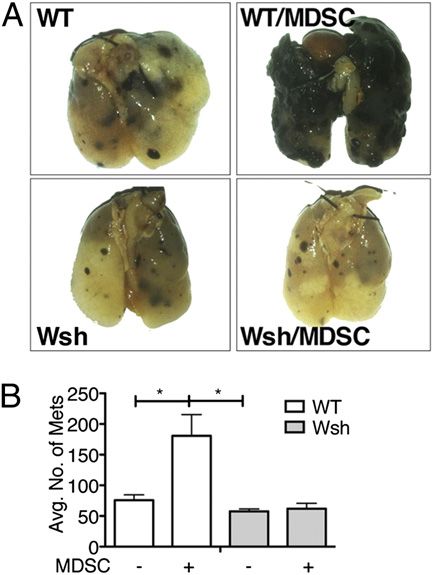
512 CUTTING EDGE: MAST CELLS REGULATE MDSC ACTIVITY
studies using either C57BL/6 (wild-type [WT]) or MC- Adoptive immunotherapy of B16 melanoma
deficient KitWsh/Wsh mice were performed in parallel. Using T cells were prepared for adoptive immunotherapy (AIT), as previously de-
two model systems, we demonstrate that the activity of scribed (13). Recipient mice were injected i.v. with 0.25 3 106 B16 cells.
MDSCs is variable, because granulocytic MDSCs enhanced One day prior to AIT, mice were treated i.p. with 2 mg cyclophosphamide
(CYP) and 1.2 mg gemcitabine (GEM) and were treated every 5 d with GEM
antiparasite immunity, whereas monocytic MDSCs promoted only. After 14 d, lung nodules were counted, as previously described (13, 14);
B16 metastasis. Intriguingly, both activities depend upon the if too many to count, they were assigned a count of 250.
presence of MCs. This intercellular communication leads to
augmentation of Th2-polarizing cytokines that are beneficial N. brasiliensis infection, GEM treatment, and AT
in the case of parasitic infection but maladaptive in neoplasia. N. brasiliensis larvae were generously provided by Joe Urban (Agriculture
Thus, our findings indicate that MDSCs have an immuno- Research Station, Beltsville, MD) and were maintained as previously de-
modulatory role that is largely dependent on the type of scribed (15). Each experimental group of mice was infected with ∼650 stage 3
larvae (L3) N. brasiliensis. Fecal egg burdens were enumerated on days 5–13,
stimuli, the MDSC population involved, and the presence and serum was collected by tail vein nick. For mice injected with GEM,
of MCs. 1.2 mg was injected i.p. starting on day 0 and repeated every 5 d throughout
the experiment. For AT studies, WT mice were injected with 5–10 3 106
naive A10Tg MDSCs via tail vein injection starting at day 0 of infection and
Materials and Methods repeated every 3 d thereafter. Additional groups of animals were sacrificed
Mice on day 7 postinfection, and the proximal halves of the small intestines were
harvested. Adult worms were purified and enumerated.
Mice were kept at Virginia Commonwealth University (VCU) in accordance
with the humane treatment of laboratory animals set forth by the National
Institutes of Health and the American Association for the Accreditation of Statistical analysis
Laboratory Animal Care. C57BL/6 ADAM10 Tgs (A10Tgs) were generated
The p values were calculated using unpaired two-tailed Student t tests or
by the VCU Transgenic Mouse Core, as previously described (12), and lit-
unpaired Mann–Whitney analysis. Error bars represent the SEM between
termates were used as controls. Pmel-1 and KitWsh/Wsh mice were purchased
samples.
from The Jackson Laboratory. All mouse protocols were approved by the
VCU Institutional Animal Care and Use Committee.
Cell lines Results and Discussion
MDSCs from A10Tg mice are phenotypically and functionally
The LLC cell line was obtained from the American Type Culture Collection. analogous to tumor-induced MDSCs
B16 melanoma and GM-CSF–B16 cells were provided by H.D.B. and
maintained as previously described (13). Mouse bone marrow-derived MCs Although MDSC accumulation is a byproduct of ADAM10
(BMMCs) were generated from mice through isolation and subsequent overexpression in early hematopoietic progenitors, ADAM10
culture of bone marrow cells in complete RPMI 1640.
expression is not altered in these cells (12). To ascertain
whether A10Tg-derived MDSCs are functional, ex vivo
Isolation of MDSCs and AT
T cell suppression assays were performed. A10Tg MDSCs
Spleens were harvested from A10Tg or LLC-bearing WT mice, depending inhibited T cell proliferation of both A10Tg and WT purified
upon the experimental protocol. They were then dispersed into single-cell
suspensions and filtered through 70-mm nylon mesh strainers (Invitrogen).
