NOVEL CLEAVAGE SITES IDENTIFIED IN SARS-COV-2 SPIKE PROTEIN REVEAL MECHANISM FOR CATHEPSIN L-FACILITATED VIRAL INFECTION AND TREATMENT STRATEGIES
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Zhao et al. Cell Discovery (2022)8:53
https://doi.org/10.1038/s41421-022-00419-w
Cell Discovery
www.nature.com/celldisc
ARTICLE Open Access
Novel cleavage sites identified in SARS-CoV-2 spike
protein reveal mechanism for cathepsin
L-facilitated viral infection and treatment strategies
Miao-Miao Zhao 1, Yun Zhu 2, Li Zhang3, Gongxun Zhong 4,5, Linhua Tai 2,6, Shuo Liu3, Guoliang Yin2,6, Jing Lu1,
Qiong He1, Ming-Jia Li1, Ru-Xuan Zhao1, Hao Wang1, Weijin Huang 3, Changfa Fan7, Lei Shuai 4,5, Zhiyuan Wen 4,5,
Chong Wang 4, Xijun He4,5, Qiuluan Chen8,9, Banghui Liu8,9, Xiaoli Xiong 8,9, Zhigao Bu 4,5 ✉, Youchun Wang 3 ✉,
Fei Sun 2,6,8,10 ✉ and Jin-Kui Yang 1 ✉
Abstract
The spike (S) protein of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is an important target for
vaccine and drug development. However, the rapid emergence of variant strains with mutated S proteins has
rendered many treatments ineffective. Cleavage of the S protein by host proteases is essential for viral infection. Here,
we discovered that the S protein contains two previously unidentified Cathepsin L (CTSL) cleavage sites (CS-1 and CS-
2). Both sites are highly conserved among all known SARS-CoV-2 variants. Our structural studies revealed that CTSL
cleavage promoted S to adopt receptor-binding domain (RBD) “up” activated conformations, facilitating receptor-
binding and membrane fusion. We confirmed that CTSL cleavage is essential during infection of all emerged SARS-
1234567890():,;
1234567890():,;
1234567890():,;
1234567890():,;
CoV-2 variants (including the recently emerged Omicron variant) by pseudovirus (PsV) infection experiment.
Furthermore, we found CTSL-specific inhibitors not only blocked infection of PsV/live virus in cells but also reduced live
virus infection of ex vivo lung tissues of both human donors and human ACE2-transgenic mice. Finally, we showed
that two CTSL-specific inhibitors exhibited excellent In vivo effects to prevent live virus infection in human ACE2-
transgenic mice. Our work demonstrated that inhibition of CTSL cleavage of SARS-CoV-2 S protein is a promising
approach for the development of future mutation-resistant therapy.
Introduction protein1,2 and the viral proteases, including the 3C-like
The coronavirus disease 2019 (COVID-19) pandemic, protease (3CLpro, the main target of Paxlovid, Pfizer’s
caused by the novel SARS coronavirus 2 (SARS-CoV-2), experimental COVID-19 pill3) and the papain-like pro-
has resulted in a global public health crisis. Most treat- tease4 (PLpro). However, the evolution of RNA viruses is
ment strategies for SARS-CoV-2 infection are focused on driven by their high mutation rates, allowing their evasion
vaccines or antiviral drugs targeting the viral spike (S) of host immune attacks5,6. Despite multiple ongoing
efforts to treat SARS-CoV-2 infections, variant strains
with mutant spikes continue to emerge in many countries,
Correspondence: Zhigao Bu (buzhigao@caas.cn) or
Youchun Wang (wangyc@nifdc.org.cn) or Fei Sun (feisun@ibp.ac.cn) or causing new waves of infections with high morbidity5–8.
Jin-Kui Yang (jkyang@ccmu.edu.cn)
1
Of note, the very recently emerged Omicron variant
Department of Endocrinology, Beijing Diabetes Institute, Beijing Tongren
(B.1.1.529) has an unusually high number of mutations,
