FLORAL AND VEGETATIVE MORPHOGENESIS IN CALIFORNIA POPPY
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Int. J. Plant Sci. 166(4):537–555. 2005.
Ó 2005 by The University of Chicago. All rights reserved.
1058-5893/2005/16604-0001$15.00
FLORAL AND VEGETATIVE MORPHOGENESIS IN CALIFORNIA POPPY
(ESCHSCHOLZIA CALIFORNICA CHAM.)
Annette Becker,1 ,* Stefan Gleissbergy, and David R. Smyth*
*School of Biological Sciences, Monash University, Melbourne, Victoria 3800, Australia; and yInstitute of
Systematic Botany, Johannes Gutenberg-Universität, Bentzelweg 9a, 55099 Mainz, Germany
For studies of the evolution of development in angiosperms, early-diverging eudicot taxa are of particular
interest for comparisons with established core eudicot model plants, such as Arabidopsis. Here we provide
a detailed description of shoot and floral development of the basal eudicot California poppy (Eschscholzia
californica). Rosette formation in the vegetative phase is accompanied by increased leaf complexity and shoot
apex size. The flowering phase is characterized by internode elongation, formation of dissected cauline leaves,
terminal flowers, and basipetal inflorescence branching. For developing flowers and fruits, we have defined 14
stages according to important landmark events, from inflorescence primordium initiation through seed
dispersal. Floral organ initiation, morphogenesis, increase in floral meristem size, and the surface structure of
mature floral organs are recorded in detail. The duration of the later floral stages, as well as the path of pollen
tube growth in the gynoecium, is documented. Comparison of California poppy floral development with that
of Arabidopsis indicates considerable differences in terms of organ fusion, whorl proliferation, and variability
of size and organ number between the two species. Transitions in meristem identity from germination to floral
organogenesis were monitored using expression of the developmental control gene EcFLO, the Eschscholzia
ortholog of FLORICAULA/LEAFY. We found that the pattern of expression of EcFLO in the flanks of the
shoot apex is maintained from late embryogenesis to flower initiation, indicating a continuous role for this
gene in meristem function. As flower organs develop, EcFLO expression becomes more restricted to petal and
stamen primordia. Development of the gynoecium occurs without EcFLO expression, indicating that EcFLO
may not be necessary for the activation of C-class genes.
Keywords: Eschscholzia californica, Papaveraceae, flower development, vegetative development, basal
eudicot, FLORICAULA expression.
Introduction A number of reasons in addition to its phylogenetic
position favor California poppy as a model for comparative
The striking diversity of vegetative and floral morphologies developmental genetics: (i) it is an easily cultivated annual-
encountered in angiosperms raises the question of how devel- to-biennial species; (ii) it is diploid, with a genome size only
opment of these traits evolved. Extensive knowledge about ca. 6.5 times larger than that of Arabidopsis (Bennett and
the control of development has been obtained in core eudicot Smith 1976); (iii) a large number of expressed sequence tags
model species such as Arabidopsis thaliana (Brassicaceae), from Eschscholzia have been sequenced by the Floral
Antirrhinum majus (Scrophulariaceae), and Petunia hybrida Genome Project and are publicly available (Soltis et al.
(Solanaceae). However, these plants represent examples from 2002); and, most important, (iv) California poppy is amena-
more advanced orders of rosids (Brassicales) and asterids ble to transgenic manipulation allowing characterization of
(Lamiales and Solanales) in the angiosperm phylogenetic gene functions (Park and Facchini 2000).
tree. To understand evolutionary processes that have led to California poppy is a qualitative long-day plant (Nanda
eudicot diversification, as well as mechanisms that govern and Sharma 1976) and is native in western North America
development in all angiosperms, it will be vital to study the from the Columbia River to Baja California and from the
molecular bases of development of species with a phylogeneti- Pacific Coast eastward into the Great Basin. Within this
cally more basal position. California poppy (Eschscholzia large area it grows in a range of different habitats, spanning
californica Cham.), a member of the Papaveraceae (order Ra- from sea level to 2000 m in altitude and from well-drained
nunculales), is well suited to serve as a model plant for early- soils of dunes to alluvial fans, river terraces, and hillsides.
diverging eudicots. It has been introduced to Chile, New Zealand, Tasmania,
and mainland Australia, where it has become naturalized
(Cook 1962).
1 Author for correspondence; e-mail annette.becker@flower-evodevo
Here we provide descriptive baselines for shoot, inflores-
.com.
cence, and flower development in California poppy to aid on-
Manuscript received November 2004; revised manuscript received January going and future developmental genetic work with this
2005. species. Development of the dissected leaves has been
537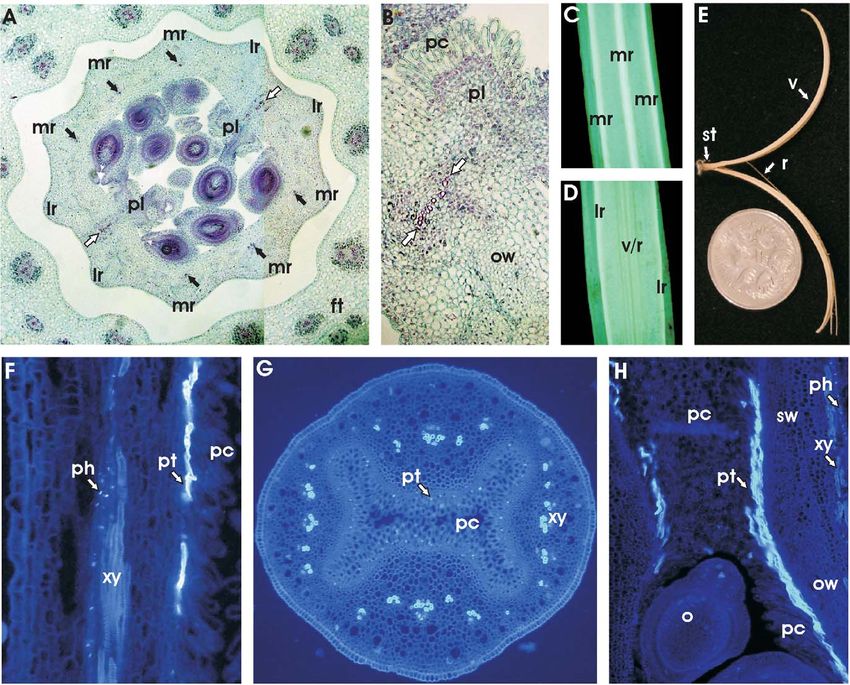
538 INTERNATIONAL JOURNAL OF PLANT SCIENCES
described elsewhere (Gleissberg 2004). Development of the in situ hybridization and SEM analysis, shoots were har-
primary (seedling) shoot through rosette establishment, inflo- vested and fixed at different stages of vegetative and floral
rescence and flower formation, and subsequent inflorescence development, as described below. Germinating embryos
branching is characterized, including leaf heteroblasty and were investigated using material sown on agar plates sup-
enlargement of the shoot apical meristem. We address dis- plemented with Gamborg’s B5 medium.
tinct morphological features of California poppy that con-
trast with those of Arabidopsis, such as phenotypic plasticity,
Analysis of Heteroblasty
terminal flowers, the occurrence of complex dissected leaves
in the flowering region, floral organ fusion, the sequence of To assess leaf form change along the primary shoot, the de-
flower organ initiation and multiplication of floral organ gree of dissection (first-order and total number of segments)
whorls, and the common and unique properties of the gynoe- and the lengths of leaf blade, petiole, and whole leaf were
cium. Inflorescence meristem formation and complete flower documented in five plants. Fully grown foliage leaves were
development until the release of seeds are characterized by individually and carefully removed from growing plants just
scanning electron microscopy and light microscopy, including before senescence to allow undisturbed further development,
a documentation of mature floral organ surface morphology. and measurements were taken on a light box after leaves
Landmarks are used to define stages of flower development were dried in a paper stack.