T cells under polyclonal (Supplemental Fig. 1A, 1B) and
Erythrocytes were lysed using an ammonium chloride potassium lysing Ag-specific (Supplemental Fig. 1C) conditions in a manner
buffer (Quality Biological). Gr-1+, Ly6G+, or Ly6C+ cells were purified from parallel to tumor-derived MDSCs (Supplemental Fig. 1D).
A10Tg spleens using the EasySep PE-Selection kit (Stem Cell Technologies), Tumor-derived MDSCs can be divided into granulocytic
according to the manufacturer’s protocol. For AT studies, 5–10 3 106 naive
A10Tg MDSCs were injected into the tail vein of each experimental group. (CD11b+Ly6Ghigh) and more immunosuppressive mono-
For T cell depletion, mice were injected i.p. with 200 mg anti-CD4 (GK1.5) cytic (CD11b+Ly6Chigh) subsets (16). Accordingly, monocytic
and anti-CD8 (2.43) Abs on days 23, 22, 21, 0, 5, and 10. MDSCs possessed more suppressive potential. Furthermore,
similar to T cells from tumor-bearing hosts, A10Tg T cells
Arginase and NO detection exhibited reduced levels of L-selectin (CD-62L), a receptor
MDSC cytoplasmic extracts were prepared from isolated A10Tg- and LLC- required for homing of naive T cells to the lymph nodes
bearing hosts and tested for urea production (DARG-200; BioAssay Systems). (Supplemental Fig. 1E, 1F) (17). MDSCs impair the T cell
NO was measured using Greiss Reagent (G7921; Molecular Probes). response through the activity of arginase 1 and inducible NO
synthase. Accordingly, A10Tg MDSCs had levels of these
T cell suppression assays enzymes that were comparable to tumor-derived MDSCs
For polyclonal T cell activation, CD90.2+ T cells were sorted from spleen (Supplemental Fig. 1G, 1H) (2). These ex vivo observations
and activated with immobilized anti-CD3 (10 mg/ml) and soluble anti-CD28 indicate that MDSCs present in tumor-free A10Tg animals
(1 mg/ml) in 96-well plates. Sorted MDSCs (Ly6G+, Gr-1+, and Ly6G2)
from A10Tg or LLC tumor-bearing mice were then added at increasing behave similarly to tumor-derived MDSCs.
T cell/MDSC ratios. For Ag-specific T cell suppression assays, soluble gp100
(1 mg/ml) was added to defined ratios of pmel-1 transgenic splenocytes and Monocytic MDSCs promote B16 metastasis
A10Tg MDSCs. After 54 h of culture, 1 mCi [3H]thymidine was added to
each well for an additional 18 h, and thymidine incorporation was measured. We used the B16 melanoma model to examine in vivo
activities of MDSCs from A10Tg mice. B16 was injected
BMMC coculture with A10Tg MDSCs i.v.; although not a true metastasis, this allows comparison
BMMCs were derived from femurs of WT naive mice cultured in complete of tumor colonization into the lung with and without MDSCs
RPMI 1640 containing 20% WEHI-3–conditioned medium over a 4-wk and is commonly used to assess AIT (13). When challenged
