JBC Papers in Press. Published on January 28, 2020 as Manuscript RA119.011440
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JBC Papers in Press. Published on January 28, 2020 as Manuscript RA119.011440
The latest version is at https://www.jbc.org/cgi/doi/10.1074/jbc.RA119.011440
Antisense oligonucleotides targeting Notch2 ameliorate the osteopenic phenotype in a mouse model of
Hajdu Cheney syndrome
Ernesto Canalis1,2,3*, Tamar R. Grossman4, Michele Carrer4, Lauren Schilling3 and Jungeun Yu1,3
Departments of Orthopaedic Surgery1 and Medicine2, and the UConn Musculoskeletal Institute3,
UConn Health, Farmington, CT, 06030 and Ionis Pharmaceuticals, Inc.4, Carlsbad, CA 92010
Running title: ASOs and Hajdu Cheney
*To whom correspondence should be addressed: Ernesto Canalis, M.D.: Departments of Orthopaedic
Surgery and Medicine, UConn Health, 263 Farmington Avenue, Farmington, CT 06030-4037;
Email: canalis@uchc.edu; Telephone: (860) 679-7978; Fax: (860) 679-1474
Keywords: Antisense oligonucleotides; Hajdu Cheney Syndrome; Notch receptor; bone; bone
resorption; osteoclast; osteoclastogenesis; cell fate; cell signaling; osteoporosis
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
ABSTRACT mice. Bone marrow–derived macrophage
Notch receptors play critical roles in cell cultures from the Notch2tm1.1Ecan mice displayed
fate decisions and in the regulation of skeletal enhanced osteoclastogenesis, which was
development and bone remodeling. Gain-of- suppressed by Notch2 ASOs. In conclusion,
function NOTCH2 mutations can cause Hajdu Notch2tm1.1Ecan mice exhibit cancellous bone
Cheney syndrome (HCS), an untreatable disease osteopenia that can be ameliorated by systemic
characterized by osteoporosis and fractures, administration of Notch2 ASOs.
craniofacial developmental abnormalities, and
acro-osteolysis. We have previously created a
mouse model harboring a point 6955C>T Notch receptors are four single-pass
mutation in the Notch2 locus upstream of the transmembrane proteins that play a critical
PEST domain and termed this model function in cell fate determination (1,2). Notch1,
Notch2tm1.1Ecan. Heterozygous Notch2tm1.1Ecan 2 and 3 and low levels of Notch4 transcripts are
mutant mice exhibit severe cancellous and detected in bone cells, where they play a key role
cortical bone osteopenia due to increased bone in osteoblast and osteoclast differentiation and
resorption. In the present work, we demonstrate function (3). Notch receptors are activated
that the subcutaneous administration of Notch2 following interactions with ligands of the Jagged
antisense oligonucleotides (ASO) down- and Delta-like families, and JAGGED1 is the
regulates Notch2 and the Notch target genes prevalent ligand expressed by skeletal cells
Hes-related family BHLH transcription factor (3,4). Interactions of NOTCH with its ligands
with YRPW motif 1 (Hey1), Hey2, and HeyL in lead to the proteolytic cleavage of the NOTCH
skeletal tissue from Notch2tm1.1Ecan mice. Results protein and to the release of the NOTCH
of microcomputed tomography experiments intracellular domain (NICD) (5,6). The NICD is
indicated that the administration of Notch2 translocated into the nucleus where it forms a
ASOs ameliorates the cancellous osteopenia of complex with recombination signal-binding
Notch2tm1.1Ecan mice, and bone protein for Ig of κ (RBPJκ) and mastermind
histomorphometry analysis revealed decreased (MAML) to induce the transcription of target
osteoclast numbers in Notch2 ASO–treated genes including those encoding Hairy Enhancer
Notch2tm1.1Ecan mice. Notch2 ASOs decreased the of Split (HES)1, 5 and 7 and HES-related with
induction of mRNA levels of TNF superfamily YRPW motif (HEY)1, 2 and L (7-9).
member 11 (Tnfsf11, encoding the Although activation of NOTCH1, 2 and
osteoclastogenic protein RANKL) in cultured 3 in the skeleton results in osteopenia, the
osteoblasts and osteocytes from Notch2tm1.1Ecan mechanisms responsible for the bone loss are
1ASOs and Hajdu Cheney
distinct (10-14). NOTCH2 has unique opportunity to correct the mechanisms
properties and impairs osteoblast maturation and responsible for the disease.
induces osteoclastogenesis by acting directly on Approaches to downregulate Notch
cells of the myeloid lineage and by inducing signaling include the use of biochemical
receptor activator of nuclear factor κB ligand inhibitors of Notch activation, thapsigargin,
(RANKL) in cells of the osteoblast lineage antibodies to nicastrin, which forms part of the
(10,12,15). γ-secretase complex, or to Notch receptors or
Hajdu Cheney Syndrome (HCS) is a their ligands, and stapled peptides that prevent
dominant inherited disease characterized by the assembly of a NICD/RBPJκ/MAML ternary
craniofacial developmental abnormalities, complex (28-32). A limitation of these
acroosteolysis, generalized osteoporosis with approaches is that either they are not specific
fractures and neurological complications (16- inhibitors of Notch signaling or they prevent the
18). HCS is associated with point mutations in, indiscriminate activation of all Notch receptors,
or short deletions of, exon 34 of NOTCH2 that leading to a generalized Notch activation knock-
lead to the creation of a stop codon upstream of down and side effects. Anti-Notch NRR
the proline (P), glutamic acid (E), serine (S) and antibodies have been effective at preventing the
threonine (T) (PEST) domain (19-23). The activation of specific Notch receptors (33-35).
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
PEST domain is required for the ubiquitination However, the pronounced downregulation of
and degradation of NOTCH2. As a Notch activation may result in gastrointestinal
consequence, the mutations result in the toxicity.
translation of a stable truncated protein product Antisense oligonucleotides (ASOs) are
and a gain-of-NOTCH2 function. Iliac crest single-stranded synthetic nucleic acids that bind
bone biopsies obtained from subjects afflicted by target mRNA by Watson-Crick pairing resulting
HCS have demonstrated the presence of in mRNA degradation by RNase H (36,37). The
osteopenia, increased bone resorption and administration of ASOs has emerged as a novel
trabecularization of cortical bone (24-26). therapeutic approach to downregulate wild type
To gain an understanding of the HCS and mutant transcripts, and has been successful
skeletal phenotype and the mechanisms in the silencing of mutant genes in the central
involved, we introduced a Notch2 mutation and peripheral nervous system, retina and liver
(6955C>T) in the mouse genome to reproduce a (38-45). ASOs have been used to downregulate
mutation (6949C>T) found in a subject with specific genes in the skeleton, although
HCS (10,20,22). The mutation creates a stop information about their possible use as a
codon in exon 34 leading to the translation of a therapeutic intervention in genetic disorders of
truncated NOTCH2 protein of 2318 amino acids the skeleton is limited (46,47).
(10). The mouse line, termed Notch2tm1.1Ecan, The purpose of the present work was to
exhibits NOTCH2 gain-of-function and answer the question as to whether the phenotype
homozygous mice display craniofacial of the Notch2tm1.1Ecan mouse model could be
developmental abnormalities and newborn ameliorated or reversed by downregulating
lethality. Heterozygous Notch2tm1.1Ecan mice Notch2 expression with Notch2-specific ASOs.
have cancellous and cortical bone osteopenia To this end, heterozygous Notch2tm1.1Ecan and
due to enhanced bone resorption. This is control littermate mice were treated with second
secondary to an increase in the number of generation phosphorothioate modified ASOs
osteoclasts due to enhanced expression of targeting Notch2 and characterized by bone
RANKL by cells of the osteoblast lineage as well microarchitectural analysis. The direct effects of
as due to direct effects of NOTCH2 on the Notch2 ASO on osteoblast, osteocyte and
osteoclastogenesis (10,15). The discovery of the osteoclast cultures from control and
mechanisms responsible for the bone loss experimental mice also were tested.
provided clues to offer improved treatments to
individuals with HCS, such as the use of the Results
RANKL antibody denosumab (27). However, Effect of Notch2 ASOs on Notch2 Expression
none of the available interventions offers the and Signaling In Vivo
2ASOs and Hajdu Cheney
In initial experiments, we tested whether administration of mouse Notch2 ASOs, control
mouse Notch2 ASOs downregulated Notch2 and Notch2tm1.1Ecan experimental mice appeared
mRNA in vivo in tissues where Notch2 is healthy although a 6% decrease in weight was
expressed and is known to have a function noted in wild type mice treated with Notch2
(10,48-52). The subcutaneous administration of ASOs when compared to control ASOs.