that will be helpful in aligning developmental observations
and to allow interpretation of gene expression patterns and
SEM and Light Microscopy
defects in California poppy lines generated by transgenic ap-
proaches. For some of the later developmental stages, an ap- Fresh floral buds and mature flowers were vacuum infil-
proximate time line is given. trated and fixed in 5% glutaraldehyde in 0.03 M Pipes buf-
In addition, we provide expression data of the develop- fer at pH 6.8–7.0 for 2 h. Subsequently, the material was
mental control gene ESCHSCHOLZIA CALIFORNICA postfixed for 2 h in 1% aqueous osmium tetroxide solution,
FLORICAULA (EcFLO) in vegetative, inflorescence, and flo- dehydrated by passing it through an ethanol series, and critical-
ral meristems. FLORICAULA/LEAFY-like genes are impor- point dried using liquid CO2. The specimens were mounted
tant developmental regulators with diverse roles in different on stubs, and several outer-whorl organs were removed when
species. Expression of EcFLO in vegetative meristems and necessary. The buds were then gold-coated, and SEM was
during leaf development indicated a role in leaf dissection performed with a Hitachi S570 microscope at 10–15 kV for
(Busch and Gleissberg 2003). Here we show that EcFLO ex- floral stages and 15–30 kV for mature organ surface struc-
hibits a characteristic and constant pattern of expression tures. Measurements of apex size on SEM images were done
through various phases of shoot apical meristem develop- as described by Gleissberg (2004).
ment and identity, indicating a general role in shoot apex For light microscopy, the buds were vacuum infiltrated in
maintenance in both vegetative and reproductive meristems. FAA (2% formaldehyde, 5% acetic acid, 60% ethanol),
EcFLO expression during flower development is discussed fixed for 24 h at 4°C, and dehydrated in an ethanol series.
with respect to the identity of floral organs. Subsequently, they were embedded in Paraplast Plus, sec-
Comparisons of the molecular basis of California poppy tioned, dewaxed, and stained overnight in 1% safranin in
development with that of model species from the rosids and 30% ethanol and then for 3 min in 0.2% alcoholic fast
asterids, as well as within the diverse Ranunculales, will help green (Clark 1981). Preparation of the tissue for fluores-
to decipher the evolution of developmental pathways on dif- cence microscopy was carried out in the same way, except
ferent systematic scales. that the sections were stained overnight in 0.01% aniline
blue in 0.15 M sodium phosphate buffer at pH 10 (Clark
1981). All photographs were processed for publication
Material and Methods
using Adobe Photoshop Elements and Corel Draw 10.
Measurements on in situ sections were done using Photo-
Plants and Growth Conditions
shop 7.0.
For analysis of flower development, Eschscholzia cali-
fornica Cham. cv. Aurantiaca Orange King seeds were pur-
In situ Hybridization
chased from B & T World Seeds, Aigues-Vive, France, and
cultivated in pots of 6 cm diameter using standard potting Tissue fixation, Paraplast embedding, and detection of Ec-
mix. After sowing, the seeds were kept at 4°C for 3 d to im- FLO transcripts on serial sections were done as described by
prove the germination rate and synchrony of development. Zachgo (2002) and Busch and Gleissberg (2003). For in vitro
The plants were then grown under natural daylight supple- transcription of the probe, a PCR fragment was amplified
mented by constant light provided by cool-white fluorescent from a plasmid containing the EcFLO sequence (GenBank
tubes at ca. 25°C. Under these conditions, plants flowered accession number AY188789) using the forward primer
early, although their vigor was somewhat compromised. For EcL01F (AGGCAGGAGCTAGTTACATAAACAAGC) and a T3
analysis of heteroblasty and gene expression, seeds of primer as reverse primer, transcribed with T3 RNA poly-
a wild-type strain of E. californica were purchased from merase. The digoxigenin-labeled antisense probe covered ca.
Larner Seeds, Bolinas, California. Seeds were sown in Jiffy 500 bps in the third exon and 39 untranslated region and was
pots at the end of March and later planted in the field at the used without hydrolysis. Hybridization with a sense probe gave
Botanical Garden of the University of Mainz, Germany. For no specific signal (not shown; Busch and Gleissberg 2003).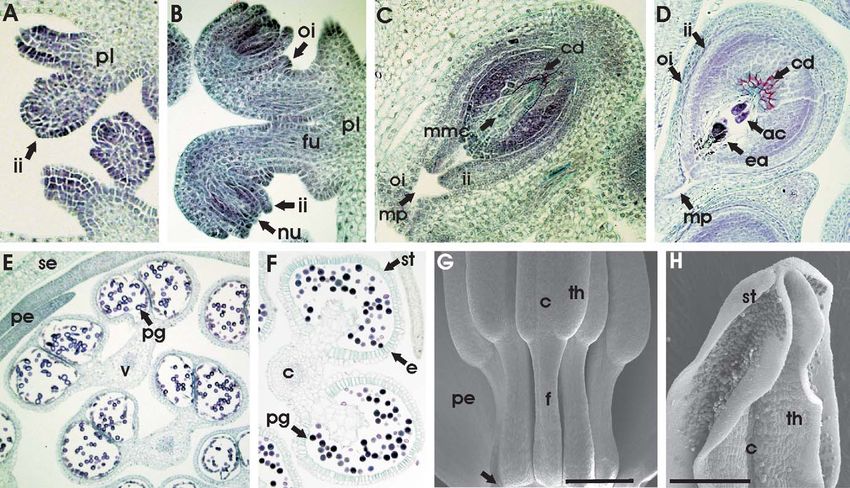
BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 539
Results pinnae ranged between five and seven and that of total seg-
ments between 37 and 98 on each side (84–196 per leaf).
California Poppy Development until Flowering Leaf length increased for several nodes, at least until node 7.
Typically, the cauline leaves were reduced, compared to
After germination, two deeply lobed cotyledons emerge
leaves at a late rosette stage, but they were still highly dis-
from the seed. The shoot apical meristem (SAM) then gener-
sected. They exhibited only a slight decrease, if at all, in the
ates between 13 and 25 highly dissected silver green leaves
number of main segments, but reduction was more obvious
arranged in an alternate fashion, forming a rosette. In contin-
for the total numbers of segments. The last leaf beneath the
uous light conditions, this takes at least 6 wk. With the onset
terminal flower had between 24 and 74 segments on each
of shoot elongation, internodes of the younger leaves expand,
side (48 and 148 per leaf) (e.g., fig. 2B).
marking the conversion of the SAM into an inflorescence
Dynamics of SAM size are illustrated in figure 2C. Because
meristem. Cauline leaves lower on the stem are arranged al-
of leaf initiation, SAM size changes periodically from plasto-
ternately, as in the rosette. The last two leaves formed below
chron to plastochron, resulting in a wide range of values for
the flower are mostly arranged in a subopposite manner,
single age stages. In addition, differences between individual
forming a pseudowhorl (fig. 1B, 1C). All leaves within the
plants may add to SAM size variations. Remarkably, we
flowering region of the shoot remain highly dissected, and no
found that SAM diameters vary approximately within the
reduced bractlike leaves are formed (i.e., it is a frondose in-
same range (between ca. 70 and 185 mm) from germination
florescence; Weberling 1989). After formation of the upper-
through a considerable time of postembryonic development
most cauline leaves, the SAM is transformed into a terminal
(at least 1 mo) (fig. 2C, left). Axillary inflorescence meristems
flower meristem, resulting in a single bud (fig. 1A–1C). Be-
had SAM diameters similar to vegetative meristems. Pro-
fore the apical flower opens, secondary inflorescence meri-
nounced increase of SAM diameter was only observed after
stems develop in the axils of one or both leaves immediately
the transition to floral identity. On the other hand, values ob-
below the primary flower. These meristems, in turn, usually
tained for SAM height (fig. 2C, right) indicate that a moder-
generate two opposite cauline leaves and then a terminal sec-
ate increase of SAM size in this dimension already occurs
ondary flower. Further inflorescence meristems may then de-
during vegetative development, with a further pronounced
velop in the axils of the leaves subtending these secondary
elevation of the apical dome upon flower initiation.