period. They were resuspended at 0.5 3 106 cells/ml, loaded with 0.5 mg/ml with B16, A10Tg animals were more susceptible to metastasis
mouse IgE, and cultured with MDSCs at a 1:1 ratio in 10 ng/ml IL-3 and than were WT controls (Fig. 1A). The A10Tg animals had
10 ng/ml GM-CSF overnight. MCs were activated by DNP-human serum
albumin (20–100 ng/ml), and supernatants were collected 6 or 18 h after tumor nodules that were too numerous to count and were
cross-linking for ELISA. consequently assigned a count of 250. Parallel to the ex vivo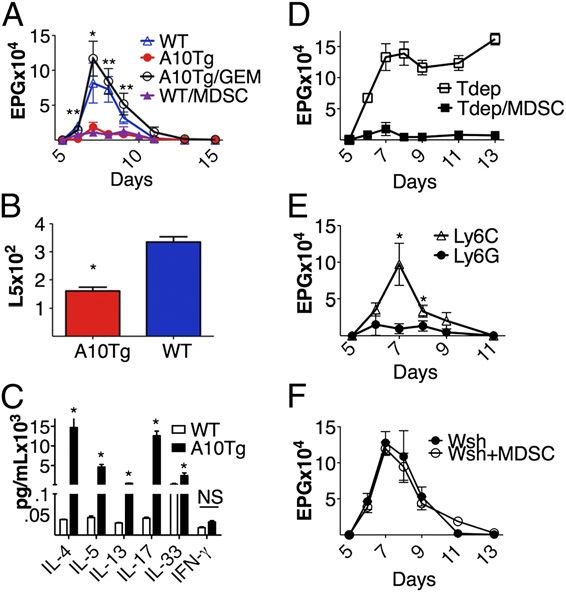
The Journal of Immunology 513
Granulocytic MDSCs enhance the immune response against
N. brasiliensis
Next, we examined the effect of MDSCs on the immune
response in antiparasite immunity. Although MDSC accu-
mulation following N. brasiliensis infection has been re-
ported (11), the direct immunomodulatory role of MDSCs
in anti-N. brasiliensis responses has not been examined. In
N. brasiliensis infection, a WT response is characterized by
production of Th2-associated cytokines that ultimately me-
diate adult worm clearance 10–12 d postinoculation (21).
Following N. brasiliensis challenge, A10Tg animals had sig-
nificantly reduced fecal egg counts compared with WT con-
trols (Fig. 2A). This is highly correlated with 50% reduction
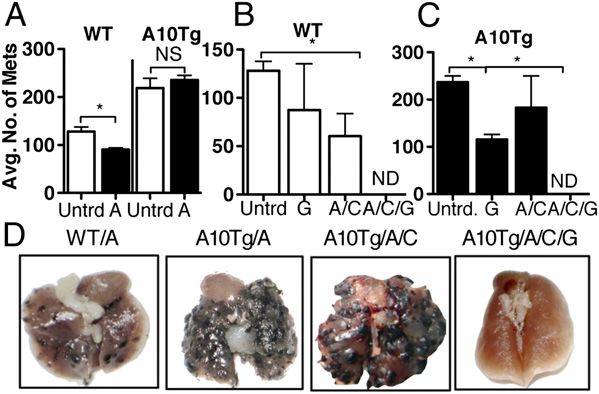
FIGURE 1. MDSC depletion restores the antitumor response and pre-
in adult worm burden in A10Tgs (Fig. 2B). This indicated
vents metastatic progression of B16 melanoma in A10Tg mice. (A) Number
of lung metastases in WT and A10Tg animals challenged with B16 mel- that MDSCs may facilitate parasite clearance. To determine
anoma with (filled bars) or without (open bars) AIT (A), as described whether the increased worm clearance correlated with an in-
in Materials and Methods. Number of lung metastases of WT (B) or A10Tg crease in Th2 cytokines, we examined a panel of Th2 cyto-
(C) mice challenged with B16 and treated as in (A), with the addition kines. Analysis of peak infection (day 7) serum levels of
of CYP (C) and GEM (G). (D) Representative lungs of WT and A10Tg IL-4, IL-5, and IL-13 demonstrated significantly enhanced
mice with AIT, with and without CYP and GEM. Data represent five levels in N. brasiliensis-infected A10Tg animals compared
mice/group. *p , 0.05 versus respective untreated controls. ND, None
detected.
with infected littermates (Fig. 2C). Additionally, IL-17 and
IL-33, which were reported to recruit MDSCs, were also ele-
vated in serum of A10Tg mice (9, 22). These serum cytokines
assay, the suppressive activity of MDSCs was attributed to the decreased as infection began to decline (data not shown).