ASOs targeting murine Notch2 to C57BL/6 wild Femoral length was not affected by Notch2
type mice at a dose of 50 mg/Kg caused a ~40 to ASOs in either control or Notch2tm1.1Ecan mice
50% downregulation of Notch2 mRNA 40 h later (Figure 3).
in the spleen, kidney and femur, and an 80% Validating previous observations, µCT
reduction of Notch2 transcripts in the liver of the distal femur revealed that 2 month old
(Figure 1). In a subsequent experiment, Notch2 Notch2tm1.1Ecan mutant male mice had a
ASOs, administered subcutaneously to wild type significant decrease in trabecular bone
C57BL/6 mice at 50 mg/Kg, downregulated volume/total volume (BV/TV) associated with
Notch2 mRNA in femur by ~40% 48 to 96 h after reduced connectivity and higher structure model
the administration of the ASO. index (SMI) (10). Trabecular number and
There was evidence of enhanced Notch thickness were both reduced in Notch2tm1.1Ecan
signaling in skeletal tissue from Notch2tm1.1Ecan mice, contributing to the decrease in BV/TV
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
mice, and the Notch target genes Hey1, Hey2 and (Figure 4). The subcutaneous administration of
HeyL were induced in bone extracts from mutant mouse Notch2 ASOs once a week at 50 mg/Kg
mice in relationship to control littermates for 4 weeks did not change microarchitectural
(Figure 2). The subcutaneous administration of parameters of femoral bone in wild type mice. In
mouse Notch2 ASOs decreased the expression contrast, Notch2tm1.1Ecan mice receiving Notch2
of Notch2 and Notch26955C>T mutant mRNA. ASOs had a BV/TV that was 30% greater than
Notch2 ASOs also decreased the Notch target in mutant mice receiving control ASOs. As a
genes Hey1 and Hey2 in bone extracts from wild consequence BV/TV in Notch2tm1.1Ecan mice was
type mice and Hey1, Hey2 and HeyL in extracts reduced by 28% when compared to control wild
from Notch2tm1.1Ecan mice demonstrating a type mice, whereas Notch2tm1.1Ecan treated with
suppressive effect of Notch2 ASOs on Notch control ASOs exhibited a 45% reduction in
signaling in the skeleton. As a result, the mRNA BV/TV compared to wild type littermate
levels of Hey1, Hey2 and HeyL in tibiae from controls (Figure 4). The partial restoration of
Notch2tm1.1Ecan mice treated with Notch2 ASOs BV/TV by Notch2 ASOs was associated with a
approached the levels found in tibiae from wild significant increase in trabecular number.
type mice treated with control ASOs. A modest Notch2tm1.1Ecan mice presented with cortical
induction of Tnfsf11 (encoding RANKL, p > osteopenia and cortical bone was thin and bone
0.05) was observed in tibiae from Notch2tm1.1Ecan area and cortical thickness were reduced (Table
mice and this was reduced by Notch2 ASOs. 1). The cortical osteopenia was not affected by
Notch2 ASOs, so that the cortical bone area and
Effect of Notch2 ASOs on General thickness in Notch2tm1.1Ecan mice treated with
Characteristics, Femoral Microarchitecture Notch2 ASOs were not different from values
and Histomorphometry of Notch2tm1.1Ecan Mice obtained in mutant mice treated with control
Heterozygous Notch2tm1.1Ecan mutant ASOs.
male mice were compared to wild type sex- Cancellous bone histomorphometric
matched littermate mice in a C57BL/6 genetic analysis revealed that osteoclast number was
background because the skeletal phenotype was increased in Notch2tm1.1Ecan mice; Notch2 ASOs
similar in both sexes, and the homozygous did not change osteoclast number in wild type
mutation of Notch2tm1.1Ecan results in perinatal mice, but significantly reduced osteoclast
lethality. Confirming prior results, number in Notch2tm1.1Ecan mice, so that osteoclast
Notch2tm1.1Ecan heterozygous mice had ~10% number was not different between Notch2tm1.1Ecan
lesser weight than littermate controls, and their mice treated with Notch2 ASOs and control
femoral length was slightly shorter than that of littermate wild type mice (Table 2). Confirming
controls (Figure 3) (10). Following the prior observations, osteoblast number was not
3ASOs and Hajdu Cheney
different between control and Notch2tm1.1Ecan was not changed in Notch2tm1.1Ecan osteoblasts
mice. Accordingly, dynamic parameters of bone (10). Notch2 ASOs decreased Notch2 mRNA in
formation were not different between wild type wild type and mutant cells, and Notch29655C>T
and mutant mice and were not affected by mRNA in osteoblasts from Notch2tm1.1Ecan mice.
Notch2 ASOs. In accordance with the cellular In addition, Notch2 ASOs decreased Hey1, Hey2
phenotype of Notch2tm1.1Ecan mice, fasting serum and Tnfsf11 mRNA in cells from Notch2tm1.1Ecan
levels of carboxy-terminal collagen crosslinks mice, without an effect on Bglap expression
(CTX) were increased from (means ± SD; n = 5 (Figure 6).
to 6) control 34.6 ± 2.4 to 49.2 ± 8.9 ng/ml (p < Notch26955C>T mRNA was present in
0.05) in Notch2tm1.1Ecan mice treated with control osteocyte-enriched cultures from Notch2tm1.1Ecan
ASOs. Notch2 ASOs reduced the serum levels mice and not in control cultures, and Hey2 and
of CTX in both wild type mice to 24.1 ± 9.7 Tnfsf11 were significantly increased in
ng/ml (p < 0.052) and Notch2tm1.1Ecan mice to Notch2tm1.1Ecan cells (Figure 7). Notch2 ASOs
23.2 ± 3.9 ng/ml (p < 0.05) demonstrating a suppressed Notch2 mRNA in wild type and
normalization of bone resorption in mutant cells and Notch26955C>T mRNA levels in
experimental mice. cells from Notch2tm1.1Ecan mice, and suppressed
Hey2 and Tnfsf11 in Notch2tm1.1Ecan cells to levels
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
Effect of Notch2 ASOs on Notch2 Expression that were similar to those found in wild type cells
and Signaling in Osteoblast and Osteocyte Cell treated with control ASOs.
Cultures
Mouse Notch2 ASOs added to the Effect of Notch2 ASOs on Notch2 Expression
culture medium of osteoblast-enriched cells and Activity in BMM Cultures and Osteoclast
from wild type C57BL/6 mice at 1 to 20 µM Formation
decreased Notch2 mRNA by ~40 to ~80% 72 h Notch2 ASOs were added to either
after ASO addition without evidence of cellular BMM cultures at the initiation of the culture
toxicity or changes in cell replication (Figure 5). period or following the addition of RANKL for
The effect of the Notch2 ASO was specific for 2 days to determine their effect in cells of the
Notch2 mRNA since, at a concentration as high myeloid lineage and in osteoclast precursors.
as 20 µM, it did not decrease the expression of Mouse Notch2 ASOs at 1 and 5µM suppressed
Notch1, 3 or 4 mRNA. The NOTCH2 Notch2 mRNA levels in BMMs from wild type
intracellular domain (N2ICD), representative of C57BL/6 mice by 85 to 95% and in osteoclast
NOTCH2 cleavage and signal activation, was precursors by 70 to 85% without evidence of
increased in Notch2tm1.1Ecan osteoblasts and the cellular toxicity and without altering cell
truncated form of NOTCH2, lacking the PEST proliferation (Figure 8). Confirming results in
domain (N2ICD∆PEST) was detected only in osteoblast cultures, the NOTCH2 intracellular
Notch2tm1.1Ecan cells. Therefore, the total levels domain (N2ICD) was increased in Notch2tm1.1Ecan
of N2ICD, intact and truncated, were ~2-fold osteoclasts and the truncated form of NOTCH2,
greater in Notch2tm1.1Ecan cells than in control lacking the PEST domain (N2ICD∆PEST) was
cells (Figure 5). Notch2 ASOs decreased the detected only in Notch2tm1.1Ecan cells.