flowers, ultimately generating tertiary flowers, and so on,
thus resulting in a compound cymose inflorescence (fig. 1A–
1D). Most flowering shoots examined in field-grown plants Flower Structure
(66%, N ¼ 57) showed two subopposite cauline leaves sub-
The California poppy flower consists of two sepals, four
tending each flower, but variations were common. Often,
free petals, four to six or more whorls of stamens, and two
a third cauline leaf joined in the pseudowhorl (28%), and
carpels united in a gynoecium (fig. 1E, 1F). The sepals are
one of them was sometimes only rudimentary (fig. 1B).
fused into a caplike structure that is pushed off by the elon-
Rarely, we observed a single cauline leaf below a flower. In
gating petals during the last stage of bud development. The
addition, in 49% of cases only one axillary inflorescence meri-
petals are bright orange in color, darker at the center of the
stem developed from the pseudowhorl; in 42% there were
flower and lighter at its periphery. They are 1–3 cm long and
two. Rarely, three or no axillary inflorescence shoots devel-
wide, depending on the condition of the plant (fig. 1E). The
oped. When there were two, the upper one was always more
petals are arranged in two whorls, with the two medial petals
developmentally advanced than the lower one (fig. 1B, 1C).
occupying the inner and the two lateral petals the outer petal
Further branching of the inflorescence occurs in basipetal
whorl.
direction along the primary shoot. These basal side shoots
Petals and stamens are attached to the top of a floral tube,
produce one to several alternate cauline leaves before they
which surrounds the bottom quarter of the ovary, forming
generate paired cauline leaves and ultimately a terminal
a perigynous flower (fig. 1F). A distinct rim encircles the flo-
flower. Higher-order inflorescence meristems then arise in the
ral tube immediately below where the sepals are inserted.
axils of these cauline leaves as before (fig. 1A, 1D).
Nectaries are not present.
The stamens consist of a short filament and a long anther,
Heteroblastic Changes and Shoot
which is bilobed and bisporangiate. The surface of the lower
Apex Corroboration
part of the filament is darkly pigmented in many flowers. Sta-
Leaf size and form along the primary shoot show consider- mens are numerous, with the number per flower observed to
able heteroblasty between the cotyledons and the terminal vary between 18 and 34. There are usually four stamens in
flower (fig. 2A, 2B). Although we observed large individual the first whorl and six in each subsequent whorl. Many flow-
differences in the measured parameters, particularly at higher ers counted had either 22 (19%) or 28 (26%) stamens, which
nodes, some general patterns are evident. Both leaf length corresponds to four and five complete whorls.
and degree of dissection increased during the vegetative The gynoecium consists of two completely fused carpels,
phase but declined again after the transition to flowering. with a large ovary, a very short style, and four long stigmatic
The maximum degree of dissection was reached at higher protrusions covered with papillae. The ovary is superior, as
nodes (later in development) than the maximum leaf size (fig. its basal region is surrounded by the floral tube but not
2A). Primary leaves started with two or three main acropetal joined to it (fig. 1F). Within the locule of the ovary, two pari-
segments and two to seven total segments on each side (four etal placentae are lined with a layer of papilla-like cells, and
to 14 per leaf). The observed maximum number of lateral two rows of ovules are attached to each placenta.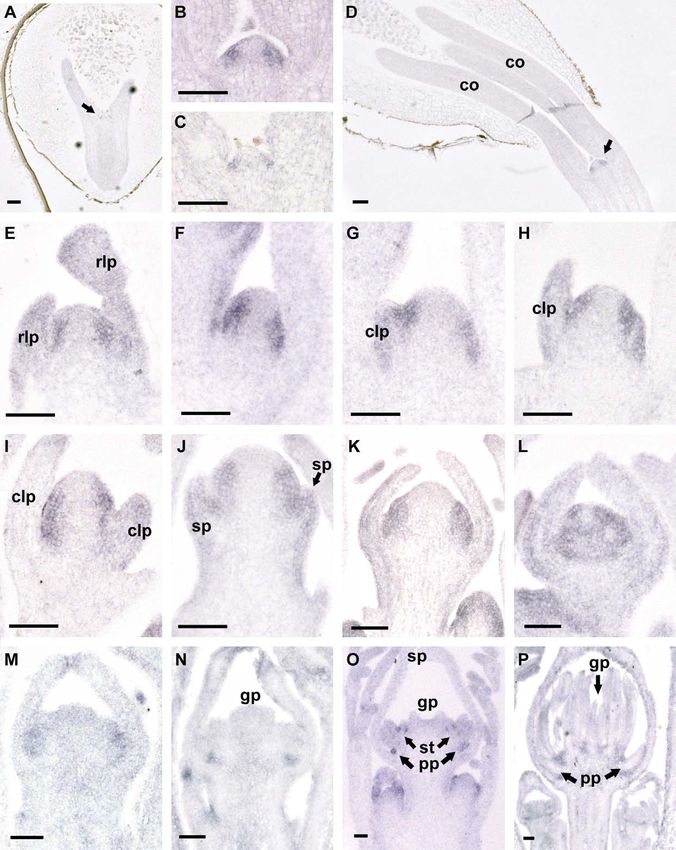
540 INTERNATIONAL JOURNAL OF PLANT SCIENCES
Early Stages of Flower Development (Stages 1–6)
Within the flower developmental stages of Buzgo et al.
(2004), other significant events have been identified (table 1).
Descriptions are based on observations of secondary inflores-
cence shoots.
Stage 1 begins with conversion of the primary SAM into
an inflorescence meristem (not shown) or the formation of
a new inflorescence meristem in the axil of a cauline leaf (fig.
3A). Inflorescence meristems increase in size by lateral out-
growth and sequentially form two (sometimes one or three)
lateral cauline leaf primordia on their flanks. The first leaf
primordium is visible as a small protrusion when the lateral
diameter of the axillary inflorescence meristem is ca. 100 mm
(fig. 3A). The second cauline leaf appears when the inflores-
cence meristem is ca. 160 mm in lateral diameter and the
central dome is ca. 80 mm in diameter (not shown). Remark-
ably, development of the inflorescence meristems in the axils
of two paired cauline leaves is antipodal, as the earlier and
larger leaf of the two paired primordia point in opposite di-
rections (fig. 3A, 3E).
Once the two cauline leaves are formed, the apical dome
above them converts into a floral meristem. When this
reaches ca. 100 mm in diameter (fig. 3B), it is hemispherical
in shape and begins to become separate from the two leaf
primordia. At that time, new axillary inflorescence meristems
may already appear in the cauline leaf axils (fig. 3C).
