LUSH Odorant-Binding Protein Mediates Chemosensory Responses to Alcohols in Drosophila melanogaster
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Copyright 1998 by the Genetics Society of America
LUSH Odorant-Binding Protein Mediates Chemosensory Responses
to Alcohols in Drosophila melanogaster
Min-Su Kim, Allen Repp and Dean P. Smith
Department of Pharmacology, University of Texas Southwestern Medical Center, Dallas, Texas 75235-9111
Manuscript received March 30, 1998
Accepted for publication June 23, 1998
ABSTRACT
The molecular mechanisms mediating chemosensory discrimination in insects are unknown. Using the
enhancer trapping approach, we identified a new Drosophila mutant, lush, with odorant-specific defects
in olfactory behavior. lush mutant flies are abnormally attracted to high concentrations of ethanol, propanol,
and butanol but have normal chemosensory responses to other odorants. We show that wild-type flies
have an active olfactory avoidance mechanism to prevent attraction to concentrated alcohol, and this
response is defective in lush mutants. This suggests that the defective olfactory behavior associated with
the lush mutation may result from a specific defect in chemoavoidance. lush mutants have a 3-kb deletion
that produces a null allele of a new member of the invertebrate odorant-binding protein family, LUSH.
LUSH is normally expressed exclusively in a subset of trichoid chemosensory sensilla located on the
ventral-lateral surface of the third antennal segment. LUSH is secreted from nonneuronal support cells
into the sensillum lymph that bathes the olfactory neurons within these sensilla. Reintroduction of a
cloned wild-type copy of lush into the mutant background completely restores wild-type olfactory behavior,
demonstrating that this odorant-binding protein is required in a subset of sensilla for normal chemosensory
behavior to a subset of odorants. These findings provide direct evidence that odorant-binding proteins
are required for normal chemosensory behavior in Drosophila and may partially determine the chemical
specificity of olfactory neurons in vivo.
I NSECTS like Drosophila detect odorants with che-
mosensory hairs or “sensilla” located primarily on
the third antennal segment (Stocker 1994). The sen-
the central nervous system where odorant information is
processed in complex neural networks called glomeruli
(reviewed in Shepherd and Greer 1990). Olfactory
silla are hollow, fluid-filled structures encasing the olfac- information is subsequently delivered to higher brain
tory neuron dendrites of one to four olfactory neurons, centers and ultimately perceived as odor. Odorant per-
and therefore provide for anatomical segregation of ception can dramatically influence animal behaviors
olfactory neurons. In Drosophila, these sensilla fall into ranging from attraction to food sources and avoidance
three distinct morphological classes: basiconic, coelo- of noxious compounds to mediation of reproductive
conic, and trichoid (Stocker 1994; Riesgo-Escovar et cues (reviewed in Halpern 1987; Bargmann et al. 1990;
al. 1997). All three classes are thought to mediate olfac- Hall 1994; Pfeiffer and Johnston 1994; Roelofs
tory responses. Odor molecules pass through pores or 1995).
grooves within the cuticle of the sensilla where they In Drosophila, each of the approximately 2000 anten-
enter the sensillum lymph bathing the olfactory neuron nal olfactory neurons project their axons directly to the
dendrites (Steinbrecht 1969; Altner and Prillinger bilateral antennal lobes, the Drosophila equivalent of
1980; Riesgo-Escovar et al. 1997). Extracellular re- the olfactory bulbs. Each neuron synapses exclusively in
cordings of the odor-induced electrical activity from one of the 35 glomeruli, either ipsilaterally or bilaterally
different regions of the Drosophila antenna reveal dif- (Stocker et al. 1983; Stocker 1994). Antennal lobe
ferent regions have differential sensitivity to specific output is routed to higher brain structures including
odorants (Siddiqi 1987; Ayer and Carlson 1992; the mushroom bodies where memory is thought to be
Dubin et al. 1995). However, the molecular mechanisms consolidated (Davis et al. 1995). Different odorants pro-
that confer odor specificity to olfactory neurons in in- duce different patterns of glomerular activation in Dro-
sects are not understood (reviewed in Smith 1996). sophila antennal lobes (Rodrigues and Buchner 1984;
In both vertebrates and insects, primary olfactory neu- Rodrigues 1988). Flies injected with 3H-labeled 2-de-
rons activated by odorants make their first synapses in oxyglucose and exposed to repetitive odorant pulses are
labeled in antennal lobe glomeruli, and the labeling
pattern is different upon exposure to different odorants
(Rodrigues 1988). Similar results have been observed
Corresponding author: Dean P. Smith, Department of Pharmacology,
University of Texas Southwestern Medical Center, 5323 Harry Hines in the vertebrate olfactory bulb (Cinelli et al. 1995).
Blvd., Dallas, TX 75235-9111. E-mail: smith15@utsw.swmed.edu Therefore, there is likely to be a correlation between
Genetics 150: 711–721 (October 1998)712 M.-S. Kim, A. Repp and D. P. Smith
the odorant specificity of the olfactory neurons and the In(2LR)O, Cy p[tau-LacZ, w1]/1; y1 D2,3 Sb/1 males were
pattern of glomerular activity in both vertebrate and recovered and mated to 5 attached-X virgin females in individ-
ual vials. Single males carrying white1(w 1), but not the
Drosophila olfactory systems. n(2LR)O, Cy p[tau-LacZ, w1] or y1 D2,3 Sb chromosomes were
One family of proteins with potential to influence recovered and used to establish stable strains carrying novel
chemosensory discrimination is invertebrate odorant- P-element integrations on the X, second, or third chromo-
binding proteins (OBPs). OBPs are produced by verte- somes by crossing each male to 10 attached-X females. Only
brate and arthropod chemosensory systems where they one male was isolated from each vial to ensure independent
insertion events were recovered and screened for LacZ expres-
are secreted from nonneuronal support cells into the sion (see below).
fluid that bathes the olfactory neuron dendrites. Odor- lush mutants were generated by mobilizing the P element
ants have been shown to bind directly to these proteins from the ET249 stock (the line carrying the enhancer trap
in both mammals and insects (Vogt and Riddiford element with trichoid sensillum-specific LacZ expression) by
1981; Pelosi et al. 1982; Pevsner et al. 1985; Pevsner crossing to flies carrying a stable source of transposase (Rob-
ertson et al. 1988) and recovering third chromosomes that
et al. 1990; Du and Prestwich 1995). In insects, mem- had lost the w 1 gene over a balancer chromosome. We recov-
bers of the invertebrate OBP family are low-molecular- ered approximately 300 independent third chromosomes
weight, chemosensory-specific proteins with six con- missing the w1 eye color marker contained within the P ele-
served cysteine residues. Unlike vertebrate odorant- ment. Homozygous strains for most of these chromosomes
binding proteins that are members of the lipocalin were generated, and genomic DNA was prepared and
screened using the polymerase chain reaction with primers
transport family (Flower 1996), the invertebrate pro- specific to the lush coding sequence (see below).
teins constitute a unique protein family. In Drosophila, b-Galactosidase expression: Enhancer trap lines were
the six previously identified invertebrate OBP members screened for b-galactosidase expression in adult heads as pre-
have surprisingly low sequence similarity and are ex- viously described (Riesgo-Escovar et al. 1992), except that
pressed in different, overlapping zones of chemosen- staining reactions were performed at 258 for 4 hr. Lines with
LacZ expression restricted to the chemosensory structures
sory sensilla. This is consistent with these proteins per- of the head were retested to confirm the staining pattern, and
forming an odor-specific function (McKenna et al. 1994; LacZ expression was simultaneously examined in the body.