total levels of N2ICD in wild type and Consequently, the total levels of N2ICD, intact
Notch2tm1.1Ecan cells demonstrating a suppression and truncated, were ~2-fold greater in
of NOTCH2 activation. Notch26955C>T Notch2tm1.1Ecan cells than in control cells (Figure
transcripts were present in cells from 8). Notch2 ASOs decreased the total levels of
Notch2tm1.1Ecan mutant mice but not in control N2ICD in wild type and Notch2tm1.1Ecan cells
cultures, and Hey1 and Hey2 transcripts were demonstrating a suppression of NOTCH2
increased in Notch2tm1.1Ecan osteoblasts activation.
confirming that Notch signaling was activated There was a significant increase in
(Figure 6). In accordance with prior osteoclast formation in BMMs from
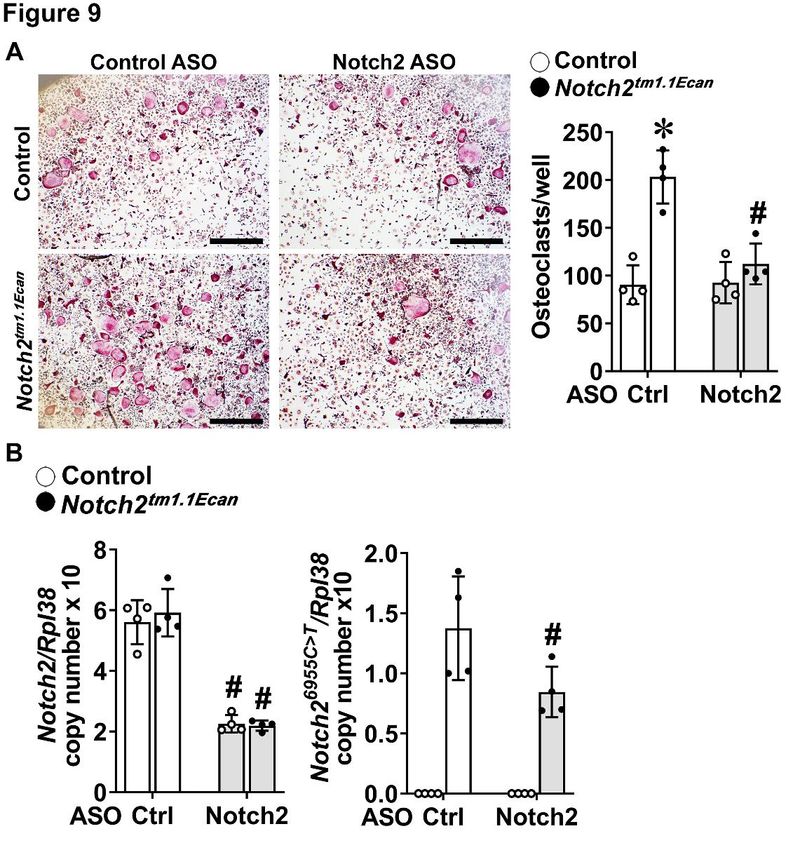
observations, tumor necrosis factor superfamily Notch2tm1.1Ecan mice cultured in the presence of
member 11 (Tnfsf11), encoding RANKL, was M-CSF and RANKL (Figure 9). The increased
induced whereas Bglap, encoding osteocalcin, osteoclastogenesis was prevented by the
4ASOs and Hajdu Cheney
addition of Notch2 ASOs to BMM cultures at 1 of RANKL by cells of the osteoblast lineage and
µM so that the osteoclastogenic potential of by inducing the differentiation of cells of the
Notch2tm1.1Ecan cells cultured with Notch2 ASOs myeloid lineage toward mature osteoclasts (15).
was no longer different from that of control cells. Notch2 ASOs decreased both effects in vitro and
The decrease in osteoclastogenesis by Notch2 decreased serum levels of CTX, a marker of
ASOs in Notch2tm1.1Ecan cells was associated with bone resorption, so that CTX levels in
a concomitant decrease in Notch2 wild type and Notch2tm1.1Ecan ASO-treated mice were not
Notch26955C>T mutant transcripts. different from those of wild type mice. These
effects would explain the amelioration of the
Discussion osteopenia observed in Notch2tm1.1Ecan mice.
Findings from the present work confirm Notch2 ASOs downregulated Notch2
that a mouse model replicating a mutation found and Notch26955C>T transcripts and decreased the
in HCS displays femoral cancellous and cortical enhanced Notch signaling found in
bone osteopenia. The osteopenic phenotype is Notch2tm1.1Ecan cells as well as in bone extracts
manifested early in life in mice of both sexes; without an effect on basal levels of Notch
and in the present study, we elected to treat 1 activation. Only Notch2tm1.1Ecan mutant cells
month old male mice with Notch2 ASOs in an synthesized the truncated form of the N2ICD
(N2ICD∆PEST) and the intact N2ICD. The
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
attempt to ameliorate the osteopenic femoral
phenotype of Notch2tm1.1Ecan mice (10). Because summation of the intact and truncated forms of
only male mice were treated one needs to be N2ICD resulted in a ~2-fold greater expression
cautious and not extrapolate the results to female of N2ICD in Notch2tm1.1Ecan mutants than in
mice. Phenotypic alterations of experimental control cells, and this was suppressed by Notch2
and control mice were assessed by µCT, and ASOs confirming the downregulation of Notch2
analyses required the ex vivo exam of bone signaling. The N2ICD∆PEST is more stable than
following the sacrifice of mice. Consequently, wild type N2ICD since it is resistant to ubiquitin-
the same animal could not be analyzed before mediated degradation, explaining the gain-of-
and after the administration of Notch2 ASOs. NOTCH2 function and the induction of Notch
Another limitation of the work is the fact that all target genes in Notch2tm1.1Ecan cells.
tm1.1Ecan
the analyses were performed in femoral bone Notch2 mice do not exhibit an increase in
since the osteopenia of Notch2tm1.1Ecan mice was osteoblast number or a bone-forming response to
established at this skeletal site (10). Whereas the increase in bone resorption, indicating a
Notch2 ASOs downregulated Notch2 wild type possible negative regulation of
and mutant transcripts in femoral bone, it was osteoblastogenesis or osteoblast function by the
not determined whether the same effect occurs at Notch2 mutation. However, in the present
other skeletal, possibly less vascularized, sites. studies we confirm that osteoblast gene markers,
The Notch2 ASO utilized is specific to Notch2 such as Bglap (osteocalcin), are not affected in
so that the results obtained should not be cells from Notch2tm1.1Ecan mice. The inactivation
attributed to the downregulation of other Notch of Notch2 in cells of the osteoblastic lineage
receptors. causes an increase in the osteogenic potential of
The phenotype of the Notch2tm1.1Ecan these cells suggesting an inhibitory role of Notch
mutant mouse recapitulates aspects of HCS signaling in osteoblastogenesis (54-56).
including osteopenia, short limbs and in the Although approaches to downregulate
homozygous state craniofacial abnormalities, Notch signaling are various, they are often not
including micrognathia, and early-lethality specific to this signaling pathway or to a specific
(10,15), and E. Canalis, unpublished Notch receptor. A recent alternative has been the
observations). However, neither Notch2tm1.1Ecan use of antibodies to the negative regulatory
nor an alternate murine model of HCS manifest region (NRR) of specific Notch receptors that
acroosteolysis (53). In the present work, we prevent the exposure of the NRR to the γ-
confirm that Notch2 has unique actions on secretase complex and thus the activation of
trabecular bone physiology and induces Notch (33-35). Recently, we demonstrated that
osteoclastogenesis by increasing the expression anti-Notch2 NRR antibodies reverse the skeletal
5ASOs and Hajdu Cheney
phenotype of Notch2tm1.1Ecan mice and anti- ASOs targeting Notch2 mRNA were
Notch3 NRR antibodies reverse the skeletal designed in silico by scanning through the
phenotype of Notch3tm1.1Ecan mice, a model of sequence of murine Notch2 pre-mRNA. The
Lateral Meningocele Syndrome (34,35). entire Notch2 pre-mRNA sequence was covered
Although anti-Notch NRR antibodies are for potential 16-mer oligonucleotides
specific, the significant downregulation of the complementary to the pre-mRNA. Sequence
Notch receptor throughout the organism may motifs that were intrinsically problematic
lead to potential side effects, such as because of unfavorable hybridization properties,
gastrointestinal toxicity. In the present studies, such as polyG stretches, or potential toxicity due
we demonstrate that downregulation of Notch to immunogenic responses, were avoided.