The first appearance of the sepal ring primordium defines
the beginning of stage 2 (fig. 3D). The central region of the
floral meristem is still hemispherical in shape and ca. 130 mm
in diameter. Sepals arise to form a ring-shaped structure en-
circling the floral dome. Later, it is clear that there are two
primordia, but initially the height of the rim is not obviously
bipartite around the circumference of the bud. At this point
the floral bud (above the cauline leaves) can be divided hori-
zontally into a pedicel zone at the bottom, the sepal zone in
the middle, and the floral dome on top (fig. 3D). During
stage 2, most of the increase in size of the floral bud can be
attributed to the outgrowth of the sepal primordia, as the flo-
ral dome does not increase significantly in size (its lateral
diameter has only increased from 130 to ca. 135 mm by the
end of stage 3). At this stage it is clear that development of
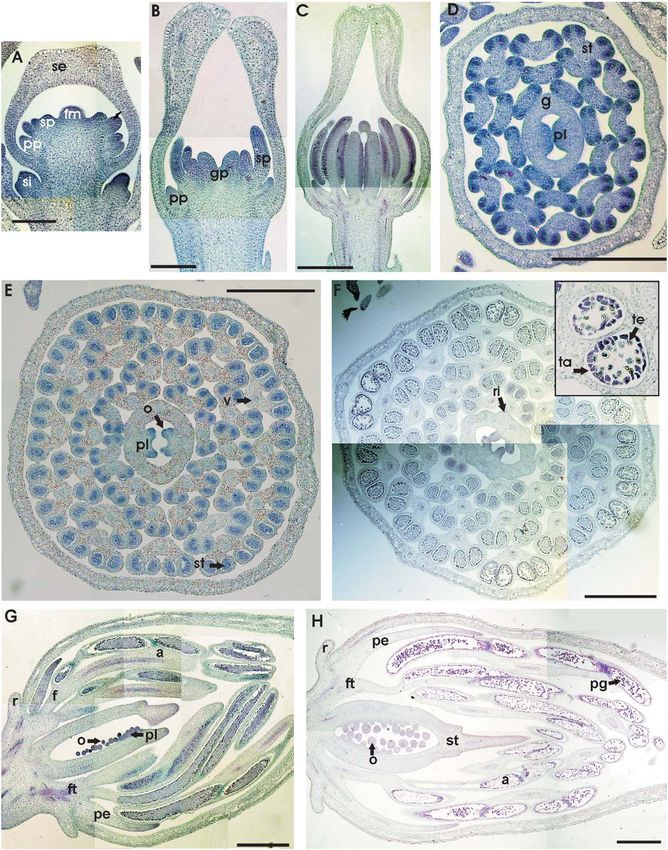
Fig. 1 California poppy morphology. A, California poppy plant the flower buds within the two opposite leaves is not syn-
before opening of the first flower (primary flower, pf ). One secondary chronous, with the lower flower lagging considerably behind
flower (sf ) with its own two cauline leaves has developed in the axil of the upper one (fig. 3E).
a cauline leaf of the primary shoot immediately below the primary Stage 3 begins with the appearance of two lateral petal pri-
flower. Another whole secondary inflorescence (si) has arisen in the mordia as protrusions from the floral apex, with the medial
axil of the cauline leaf further down. This plant is ca. 25 cm tall. B–C, petals initiating soon afterward. The axis connecting the two
Close-ups of pairs of lateral inflorescence shoots within axils of the lateral petal primordia is displaced by an angle of 30° in rela-
uppermost cauline leaves (cl). These leaves are inserted in subopposite
tion to the two cauline leaves (fig. 3F). Subsequently, the petals
orientation on the shoot, in contrast to cauline leaves farther down
are separated from the floral apex by grooves. The lateral
that arise with alternate orientation. Development of the axillary
bud of the upper cauline leaf (on the right in each case) is accelerated. petal primordia are narrower, and the medial petal primordia
Note that in B, a rudimentary third leaf has developed in the are relatively wide. During this stage, the floral dome increases
pseudowhorl. D, Diagram of the California poppy inflorescence
structure. Dark green, rosette leaves; light green, cauline leaves; red,
primary flower; orange, secondary flowers; yellow, tertiary flowers.
Arrows show newly arising inflorescence shoots. E, Top view of stamens (st). Pollen grains have not yet attached to the four long
a California poppy flower at anthesis. The sepals have already stigmatic extensions (sa; front stigmatic branch has been removed).
abscised, and the stigma (sa) extends above the stamens (st); petals The short style is indicated (sy). The lower quarter of the ovary (o) is
(pe) are indicated. F, Lateral view of a longitudinal section of surrounded by the floral tube (ft). A protruding rim (r) surrounds the
a California poppy flower at the beginning of pollen shed from the outside of the top of the floral tube.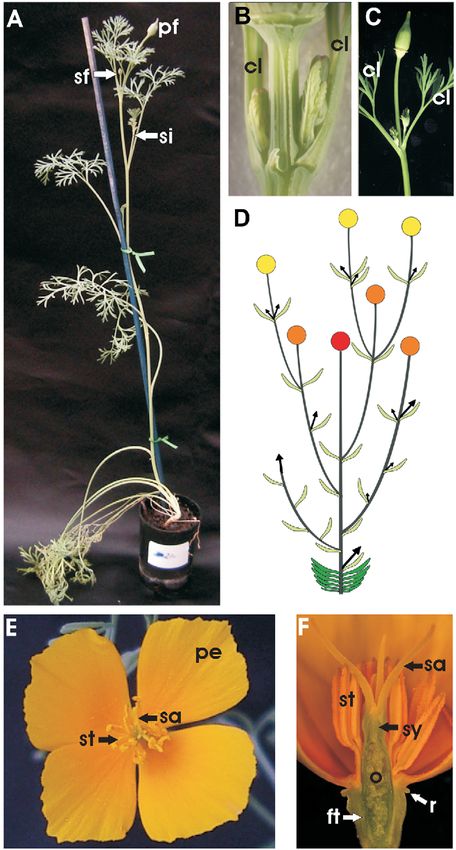
BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 541
are still being initiated, the outermost primordia are already
well formed (figs. 3H, 4A). Sometimes naturally occurring
variability can be observed in stamen number (see above).
This is characterized by the lack of regular stamen formation,
especially in the inner whorls, and results in disruption of the
usual twofold axis of symmetry (fig. 3I). At the end of stage
4, the bud has reached a lateral diameter of ca. 370 mm.
During stage 5, considerable changes occur in the mor-
phology of the central part of the developing bud. In the be-
ginning, the floral meristem resolves into a small dome
separate from the stamen primordia, from which the gynoe-
cium develops (figs. 3H, 4A). Stamen primordia are still
being generated outside this dome. Once the gynoecial pri-
mordium starts elongating with vertical edges, it also devel-
ops an indentation in its center (figs. 3J, 4B).
During stage 5, the perianth organs are developing. The
fused sepals, enclosing the bud fully from the end of stage 4,
elongate lengthwise, leaving a hollow cavity up to 550 mm
high above the gynoecium and stamen primordia. The sepals
do not fuse completely at the tip of the bud, leaving a narrow,
open slit (fig. 4B). No significant longitudinal growth is de-
tectable in the petals during stage 5 (fig. 4A, 4B).
At the end of stage 5, the bud’s lateral diameter is ca. 650
Fig. 2 Leaf heteroblasty and change in size of the shoot apex. A, mm. The sepals have grown to a thickness of ca. 40–70 mm
Heteroblasty of the 19 leaves of a typical primary shoot between (fig. 4B). The oval stamen primordia of the first stamen
cotyledons (left) and the terminal flower (right). Primary leaves, but whorl are ca. 60 mm wide, while the primordia of the inner-
also those preceding the terminal flower, are smaller and less complex. most whorl of stamens have just initiated (fig. 3J).
Open diamonds, number of primary lateral segments (pinnae) of one The beginning of stage 6 is characterized by the appear-
leaf half; filled diamonds, total number of segments on one leaf half ance of microsporangia in the outermost stamen primordia.
including leaf tip; solid line, leaf size in centimeters. B, Examples of The anthers adopt a concave profile (viewed from the out-
leaves, showing the range of complexity within a heteroblastic series. side), corresponding with the appearance of locules and locu-
The large climax leaf is much more complex than the first three foliage
lar ridges (fig. 3K; fig. 4C, 4D). By the end of stage 6, the
leaves shown from right to left. C, Diameter (left) and height (right) of
the shoot apical meristem (SAM) during vegetative development up to
stamens in all whorls have developed locules (fig. 4D). The
flowering. Measurements are, from left to right, primary shoots in stamens grow laterally, and those in the outer whorls are lon-
germinating seeds (G), SAMs 10 (10d) and 30 d (30d) after ger- ger than the inner ones. The stamens also become constricted
mination, lateral inflorescence meristems (IFM), and stage 2 flowers at their base (fig. 3L). The gynoecium continues to grow ver-
(FM2). Individual values (diamonds) are connected by lines represent- tically but is shorter than the outer stamens. Ovules have not
ing the observed range. yet initiated, but the two placental regions are bulging in-
ward, resulting in a gynoecium that is shaped like a hollow
tube with a very narrow center (fig. 4C, 4D).
in size laterally, leading to an elliptical shape (fig. 3F). By The bud has now grown to a diameter of ca. 1.25 mm. The
mid–stage 3 it is ca. 200 mm in lateral diameter, ca. 150 mm lengthwise growth of the gynoecium and stamens begins to fill
in medial diameter, and ca. 120 mm in height. At first, the se- the cavity formed by sepal extension. In comparison, the pet-
pal primordial ring is relatively shallow at the positions where als do not show a significant increase in length (fig. 4C).
the lateral petals arise, indicating that there are two sepals.