Pikielny et al. 1994). Moth pheromone-binding protein Larvae were stained as described Riesgo-Escovar et al.
members of this family have been shown to bind directly (1992). To examine LacZ expression in tissue sections, 10-mm-
to pheromone with chemical selectivity indicating mem- thick frozen sections were fixed for 10 min in 1% glutaralde-
hyde (EM Grade, EM Science), washed in PBS, and stained
bers of this family interact directly with odorant mole- as described above.
cules (Du and Prestwich 1995). No mutants defective DNA, RNA, sequencing and PCR: Genomic DNA flanking
for any odorant-binding protein gene have been pre- the P-element insertion was cloned by plasmid rescue as de-
viously described; therefore the in vivo function of these scribed by Pirrota (1986). Genomic DNA was prepared as
proteins is unknown. Possible functions include solubi- described by Lis (1983). Library screening, restriction map-
ping, and mRNA isolation were performed as described in
lizing or concentrating odorants in the sensillum lymph,
Maniatis et al. (1982). Appendage cDNA was prepared from
or mediating odorant removal (reviewed in Pelosi mRNA using a reverse transcription kit (Invitrogen, San Di-
1994). We report here the identification and character- ego, CA) using appendages isolated as described by Oliver
ization of lush, a gene encoding a new member of the (Oliver and Philips 1970). Hybridizations were performed
invertebrate odorant-binding protein family in Dro- at 658 in 750 mm NaCl, 100 mm NaH2PO4 (pH 6.8), 75 mm
sodium citrate, 0.4% Ficoll, and 0.5% sodium dodecyl sulfate.
sophila. LUSH is expressed in a subset of trichoid sen-
Filters were washed in 0.23 SSC (13 SSC is 150 mm NaCl,
silla and is required for normal olfactory behavior re- 15 mm sodium citrate) and 0.5% SDS at 658. Sequence analysis
sponses to a small subset of chemically related odorants. was performed using an ABI automated sequencer (ABI Adv.
Our results support models in which odorant-binding Biotechnologies, Columbia, MD). PCR reactions to identify
proteins participate in determining the chemical speci- lush mutants were performed using the method of Saiki et al.
ficity of olfactory neurons in Drosophila. (1985) with oligonucleotides 59 GAAGCTTGTAGGGATACG
and 59 TTAAGGCCACATGAACTG. PCR conditions were 948
for 30 sec, 508 for 30 sec, and 728 for 2 min, repeated for 35
MATERIALS AND METHODS cycles. Control primers specific to unlinked sequences were
included in each PCR reaction to control for presence of
Drosophila stocks, generation of enhancer trap lines, lush template DNA.
mutants: Flies carrying the Tau-LacZ P element were obtained In situ hybridization to polytene chromosomes and tissue
from John Thomas and Chris Callahan (Salk Institute). TAU sections: Polytene chromosomes were prepared from salivary
is a microtubule-binding protein that localizes the fusion pro- glands of late third instar larvae of the Oregon R wild-type
tein to axons when expressed in neurons (Callahan and strain and hybridized as described by Langer-Sofer et al.
Thomas 1994). Genetic crosses were carried out under stan- (1982). DNA fragments to be mapped were labeled with [bio-
dard laboratory conditions using balancer stocks (Lindsley 16]dUTP (Enzo Biochemicals) by nick translation. Signal de-
and Zimm 1992). After isogenic strains were created, z2500 tection was performed with streptavidin-conjugated horserad-
lines carrying novel insertions were generated as follows: y ish peroxidase (Enzo Biochemicals) and diaminobenzidine.
w; 1/1; y1 D2,3 Sb/TM2 males carrying the D2,3 activated Generation of antiserum, immunofluorescence, Western
transposase (Robertson et al. 1988) were crossed to w/w; blotting: Rabbit polyclonal antiserum was raised to a six histi-
In(2LR)O, Cy p[tau-LacZ, w1]/In2LR Gla; 1/1 virgin females. dine-tagged LUSH protein expressed in bacteria. Serum was
In(2LR)O, Cy and TM2 are balancer chromosomes. Single affinity purified on LUSH Affi-gel columns (Bio-Rad, Rich-LUSH Mediates Alcohol Responses 713
mond, CA) according to the instructions of the manufacturer. the LacZ gene is fused in frame to the gene encoding
Immunofluorescence was performed as described in (Smith the microtubule-associated protein tau, expression of
et al. 1991). Western Blots were performed as described in
Stamnes et al. (1991) except that antibodies were detected the reporter gene in neurons results in LacZ staining
using ECL kits (Amersham, Arlington Heights, IL). Forty an- of axonal projections when expressed in these cells
tennae equivalents were run per lane. Canton-S and w 1118 were (Callahan and Thomas 1994).
used as controls. We generated several thousand lines of flies carrying
P-element-mediated DNA transformations: Drosophila stable, independent P-element insertions. Members of
transformations were carried out as described by Karess and
Rubin (1984). Transposase DNA was used at a concentration each line were screened for reporter gene expression
of 200 mg/ml and sample DNA was used at 1 mg/ml. The restricted to the chemosensory structures. ET249 was
rescue fragment used to restore wild-type lush function ex- one of several lines with adult LacZ expression restricted
tended from the left end of l249 to the first BamHI site (see to a subset of chemosensory sensilla on the third anten-
Figure 2). nal segment (Figure 1A). Olfactory neuron axons, visi-
Olfactory behavioral assays: Isogenized w 1118 flies were the
parental background for all experiments. w 1118 or ET249 flies ble in other enhancer trap lines expressing tau-LacZ in
were chosen as olfactory normal controls for testing lush mu- olfactory neurons, were not stained in ET249 indicating
tants to minimize differences in genetic background that expression of tau-LacZ was restricted to support cells.
are well known to influence olfactory behavioral responses To more precisely identify the cells that were expressing
(Alcorta and Rubio 1988; Alcorta and Rubio 1989; Monte LacZ in the antennae of ET249 flies, we stained frozen
et al. 1989; Dubin et al. 1995). ET249 flies express LUSH
at normal levels indicating the P element does not disrupt tissue sections from these structures. LacZ expression
expression of this gene. was prominent in cells associated with trichoid sensilla
Olfactory trap assays were performed essentially as described on the ventral-lateral surface of the third antennal seg-
in (Woodard et al. 1989) except that 5 male and 5 female ment (Figure 1B). Based on their relative position in
flies were tested in each plate. No sex-specific differences in the epithelium, the support cells expressing LacZ in
olfactory behavior were observed in lush mutants. One- to
three-day-old flies were tested and a minimum of 100 flies ET249 flies are trichogen support cells that secrete the
(10 plates) were tested for each odorant concentration and sensillum lymph that bathes the olfactory neuron den-
genotype. A total of 10 ml of the diluted odorant was vortexed drites (Keil and Steinbrecht 1984; Hartenstein and
with 1 ml of 1% agarose at 458, and 100-ml aliquots were Posakony 1989; Ray and Rodrigues 1995). We exam-
distributed to 10 traps on ice to rapidly solidify the agarose. ined third-instar larvae for LacZ expression and found
The concentrations noted in the figures are the concentration
within the agarose. The actual odorant concentrations in air expression restricted to the larval olfactory organs, the
are significantly less. Odorants were obtained from Aldrich antennomaxillary complex (Figure 1C). We mapped
Chemical (Milwaukee, WI) and were the highest purity avail- the P-element insertion to position 76C on the third
able. Differences in the means were tested for significance chromosome (data not shown). No previously identified
using t -tests for independent samples and ANOVA was used olfactory mutants have been mapped to this genomic
for comparison of more than two means (Statistica Software;
StatSoft Inc., Tulsa, OK). region.