monocytic population. When WT mice were challenged with To determine whether enhanced N. brasiliensis clearance
B16 and AT of either monocytic or granulocytic MDSCs, is dependent on MDSC activity, MDSCs were depleted from
melanoma metastasized more aggressively in mice given the A10Tg mice. GEM treatment resulted in elevated egg counts
monocytic subset, and granulocytic MDSCs had no effect
(Supplemental Fig. 2A). AIT exerted a minimal effect on A10Tg
mice, suggesting that the presence of MDSCs diminished the
activity of adoptively transferred T cells. Although AIT
treatment did not completely abolish metastasis in WT lungs,
it induced a significant decrease in metastatic lesions. Given
that lymphodepleting chemotherapy is known to enhance
AIT (18), the protocol was modified to incorporate CYP
and GEM. CYP lymphodepletes recipient mice, permitting
homeostatic proliferation of transferred T cells; GEM prefer-
entially decreases MDSC levels in tumor-bearing mice (19,
20). CYP and GEM alone and in combination in WT mice did
not significantly alter metastasis, indicating that they have
minimal effects on tumor cells (Supplemental Fig. 2C). How-
ever, GEM selectively diminished the MDSC population in
A10Tg mice without affecting lymphocyte levels (Supple-
mental Fig. 2B). A10Tg mice treated with GEM alone
exhibited an antitumor response comparable to WT counter-
parts, demonstrating that T cell function is restored upon
MDSC depletion (Fig. 1B, 1C). Additionally, the combination
of GEM, CYP, and AIT (tritherapy) resulted in complete re-
gression of metastasis in both WT and A10Tgs (Fig. 1B, 1C).
Fig. 1D illustrates the significant abrogation of metastasis FIGURE 2. A10Tg mice are resistant to N. brasiliensis infection. (A) Eggs/g
feces were determined at the indicated times in WT and A10Tg mice, as well
arising from tritherapy. Ideal AIT would use tumor-specific
as A10Tgs treated with GEM or WT mice with AT of MDSCs upon in-
T cells and chemotherapy. Thus, the AIT protocol was ad- fection with 650 N. brasiliensis L3 worms. (B) WT and A10Tg mice were
justed to incorporate pmel-1 mice that are TCR transgenic for examined on day seven for adult worm recovery, as described in Materials and
gp100 melanoma peptide (13). Similar results were observed, Methods. (C) Serum was collected at day seven and cytokine analysis was
in that successful AIT required MDSC depletion (Supple- performed using a Milliplex Mouse Cytokine Kit. (D) Eggs/g feces of WT
mental Fig. 2C). Although not shown, A10Tg mice were also mice, with or without MDSC AT, and depleted of T cells, as described in
Materials and Methods. (E) Enumerated eggs/g feces of infected WT mice with
more susceptible to Lewis lung carcinoma compared with WT
AT of either the CD11b+Ly6highLy6Clow or CD11b+Ly6lowLy6Chigh pop-
controls. Thus, MDSCs in A10Tg mice are functionally ulation of MDSCs. (F) Enumerated eggs/g feces of MC-deficient KitWsh/Wsh
analogous to tumor-derived MDSCs, promote tumor me- mice, with and without MDSCs, upon infection with 650 N. brasiliensis
tastasis, and compromise the efficacy of tumor immuno- L3 worms. The data represent five mice/group. *p , 0.05, **p , 0.005 versus
therapy. infected WT controls.
514 CUTTING EDGE: MAST CELLS REGULATE MDSC ACTIVITY
in A10Tg mice similar to WT levels (Fig. 2A). Additionally, to the enhanced N. brasiliensis expulsion. Surprisingly, AT of
N. brasiliensis infection resulted in a modest 4–5-fold in- MDSCs into MC-deficient mice did not enhance N. brasiliensis
crease in MDSCs in WT spleens from days 0 to 14. This expulsion, indicating that an MDSC/MC interaction is critical
level of WT MDSCs is sufficient to promote N. brasiliensis for MDSC-mediated anti-N. brasiliensis responses (Fig. 2F).
clearance and prevent a chronic infection. However, GEM- To assess this interaction ex vivo, MCs were cocultured with
mediated MDSC depletion in WT mice significantly exac- MDSCs. This resulted in a synergistic increase in IL-6, IL-13,
erbated the peak level of N. brasiliensis infection (data not TNF-a, and MIP-1a (Fig. 3).