expression by specific Notch ASOs is a suitable Notch2 ASOs were tested for activity in vitro for
alternative to decrease Notch activation in downregulation of Notch2 mRNA in HEPA 1-6
conditions of Notch gain-of-function. Although cells at Ionis Pharmaceuticals (Carlsbad, CA),
the effect of Notch2 ASOs was less pronounced and 14 ASOs targeting Notch2 mRNA were
than the one reported with anti-Notch2 NRR screened for activity and toxicity in vivo at the
antibodies, Notch2 ASOs were effective at Korea Institute of Toxicology (KIT, Daejeon,
ameliorating the skeletal phenotype of Korea). To this end, 7-week-old BALB/c male
Notch2tm1.1Ecan mice and appeared to be well
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
mice were administered ASOs at a dose of 50
tolerated by this experimental model of HCS. mg/Kg once a week by subcutaneous injection
Although attempts have been made to for a total of 3.5 weeks (4 doses). Body weights
transport ASOs to bone, complex delivery were measured weekly and mice were
systems were necessary and the technology has euthanized 48 hours after the last dose of ASO.
not been applied to the correction of gene Liver, kidney and spleen were weighed,
mutations in the skeleton (57). In the present normalized to body weight and compared to
studies, we used a practical systemic approach to organs from control mice. Blood was obtained
downregulate Notch2 in skeletal and non- by cardiac puncture, and plasma was collected
skeletal tissue. We demonstrate that a second for the measurement of alanine
generation phosphorothioate modified murine aminotransferase, aspartate aminotransferase,
Notch2 ASO downregulated Notch2 in tissues total bilirubin, albumin and blood urea nitrogen.
where the gene is expressed and has a function, Total RNA was extracted from liver samples to
including bone. The decrease in Notch2 in a determine Notch2 mRNA levels corrected for
mouse model of Notch2 gain-of-function was cyclophilin A expression. Based on the
associated with a concomitant decrease in Notch information obtained, ASOs found to
target gene expression in skeletal cells downregulate Notch2 liver mRNA by more than
documenting a tempering effect on Notch 75% compared to a control mismatched ASO
activation. As a consequence, a recovery of without toxicity in vivo were selected.
bone mass was observed. Although this was not Procedures performed at KIT were approved by
complete, a significant effect on BV/TV was the KIT animal care and use committee. For the
achieved with amelioration of the Notch2tm1.1Ecan present studies, mouse Notch2 ASO Ionis
skeletal phenotype. 977472 of sequence GTTATATAATCTTCCA
In conclusion, Notch2 ASOs and control mismatched ASO Ionis 549144 of
downregulate Notch2 expression and signal sequence GGCCAATACGCCGTCA were
activation, and decrease RANKL and selected.
osteoclastogenesis in a model of HCS, and
consequently ameliorate its osteopenic Notch2tm1.1Ecan Mutant Mice
phenotype. The downregulation of NOTCH2 A mouse model of HCS, termed
may offer a potential therapeutic opportunity for Notch2tm1.1Ecan, harboring a 6955C>T
subjects with HCS in the future. substitution in exon 34 of Notch2 was previously
reported and validated (10). Notch2tm1.1Ecan mice
Experimental procedures were backcrossed into a C57BL/6J background
Notch2 Antisense Oligonucleotides for 8 or more generations and genotyping was
6ASOs and Hajdu Cheney
conducted in tail DNA extracts by polymerase hydroxyapatite (58,59). For analysis of femoral
chain reaction (PCR) using forward primer cortical bone, contours were iterated across 100
Nch2Lox gtF 5’–CCCTTCTCTCTGTGCGG slices along the cortical shell of the femoral
TAG-3’ and reverse primer Nch2Lox gtR 5’– midshaft, excluding the marrow cavity.
CTCAGAGCCAAAGCC TCACTG-3’. In the Analysis of bone volume/total volume, porosity,
present study, one month old mice heterozygous cortical thickness, total cross sectional and
for the Notch26955C>T allele and control mice cortical bone area, periosteal perimeter,
were obtained by crossing heterozygous mutants endosteal perimeter and material density were
with wild type mice to assess the impact of performed using a Gaussian filter (σ = 0.8,
Notch2 ASOs on the Notch2tm1.1Ecan skeletal support = 1), and a threshold of 400 permil
phenotype. One month old male Notch2tm1.1Ecan equivalent to 704.7 mg/cm3 hydroxyapatite.
heterozygous mutant and control sex-matched
littermate mice were treated with Notch2 ASO Bone Histomorphometric Analysis
(Ionis 977472) or control ASO (Ionis 549144) Static cancellous bone
suspended in phosphate buffered saline, and histomorphometry was carried out on
administered subcutaneously at a dose of 50 experimental and control mice. Five micron
mg/Kg once a week for 4 consecutive weeks. longitudinal sections of undecalcified femurs
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
Mice were euthanized at 2 months of age. embedded in methyl methacrylate were cut on a
Studies were approved by the Institutional microtome (Microm, Richards-Allan Scientific,
Animal Care and Use Committee of UConn Kalamazoo, MI), and stained with 0.1%
Health. toluidine blue. Static and dynamic parameters of
bone formation and resorption were measured in
Microcomputed Tomography (µCT) a defined area between 360 µm and 2160 µm
Bone microarchitecture of femurs from from the growth plate, using an OsteoMeasure
experimental and control mice was determined morphometry system (OsteoMetrics, Atlanta,
using a microcomputed tomography instrument GA). Stained sections were used to measure
(µCT 40; Scanco Medical AG, Bassersdorf, osteoblast and osteoclast number and eroded
Switzerland), which was calibrated periodically surface. Mineralizing surface per bone surface
using a phantom provided by the manufacturer and mineral apposition rate were measured on
(58,59). Femurs were scanned in 70% ethanol at unstained sections visualized under UV light and
high resolution, energy level of 55 kVp, intensity a triple diamidino-2-phenylindole/fluorescein/
of 145 µA and integration time of 200 ms. A Texas red set long pass filter, and bone formation
total of 100 slices at midshaft and 160 slices at rate was calculated. The terminology and units
the distal metaphysis were acquired at an used are those recommended by the
isotropic voxel size of 216 µm3 and a slice Histomorphometry Nomenclature Committee of
thickness of 6 µm, and chosen for analysis. the American Society for Bone and Mineral
Trabecular bone volume fraction and Research (60,61).
microarchitecture were evaluated starting
approximately 1.0 mm proximal from the Osteoblast-enriched Cell Cultures
femoral condyles. Contours were manually The parietal bones of 3 to 5 day old
drawn a few voxels away from the endocortical control and Notch2tm1.1Ecan mutant mice were
boundary every 10 slices to define the region of exposed to Liberase TL 1.2 U/ml (Sigma-
interest for analysis. The remaining slice Aldrich St. Louis, MO) for 20 min at 37°C and
contours were iterated automatically. cells extracted in 5 consecutive reactions (62).