Electroantennograms: Extracellular recordings of electrical P-element excision mutants have abnormal olfactory
responses of the antenna were obtained essentially as de- behavior to a subset of odorants: To create loss-of-func-
scribed by Dubin et al. (1995) using an EX-1 single channel tion mutations in the putative chemosensory-specific
extracellular amplifier (Degan, Minneapolis, MN) and Mac- gene identified by the ET249 P element, we generated
Adios II hardware and Superscope software (GW Instruments,
Somerville, MA) with an automated odorant delivery system small deletions at the P-element integration site by mobi-
(Alcorta 1991) providing 1-sec odorant pulses. The record- lizing the transposon from ET249 flies and recovering
ing electrode was placed on the ventral lateral surface of the chromosomes from which excision had occurred (for
antenna, and the ground electrode was placed in the brain example see Sass et al. 1993). We identified five putative
through the vertex of the head. deletions based on the absence of a PCR product using
primers specific to DNA sequences flanking the P ele-
ment. The largest deletion eliminated 3 kb of genomic
RESULTS
DNA flanking the P element. This lesion completely
ET249: an enhancer trap line expressing LacZ exclu- removed the lush transcription unit (see below). Flies
sively in the chemosensory system: LUSH was identified homozygous for this deletion are viable and fertile and
using the enhancer detection approach (Bellen et al. were named lush mutants (see below).
1989) as a gene expressed exclusively in the olfactory To compare olfactory discrimination between lush
organs. Briefly, single P-transposable elements (P ele- and wild-type adults, we employed the olfactory trap
ments) modified to express a tau-LacZ fusion reporter assay (Woodard et al. 1989). Briefly, 10 wild-type or
gene (Callahan and Thomas 1994) were randomly mutant flies were placed in a petri plate with a single
inserted in the genome. Expression of the reporter fu- odorant trap, and the number of flies within the trap
sion gene is dependent on enhancer elements acquired was determined after a set time period (see materials
upon integration and can mimic the temporal and spa- and methods). We screened a panel of 60 simple vola-
tial expression pattern of individual genes located at or tile organic compounds at different concentrations to
near the integration site (Bellen et al. 1989). Because test for differences in distribution between control and714 M.-S. Kim, A. Repp and D. P. Smith
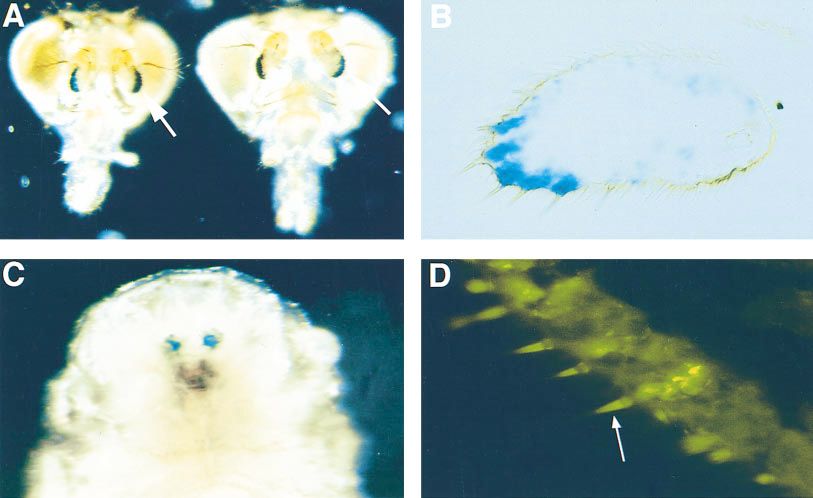
Figure 1.—Expression of
Tau-LacZ in ET249. (A)
Male (left) and female
ET249 adult heads were
stained for b-galactosidase
activity. LacZ expression is
visible on the ventral-lateral
surface of each third anten-
nal segment (arrows). No
staining is present in the
brain, thorax or abdomen,
wings or legs. No olfactory
neurons are stained. (B)
Frozen tissue section
through ET249 antenna
stained for b-galactosidase
activity. b-Galactosidase is
associated with support cells
in a subset of trichoid sen-
silla. (C) b-Galactosidase ac-
tivity is also present in the
larval olfactory organ in
ET249, the antennomaxil-
lary complex. (D) LUSH
protein is expressed in the same region as b-galactosidase in ET249 flies. Immunofluorescence image of frozen tissue section
of wild-type fly reacted with affinity-purified LUSH antiserum. Note secretion of LUSH protein into the sensillum lymph of the
trichoid sensilla (arrow).
lush flies. Table 1 shows results for representative odor- Wild-type flies have endogenous mechanisms to avoid
ants tested. Odorants were tested at 1:1000 and 1:4 dilu- concentrated ethanol that are defective in lush mutants:
tions in agarose. As expected from the restricted expres- The increased likelihood of lush mutant flies entering
sion pattern of LacZ in a subset of sensilla, the majority traps containing high concentrations of these alcohols
of the compounds attract similar proportions of wild- could result from either increased attraction to these
type and lush flies, indicating there is no global olfactory odorants or a defect in avoidance of high concentrations
defect associated with the deletion. However, odor-spe- of these compounds. If there is a defect in chemoavoi-
cific defects in chemosensory behavior are observed in dance to ethanol in lush mutants, we should be able to
lush flies when challenged with three chemically related demonstrate this behavioral response in wild-type flies.
odors. We observe a significant increase in the number To determine if wild-type flies have endogenous mecha-
of mutant flies in traps containing high concentrations nisms to avoid high-ethanol environments that are de-
of ethanol, propanol, and butanol compared to control fective in the mutants, we tested the effects of mixing
flies. Their responses to a variety of other alcohols are ethanol with yeast extract, a strong chemoattractant.
not different from those of wild type (Figure 2A). Inter- Figure 2C shows that wild-type flies are attracted to di-
estingly, the apparent increased attraction of lush flies lute yeast extract (left panel, open bars). However, when
for ethanol, propanol, and butanol is specific to high- the same amount of yeast is mixed with concentrated
odorant concentrations. Figure 2B reveals the dose- ethanol, wild-type flies are significantly less likely to en-
dependent, abnormal attraction of lush mutants for eth- ter these traps (compare open bars). Therefore, the
anol. The extent of the attraction of lush flies to yeast presence of high levels of ethanol reduces attraction
extract, ethyl acetate, and low concentrations of ethanol for yeast in wild-type flies. This demonstrates that there
is similar to that of wild-type. However, the mutant flies is an active avoidance mechanism in wild-type flies that
display an abnormal attraction to traps containing high is stimulated by high concentrations of ethanol. lush
concentrations of ethanol (1:100, 1:4; Figure 3B). mutants are equally attracted to yeast compared to wild-
ET249 flies (that carry the P element but not the dele- type flies (filled bar, left graph) but are defective for
tion) and the w1118 strain from which these lines were the avoidance behavioral response (Figure 2C, filled
derived have normal chemosensory responses to these bars). In fact the lush mutants are significantly more
alcohols, as do third-instar larvae from lush flies (data attracted to the mixture of yeast and concentrated etha-
not shown). We named this deletion mutant “lush” to nol than to yeast alone.