shown). Taken together, the data indicate that the enhanced
immune response in A10Tgs is a consequence of elevated MCs also contribute to MDSC-mediated immune suppression in B16
MDSC levels. Next, to eliminate off-target effects of GEM, melanoma
A10Tg MDSCs were purified and adoptively transferred to Given that MDSCs require MCs to enhance anti-N. brasi-
WT mice over the course of N. brasiliensis infection. Like liensis immunity, the B16 metastasis study was re-evaluated in
the MDSCs that were observed accumulating following MC-deficient mice. Several groups demonstrated that MCs
N. brasiliensis infection, the transferred MDSCs also con- recruit MDSCs, which accumulate in the tumor microen-
tained both Ly-6G+ and Ly6C+ populations. AT of MDSCs vironment and correlate with poor prognosis. In addition,
resulted in significantly reduced egg counts, comparable to MC-deficient mice have decreased rates of tumor growth that
A10Tgs (Fig. 2A). To determine which MDSC subset was increases to WT levels subsequent to MC reconstitution (26).
responsible for this observation, MDSCs were purified based Therefore, we anticipated that B16 melanoma cells would
upon Ly6G and Ly6C expression and adoptively trans- metastasize more slowly in KitWsh/Wsh mice compared with
ferred into WT N. brasiliensis-infected mice. The Ly6Ghigh WT controls, but it would be enhanced with MDSC AT.
population had the same effect as did total Gr-1+–purified KitWsh/Wsh and WT mice were injected with B16, with and
MDSCs, whereas the adoptively transferred Ly6Chigh pop- without AT of MDSCs. WT mice were more susceptible to
ulation had no effect. This indicates that the granulocytic B16 metastasis compared with KitWsh/Wsh mice. Upon MDSC
population of MDSCs mediates the anti-N. brasiliensis im- AT, WT mice exhibited similar levels of tumor nodules as did
munity (Fig. 2E). The direct role of MDSCs was further B16 A10Tgs, indicating the direct contribution of MDSCs
supported in WT mice that were T cell depleted. These mice to the immune response. However, much to our surprise, AT
exhibited the same rapid rate of clearance as WT adoptively of MDSCs into KitWsh/Wsh mice failed to increase B16 metas-
transferred with MDSCs (Fig. 2A). As expected, N. brasiliensis tasis (Fig. 4). Thus, these results demonstrate that MDSCs
clearance was significantly slower in T cell-depleted WT mice promote B16 melanoma metastasis in an MC-dependent
without AT of MDSCs (Fig. 2D). manner and indicate that MDSC/MC interactions signifi-
cantly enhance the MDSC-mediated suppression of anti-
MCs are required for MDSC-mediated N. brasiliensis clearance
tumor immunity.
Mucosal MC hyperplasia is a hallmark of gastrointestinal hel- The A10Tg mouse affords a unique tool to characterize
minth infection that enhances the immune response, leading the immune-modulatory potential of MDSCs in an envi-
to parasite clearance (23, 24). Mice deficient in MCs exhibit ronment free of established tumor. Because the A10Tgs suffer
delayed clearance kinetics (25). Given that MCs mainly pro- no confounding pathology, this was an ideal system in which
duce Th2-polarizing and proinflammatory cytokines and that to elucidate the immunomodulatory roles of MDSCs. We
MCs chemoattract MDSCs, we examined whether the in-
teraction between the two cell types could be contributing
FIGURE 3. MC/MDSC interaction augments cytokine production.
BMMCs were cocultured with A10Tg MDSCs, as described in Materials FIGURE 4. MC/MDSC interaction is required for MDSC-mediated
and Methods. Supernatants were collected and analyzed for the produc- immune suppression. Photographic representation (A) and quantification
tion of IL-6 (A), IL-13 (B), MIP-1a (C), and TNF-a (D). The data (B) of B16 lung metastasis in control C57 and KitWsh/Wsh mice, with and
represent pooled spleens of at least three mice/group. *p , 0.05 versus without AT of MDSCs. The data are representative of at least five mice/
MCs alone. group. *p , 0.05 versus WT alone or KitWsh/Wsh with MDSCs.The Journal of Immunology 515
used this system to investigate roles for MDSCs in both 9. Yang, Z., B. Zhang, D. Li, M. Lv, C. Huang, G.-X. Shen, and B. Huang. 2010.