Trabecular regions were assessed for total Cells from the last 3 digestions were pooled and
volume, bone volume, bone volume fraction seeded at a density of 10 x 104 cells/cm2, as
(bone volume/total volume), trabecular described (63). Osteoblast-enriched cells were
thickness, trabecular number, trabecular cultured in Dulbecco’s modified Eagle’s
separation, connectivity density and SMI, using medium (DMEM) supplemented with non-
a Gaussian filter (σ = 0.8), and a threshold of 240 essential amino acids (both from Life
permil equivalent to 355.5 mg/cm3 Technologies, Grand Island, NY) and 10% heat-
7ASOs and Hajdu Cheney
inactivated fetal bovine serum (FBS; Atlanta For osteoclast formation, cells were
Biologicals, Norcross, GA) in a humidified 5% collected following treatment with 0.05%
CO2 incubator at 37°C. Confluent osteoblast- trypsin/EDTA and seeded at a density of 4.7 x
enriched cells were exposed to DMEM 104 cells/cm2 on tissue culture plates in the
supplemented with 10% heat-inactivated FBS, presence of M-CSF at 30 ng/ml and murine
100 µg/ml ascorbic acid and 5 mM β- RANKL at 10 ng/ml until the formation of
glycerophosphate (both from Sigma-Aldrich) in multinucleated tartrate-resistant acid
the presence of Notch2 ASO or control ASO at phosphatase (TRAP)-positive cells. RANKL
various doses as indicated in figure legends. cDNA and expression vector were obtained from
M. Glogauer (Toronto, ON, Canada), and
Osteocyte-enriched Cultures glutathione S-transferase-tagged RANKL was
Femurs from 6 to 7 week old wild type expressed and purified as described (67). TRAP
or Notch2tm1.1Ecan mice were collected after enzyme histochemistry was conducted using a
sacrifice, surrounding tissues dissected, the commercial kit (Sigma Aldrich), in accordance
proximal epiphysis excised and the bone marrow with manufacturer’s instructions. TRAP-
removed by centrifugation. The distal epiphysis positive cells containing more than 3 nuclei were
was removed and, to release the endosteal and considered osteoclasts. Cultures were carried out
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
periosteal cellular layers, the femoral fragments in the presence of Notch2 or control ASO at
were sequentially exposed for 20 min periods to various doses as indicated in figure legends.
type II collagenase pretreated with Nα-Tosyl-L-
lysine chloromethylketone hydrochloride 17 Cell Proliferation Assay
µg/ml and EDTA 5 mM (Life Technologies) at Cell replication was determined using
37°C, as described (4,64). Osteocyte-enriched the Cell Counting Kit-8 (CCK-8). In this kit, the
bone fragments were obtained and cultured tetrazolium salt WST-8 [2-(2-methoxy-4-
individually in DMEM supplemented with nitrophenyl)-3-(4-nitrophenhyl)-5-(2,4-
nonessential amino acids, 100 µg/ml ascorbic disulfophenyl)-2H-tetrazolium, monosodium
acid and heat-inactivated 10% FBS for 72 hours salt] produces a formazan dye, measured at an
in a humidified 5% CO2 incubator at 37°C in the absorbance of 450 nm, upon reduction by
presence of control or Notch2 ASOs, as cellular dehydrogenases. The assay quantifies
indicated in figure legends (4,65). viable cells and was used in accordance with
manufacturer’s instructions (Dojindo Molecular
Bone Marrow-derived Macrophage (BMM) Technologies, Rockville, MD).
Cultures and Osteoclast Formation
To obtain BMMs, bone marrow cells Quantitative Reverse Transcription (qRT)-PCR
were isolated from long bones by flushing the Total RNA was extracted from either
marrow with a 26 gauge needle. Red blood cells cultured cells or tibiae following the removal of
were lysed in lysis buffer containing 150 mM the bone marrow by centrifugation, and mRNA
NH4Cl, 10 mM KHCO3 and 0.1 mM EDTA (pH levels determined by qRT-PCR (68,69). For this
7.4). The cell suspension was centrifuged and purpose, equal amounts of RNA were reverse-
the pellet suspended in α-minimum essential transcribed using the iScript RT-PCR kit
medium (α-MEM) (Life Technologies) (BioRad, Hercules, CA), according to
containing 10% heat-inactivated FBS and manufacturer’s instructions, and were amplified
recombinant human macrophage colony in the presence of specific primers (Table 3, all
stimulating factor (M-CSF) at 30 ng/ml. M-CSF primers from Integrated DNA Technologies
complementary DNA (cDNA) and expression (IDT), Coralville, IA), and iQ SYBR Green
vector were obtained from D. Fremont (St. Supermix (BioRad), at 60ºC for 35 cycles.
Louis, MO) and M-CSF was purified as Transcript copy number was estimated by
previously reported (66). Cells were seeded at a comparison with a serial dilution of cDNA for
density of 3 x 105 cells/cm2 on uncoated Petri Bglap (from J. Lian, Burlington, VT) Hey1 and
dishes and cultured for 3 days. Hey2 (both from T. Iso, Gunma, Japan), HeyL
(from D. Srivastava, San Francisco, CA), Notch2
8ASOs and Hajdu Cheney
(from Thermo Fisher Scientific), Notch1 (from Immunoblotting
J.S. Nye, Cambridge, MA), Notch4 (from Y. Pre-osteoclasts or osteoblasts from
Shirayoshi, Tottori, Japan) or Tnfsf11 (from control or Notch2tm1.1Ecan mice were extracted in
Source BioScience, Nottingham, UK) (70-75). buffer containing 25 mM Tris-HCl (pH 7.5), 150
Notch3 copy number was estimated by mM NaCl, 5% glycerol, 1 mM EDTA, 0.5%
comparison to a serial dilution of a ~100 base Triton X-100, 1 mM sodium orthovanadate, 10
pairs synthetic DNA template (IDT) cloned into mM NaF, 1 mM phenyl methyl sulfonyl fluoride
pcDNA3.1 (Thermo Fischer Scientific) by and protease inhibitor cocktail (all from Sigma
isothermal single reaction assembly using Aldrich). Quantified total cell lysates (35 µg of
commercially available reagents (New England total protein) were separated by sodium dodecyl
BioLabs, Ipswich, MA) (76). sulfate (SDS)-polyacrylamide gel electro-
To measure levels of the Notch26955C>T phoresis (PAGE) in 8% polyacrylamide gels and
mutant transcript, total RNA was reverse transferred to Immobilon-P membranes
transcribed with Moloney murine leukemia virus (Millipore, Billerica, MA). The blots were
reverse transcriptase in accordance with probed with anti-NOTCH2 (C651.6DbHN)
manufacturer’s instructions (Life Technologies) antibodies (Developmental Studies Hybridoma
in the presence of reverse primers for Notch2 and Bank (DSHB C651.6DbHN, University of Iowa,
Iowa City, IA)) and β-actin (3700) antibodies
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
of reverse primers for ribosomal protein L38
(Rpl38) (Table 3). Notch2 cDNA was amplified (Cell Signaling Technology, Danvers, MA) and
by PCR in the presence of specific primers, a exposed to anti-rabbit IgG and anti-rat IgG
TET labeled DNA probe of sequence 5’- conjugated to horseradish peroxidase (HRP)
CATTGCCTAGGCAGC-3’ covalently bound (Sigma-Aldrich) and incubated with a
to a 3’-minor groove binder quencher (Life chemiluminescence detection reagent (Bio-
Technologies), and SsoAdvanced Universal Rad). Chemiluminescence was detected by
Probes Supermix (BioRad) at 60ºC for 45 cycles ChemiDoc™ XSR+ molecular imager (Bio-
(10,77). Notch26955C>T transcript copy number Rad) with Image Lab™ software (version 5.2.1)
was estimated by comparison with a serial and the amount of protein in individual bands
dilution of a synthetic DNA fragment (IDT) was quantified (15).
containing ~200 bp surrounding the 6955C>T
mutation in the Notch2 locus, and was cloned Serum Carboxy-terminal Collagen Crosslinks
into pcDNA3.1(-) (Life Technologies) by Assay
isothermal single-reaction assembly using Serum samples from control and
commercially available reagents (New England experimental mice were obtained after an
Biolabs, Ipswich, MA) (76). overnight fast. CTX levels were measured using
Amplification reactions were conducted an enzyme-linked immunosorbent assay kit
in a CFX96 qRT-PCR detection system according to manufacturer’s instructions
(BioRad), and fluorescence was monitored (Immunodiagnostic Systems, Gaithersburg,
during every PCR cycle at the annealing step. MD).