reflect their increased affinity for ethanol-rich environ- One model that could explain the increased affinity
ments. We conclude that lush flies have odor-specific of lush mutants for high concentrations of alcohol is a
defects in chemosensory discrimination and are abnor- specific defect in active avoidance behavior to ethanol
mally attracted to high concentrations of a subset of mediated through the lush gene product. In an alterna-
odorants including ethanol, propanol, and butanol. tive model, the same phenotype could arise if the lushLUSH Mediates Alcohol Responses 715
TABLE 1
Olfactory behavioral responses of lush and control flies (w1118) to a variety of odorants
Odorant w 1118 lush P valueb
Ethanol
1:1000 1.1 6 0.45 (150) 0.8 6 0.32 0.6
1:4a 2.6 6 0.55 (150) 5.6 6 0.48 0.0002a
Propanol
1:1000 2.7 6 0.76 (100) 2.5 6 0.68 0.85
1:4a 0.1 6 0.1 (100) 1.2 6 0.25 0.0007a
Butanone
1:1000 1.9 6 0.34 (100) 1.1 6 0.31 0.11
1:4 1.8 6 0.47 (100) 1.8 6 0.34 1.0
Acetone
1:1000 2.3 6 0.57 (100) 2.2 6 0.46 0.89
1:4 2.5 6 0.58 (100) 3.7 6 0.76 0.23
Ethyl acetate
1:1000 2.5 6 0.5 (100) 1.9 6 0.5 0.41
1:4 2.5 6 0.61 (100) 2.1 6 0.58 0.64
Isoamyl acetate
1:1000 3.2 6 0.42 (100) 2.7 6 0.37 0.38
1:4 0.4 6 0.22 (100) 0.5 6 0.22 0.75
Acetic acid
1:1000 4.6 6 0.6 (100) 5.3 6 0.7 0.89
1:4 1.1 6 0.3 (100) 0.8 6 0.2 0.23
Benzaldehyde
1:1000 3.0 6 0.73 (100) 2.7 6 0.36 0.73
1:4 0.09 6 0.09 (100) 0.0 6 0.0 0.94
Yeast
1:100 5.0 6 0.6 (100) 4.7 6 0.8 0.70
Values are mean number of flies (out of 10 possible) attracted to odorant traps. Parentheses denote total
flies tested.
a
Significant difference between genotypes (two-tailed t-test, independent samples).
b
Probability that the difference between the means for the two genotypes is the same by chance.
gene product were required to deactivate or desensitize P-element insertion site was cloned by plasmid rescue
neurons mediating chemoattraction. If this latter model (Pirrotta 1986). We recovered 2.5 kb of genomic DNA
is correct, lush mutants should have a delay in termina- flanking the P-element insertion site and used these
tion of the ethanol-induced electrical responses com- sequences to isolate genomic and cDNA clones (Figure
pared to control flies. In an attempt to distinguish be- 4). Two transcription units were mapped to the region
tween these models, we recorded electroantennograms of the P-element integration site, one of which mimics
(EAG) using a computer-triggered odorant delivery sys- the expression profile of LacZ in ET249 and is specifi-
tem (Alcorta 1991) to analyze the electrical responses cally deleted in the mutant (see below).
of the ethanol-sensitive olfactory neurons in wild-type We determined the entire nucleotide sequence of a
and lush mutant flies. We observed no significant differ- putative lush cDNA and z3 kb of genomic DNA flanking
ence between wild-type and lush mutants in amplitude the P-element insertion site. The P element inserted
or the time required to deactivate the response to 75% 373 base pairs downstream from the polyadenylation
of baseline over a wide range of ethanol concentrations site of the lush transcription unit (Figure 4) and did
(see Figure 3). These results argue against a defect in not disrupt expression of this gene (data not shown).
adaptation or deactivation in neurons mediating attrac- This is consistent with the observation that ET249 flies
tion that would produce prolonged ethanol-induced avoid concentrated ethanol (Figure 2). The predicted
electrical responses in lush mutants. protein encoded by the lush gene is 153 residues in
Deletion mutants are missing the lush gene: a new length with a series of hydrophobic residues near the
member of the invertebrate odorant-binding protein N terminus typical of a signal sequence (von Heijne
family: We characterized the genomic DNA at the 1986). Database comparison with previously identified
P-element insertion site in ET249 flies to define the proteins revealed significant homology (24% overall
gene responsible for the abnormal chemosensory re- identity) with OS-F/PB-PRP3 (McKenna et al. 1994;
sponses in lush mutant flies. Genomic DNA was pre- Pikielny et al. 1994), a Drosophila member of the inver-
pared from ET249 flies, and the DNA flanking the tebrate odorant-binding protein family (Figure 5). LUSH716 M.-S. Kim, A. Repp and D. P. Smith
Figure 2.—lush mutant flies have ab-
normal behavioral responses to ethanol,
propanol, and butanol in olfactory trap
assays. (A) Olfactory trap data compar-
ing ET249 (control) and lush mutant re-
sponses to short-chain alcohols. Bars
represent average number of flies enter-
ing traps containing the substances
noted. Averages represent a minimum
of 10 experiments. Standard errors are
depicted above the bars. Asterisk de-
notes statistical significance between the
means of control (ET249) and lush mu-
tant flies (two-tailed t -test, independent
samples). Concentrations noted are the
initial concentration of odorant mixed
in agarose. lush mutants have normal re-
sponses to methanol and isopropanol,
but are more likely to enter traps con-
taining concentrated propanol than are
control flies. (B) lush mutant flies are
more likely to enter traps containing
high levels of ethanol in a dose-depen-
dent manner. The sensitivity of lush mu-
tants for ethanol is not significantly dif-
ferent from controls at low concen-
trations (1:500). Wild-type responses are
restored to the mutants by the lush
transgene (hatched bars). (C) lush mu-
tants are defective for chemoavoidance
of concentrated ethanol. Wild-type and
lush flies are equally attracted to traps
containing 1% yeast extract. Wild-type and lush mutants expressing a transgenic lush gene (rescue) are significantly less likely
to enter traps containing the same amount of yeast when it is mixed with 25% ethanol (P , 0.001, t -test, independent samples).
lush mutants are defective for this avoidance response, are significantly more likely to enter the traps containing the mixture
than are the controls, and are significantly more likely to enter the traps containing the mixture than traps containing yeast
alone (P , 0.001). Asterisk denotes significant differences between yeast and yeast 1 ethanol for each genotype.
shares all features of this protein family including a firmed that the 3-kb deletion removes the entire protein-
signal sequence to direct polypeptides to the secretory coding region of the lush gene (Figure 4, lower panel).
pathway, chemosensory-specific expression pattern, and This suggests that loss of this odorant-binding protein
six conserved cysteine residues with the spacing between gene is responsible for the chemosensory defects in the
cysteines 2 and 3 and 5 and 6 completely conserved in lush mutants.