Mast cells mobilize myeloid-derived suppressor cells and Treg cells in tumor mi-
neoplasia and helminth infection. In B16 metastasis, mono- croenvironment via IL-17 pathway in murine hepatocarcinoma model. PLoS ONE
cytic MDSCs strongly suppressed the immune response, 5: e8922.
10. Cheon, E. C., K. Khazaie, M. W. Khan, M. J. Strouch, S. B. Krantz, J. Phillips,
whereas granulocytic MDSCs failed to promote metastasis. N. R. Blatner, L. M. Hix, M. Zhang, K. L. Dennis, et al. 2011. Mast cell
However, in N. brasiliensis infection, the granulocytic MDSCs 5-lipoxygenase activity promotes intestinal polyposis in APCDelta468 mice.
enhanced the antiparasitic immune response, and the mono- Cancer Res. 71: 1627–1636.
11. Van Ginderachter, J. A., A. Beschin, P. De Baetselier, and G. Raes. 2010.
cytic MDSCs had no effect. AT studies demonstrated that Myeloid-derived suppressor cells in parasitic infections. Eur. J. Immunol. 40:
this differential immune regulation by MDSCs occurs in an 2976–2985.
12. Gibb, D. R., S. J. Saleem, D.-J. Kang, M. A. Subler, and D. H. Conrad. 2011.
MC-dependent manner. The ex vivo coculture studies indi- ADAM10 overexpression shifts lympho- and myelopoiesis by dysregulating site
cate that MDSC/MC interactions may modulate immune 2/site 3 cleavage products of Notch. J. Immunol. 186: 4244–4252.
13. Le, H. K., L. Graham, C. H. Miller, M. Kmieciak, M. H. Manjili, and H. D. Bear.
responses by increasing cytokine production. MDSCs traffic 2009. Incubation of antigen-sensitized T lymphocytes activated with bryostatin 1 +
to MCs, affording a synergistic Th2-skewed immune re- ionomycin in IL-7 + IL-15 increases yield of cells capable of inducing regression of
melanoma metastases compared to culture in IL-2. Cancer Immunol. Immunother.
sponse. Although undesirable in the context of neoplasia, the 58: 1565–1576.
observed response is ideal in helminth infections. Overall, 14. Lipshy, K. A., P. J. Kostuchenko, G. G. Hamad, C. E. Bland, S. K. Barrett, and
our data indicate that inhibition of MC function may serve H. D. Bear. 1997. Sensitizing T-lymphocytes for adoptive immunotherapy by
vaccination with wild-type or cytokine gene-transduced melanoma. Ann. Surg.
as a novel target to inhibit MDSC activity. Oncol. 4: 334–341.
15. Camberis, M., G. Le Gros, and J. Urban, Jr. 2003. Animal model of Nippostrongylus
brasiliensis and Heligmosomoides polygyrus. Curr. Protoc. Immunol. Chapter 19: Unit
Acknowledgments 19.12.
16. Peranzoni, E., S. Zilio, I. Marigo, L. Dolcetti, P. Zanovello, S. Mandruzzato, and
We thank Natalie Allen, Suzanne Barbour, Madiha Laiq, and Kahley Stanco V. Bronte. 2010. Myeloid-derived suppressor cell heterogeneity and subset defini-
for technical assistance. tion. Curr. Opin. Immunol. 22: 238–244.
17. Hanson, E. M., V. K. Clements, P. Sinha, D. Ilkovitch, and S. Ostrand-Rosenberg.
2009. Myeloid-derived suppressor cells down-regulate L-selectin expression on CD4
+ and CD8+ T cells. J. Immunol. 183: 937–944.
Disclosures 18. Bracci, L., F. Moschella, P. Sestili, V. La Sorsa, M. Valentini, I. Canini,
The authors have no financial conflicts of interest. S. Baccarini, S. Maccari, C. Ramoni, F. Belardelli, and E. Proietti. 2007. Cyclo-
phosphamide enhances the antitumor efficacy of adoptively transferred immune
cells through the induction of cytokine expression, B-cell and T-cell homeostatic
proliferation, and specific tumor infiltration. Clin. Cancer Res. 13: 644–653.