Data are expressed as copy number corrected for
Rpl38 copy number, estimated by comparison Statistics
with a serial dilution of Rpl38 cDNA (from Data are expressed as means ± standard
ATCC) (78). In selected experiments, control deviations (SD). Statistical differences were
data were normalized to 1 following correction determined by analysis of variance with Holm-
for Rpl38 expression. Sidak’s post-hoc analysis for pairwise or
multiple comparisons.
9ASOs and Hajdu Cheney
Acknowledgments: The authors thank D, Srivastava for HeyL cDNA, D. Fremont for M-CSF cDNA,
M. Glogauer for RANKL cDNA, T. Iso for Hey1 and Hey2 cDNA, J. Lian for Bglap cDNA, J.S. Nye for
Notch1 cDNA, Y. Shirayoshi for Notch4 cDNA, T. Eller for technical assistance and Mary Yurczak for
secretarial support. This work was supported by grants DK045227 (EC) from the National Institute of
Diabetes and Digestive and Kidney Diseases and AR076747 (EC) from the National Institute of Arthritis
and Musculoskeletal and Skin Diseases.
Disclosure statement: Tamar R. Grossman and Michele Carrer are employed by Ionis Pharmaceuticals,
Inc., Ernesto Canalis, Lauren Schilling and Jungeun Yu have nothing to disclose.
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
10ASOs and Hajdu Cheney
REFERENCES
1. Siebel C, Lendahl U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol
Rev. 2017;97(4):1235-1294.
2. Zanotti S, Canalis E. Notch Signaling and the Skeleton. Endocr Rev. 2016;37(3):223-253.
3. Canalis E. Notch in skeletal physiology and disease. Osteoporos Int. 2018;29(12):2611-2621.
4. Zanotti S, Canalis E. Parathyroid hormone inhibits Notch signaling in osteoblasts and osteocytes.
Bone. 2017;103:159-167.
5. Zanotti S, Canalis E. Notch and the Skeleton. Mol Cell Biol. 2010;30(4):886-896.
6. Schroeter EH, Kisslinger JA, Kopan R. Notch-1 signalling requires ligand-induced proteolytic
release of intracellular domain. Nature. 1998;393(6683):382-386.
7. Kovall RA. More complicated than it looks: assembly of Notch pathway transcription complexes.
Oncogene. 2008;27(38):5099-5109.
8. Nam Y, Sliz P, Song L, Aster JC, Blacklow SC. Structural basis for cooperativity in recruitment
of MAML coactivators to Notch transcription complexes. Cell. 2006;124(5):973-983.
9. Wilson JJ, Kovall RA. Crystal structure of the CSL-Notch-Mastermind ternary complex bound
to DNA. Cell. 2006;124(5):985-996.
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
10. Canalis E, Schilling L, Yee SP, Lee SK, Zanotti S. Hajdu Cheney Mouse Mutants Exhibit
Osteopenia, Increased Osteoclastogenesis and Bone Resorption. J Biol Chem. 2016;291:1538-
1551.
11. Bai S, Kopan R, Zou W, Hilton MJ, Ong CT, Long F, Ross FP, Teitelbaum SL. NOTCH1
regulates osteoclastogenesis directly in osteoclast precursors and indirectly via osteoblast lineage
cells. J Biol Chem. 2008;283(10):6509-6518.
12. Fukushima H, Nakao A, Okamoto F, Shin M, Kajiya H, Sakano S, Bigas A, Jimi E, Okabe K.
The association of Notch2 and NF-kappaB accelerates RANKL-induced osteoclastogenesis. Mol
Cell Biol. 2008;28(20):6402-6412.
13. Canalis E, Yu J, Schilling L, Yee SP, Zanotti S. The lateral meningocele syndrome mutation
causes marked osteopenia in mice. J Biol Chem. 2018;293(36):14165-14177.
14. Canalis E, Parker K, Feng JQ, Zanotti S. Osteoblast Lineage-specific Effects of Notch Activation
in the Skeleton. Endocrinology. 2013;154(2):623-634.
15. Yu J, Canalis E. The Hajdu Cheney mutation sensitizes mice to the osteolytic actions of tumor
necrosis factor alpha. J Biol Chem. 2019;294(39):14203-14214.
16. Cheney WD. Acro-Osteolysis. Am J Roentgenol Radium Ther Nucl Med. 1965;94:595-607.
17. Hajdu N, Kauntze R. Cranio-skeletal dysplasia. Br J Radiol. 1948;21(241):42-48.
18. Canalis E. Clinical and experimental aspects of notch receptor signaling: Hajdu-Cheney
syndrome and related disorders. Metabolism. 2018;80:48-56.
19. Gray MJ, Kim CA, Bertola DR, Arantes PR, Stewart H, Simpson MA, Irving MD, Robertson SP.
Serpentine fibula polycystic kidney syndrome is part of the phenotypic spectrum of Hajdu-
Cheney syndrome. Eur J Hum Genet. 2012;20(1):122-124.
20. Isidor B, Lindenbaum P, Pichon O, Bezieau S, Dina C, Jacquemont S, Martin-Coignard D,
Thauvin-Robinet C, Le MM, Mandel JL, David A, Faivre L, Cormier-Daire V, Redon R, Le CC.
Truncating mutations in the last exon of NOTCH2 cause a rare skeletal disorder with
osteoporosis. Nat Genet. 2011;43(4):306-308.
21. Majewski J, Schwartzentruber JA, Caqueret A, Patry L, Marcadier J, Fryns JP, Boycott KM, Ste-
Marie LG, McKiernan FE, Marik I, Van EH, Michaud JL, Samuels ME. Mutations in NOTCH2
in families with Hajdu-Cheney syndrome. Hum Mutat. 2011;32(10):1114-1117.
22. Simpson MA, Irving MD, Asilmaz E, Gray MJ, Dafou D, Elmslie FV, Mansour S, Holder SE,
Brain CE, Burton BK, Kim KH, Pauli RM, Aftimos S, Stewart H, Kim CA, Holder-Espinasse M,
Robertson SP, Drake WM, Trembath RC. Mutations in NOTCH2 cause Hajdu-Cheney syndrome,
a disorder of severe and progressive bone loss. Nat Genet. 2011;43(4):303-305.
11ASOs and Hajdu Cheney
23. Zhao W, Petit E, Gafni RI, Collins MT, Robey PG, Seton M, Miller KK, Mannstadt M. Mutations
in NOTCH2 in patients with Hajdu-Cheney syndrome. Osteoporos Int. 2013;24(8):2275-2281.
24. Udell J, Schumacher HR, Jr., Kaplan F, Fallon MD. Idiopathic familial acroosteolysis:
histomorphometric study of bone and literature review of the Hajdu-Cheney syndrome. Arthritis
Rheum. 1986;29(8):1032-1038.
25. Blumenauer BT, Cranney AB, Goldstein R. Acro-osteolysis and osteoporosis as manifestations
of the Hajdu-Cheney syndrome. Clin Exp Rheumatol. 2002;20(4):574-575.
26. Sakka S, Gafni RI, Davies JH, Clarke B, Tebben P, Samuels M, Saraff V, Klaushofer K, Fratzl-
Zelman N, Roschger P, Rauch F, Hogler W. Bone Structural Characteristics and Response to
Bisphosphonate Treatment in Children With Hajdu-Cheney Syndrome. J Clin Endocrinol Metab.
2017;102(11):4163-4172.
27. Adami G, Rossini M, Gatti D, Orsolini G, Idolazzi L, Viapiana O, Scarpa A, Canalis E. Hajdu
Cheney Syndrome; report of a novel NOTCH2 mutation and treatment with denosumab. Bone.
2016;92:150-156.
28. Ryeom SW. The cautionary tale of side effects of chronic Notch1 inhibition. J Clin Invest.
2011;121(2):508-509.
29. De Strooper B, Annaert W, Cupers P, Saftig P, Craessaerts K, Mumm JS, Schroeter EH,
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
Schrijvers V, Wolfe MS, Ray WJ, Goate A, Kopan R. A presenilin-1-dependent gamma-
secretase-like protease mediates release of Notch intracellular domain. Nature.
1999;398(6727):518-522.
30. Duggan SP, McCarthy JV. Beyond gamma-secretase activity: The multifunctional nature of
presenilins in cell signalling pathways. Cell Signal. 2016;28(1):1-11.