all members. To prove the chemosensory defects associated with
Rabbit polyclonal antiserum was raised to bacterially lush mutants are due entirely and specifically to loss of
expressed LUSH protein for direct examination of the LUSH protein, we introduced a cloned wild-type copy
expression of this protein in wild-type and lush flies (see of this gene into the mutant flies by germ-line transfor-
materials and methods). Affinity-purified anti-LUSH mation (see materials and methods). Expression of
antibodies recognize protein in accessory cells of tri- a lush transgene under control of its own promoter in
choid sensilla on the ventral-lateral portion of the third- the mutant background restores LUSH expression to
antennal segment in wild-type males and females, in a normal levels (Figure 6, rescue). Furthermore, the
pattern identical to LacZ expression in ET249 (Figure transgene completely restores wild-type olfactory behav-
1D). In contrast to the LacZ that is localized to the ioral responses to the lush mutants (Figure 2, B and C,
support cell cytoplasm in ET249 flies, LUSH protein was striped bars). Therefore, the abnormal chemoattraction
clearly present within the shafts of the trichoid sensilla, to high levels of alcohol associated with the deletion
confirming it was secreted into the sensillum lymph results specifically from loss of LUSH. Flies carrying six
(compare Figure 1B and Figure 1D). No labeling of lush genes overexpress LUSH in the trichoid sensilla
olfactory neurons was observed. Western blots of anten- (see Figure 6) and behave indistinguishably from con-
nal extracts from wild-type and lush mutant flies probed trols in response to ethanol (data not shown) suggesting
with anti-LUSH antiserum revealed that the mutants are that the levels of LUSH are not the rate-limiting compo-
completely defective for LUSH expression (Figure 6, nent of this behavior. We conclude that lush mutants
LUSH). Southern blot analysis of lush mutant DNA con- have defective chemosensory responses to a subset ofLUSH Mediates Alcohol Responses 717
Figure 3.—lush mutants deactivate ethanol-sensitive neu-
rons normally. lush mutants and control flies terminate etha-
nol-induced olfactory responses with similar time courses. The
time to deactivate the response from peak to 75% return-to-
baseline was determined for lush mutants, wild-type controls Figure 4.—Map of 76C genomic region. The ET249 P ele-
(ET249), and lush mutants carrying two wild-type copies of ment integrated into the genomic region corresponding to
the lush gene (rescue). Recordings from extracellular re- l249. In ET249 flies, the P element integrated into the R1-RV
sponses were measured and averaged. SEM is depicted by fragment. The lower panel depicts the restriction fragments
the error bars. Responses from a minimum of five flies were completely (2) or partially (1/2) deleted in lush mutants.
measured for each genotype for each odor. There is no sig- The genomic structure of the lush cDNA is shown below the
nificant difference in time required to deactivate the ethanol l249 map with an expanded view of the lush locus. The posi-
response in lush and control flies. EA, ethyl acetate. tion of the ET249 P element, the lush gene, and the ash-1
gene (Tripoulas et al. 1994) are depicted above l249. All
cloned sequences were confirmed to map to position 76C on
odorants resulting from loss of a single odorant-binding the polytene chromosome.
protein in the sensillum lymph of a small subset of
trichoid chemosensory sensilla.
edge, lush is the first odorant-binding protein mutant
described for any organism. The specific olfactory de-
DISCUSSION
fects associated with the lush mutant provide the first
LUSH has the hallmark features of a member of the direct evidence that a member of this protein family is
invertebrate odorant-binding protein family. These fea- required for normal olfactory behavior.
tures include chemosensory-specific expression, the Specific features of the arthropod chemosensory sys-
presence of a signal sequence for secretion from the tem not present in mammalian systems may allow OBPs
nonneuronal support cells in which it is expressed, and to play a unique role in chemosensory discrimination.
the presence of six cysteine residues with conserved Unlike mammals, whose olfactory cilia are bathed in a
spacing, especially between cysteines 2 and 3 and 5 and common overlying fluid, most arthropods (including
6 where the spacing is absolutely conserved in all mem- insects) have compartmentalized their olfactory neu-
bers. The conservation of these cysteines suggests that rons into sensilla. Segregation of individual or small
the members of this family share a common disulfide groups of olfactory neurons in separate compartments
bonding pattern that may impart a similar tertiary struc- provides the opportunity to independently regulate the
ture (Raming et al. 1990). composition of the fluid bathing the olfactory neuron
We have shown that LUSH is secreted into the sensil- dendrites. Indeed, the seven Drosophila members iden-
lum lymph (Figure 1D). Electron microscopy studies tified to date are expressed in specific subsets of sensilla,
have previously demonstrated secretion of moth phero- and none are expressed in all sensilla (McKenna et al.
mone-binding proteins and the Drosophila OS-E and 1994; Pikielny et al. 1994; Hekmat-Scafe et al. 1997).
OS-F into the sensillum lymph of the sensilla in which More than one odorant-binding protein can be ex-
they are expressed (Steinbrecht 1996; Hekmat-Scafe pressed within a single sensillum (Hekmat-Scafe et al.
et al. 1997). It is likely, therefore, that all members of 1997). Differential expression of a family of odorant-
this family function in the sensillum lymph. Six other binding proteins, therefore, is a feasible mechanism
members of the invertebrate OBP family have been for influencing the chemical specificity of the olfactory
identified in Drosophila through the use of differential neurons within those sensilla, perhaps by regulating
screening methods (McKenna et al. 1994; Pikielny et access of odorants to the neuron. The defective olfactory
al. 1994). However, there are no corresponding mutants behavior associated with the lush mutant is consistent
that specifically disrupt these genes to provide insight with this idea.
into the in vivo function of these proteins. To our knowl- How does a protein secreted into the fluid that bathes718 M.-S. Kim, A. Repp and D. P. Smith
Figure 5.—Alignment of
LUSH with other members
of the invertebrate odor-
ant-binding protein family.
LUSH aligned with four
moth and four Drosophila
members of the inverte-
brate odorant-binding pro-
tein family. Conserved cys-
teines are denoted with an
asterisk above the align-
ment. Lush, LUSH; PB-
PRP1, Drosophila PBP re-
lated protein 1 (Pikielny
et al. 1994); PB-PRP2, Dro-
sophila PBP related protein
2 (Pikielny et al. 1994); PB-
PRP3, Drosophila PBP re-
lated protein 3 (McKenna
et al. 1994; Pikielny et al.
1994); PB-PRP5, Drosoph-
ila PBP related protein
5 (Pikielny et al. 1994);
PBP-1, pheromone-bind-
ing protein from moth A.
polyphemus (Raming et al.
1989); PBP-2, pheromone-
binding protein from Man-
duca sexta (Gyorgyi et al.
1988); PBP-6 and PBP-8,
general odorant-binding
proteins from Manduca
sexta (Vogt et al. 1991).
olfactory neuron dendrites (but is not synthesized by known, our data provide important clues about the in
olfactory neurons) affect chemosensory behavior? First, vivo role of these proteins. First, ethanol is very soluble
the olfactory defects observed in lush mutants do not in both aqueous and lipid environments. It is unlikely,
arise from the loss or global disruption of the function of therefore, that LUSH functions simply to solubilize
the support cells that secrete LUSH. These cells appear these alcohols in the sensillum lymph. Furthermore, we
morphologically normal in the mutants and are able to think it is unlikely that LUSH simply removes these
secrete other members of the invertebrate OBP family odorants from the lymph. If LUSH removed ethanol
normally (M.-S. Kim and D. P. Smith, unpublished re- from the sensillum lymph, and the olfactory neurons
sults). Therefore, LUSH is not required for the pres- in the trichoid sensilla mediated attraction, increased
ence, determination of cell fate, or functioning of the attraction could result from the persistence of these
support cells in which it is expressed. Similarly, LUSH is alcohols in the lymph. However, if odorant removal is
not expressed in the antennal lobe, the central nervous the sole function of LUSH, we would expect to find
system, or the motor pathways, indicating it does not increased sensitivity to alcohol in lush mutants because
mediate chemosensory information processing or effer- they would have increased alcohol levels in the sensillum
ent behavioral responses to odorants. This narrows the lymph compared to normal flies at low alcohol concen-
site of action of LUSH to effects on the primary olfactory trations. lush mutants have normal sensitivity to low lev-
neurons. els of ethanol (Figure 2B). Finally, LUSH could act by
Given their location in the sensillum lymph, the fact desensitizing the trichoid olfactory neurons mediating
that moth pheromone-binding proteins bind directly to attraction to alcohol, perhaps by acting as a neuronal
pheromone odorant (Du and Prestwich 1995), and receptor antagonist when bound to alcohol. However, in
that lush mutants have odor-specific defects in olfactory the absence of LUSH this model predicts that olfactory
behavior, LUSH probably modulates the activity of pri- neurons will be active over a longer time course than
mary olfactory neurons in the trichoid sensilla through wild-type controls, and we do not observe this in extra-
a mechanism involving a direct interaction with odor- cellular recordings from the antenna (Figure 3). How-
ants in the sensillum lymph. For example, LUSH could ever, LUSH-dependent olfactory neurons may be a small
regulate odorant concentration, transport, or metabo- fraction of the ethanol-sensitive neurons, and their con-
lism within the lymph. While the exact mechanism by tribution to the EAG may not be detectable.