The sepal primordia are ca. 30 mm thick, and they grow rap-
Later Stages of Flower Development to
idly and evenly around the floral apex, forming a tube. They
Anthesis (Stages 7–11)
then curve inward to cover the floral apex in a bowl-shaped
manner, leaving a small medial opening (not shown). Stage 7 begins with the initiation of two files of ovule pri-
Very shortly after the initiation of the petal primordia, mordia as small protrusions from each of the two placental
stage 4 begins when four stamen primordia initiate in posi- regions within the gynoecium (fig. 4E). At this stage, the gy-
tions between and internal to the petal primordia (fig. 3G). noecium increases in lateral diameter to ca. 0.5 mm (from
Subsequently, up to four additional whorls of stamens, each ca. 350 mm in stage 6) to harbor the emerging ovule primor-
usually containing six stamen primordia, arise centripetally. dia. The connectives of the stamens begin to show vascula-
In the second stamen whorl, single primordia are formed op- ture (fig. 4E). In the outer stamen whorls, the locular tissue
posite the two lateral petal primordia, whereas paired stamen differentiates into compact sporogenous tissue in the center
primordia fill the gap opposite each of the two wider medial and a surrounding single layer of tapetal cells. In the inner-
petal primordia (fig. 3H). Next, six stamen primordia arise whorl stamens, sporogenous cells have not yet developed. Be-
in whorl 3 in positions alternate with those in the second sta- cause of the start of pedicel elongation, the bud is no longer
men whorl (fig. 3H). While the innermost stamen primordia constricted to the space between the cauline leaf and pedicel542 INTERNATIONAL JOURNAL OF PLANT SCIENCES
Table 1
Landmarks of Floral Development (according to Buzgo et al. 2004) of Eschscholzia californica
That Divide Floral and Fruit Development into 14 Stages
Observed range of bud Method of
Stage, developmental landmark Additional developmental events diameters during stage observation
1, Inflorescence meristem forms Inflorescence meristem emerges; grooves 80–120 mm SEM
separate the floral meristem from two
cauline leaf primordia; secondary
inflorescence meristems may appear
2, Sepal primordium emerges Sepal primordium encircles the floral 130–190 mm SEM
primordium
3, Petal primordia emerge Lateral petal primordia emerge; medial 170–240 mm SEM
petal primordia emerge
4, First whorl of stamens initiates Four stamen primordia initiate in the first 250–370 mm SEM, sections
stamen whorl; six stamen primordia
initiate in the subsequent 3–4 whorls
5, Carpel initiation Continuous stamen initiation; gynoecium 390–650 mm SEM, sections
becomes indented
6, Microsporangia initiate Stamens stalked at their base; locular 1–1.25 mm SEM, sections
ridges
7, Ovule initiation Male sporogenous tissue forms 1.65–2.25 mm Sections
8, Male meiosis Ovary valves develop ridges; first (inner) 2.3–2.8 mm Sections
ovule integument initiates; second
(outer) ovule integument initiates;
pollen grains mature; megaspore
mother cell differentiates
9, Female meiosis Embryo sac development occurs 3.5–5.5 mm Sections, macroscopic
(may continue into stage 10)
10, Sepal cap abscises Flower opens; stamens extend above Macroscopic
stigma
11, Anthesis Pollen shed; gynoecium becomes Macroscopic
receptive; stigma extends above
anthers; petals and stamens wither
12, Petals and stamens abscise Capsule elongates Macroscopic
13, Capsule fully extended Capsule dries out Macroscopic
14, Valves separate from dry capsule
and seeds fall Macroscopic
of the primary flower, and its originally oval shape becomes ameter and consist of densely packed small cells (fig. 5A). The
more rounded (fig. 4E, 4G). At the end of stage 7, the bud is gynoecium continues its medial and lateral expansion and is
ca. 2.0 mm in diameter. now ca. 0.8 mm wide. As this stage progresses, tetrads appear
Stage 8 is characterized by the commencement of meiosis in all stamen whorls (fig. 4F). In the outer-whorl stamens the
in the outer-whorl stamens. This occurs relatively rapidly, and tapetum layer begins to disintegrate. It becomes apparent now
tetrads of microspores soon appear (fig. 4F). Tapetal tissue that the inner-whorl anthers will not grow in diameter to
and sporogenous cells have also differentiated in the inner match those in the outer whorl but will remain at about one-
stamens by now. The anthers elongate substantially during third of their size (fig. 4E). The petals have enlarged laterally
this stage, whereas the filaments shows no significant length- and show several differentiated vascular bundles.
wise growth. At the beginning of stage 8, the gynoecium is Later in this stage, the second (outer) ovule integument ini-
ca. 0.5 mm wide. Each carpel develops five longitudinal tiates. On the adaxial side of the ovule, the second integu-
ridges, three medial and two lateral, resulting in a 10-pointed ment elongates and will eventually cover the entire nucellus.
star when the gynoecium is viewed in cross section (fig. 4F). On the abaxial side, the second integument initiates as a small
The gynoecium is significantly shorter than even the inner- protrusion (about two or three cells thick) but then fuses to
most stamens, and its base is surrounded by a floral tube (fig. the funiculus, which has elongated to approximately the
4G). Petals and stamens are attached to this floral tube, same length as the nucellus, including integuments. Associ-
which encloses about one-quarter of the gynoecium (fig. 4H). ated with the unequal growth of the second integument, the
Inside the ovary, the ovule primordia elongate and bend side- ovules now appear anatropous and curve back to the pla-
ways into the lateral part of the locule of the ovary (fig. 4F). centa (fig. 5B). The gynoecium has now grown to a lateral
Next, the first (inner) ovule integuments initiate as small diameter of ca. 1.2 mm, and a large cavity has appeared
bulges approximately between the middle and the top third of within the ovary, of which only a small portion is occupied
the ovule primordium. The ovules increase in length and di- by the ovules at this stage.BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 543
The outer-whorl anthers now carry pollen grains that ap- shortly after the stomia open (protandry), with pollen stick-
pear to be approaching maturity, and the tapetum layers ing to its stigmatic tissue. After the pollen grains have at-
have completely disintegrated (fig. 5E). In the inner-whorl tached to the stigmatic papillae, the pollen tubes start
stamens, the tapetum is visible, and most pollen grains are growing down the stigma toward the ovary (fig. 6F–6H).
still immature. The filaments of the innermost stamen whorls They choose a path along the inside of the solid style wall at
are now longer than those in the outer whorls, so that their the base of the papilla-like cells (fig. 6G). Once they have
shorter anthers are elevated to the same total height. The pet- reached the ovary, the pollen tubes are apparently directed
als have extended longitudinally and are about one-third of to grow down inside the two placentae before fertilizing the
the length of the outer-whorl stamens (fig. 4H). ovules (fig. 6H).