References 19. Teitz-Tennenbaum, S., Q. Li, M. A. Davis, K. Wilder-Romans, J. Hoff, M. Li, and
1. Gabrilovich, D. I., and S. Nagaraj. 2009. Myeloid-derived suppressor cells as reg- A. E. Chang. 2009. Radiotherapy combined with intratumoral dendritic cell vac-
ulators of the immune system. Nat. Rev. Immunol. 9: 162–174. cination enhances the therapeutic efficacy of adoptive T-cell transfer. J. Immunother.
2. Ostrand-Rosenberg, S. 2010. Myeloid-derived suppressor cells: more mechanisms 32: 602–612.
for inhibiting antitumor immunity. Cancer Immunol. Immunother. 59: 1593–1600. 20. Le, H. K., L. Graham, E. Cha, J. K. Morales, M. H. Manjili, and H. D. Bear. 2009.
3. Cuenca, A. G., M. J. Delano, K. M. Kelly-Scumpia, C. Moreno, P. O. Scumpia, Gemcitabine directly inhibits myeloid derived suppressor cells in BALB/c mice
D. M. Laface, P. G. Heyworth, P. A. Efron, and L. L. Moldawer. 2011. A paradoxical bearing 4T1 mammary carcinoma and augments expansion of T cells from tumor-
role for myeloid-derived suppressor cells in sepsis and trauma. Mol. Med. 17: 281–292. bearing mice. Int. Immunopharmacol. 9: 900–909.
4. Bronte, V., E. Apolloni, A. Cabrelle, R. Ronca, P. Serafini, P. Zamboni, 21. Else, K. J., and F. D. Finkelman. 1998. Intestinal nematode parasites, cytokines and
N. P. Restifo, and P. Zanovello. 2000. Identification of a CD11b(+)/Gr-1(+)/ effector mechanisms. Int. J. Parasitol. 28: 1145–1158.
CD31(+) myeloid progenitor capable of activating or suppressing CD8(+) T cells. 22. Perrigoue, J. G., F. A. Marshall, and D. Artis. 2008. On the hunt for helminths:
Blood 96: 3838–3846. innate immune cells in the recognition and response to helminth parasites. Cell.
5. Narita, Y., D. Wakita, T. Ohkur, K. Chamoto, and T. Nishimura. 2009. Potential Microbiol. 10: 1757–1764.
differentiation of tumor bearing mouse CD11b+Gr-1+ immature myeloid cells into 23. Turnquist, H. R., Z. Zhao, B. R. Rosborough, Q. Liu, A. Castellaneta, K. Isse,
both suppressor macrophages and immunostimulatory dendritic cells. Biomed. Res. Z. Wang, M. Lang, D. B. Stolz, X. X. Zheng, et al. 2011. IL-33 expands suppressive
30: 7–15. CD11b+ Gr-1(int) and regulatory T cells, including ST2L+ Foxp3+ cells, and
6. Pastuła, A., and J. Marcinkiewicz. 2011. Myeloid-derived suppressor cells: a double- mediates regulatory T cell-dependent promotion of cardiac allograft survival.
edged sword? Int. J. Exp. Pathol. 92: 73–78. J. Immunol. 187: 4598–4610.
7. Oldford, S. A., I. D. Haidl, M. A. Howatt, C. A. Leiva, B. Johnston, and 24. Miller, H. R. 1996. Mucosal mast cells and the allergic response against nematode
J. S. Marshall. 2010. A critical role for mast cells and mast cell-derived IL-6 in parasites. Vet. Immunol. Immunopathol. 54: 331–336.
TLR2-mediated inhibition of tumor growth. J. Immunol. 185: 7067–7076. 25. Mitchell, L. A., R. B. Wescott, and L. E. Perryman. 1983. Kinetics of expulsion
8. Huang, B., Z. Lei, G.-M. Zhang, D. Li, C. Song, B. Li, Y. Liu, Y. Yuan, of the nematode, Nippostrongylus brasiliensis, in mast-cell deficient W/WV mice.
J. Unkeless, H. Xiong, and Z.-H. Feng. 2008. SCF-mediated mast cell infiltration Parasite Immunol. 5: 1–12.
and activation exacerbate the inflammation and immunosuppression in tumor 26. Starkey, J. R., P. K. Crowle, and S. Taubenberger. 1988. Mast-cell-deficient W/Wv
microenvironment. Blood 112: 1269–1279. mice exhibit a decreased rate of tumor angiogenesis. Int. J. Cancer 42: 48–52.You can also read