which LUSH affects chemosensory behavior is not We think the most likely possibility is that LUSH isLUSH Mediates Alcohol Responses 719
We have shown that LUSH is required for normal che-
mosensory responses to specific odorants. This implies
a correlation between the odorant-binding protein ex-
pression zone and the odor specificity of olfactory neu-
rons. Previous workers have demonstrated a relation-
ship between odorant sensitivity and position on the
surface of the antenna (Siddiqi 1987; Ayer and Carl-
son 1992; Dubin et al. 1995) and these zones of sensitiv-
ity could correspond to odorant-binding protein expres-
sion zones. Cobalt backfilling experiments labeling the
projections of the olfactory neurons from the LUSH
expression zone (the ventral-lateral surface) revealed
these olfactory neurons synapse primarily in only 2 of
the 35 anatomically identified glomeruli in the antennal
lobe, VA-1 and DA-1 (Stocker et al. 1983). It will be
interesting to determine if one or both of these glomer-
uli specifically function in chemosensory avoidance, and
if neurons associated with other odorant-binding pro-
Figure 6.—Expression of LUSH protein in wild-type flies,
tein zones project to common subsets of glomeruli. The
deletion mutants, and deletion mutants transformed with a
wild-type lush gene. Anti-LUSH antiserum recognizes a 14-kD lush expression zone overlaps several other Drosophila
LUSH protein in control antennae (Canton S and w1118) that odorant-binding proteins, specifically PB-PRP-1 and PB-
is absent in the lush mutants (LUSH). Mutants transformed PRP-3/OS-F (McKenna et al. 1994; Pikielny et al. 1994).
with two copies of the wild-type lush gene are restored for When mutants defective for these gene products be-
LUSH expression (Rescue). LUSH is overexpressed in flies
come available, it will be important to determine if they
carrying six copies of lush (3xRescue).
have defective avoidance responses, but to a different
subset of odorants. Similarly, we predict mutations in
required to activate a small subset of olfactory neurons OBPs expressed in the other classes of sensilla will have
in the trichoid sensilla that specifically mediate chemo- defective attraction to a subset of odorants.
avoidance. Olfactory neurons specific for chemoavoid- Our data implicate members of the invertebrate odor-
ance are well documented in Caenorhabditis elegans ant-binding protein family in odorant discrimination in
(Bargmann and Horvitz 1991; Bargmann et al. 1993; Drosophila. However, neuronal receptors are also likely
Troemel et al. 1997). For example, LUSH might con- to contribute to chemosensory discrimination in vivo.
centrate or prevent the rapid metabolism of these alco- Seven transmembrane receptors mediate odorant re-
hols in the sensillum lymph thus increasing the steady- sponses in C. elegans (Sengupta et al. 1996) and in
state concentration of these odorants in the trichoid vertebrates (Buck and Axel 1991; Zhou et al. 1998).
sensillum lymph of wild-type flies. This could trigger In Drosophila the dGqa-3 heterotrimeric G protein
activation of olfactory neurons mediating avoidance and a-subunit is expressed in the dendritic portion of a sub-
altering the perception of ethanol so it “smells bad.” set of olfactory neurons, consistent with a role in trans-
Pheromone-binding proteins may perform a similar role ducing a subset of odorant responses through seven
in sensitizing chemosensory neurons to pheromone in transmembrane receptors (Talluri and Smith 1995).
moths (Vogt et al. 1985; Kaissling 1997). If true, this Furthermore, insects often package several olfactory
model predicts that these LUSH-dependent olfactory neurons within a single sensillum, and there is evidence
neurons would not be activated by ethanol in lush mu- to suggest these neurons are not functionally identical.
tants, but would be activated in wild-type flies. However, Analysis of the electrical responses of Antherea polyphemus
we see no significant differences in the EAG recordings, moths revealed that two neurons in the same phero-
again perhaps because these neurons are not detectable. mone-sensitive sensillum responded preferentially ei-
Alternatively, LUSH may affect olfactory behavior by ther to the pheromone acetate or to the aldehyde (Gan-
regulating processes that occur on a slower time scale jian et al. 1978). This suggests that the olfactory neurons
apparent in chemosensory behavior assays, but not EAG within a sensillum are not functionally identical, and
recordings. Additional experiments will be required to these differences probably correspond to differential
identify the exact biochemical function of the LUSH expression of receptor proteins on the dendritic surface
protein and the behavioral specificity of the chemosen- of the neuron. Therefore, we suggest that chemical spec-
sory neurons within the trichoid sensilla. ificity of olfactory neurons in Drosophila results from a
Each of the seven members of the Drosophila odor- combination of interaction of odorants with odorant-
ant-binding protein family are expressed in specific binding proteins in the sensillum lymph and the speci-
zones on the surface of the antenna. Therefore, there ficity of receptor proteins present on the olfactory neu-
is a topographic map on the surface of the antenna rons. A diverse family of odorant-binding proteins could
defined by zones of odorant-binding protein expression. enable a relatively small family of neuronal recep-720 M.-S. Kim, A. Repp and D. P. Smith
tors to respond differentially to a broad range of com- Buck, L., and R. Axel, 1991 A novel multigene family may encode
odorant receptors: a molecular basis for odor recognition. Cell
pounds through a combinatorial mechanism. For exam- 65: 175–187.
ple, two neurons in different sensilla expressing the Callahan, C. A., and J. B. Thomas, 1994 Tau-beta-galactosidase,
same broadly tuned neuronal receptor might be acti- an axon-targeted fusion protein. Proc. Natl. Acad. Sci. USA 91:
5972–5976.
vated by different subsets of odorants if each sensillum Chakir, M., O. Peridy, P. Capy, E. Pla and J. R. David, 1993 Adapta-
expressed a different repertoire of binding proteins. tion to alcoholic fermentation in Drosophila: a parallel selection
Elucidation of the relative size of the receptor and bind- imposed by environmental ethanol and acetic acid. Proc. Natl.
Acad. Sci. USA 90: 3621–3625.
ing protein families and determination of their spatial Cinelli, A. R., K. A. Hamilton and J. S. Kauer, 1995 Salamander
expression relationships will provide further insight into olfactory bulb neuronal activity observed by video rate, voltage-
the question of insect chemoreception. sensitive dye imaging. III. Spatial and temporal properties of
responses evoked by odorant stimulation. J. Neurophysiol. 73:
Finally, it should be noted that the alcohols that in- 2053–2071.
duce abnormal chemosensory reponses in lush mutants Davis, R., J. Cherry, B. Dauwalder, P. Han and E. Skoulakis,
are biologically relevant odorants for Drosophila melano- 1995 The cyclic AMP system and Drosophila learning. Mol. Cell.