31. Ilagan MX, Kopan R. Selective blockade of transport via SERCA inhibition: the answer for
oncogenic forms of Notch? Cancer Cell. 2013;23(3):267-269.
32. Moellering RE, Cornejo M, Davis TN, Del BC, Aster JC, Blacklow SC, Kung AL, Gilliland DG,
Verdine GL, Bradner JE. Direct inhibition of the NOTCH transcription factor complex. Nature.
2009;462(7270):182-188.
33. Wu Y, Cain-Hom C, Choy L, Hagenbeek TJ, de Leon GP, Chen Y, Finkle D, Venook R, Wu X,
Ridgway J, Schahin-Reed D, Dow GJ, Shelton A, Stawicki S, Watts RJ, Zhang J, Choy R, Howard
P, Kadyk L, Yan M, Zha J, Callahan CA, Hymowitz SG, Siebel CW. Therapeutic antibody
targeting of individual Notch receptors. Nature. 2010;464(7291):1052-1057.
34. Canalis E, Sanjay A, Yu J, Zanotti S. An Antibody to Notch2 Reverses the Osteopenic Phenotype
of Hajdu-Cheney Mutant Male Mice. Endocrinology. 2017;158(4):730-742.
35. Yu J, Siebel CW, Schilling L, Canalis E. An antibody to Notch3 reverses the skeletal phenotype
of lateral meningocele syndrome in male mice. J Cell Physiol. 2020;235(1):210-220.
36. Bennett CF, Baker BF, Pham N, Swayze E, Geary RS. Pharmacology of Antisense Drugs. Annu
Rev Pharmacol Toxicol. 2017;57:81-105.
37. Cerritelli SM, Crouch RJ. Ribonuclease H: the enzymes in eukaryotes. FEBS J.
2009;276(6):1494-1505.
38. Murray SF, Jazayeri A, Matthes MT, Yasumura D, Yang H, Peralta R, Watt A, Freier S, Hung
G, Adamson PS, Guo S, Monia BP, LaVail MM, McCaleb ML. Allele-Specific Inhibition of
Rhodopsin With an Antisense Oligonucleotide Slows Photoreceptor Cell Degeneration. Invest
Ophthalmol Vis Sci. 2015;56(11):6362-6375.
39. Shy ME. Antisense oligonucleotides offer hope to patients with Charcot-Marie-Tooth disease
type 1A. J Clin Invest. 2018;128(1):110-112.
40. Carroll JB, Warby SC, Southwell AL, Doty CN, Greenlee S, Skotte N, Hung G, Bennett CF,
Freier SM, Hayden MR. Potent and selective antisense oligonucleotides targeting single-
nucleotide polymorphisms in the Huntington disease gene / allele-specific silencing of mutant
huntingtin. Mol Ther. 2011;19(12):2178-2185.
12ASOs and Hajdu Cheney
41. Limmroth V, Barkhof F, Desem N, Diamond MP, Tachas G, Group ATLS. CD49d antisense drug
ATL1102 reduces disease activity in patients with relapsing-remitting MS. Neurology.
2014;83(20):1780-1788.
42. McCampbell A, Cole T, Wegener AJ, Tomassy GS, Setnicka A, Farley BJ, Schoch KM, Hoye
ML, Shabsovich M, Sun L, Luo Y, Zhang M, Comfort N, Wang B, Amacker J, Thankamony S,
Salzman DW, Cudkowicz M, Graham DL, Bennett CF, Kordasiewicz HB, Swayze EE, Miller
TM. Antisense oligonucleotides extend survival and reverse decrement in muscle response in
ALS models. J Clin Invest. 2018;128(8):3558-3567.
43. Zhao HT, Damle S, Ikeda-Lee K, Kuntz S, Li J, Mohan A, Kim A, Hung G, Scheideler MA,
Scherer SS, Svaren J, Swayze EE, Kordasiewicz HB. PMP22 antisense oligonucleotides reverse
Charcot-Marie-Tooth disease type 1A features in rodent models. J Clin Invest. 2018;128(1):359-
368.
44. Zhu C, Kim K, Wang X, Bartolome A, Salomao M, Dongiovanni P, Meroni M, Graham MJ,
Yates KP, Diehl AM, Schwabe RF, Tabas I, Valenti L, Lavine JE, Pajvani UB. Hepatocyte Notch
activation induces liver fibrosis in nonalcoholic steatohepatitis. Sci Transl Med. 2018;10(468).
45. Crooke ST, Witztum JL, Bennett CF, Baker BF. RNA-Targeted Therapeutics. Cell metabolism.
2018;27(4):714-739.
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
46. Wang FS, Wu RW, Ko JY, Tai MH, Ke HC, Yeh DW, Wu SL, Chen MW. Heat shock protein
60 protects skeletal tissue against glucocorticoid-induced bone mass loss by regulating osteoblast
survival. Bone. 2011;49(5):1080-1089.
47. Wang FS, Ko JY, Lin CL, Wu HL, Ke HJ, Tai PJ. Knocking down dickkopf-1 alleviates estrogen
deficiency induction of bone loss. A histomorphological study in ovariectomized rats. Bone.
2007;40(2):485-492.
48. Yu J, Zanotti S, Walia B, Jellison E, Sanjay A, Canalis E. The Hajdu Cheney Mutation Is a
Determinant of B-Cell Allocation of the Splenic Marginal Zone. The American journal of
pathology. 2018;188(1):149-159.
49. Saito T, Chiba S, Ichikawa M, Kunisato A, Asai T, Shimizu K, Yamaguchi T, Yamamoto G, Seo
S, Kumano K, Nakagami-Yamaguchi E, Hamada Y, Aizawa S, Hirai H. Notch2 is preferentially
expressed in mature B cells and indispensable for marginal zone B lineage development.
Immunity. 2003;18(5):675-685.
50. Sparks EE, Huppert KA, Brown MA, Washington MK, Huppert SS. Notch signaling regulates
formation of the three-dimensional architecture of intrahepatic bile ducts in mice. Hepatology.
2010;51(4):1391-1400.
51. McCright B, Gao X, Shen L, Lozier J, Lan Y, Maguire M, Herzlinger D, Weinmaster G, Jiang R,
Gridley T. Defects in development of the kidney, heart and eye vasculature in mice homozygous
for a hypomorphic Notch2 mutation. Development. 2001;128(4):491-502.
52. Liu Z, Chen S, Boyle S, Zhu Y, Zhang A, Piwnica-Worms DR, Ilagan MX, Kopan R. The
extracellular domain of Notch2 increases its cell-surface abundance and ligand responsiveness
during kidney development. Developmental cell. 2013;25(6):585-598.
53. Vollersen N, Hermans-Borgmeyer I, Cornils K, Fehse B, Rolvien T, Triviai I, Jeschke A, Oheim
R, Amling M, Schinke T, Yorgan TA. High Bone Turnover in Mice Carrying a Pathogenic
Notch2 Mutation Causing Hajdu-Cheney Syndrome. J Bone Miner Res. 2018;33(1):70-83.
54. Yorgan T, Vollersen N, Riedel C, Jeschke A, Peters S, Busse B, Amling M, Schinke T.
Osteoblast-specific Notch2 inactivation causes increased trabecular bone mass at specific sites of
the appendicular skeleton. Bone. 2016;87:136-146.
55. Zanotti S, Smerdel-Ramoya A, Stadmeyer L, Durant D, Radtke F, Canalis E. Notch inhibits
osteoblast differentiation and causes osteopenia. Endocrinology. 2008;149(8):3890-3899.
56. Deregowski V, Gazzerro E, Priest L, Rydziel S, Canalis E. Notch 1 Overexpression Inhibits
Osteoblastogenesis by Suppressing Wnt/beta-Catenin but Not Bone Morphogenetic Protein
Signaling. J Biol Chem. 2006;281(10):6203-6210.
13ASOs and Hajdu Cheney
57. Zhang G, Guo B, Wu H, Tang T, Zhang BT, Zheng L, He Y, Yang Z, Pan X, Chow H, To K, Li
Y, Li D, Wang X, Wang Y, Lee K, Hou Z, Dong N, Li G, Leung K, Hung L, He F, Zhang L, Qin
L. A delivery system targeting bone formation surfaces to facilitate RNAi-based anabolic therapy.
Nature medicine. 2012;18(2):307-314.