Subsequently, the ovules differentiate a long and narrow
megaspore mother cell. This is embedded in five to eight cell
Surface Morphology of the Mature Floral Organs
layers of nucellar tissue (i.e., the ovules are crassinucellate)
and is enclosed in callose deposits toward its base. Also, the To document the surface structure of mature floral organs,
two integuments are now fully developed: the inner integu- SEM was used. Both sepal surfaces are composed of large, ir-
ment consists of three cell layers, and the outer integument regularly shaped cells that can extend up to 50 mm in length
can grow up to five layers thick, with an even larger number and more than 30 mm in width. Many stomata are inter-
of cell layers toward the micropyle. The funiculus shows vas- spersed between these cells on the outer surface (fig. 7A).
culature and does not further increase in length or diameter Both surfaces of the petals are covered with highly elongated
(fig. 5C). In the ovary, lignification occurs in the walls of (ca. 75 mm long), narrow (ca. 5 mm wide) cells. They are ar-
a single line of cells that extends from the center of the pla- ranged in long, regular rows. Thick deposits likely to be cel-
centa to the outside of the ovary wall (fig. 6A, 6B). This indi- lulose create a ridge along the whole length of the cell. This
cates the site of dehiscence of the two valves that occurs ridge even continues over the plane of contact between two
much later in the mature fruit. The lateral diameter of a bud adjacent cells (fig. 7B). The cells on the abaxial side of the
at the end of stage 8 ranges from ca. 2.3 to 2.8 mm. petals have less pronounced ridges (not shown). Petals do not
From the commencement of stage 9, the megaspore mother have stomata. The surface of the stamen filament consists of
cell undergoes meiosis. One resulting megaspore develops cells with approximately the same width as the petal cells,
into the embryo sac, which contains the egg apparatus (con- but they are considerably shorter (a maximum of 30 mm
sisting of the egg cell and synergids), central cell, and anti- long) and lack pronounced ridges (fig. 7C). The cells of the
podals (fig. 5D). Substantial amounts of callose are deposited anther surface are comparably large, up to 60 mm long and
in the cells surrounding the megaspore mother cell within the up to 20 mm wide. Stomata were observed on the surface of
nucellus before megasporogenesis begins, and these callose the filament but not on the anther (fig. 7C, 7D). Stigmatic
deposits are maintained until anthesis (fig. 5C, 5D). The papillae densely cover the whole length of the dry stigma.
ovules do not fill all available space in the ovary, as spaces They are club shaped, with the rounded end (ca. 15 mm in
are apparent even close to anthesis (fig. 6A). diameter) bulging outward (fig. 7E). The surface of the ovary
The pollen grains in all anthers have matured in stage 9, wall is composed of small, irregularly shaped cells with in-
and the tapetum has disintegrated, as has the cell layer sepa- terspersed stomata (fig. 7F). Two distinct cell surface types
rating the two locules in each theca from each other. Every occur: cells with a smooth, bulging surface that contribute to
pollen sac has developed a thickened endodermis and a sto- the regions between the ovary ridges and cells with a flat,
mium, through which pollen is released later at anthesis (fig. ribbed surface that form the ridge’s surface. Deposits of
5F, 5H). waxy granules can be observed as white dots (fig. 7F). Tri-
Stage 10 begins with the abscission of the sepals as a single chomes are not present on any mature floral organ of Cali-
cap, pushed off by the elongating petals. The bud size at the fornia poppy.
beginning of this stage varies considerably, falling between
ca. 3.5 and 5.5 mm (average 6 standard error ¼ 4:37 6 0:11
Postfertilization Development and Fruit
mm). Subsequently, the flower opens, and the petals, wrin-
Maturation (Stages 12–14)
kled at first, straighten. The stamens extend above the
stigma. Pedicel elongation stops during stage 10, and final During late stage 11, the petals and stamens start to
pedicel lengths were observed to range between 4 and 8.25 wither, and at the beginning of stage 12, they abscise from
cm, possibly associated with the position of the flower on the the floral tube. In most cases, one petal and a few stamens
plant and/or plant growth conditions. positioned abaxially of it remain attached and abscise to-
Stage 11 is characterized by anthesis. The anthers dehisce gether. The gynoecium has grown to a length of 8:25 6 0:43
simultaneously through long slits (stomia) at their abaxial mm, on average, at the beginning of stage 12, and it contin-
side, ca. 1 d after flower opening (fig. 5H), and release bright ues its rapid elongation throughout this stage. Prominent
yellow pollen grains. The staining properties of the endothe- ridges run the length of each capsule, three in medial regions
cium layer in the anticlinal walls of the anthers (fig. 4F) indi- of each valve (fig. 6C) and two in lateral regions (fig. 6D).
cate that these thickened cell walls contain cellulose rather Stage 13 commences once the elongation of the capsule is
than lignin. However, the endothecium cell walls oriented to- complete. Mature capsules ranged in final length between 3.1
ward the anther center contain lignin as well as cellulose. and 6.6 cm, with an average of 5:0 6 0:2 cm. The capsules
The gynoecium continues growing, and soon the style ex- then dry out while the seeds inside mature. Parallel to the
tends above the anthers. The gynoecium becomes receptive drying process of the capsule, the pedicel dries out as well.BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 545
Stage 14 is marked by the explosive release of the seeds. flowers are open for 3:16 6 0:17 d, on average (range 3–4 d),
This occurs when the now dry valves separate basipetally until the petals and stamens abscise (stage 12), although
from each other. During this separation process, the bottom these organs start withering ca. 2 d after anthesis. Following
parts of the two valves bend toward their abaxial side, re- petal and stamen loss, the capsule grows longitudinally for
maining connected at the style. In each case, the valve also 5:8 6 0:17 d (range 5–7 d), on average, until the final length
usually separates from the two lateral placental regions along is reached (stage 13).
each of its edges. Thus, when both valves have dehisced, four
placental remnants remain, partially separated and partially
Expression of EcFLO during the Life Cycle
attached to the valves. These replum-like frames may also re-
of Eschscholzia
main attached to the style but not to the receptacle, so that
the capsule abscises as a whole (see fig. 6E). Expression of EcFLO in apical meristems was monitored
from before germination to the development of axillary in-
florescences and flowers (fig. 8). Expression was first detected
Duration of Some Flower Development Progressions
in imbibed seeds that usually germinated after 5 d. After
Individual poppy plants varied in respect to luxuriance, breaking the seed coat, the root pole grows outward, being
flowering time, and flower numbers at any given time. Even pushed by the elongating cotyledons that remain within the
so, some time course data could be collected once buds were seed coat at this stage (fig. 8A, 8D). EcFLO expression oc-
sufficiently large, so that measurements of the diameter of in- curs in a ringlike zone around the central zone of the seedling
dividual buds could be followed over time. The stage reached SAM at various germination stages (fig. 8B, 8C).
by buds of a specific diameter was estimated from sections of During development of the rosette, the position of the ex-
equivalent buds. This was relatively constant at first, but, by pression domain in the flank meristem remains the same but
the end of stage 8, variability between buds meant that the the domain becomes larger with the overall increase of SAM
diameter was not a good predictor of bud stage (table 1). size. As previously reported (Busch and Gleissberg 2003),
The diameter of 10 buds was measured each day, starting leaf initiation is associated with expression gaps correspond-
from 1.25 mm (micorsporangia initiation, stage 6) until they ing to the center of prospective and emerging leaf primordia
reached 2.5 mm, at which size male meiosis occurs (stage 8). (fig. 8E). Initiation of axillary shoots is also marked by
These buds needed 6.95 d, on average (60.39 d standard er- downregulation in the prospective central zone of the axillary
ror, range 6–8 d), to pass through these stages. meristem (not shown; see Busch and Gleissberg 2003).