Biochem. 149: 271–278.
gaster. In nature, fruit flies feed and deposit eggs on Du, G., and G. D. Prestwich, 1995 Protein structure encodes the
fermenting plant materials in which ethanol is the ligand binding specificity in pheromone binding proteins. J. Am.
most abundant short-chain alcohol (McKechnie and Chem. Soc. 34: 8726–8732.
Dubin, A. E., N. L. Heald, B. Cleveland, J. R. Carlson and G. L.
Morgan 1982; Van Delden 1982). The ability to detect Harris, 1995 Scutoid mutation of Drosophila melanogaster spe-
ethanol is important for chemotaxis toward food cifically decreases olfactory responses to short chain acetate esters
sources. However, adult flies are also susceptible to in- and ketones. J. Neurobiol. 28: 214–233.
Flower, D. R., 1996 The lipocalin protein family—structure and
toxication and death in high ethanol environments function. Biochem. J. 318: 1–14.
(Chakir et al. 1993). Therefore, there is a selective ad- Ganjian, I., M. J. Pettei, K. Nakanishi and K.-E. Kaissling, 1978
vantage for the ability to avoid environments with dan- A photoaffinity-labelled insect sex pheromone for the moth An-
theraea polyphemus. Nature 271: 157–158.
gerously high alcohol concentrations, and LUSH is re- Gyorgyi, T. K., A. J. Roby-Shemkovitz and M. R. Lerner, 1988
quired for this response. The behavioral response to Characterization and cDNA cloning of the pheromone-binding
alcohol, therefore, reflects a finely tuned olfactory re- protein from the tobacco hornworm, Manduca sexta: a tissue-
specific developmentally regulated protein. Proc. Natl. Acad. Sci.
sponse. USA 85: 9851–9855.
We thank Lisa Montgomery for excellent technical assistance, Ro- Hall, J. C., 1994 The mating of a fly. Science 264: 1702–1714.
Halpern, M., 1987 The organization and function of the vomerona-
mayleh Behrouzi for EAGs, Al Gilman and Leon Avery for helpful
sal system. Annu. Rev. Neurosci. 10: 325–362.
discussions, Helmut Kramer for use of injection facilities, and Charles
Hartenstein, V., and J. W. Posakony, 1989 Development of adult
Zuker, Leon Avery, and Steve Wasserman for helpful comments on sensilla on the wing and notum of Drosophila melanogaster. Devel-
the manuscript. This work was supported by the National Institutes opment 107: 389–405.
of Health grant DC-02539-2. Sequence of the lush gene is available Hekmat-Scafe, D., R. Steinbrecht and J. R. Carlson, 1997 Coex-
in GenBank accession no. AF001621. pression of two odorant binding homologs in Drosophila: implica-
tions for olfactory coding. J. Neurosci. 17: 1616–1624.
Kaissling, K.-E., 1998 Olfactory transduction in moths. I. Genera-
tion of receptor potentials and nerve impulses, II. Extracellular
LITERATURE CITED transport, deactivation and degradation of stimulus molecules,
pp. 93–112 in From Structure to Information in Sensory Systems, edited
Alcorta, E., 1991 Characterization of the electroantennogram in by C. Taddei-Ferretti and C. Musio. World Scientific, Singa-
Drosophila melanogaster and its use for identifying olfactory capture pore/New Jersey/London/Hong Kong.
and transduction mutants. J. Neurophysiol. 65: 702–714. Karess, R. E., and G. M. Rubin, 1984 Analysis of P transposable
Alcorta, E., and J. Rubio, 1988 Genetical analysis of intrapopula- element functions in Drosophila. Cell 38: 135–146.
tional variation in olfactory responses in Drosophila melanogaster. Keil, T. A., and R. A. Steinbrecht, 1984 Mechanosensitive and
Heredity 60: 7–14. olfactory sensilla of insects, pp. 477–516 in Insect Ultrastructure,
Alcorta, E., and J. Rubio, 1989 Intrapopulational variation of olfac- edited by R. C. King and H. Akai. Plenum Press, New York.
tory responses in Drosophila melanogaster. Behav. Genet. 19: 285– Langer-Sofer, P. R., M. Levine and D. C. Ward, 1982 Immunologi-
299. cal method for mapping genes on Drosophila polytene chromo-
Altner, H., and L. Prillinger, 1980 Ultrastructure of invertebrate somes. Proc. Natl. Acad. Sci. USA 79: 4381–4385.
chemo-, thermo-, and hygroreceptors and its functional signifi- Lindsley, D. L., and G. Zimm, 1992 The genome of Drosophila mela-
cance. Int. Rev. Cytol. 67: 69–139. nogaster. Academic Press, San Diego.
Ayer, R. J., and J. Carlson, 1992 Olfactory physiology in the Dro- Lis, J. T., J. A. Simon and C. A. Sutton, 1983 New heat shock puffs
sophila antenna and maxillary palp: acj6 distinguishes two classes and beta-galactosidase activity resulting from transformation of
of odorant pathways. J. Neurobiol. 23: 965–982. Drosophila with an hsp70-LacZ hybrid gene. Cell 35: 403–410.
Bargmann, C. I., and H. R. Horvitz, 1991 Chemosensory neurons Maniatis, T., E. F. Fritsch and J. Sambrook, 1982 Molecular Clon-
with overlapping functions direct chemotaxis to multiple chemi- ing: A Laboratory Manual. Cold Spring Harbor Laboratory Press,
cals in C. elegans. Neuron 7: 729–742. Cold Spring Harbor, NY.
Bargmann, C. I., J. H. Thomas and H. R. Horvitz, 1990 Chemosen- McKechnie, S. W., and P. Morgan, 1982 Alcohol dehydrogenase
sory cell function in the behavior and development of Caenorhab- polymorphism of Drosophila melanogaster: aspects of alcohol and
ditis elegans. Cold Spring Harbor Symp. Quant. Biol. 55: 529–538. temperature variation in the larval environment. Aust. J. Biol.
Bargmann, C. I., E. Hartweig and H. R. Horvitz, 1993 Odorant- Sci. 35: 85–93.
selective genes and neurons mediate olfaction in C. elegans. Cell McKenna, M. P., D. S. Hekmat-Scafe, P. Gaines and J. R. Carlson,
74: 515–527. 1994 Putative pheromone-binding proteins expressed in a sub-
Bellen, H. J., C. J. O’Kane, C. Wilson, U. Grossniklaus, R. K. region of the olfactory system. J. Biol. Chem. 269: 1–8.
Pearson et al., 1989 P-element-mediated enhancer detection: Monte, P., C. Woodard, R. Ayer, M. Lilly, H. Sun et al., 1989
a versatile method to study development in Drosophila. Genes Characterization of the larval olfactory response in Drosophila
Dev. 3: 1288–1300. and its genetic basis. Behav. Genet. 19: 267–283.LUSH Mediates Alcohol Responses 721
Oliver, D. V., and J. P. Philips, 1970 Technical note. Dros. Info. Shepherd, G. M., and C. A. Greer, 1990 Olfactory bulb, pp. 133–169
Serv. 45: 58. in The Synaptic Organization of the Brain, edited by G. M. Shepherd.
Pelosi, P., 1994 Odorant-binding proteins. Crit. Rev. Biochem. Mol. Oxford University Press, New York.