58. Bouxsein ML, Boyd SK, Christiansen BA, Guldberg RE, Jepsen KJ, Muller R. Guidelines for
assessment of bone microstructure in rodents using micro-computed tomography. J Bone Miner
Res. 2010;25(7):1468-1486.
59. Glatt V, Canalis E, Stadmeyer L, Bouxsein ML. Age-Related Changes in Trabecular Architecture
Differ in Female and Male C57BL/6J Mice. J Bone Miner Res. 2007;22(8):1197-1207.
60. Dempster DW, Compston JE, Drezner MK, Glorieux FH, Kanis JA, Malluche H, Meunier PJ, Ott
SM, Recker RR, Parfitt AM. Standardized nomenclature, symbols, and units for bone
histomorphometry: a 2012 update of the report of the ASBMR Histomorphometry Nomenclature
Committee. J Bone Miner Res. 2013;28(1):2-17.
61. Parfitt AM, Drezner MK, Glorieux FH, Kanis JA, Malluche H, Meunier PJ, Ott SM, Recker RR.
Bone histomorphometry: standardization of nomenclature, symbols, and units. Report of the
ASBMR Histomorphometry Nomenclature Committee. J Bone Miner Res. 1987;2(6):595-610.
62. Yesil P, Michel M, Chwalek K, Pedack S, Jany C, Ludwig B, Bornstein SR, Lammert E. A new
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
collagenase blend increases the number of islets isolated from mouse pancreas. Islets.
2009;1(3):185-190.
63. McCarthy TL, Centrella M, Canalis E. Further biochemical and molecular characterization of
primary rat parietal bone cell cultures. J Bone Miner Res. 1988;3(4):401-408.
64. Halleux C, Kramer I, Allard C, Kneissel M. Isolation of mouse osteocytes using cell fractionation
for gene expression analysis. Methods Mol Biol. 2012;816:55-66.
65. Canalis E, Schilling L, Zanotti S. Effects of Sex and Notch Signaling on the Osteocyte Cell Pool.
J Cell Physiol. 2017;232(2):363-370.
66. Lee SH, Rho J, Jeong D, Sul JY, Kim T, Kim N, Kang JS, Miyamoto T, Suda T, Lee SK, Pignolo
RJ, Koczon-Jaremko B, Lorenzo J, Choi Y. v-ATPase V0 subunit d2-deficient mice exhibit
impaired osteoclast fusion and increased bone formation. Nature medicine. 2006;12(12):1403-
1409.
67. Wang Y, Lebowitz D, Sun C, Thang H, Grynpas MD, Glogauer M. Identifying the relative
contributions of Rac1 and Rac2 to osteoclastogenesis. J Bone Miner Res. 2008;23(2):260-270.
68. Nazarenko I, Pires R, Lowe B, Obaidy M, Rashtchian A. Effect of primary and secondary
structure of oligodeoxyribonucleotides on the fluorescent properties of conjugated dyes. Nucleic
Acids Res. 2002;30(9):2089-2195.
69. Nazarenko I, Lowe B, Darfler M, Ikonomi P, Schuster D, Rashtchian A. Multiplex quantitative
PCR using self-quenched primers labeled with a single fluorophore. Nucleic Acids Res.
2002;30(9):e37.
70. Iso T, Sartorelli V, Chung G, Shichinohe T, Kedes L, Hamamori Y. HERP, a new primary target
of Notch regulated by ligand binding. Mol Cell Biol. 2001;21(17):6071-6079.
71. Nakagawa O, Nakagawa M, Richardson JA, Olson EN, Srivastava D. HRT1, HRT2, and HRT3:
a new subclass of bHLH transcription factors marking specific cardiac, somitic, and pharyngeal
arch segments. Dev Biol. 1999;216(1):72-84.
72. Wong BR, Rho J, Arron J, Robinson E, Orlinick J, Chao M, Kalachikov S, Cayani E, Bartlett FS,
3rd, Frankel WN, Lee SY, Choi Y. TRANCE is a novel ligand of the tumor necrosis factor
receptor family that activates c-Jun N-terminal kinase in T cells. J Biol Chem.
1997;272(40):25190-25194.
73. Nye JS, Kopan R, Axel R. An activated Notch suppresses neurogenesis and myogenesis but not
gliogenesis in mammalian cells. Development. 1994;120(9):2421-2430.
74. Shirayoshi Y, Yuasa Y, Suzuki T, Sugaya K, Kawase E, Ikemura T, Nakatsuji N. Proto-oncogene
of int-3, a mouse Notch homologue, is expressed in endothelial cells during early embryogenesis.
Genes Cells. 1997;2(3):213-224.
14ASOs and Hajdu Cheney
75. Lian J, Stewart C, Puchacz E, Mackowiak S, Shalhoub V, Collart D, Zambetti G, Stein G.
Structure of the rat osteocalcin gene and regulation of vitamin D-dependent expression. Proc Natl
Acad Sci U S A. 1989;86(4):1143-1147.
76. Gibson DG, Young L, Chuang RY, Venter JC, Hutchison CA, 3rd, Smith HO. Enzymatic
assembly of DNA molecules up to several hundred kilobases. Nat Methods. 2009;6(5):343-345.
77. Kutyavin IV, Afonina IA, Mills A, Gorn VV, Lukhtanov EA, Belousov ES, Singer MJ, Walburger
DK, Lokhov SG, Gall AA, Dempcy R, Reed MW, Meyer RB, Hedgpeth J. 3'-minor groove
binder-DNA probes increase sequence specificity at PCR extension temperatures. Nucleic Acids
Res. 2000;28(2):655-661.
78. Kouadjo KE, Nishida Y, Cadrin-Girard JF, Yoshioka M, St-Amand J. Housekeeping and tissue-
specific genes in mouse tissues. BMC Genomics. 2007;8:127.
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
15ASOs and Hajdu Cheney
FOOTNOTES
This work was supported by grant DK045227 (EC) from the National Institute of Diabetes and Digestive
and Kidney Diseases and grant AR076747 (EC) from the National Institute of Arthritis and
Musculoskeletal and Skin Diseases. The content is solely the responsibility of the authors and does not
necessarily represent the official views of the National Institutes of Health.
The abbreviations used are: αMEM, α-minimum essential medium; ASO, antisense oligonucleotide;
BMM, bone marrow macrophage; BV/TV, bone volume/tissue volume; CTX, carboxy-terminal collagen
crosslinks; CCK, Cell Counting Kit; cDNA, complementary DNA; Ctrl, control; Conn.D, connectivity
density; DMEM, Dulbecco’s modified Eagle’s medium; FBS, fetal bovine serum; HCS, Hajdu Cheney
Syndrome; HES, Hairy Enhancer of Split; HEY, Hes-related with YRPW; HRP, horseradish peroxidase;
HA, hydroxyapatite; M-CSF, macrophage colony stimulating factor; MAML, mastermind; μCT,
microcomputed tomography; NICD, NOTCH intracellular domain; N2ICD, NOTCH2 intracellular
domain; NRR, negative regulatory region: Ob, osteoblasts; OCP, osteoclast precursors; PEST, proline
(P), glutamic acid (E), serine (S) and threonine (T); PAGE, polyacrylamide gel electrophoresis; PCR,
polymerase chain reaction; qRT, quantitative reverse transcription; RANKL, receptor activator of nuclear
factor Kappa B ligand; RBPJκ, recombination signal-binding protein for Ig of κ; Rpl38, ribosomal protein
Downloaded from http://www.jbc.org/ by guest on March 25, 2020
L38; SDS, sodium dodecyl sulfate; SD, standard deviation; SMI, structure model index; TRAP, tartrate-
resistant acid phosphatase; Tb.Th, trabecular thickness; Tb.N, trabecular number; Tb.Sp, trabecular
separation; Tnfsf11, tumor necrosis factor superfamily member 11
16You can also read

















