Four buds were measured starting from a 1.5-mm diameter Lateral inflorescence meristems arising from the axils of
(in stage 6) until sepal abscission (stage 10). They needed an the uppermost cauline leaves show the same expression pat-
average of 11:37 6 1:49 d (range 9.5–13 d) to pass from tern restricted to the flank meristem before (fig. 8F) and dur-
stage 6 to stage 10. These four buds showed pronounced dif- ing sequential initiation of the cauline leaf pairs (fig. 8G–8I).
ferences in growth rate, especially apparent after commence- As in rosette leaf primordia, expression is discontinued in the
ment of male meiosis in stage 8, resulting in bud diameter center of arising cauline leaf primordia (fig. 8H, 8I).
before flower opening ranging from 3.2 to 5.2 mm. Interest- After transition to floral meristem identity, the pedicel
ingly, the steady increase in bud diameter was usually ob- starts to develop on the enlarging floral apex that has initi-
served to slow ca. 2 d before sepal abscission. ated the sepal ring primordium (stage 2; fig. 8J). EcFLO
After sepal abscission (stage 10), the flower opens, and the transcripts extend into the periphery of the sepal ring. Above
petals extend fully. After that, 0:9 6 0:1 d (range 0–2 d) pass the sepal primordia, expression is seen in a clearly defined
until anthesis begins (stage 11). Following abscission, the peripheral ring excluding a broad central area, similar to
Fig. 3 Scanning electron micrographs of early stages of floral development. A, Vertical view of two primary inflorescence meristems (im) in
early stage 1. These are developing from the uppermost cauline leaves below the terminal primary flower, which has been removed from the center.
Bar ¼ 75 mm. B, C, Vertical view of buds in late stage 1. Cauline leaf primordia (lp) and the floral meristem (fm) are indicated. In C, an axillary
inflorescence meristem (im) has initiated from the axil of the lower cauline leaf (lp). Bar ¼ 75 mm. D, Lateral view of a bud at stage 2. The dome of
the floral meristem (fm) is now encircled by the sepal primordia (sp). Bar ¼ 100 mm. E, Vertical view of two secondary inflorescences that have
reached early stage 2 (left side) and late stage 2 (right side). The primary flower has been removed. Bar ¼ 100 mm. F, Vertical view of a late stage 3
bud, showing petal primordia (pp) already separated from the remaining floral meristem (fm) and showing that the sepal primordia (sp) are
starting to overgrow the floral dome. The axis connecting the two lateral petal primordia is displaced by an angle of ca. 30° in relation to the
cauline leaf primordium (lp). Bar ¼ 100 mm. G, Vertical view of a bud in stage 4. The sepals enclosing the bud at this stage have been removed.
Four stamen primordia arise in the first stamen whorl (1). More stamen whorls are being initiated around the floral dome (fd), and there is not yet
a distinct gynoecium primordium. Lateral and medial petal primordia (lpp and mpp, respectively) are shown. Bar ¼ 100 mm. H, Bud at the
beginning of stage 5 in which the gynoecium (g) is separated from the region producing stamen primordia. Stamen whorls are numbered from the
oldest (1) to the youngest (3 in H and I, 4 in J). Bar ¼ 100 mm. I, View of a bud between stage 4 and 5 showing irregular stamen initiation. In
stamen whorls 1 and 2, the stamens have initiated regularly, but those in whorl 3 and the following whorls grow in a disordered fashion.
Bar ¼ 100 mm. J, Vertical view of a late stage 5 bud showing the indented gynoecium (g) separated by a deep groove from the stamen primordia.
The latter do not yet show locular ridges on their abaxial surfaces. Bar ¼ 120 mm. K, Vertical view of a bud in stage 6 showing clearly visible
locular ridges (arrows) in the outer stamens. The gynoecium has begun growing vertically as a tube. Bar ¼ 150 mm. L, Lateral view of the bud in
K, showing triangular petals (p) and stamen primordia that are constricted at the base (arrows). The height of the stamen primordia is in
proportion to when they arose. Bar ¼ 150 mm.BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 547
vegetative SAMs. Expression also extends downward around were observed in SAM size, floral meristem size, bud diame-
the periphery of the elongating pedicel. At a slightly later ter at various floral stages, and stamen number in plants
stage, before extension of sepal primordia to form a cap, Ec- grown under the same conditions (fig. 2C; table 1; Cook
FLO expression corresponds to a domain from which petals 1962). Vigorous plants are more procumbent, with only the
and stamens start to initiate. This ring-shaped area is slightly pedicel elongated, and they have several secondary inflores-
elevated, and it is separated from the gynoecial initiation cences in more basal positions (Günther 1975). Much of the
field in the center by a smooth sinus (figs. 4G, 8K). Isolated flexibility in developmental program seems to be intimately as-
gaps within the peripheral expression domain appear to cor- sociated with environmental factors, but some growth habit
respond to initiating stamens, as seen in tangential sections differences are genetically determined, including annual/
(fig. 8L). Up to this point, weak EcFLO expression is main- perennial and erect/procumbent growth habits (Beatty 1936;
tained in the periphery of the sepals and pedicel. Günther 1975).
During further initiation of stamens and the beginning of The observed phenotypic variation could be associated
gynoecium elevation and outgrowth (stage 5), EcFLO tran- with the strong genetic self-incompatibility (SI) system pres-
script accumulation retreats to smaller areas at the base of ent in California poppy (Beatty 1936). This is a gametophytic
petal primordia, near the base, and around the insertion of system, and a single genetic factor (controlled by the S-locus)
stamen primordia (fig. 8N, 8O). Expression in the receptacle apparently acts in the stigma and inhibits pollen tube growth
between developing sepals, petals, stamens, and the gynoe- soon after pollen germination. Outbreeding systems like this
cium is still visible in stage 6 flower buds. No EcFLO expres- can lead to high levels of heterozygosity in natural pop-
sion could be observed in gynoecium tissue at any stage of ulations (Wright 1979). Even though some Eschscholzia
flower development. populations, especially naturalized ones, are somewhat self-
compatible, the extent of inbreeding is still very low. In rare
Discussion cases of self-compatibility, the resulting seed set and viability
of the F1 generation was low, and the few F1 plants that
Our observations on the development of Eschscholzia cali- flowered were sterile (Beatty 1936). It will be of interest to
fornica are to a large extent consistent with previous, less de- determine if the SI system in Eschscholzia involves the same
tailed work on California poppy flower development (Ronse mechanism of action, programmed cell death of incompatible
Decraene and Smets 1990; Karrer 1991). In addition, we pollen, as occurs in the related poppy Papaver rhoeas
studied the expression pattern of EcFLO, the Eschscholzia (Thomas and Franklin-Tong 2004).
ortholog of the Arabidopsis flower meristem identity gene LFY,
to understand its role in conferring inflorescence and flower
Comparison of Flower Development in Californian
meristem identity and floral organ properties in the poppy.
Poppy and Other Poppy Species
The poppy family comprises 23 genera with ca. 240 species,
Californian Poppy Morphology Shows a
which are distributed mainly in the Northern Hemisphere of
High Degree of Plasticity
the Old and New World. They have actinomorphic, bisexual
Extensive variation in growth habit, floral structure, and flowers. Usually, there are two or three free sepals, which ab-
plant longevity can be observed in Eschscholzia specimens scise at or shortly after flower opening. Typically, Papavera-
collected in a single population and within populations ceae species have two whorls of two or three petals, which
(Beatty 1936; Cook 1962). In this study, plants grown in are crumpled in the bud. Numerous free stamens arise cen-
constant light in the glasshouse showed elongated internodes tripetally in whorls, with basifixed anthers containing four
and very few basal branches (fig. 1A), in comparison to sporangia. The ovary is syncarpous and made up of two to
specimens grown in spring in the field. Large differences 20 carpels, with many ovules arising from parietal placentas.