Biol. 29: 199–228. Siddiqi, O., 1987 Neurogenetics of olfaction in Drosophila melanogas-
Pelosi, P., N. E. Baldaccini and A. M. Pisanelli, 1982 Identifica- ter. Trends Neurosci. 3: 137–142.
tion of a specific olfactory receptor for 2-isobutyl-3-methoxypyra- Smith, D. P., 1996 Olfactory mechanisms in Drosophila melanogaster.
zine. Biochem. J. 201: 245–248. Curr. Opin. Neurobiol. 6: 500–505.
Pevsner, J., R. R. Trifiletti, S. M. Strittmater and S. H. Snyder, Smith, D. P., R. Ranganathan, R. W. Hardy, J. Marx, T. Tsuchida
1985 Isolation and characterization of an olfactory protein for et al., 1991 Photoreceptor deactivation and retinal degeneration
odorant pyrazines. Proc. Natl. Acad. Sci. USA 82: 3050. mediated by a photoreceptor-specific protein kinase C. Science
Pevsner, J., V. Hou, A. M. Snowman and S. H. Snyder, 1990 Odor- 254: 1478–1484.
ant-binding protein. Characterization of ligand binding. J. Biol. Stamnes, M. A., B.-H. Shieh, L. Chuman, G. L. Harris and C. S.
Chem. 265: 6118–6125. Zuker, 1991 The cyclophilin homolog ninaA is a tissue-specific
Pfeiffer, C. A., and R. E. Johnston, 1994 Hormonal and behavioral integral membrane protein required for the proper synthesis of
responses of male hamsters to females and female odors: roles a subset of Drosophila rhodopsins. Cell 65: 219–227.
of olfaction, the vomeronasal system, and sexual experience. Phys- Steinbrecht, R. A., 1969 Comparative morphology of olfactory re-
iol. Behav. 55: 129–138. ceptors, pp. 3–21 in Olfaction and Taste III, edited by C. Pfaffman.
Pikielny, C. W., G. Hasan, F. Rouyer and M. Rosbash, 1994 Mem- Rockefeller University Press, New York.
bers of a family of Drosophila putative odorant-binding proteins Steinbrecht, R. A., 1996 Are odorant binding proteins involved in
are expressed in different subsets of olfactory hairs. Neuron 12: odorant discrimination? Chem. Senses 21: 719–727.
35–49. Stocker, R. F., 1994 The organization of the chemosensory system
Pirrotta, V., 1986 Cloning Drosophila genes, pp. 83–110 in Drosoph- in Drosophila melanogaster: a review. Cell Tissue Res. 275: 3–26.
ila: A Practical Approach, edited by D. B. Roberts. IRL Press, Stocker, R. F., R. N. Singh, M. Schorderet and O. Siddiqi, 1983
Oxford. Projection patterns of different types of antennal sensilla in the
Raming, K., J. Kreiger and H. Breer, 1989 Molecular cloning of antennal glomeruli of Drosophila melanogaster. Cell Tissue Res.
an insect pheromone-binding protein. FEBS Lett. 256: 215–218. 232: 237–248.
Raming, K., J. Krieger and H. Breer, 1990 Primary structure of a Talluri, S., and D. P. Smith, 1995 Identification of a Drosophila
pheromone-binding protein from Antheraea pernyi: homologies G protein a subunit (dGqa-3) expressed in chemosensory cells
with other ligand carrying proteins. J. Comp. Physiol. B 160: and in central neurons. Proc. Natl. Acad. Sci. USA 92: 11475–
503–509. 11479.
Ray, K., and V. Rodrigues, 1995 Cellular events during develop- Tripoulas, N. A., E. Hersperger, D. La Jeunesse and A. Shearn,
ment of the olfactory sense organs in Drosophila melanogaster. Dev. 1994 Molecular genetic analysis of the Drosophila melanogaster
Biol. 167: 426–438. gene absent, small or homeotic discs1 (ash1). Genetics 137: 1027–
Riesgo-Escovar, J., C. Woodward, P. Gaines and J. Carlson, 1992 1038.
Development and organization of the Drosophila olfactory Troemel, E. R., B. E. Kimmel and C. I. Bargmann, 1997 Reprogram-
system: an analysis using enhancer traps. J. Neurobiol. 23: 947– ming chemotaxis responses: Sensory neurons define olfactory
964. preferences in C. elegans. Cell 91: 161–169.
Riesgo-Escovar, J. R., W. B. Piekos and J. R. Carlson, 1997 The Van Delden, W., 1982 The alcohol dehydrogenase polymorphism
Drosophila antenna: ultrastructural and physiological studies in in Drosophila melanogaster: Selection at an enzyme locus. Evol. Biol.
wildtype and lozenge mutants. J. Comp. Physiol. A 180: 151–160. 15: 187–222.
Robertson, H. M., C. R. Preston, R. W. Phillis, D. M. Johnson- Vogt, R. G., and L. M. Riddiford, 1981 Pheromone binding and
Schlitz, W. K. Benz et al., 1988 A stable source of P element inactivation by moth antennae. Nature 293: 161–163.
transposase in Drosophila melanogaster. Genetics 118: 461–470. Vogt, R. J., L. M. Riddiford and G. D. Prestwich, 1985 Kinetic
Rodrigues, V., 1988 Spatial coding of olfactory information in the properties of a pheromone degrading enzyme: the sensillar ester-
antennal lobe of Drosophila melanogaster. Brain Res. 453: 299–307. ase of Antherea polyphemus. Proc. Natl. Acad. Sci. USA 82: 8827–
Rodrigues, V., and E. Buchner, 1984 [3H] 2-deoxyglucose mapping 8831.
of odor-induced neuronal activity in the antennal lobes of Drosoph- Vogt, R. G., R. Rybczynski and M. R. Lerner, 1991 Molecular
ila melanogaster. Brain Res. 324: 374–378. cloning and sequencing of general odorant-binding proteins
Roelofs, W. L., 1995 Chemistry of sex attraction. Proc. Natl. Acad. GOBP1 and GOBP2 from the tobacco hawk moth Manduca sexta:
Sci. USA 92: 44–49. comparisons with other insect OBPs and their signal peptides. J.
Saiki, R. K., S. Scharf, F. Faloona, K. B. Mullis, G. T. Horn et al., Neurosci. 11: 2972–2984.
1985 Enzymatic amplification of b-Globin genomic sequences von Heijne, G., 1986 A new method for predicting signal sequence
and restriction site analysis for diagnosis of sickle cell anemia. cleavage. Nucleic Acids Res. 14: 4683–4690.
Science 230: 1350–1354. Woodard, C., T. Huang, H. Sun, S. Helfand and J. Carlson, 1989
Sass, G. L., J. D. Mohler, R. C. Walsh, L. J. Kalfayan and L. L. Genetic analysis of olfactory behavior in Drosophila: a new screen
Searles, 1993 Structure and expression of hybrid dysgenesis- yields the ota mutants. Genetics 123: 315–326.
induced alleles of the ovarian tumor (otu) gene in Drosophila Zhou, H., L. Ivic, J. M. Otaki, M. Hashimoto, K. Mikoshiba et al.,
melanogaster. Genetics 133: 253–263. 1998 Functional expression of a mammalian odorant receptor.
Sengupta, P., J. H. Chou and C. I. Bargmann, 1996 odr-10 encodes Science 279: 237–242.
a seven transmembrane domain receptor required for responses
to the odorant diacetyl. Cell 84: 899–909. Communicating editor: T. F. C. MackayYou can also read

















