Fig. 4 Sections of California poppy buds stained with safranin and fast green. A, Longitudinal section of a bud in stage 4, showing the dome-
shaped floral meristem (fm), stamen primordia (sp), petal primordia (pp), and elongating sepal cap (se). Secondary inflorescence meristems (si)
have formed in the axils of the cauline leaves of the primary bud. Bar ¼ 200 mm. B, Longitudinal section of a stage 5 bud, showing the indented
gynoecium primordium (gp) and the petal (pp) and stamen (sp) primordia. Bar ¼ 200 mm. C, Longitudinal section of a bud in stage 6, showing the
elongating tubelike gynoecium in the center. Locular regions in the stamens surrounding the gynoecium are darker in color. Bars in C–H ¼
500 mm. D, Transverse section of a stage 6 bud, showing the narrow gynoecium (g) in the center with two placentae (pl) protruding inward and
the four locules containing microsporangia in each stamen (st). The petals are not visible because of the position of the cross section. E, Transverse
section of a bud in stage 7. Ovule primordia (o) are initiated at this stage and are visible as small protrusions extending from the placenta (pl). In
the anthers, the vasculature (v) of the connectives is indicated, as well as the sporogenous tissue (st) in the locules. F, Transverse section of a bud in
stage 8, which commences when male meiosis begins. The valves of the gynoecium show 10 protruding ridges (ri). The inset shows an enlargement
of an outer-whorl anther. Tetrads of microspores (te) appear as an agglomeration of light blue–stained cells, and the tapetum (ta) is visible
surrounding the tetrads. G, Longitudinal section of a bud in stage 7. Within the gynoecium, part of the placenta (pl) with initiating ovules (o) is
visible. The gynoecium is surrounded at its base by the floral tube (ft), which is encircled by a rim (r) on its outside. Petals (pe) and stamens are
attached to the top of the floral tube. Stamens consist of a short filament (f ) and an anther (a) that contains densely packed sporogenous cells
surrounded by the dark blue–stained tapetum. H, Longitudinal section of a late stage 8 bud. Within the anthers (a), the tapetum layer disintegrates
and pollen grains (pg) mature. The style of the gynoecium (st) elongates, and the ovary harbors ovules (o), now consisting of two integuments and
a nucellus with a megaspore mother cell in the center. The floral tube (ft), expanding petals (pe), and outer rim (r) are also indicated.548 INTERNATIONAL JOURNAL OF PLANT SCIENCES Fig. 5 Ovule and stamen development. A, Transverse section showing ovules protruding from the placenta (pl) of a bud late in stage 8, when the first (inner) integument (ii) arises. B, Transverse section of two ovules from a bud late in stage 8, where the second integument has already initiated and the ovules adopt their anatropous shape. The nucellus (nu) is embedded in the inner (ii) and outer (oi) integuments. The ovule is connected to the placenta (pl) by the funiculus (fu). C, Transverse section of an ovule from a bud in stage 8. The long and narrow megaspore mother cell (mmc) is surrounded by callose deposits (cd, stained pink). Inner (ii) and outer (oi) integuments embed the nucellus and are open at the micropylar end (mp). D, Transverse section of an ovule from a bud in stage 9, after megasporogenesis (female meiosis) has occurred. The egg apparatus (ea) and antipodal cells (ac) are part of the embryo sac, which is still surrounded by callose deposits (cd). E, Transverse section of anthers from a bud late in stage 8, when the first immature pollen grains (pg) appear. The vasculature (v) in the connective, petals (pe), and sepals (se) are indicated. F, Transverse section of an anther from a bud at the end of stage 8. Mature pollen grains (pg) are stained dark blue. The endothelium (e), stomium (st), and connective (c) are indicated. G, Scanning electron micrograph showing the lower part of mature stamens before anthesis. The bases of the filaments (f ) are attached to the petal (pe) as indicated by an arrow, and they will dehisce together at anthesis. The connective (c) is situated between the two thecae (th). Bar ¼ 75 mm. H, Scanning electron micrograph showing the top part of an anther at anthesis (stage 11). The anthers open through the long, slit-shaped stomium (st) to release pollen grains. The connective (c) between the two thecae (th) is also indicated. Bar ¼ 0:5 mm. The stigma is often sessile and merges with the style, and two-carpel theory, the gynoecium consists of two carpels, poppy fruits normally dehisce as dry capsules (Kadereit 1993). with the placental regions at their points of fusion (Karrer In most aspects of flower morphology, California poppy is 1991; Brückner 2000). However, a four-carpel theory has a typical representative of the Papaveraceae (Hoot et al. been proposed to account for the presence of four stigmatic 1997). However, its fused sepals are typical only for members lobes. This assumes the presence of two types of carpels of the genus Eschscholzia and for Eomecon, a genus in sub- within two-valved gynoecium, one type being fertile and ex- family Chelidonioideae. Most poppy species have hypogy- panding, the other fertile and contracting (or, according to nous flowers, and the presence of a floral tube (hypanthium) older ideas, two fertile and two sterile carpels) in two whorls. is specific to the genus Eschscholzia. In addition, a conspicu- However, our results and the work of Karrer (1991) have ous torus rim developing below the sepal ring before anthesis shown that the gynoecium initiates as a single hemispherical is characteristic of E. californica (Ernst 1962). The California structure and that additional carpel whorls were never ob- poppy gynoecium consists of two fused carpels with entirely served. Also, the stigmatic lobes above the placentas develop free carpel tips (free carpel tips are also found in the genus only late in floral development (during or after the beginning Platystemon and allies in subfamily Papaveroideae). Two of stage 8), are extremely variable in length, and in some long stylar protrusions occur above the valves, and two more cases do not develop at all. are usually present above the placentae in all members of the Nectar is absent in all Papaveraceae, but pollen is provided Eschscholzioideae (Kadereit 1993). According to the classical in abundance as a pollination reward. California poppy is
BECKER ET AL.—CALIFORNIA POPPY DEVELOPMENT 549 Fig. 6 Late stages of gynoecium and fruit development. A, Transverse section through a gynoecium just before female meiosis begins in stage 9. The gynoecium, containing the ovules (o), is surrounded by the floral tube (ft). Each of the two valves includes three median (mr) and two lateral ridges (lr) within the ovary wall. The replum region is situated between the two neighboring lateral ridges. The black arrows indicate six vascular bundles in the ovary wall, and the white arrows show the file of lignified cells in the valve/replum border region. B, Enlargement of the valve/ replum border in the ovary wall (ow) of a bud in stage 9. Lignified cells with pink-stained cell walls (white arrows) are arranged transversely from the replum to the placenta (pl). Later in capsule development, the dehiscence zone between the two rows of seeds attached to the placenta will form in this position. The placenta is lined with papillae-like cells (pc). C, View of the valve region of a mature California poppy capsule. The median ridges (mr) are indicated. D, View of the valve/replum border of a mature capsule. Lateral ridges (lr) and the zone of the valve/replum (v/r) border are indicated. E, Dry silique with valves (v) separated from the replum-like frame (r). Californian poppy siliques open explosively from bottom to top, and the two valves remain attached below the style (st). Replum material also dehisces from the receptacle and usually remains attached to the valves. The coin is 19 mm in diameter. F, Longitudinal section of the stigma after pollination, stained with aniline blue. Callose appears brightly fluorescent. Pollen tubes (pt) are visible, growing internal to the papillae cells (pc) that cover the surface of the stigma. Callose is also present in the sieve plates of the phloem (ph). The cell walls of the xylem (xy) are also weakly fluorescent. G, Transverse section of the style region after pollination, stained with aniline blue. The pollen tubes (pt) are visible as small circles internal to the papillae cells (pc) that line the hollow internal space of the style. The xylem vessels (xy) are also fluorescent. H, Longitudinal section of the base of the style and top part of the ovary (o). Pollen tubes (pt) grow down the ovary at the inside of the papillae-like tissue (pc) lining the placenta. The ovary wall (ow) and style wall (sw) are also indicated. pollinated by a variety of insects, the most significant being to the two valves, and seeds may be scattered in a radius of honeybees, bumblebees, beetles, and some other insect spe- 1.5 m by this action (Cook 1962). The fruits of the Papavera- cies (Cook 1962). ceae are very variable; some dehisce basipetally or laterally, California poppy capsules dehisce explosively in an acrop- however, and some more derived poppy species, such as Pa- etal direction, leaving the replum-like frames partly attached paver somniferum (opium poppy), have indehiscent capsules
You can also read

















