Hospital, Capital Medical University, Beijing, China
2
National Key Laboratory of Biomacromolecules, CAS Center for Excellence in with more than 30 in S protein, 1 in 3CLpro, and 5 in
Biomacromolecules, Institute of Biophysics, Chinese Academy of Sciences, PLpro. Therefore, antiviral drugs or vaccines targeting the
Beijing, China
viral proteins are likely to encounter the problem of
Full list of author information is available at the end of the article
These authors contributed equally: Miao-Miao Zhao, Yun Zhu, Li Zhang, resistant mutations. Indeed, three vaccines (Janssen,
Gongxun Zhong
© The Author(s) 2022
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction
in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if
changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If
material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain
permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.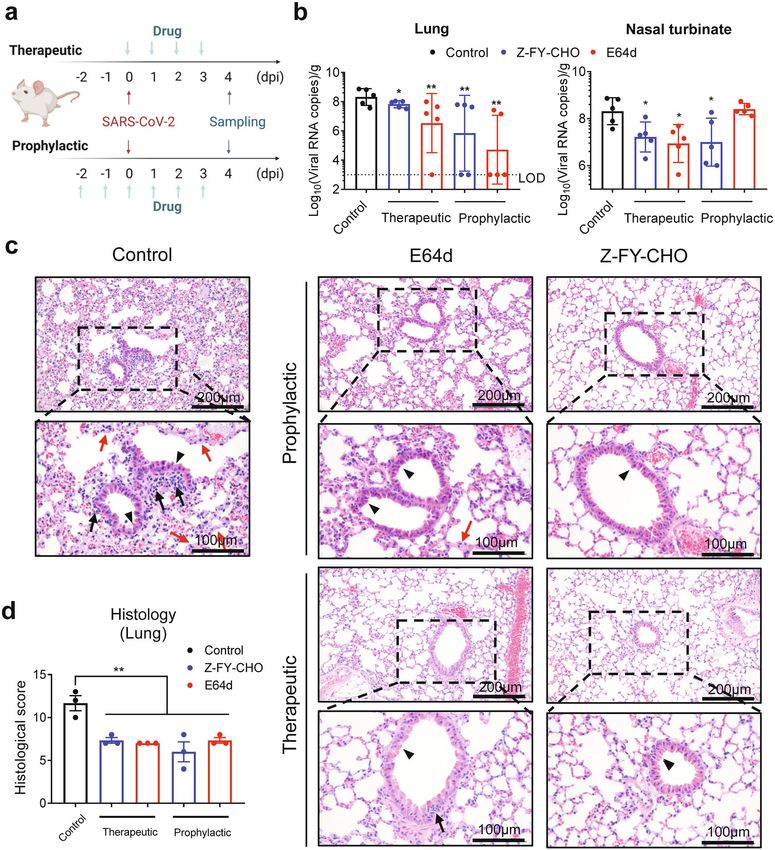
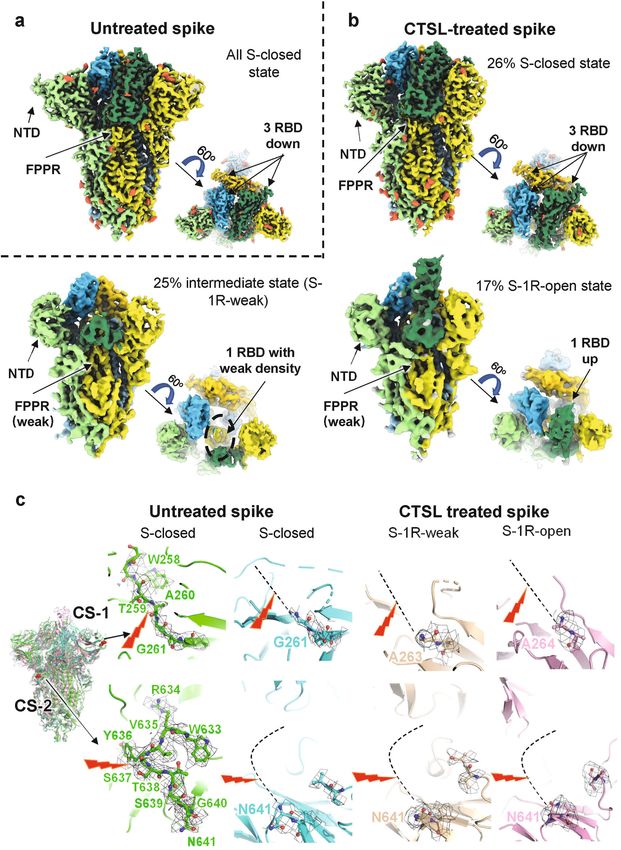
Zhao et al. Cell Discovery (2022)8:53 Page 2 of 18 Novavax, and AstraZeneca) tested against the Beta variant Results (B.1.351) exhibited reduced effects on preventing symp- CTSL cleaves the SARS-CoV-2 S protein at two specific sites tomatic infection, with effectiveness estimates of 57%, The trimeric S protein that incorporated into the 49%, and even a statistically insignificant percentage, SARS-CoV-2 envelope contains S1 and S2 subunits (Fig. respectively9. The serum neutralizing activity against the 1a, b). The S1 subunit binds to the host cellular receptor, Beta (B.1.351) variant among vaccinated persons was while the S2 subunit is involved in the virus-cell mem- reduced by 6.5- and 8.6-fold for the BNT162b2 (Pfizer) brane fusion process22, which is followed by the release of and mRNA-1273 (Moderna) mRNA vaccines, respec- viral genetic material into target cells23. To study the tively10. Molnupiravir is the chemical antiviral for the structural and functional changes in the S protein upon treatment of COVID-19 approved by the UK’s Medicines treatment with CTSL, we cloned the mammalian codon- and Healthcare products Regulatory Agency (MHRA). optimized nucleotide sequence encoding the SARS-CoV- However, the final study results Merck announced very 2 (Wuhan-Hu-1 strain, GenBank ID: MN908947.3) S recently raised questions about the drug’s benefit11. Thus, protein ectodomain (residues M1–Q1208) with proline there is an urgent need to develop broad-spectrum and substitutions at K986 and V987 and a “GSAS” substitu- mutation-resistant treatment strategies for SARS-CoV-2 tion at the furin cleavage site (S1/S2 site, R682–R685) infection. and purified the resulting protein, named S2P protein. During coronavirus infection, multiple host cell pro- This mutant S protein is widely used for structural and teases, such as furin, transmembrane serine protease 2 functional analysis due to its enhanced stability24. After (TMPRSS2), and cathepsin L (CTSL)12–14 are known to co-incubation with CTSL, the purified S protein was process S protein; however, the exact sequence of pro- cleaved into three major fragments, and the reaction tease cleavage and the interplay between host proteases exhibited a dose-dependent relationship with the CTSL remain poorly understood15. CTSL is a member of the concentration (Fig. 1c). Based on N-terminal amino acid lysosomal cysteine protease family, whose major function sequencing of these three fragments, we identified two is proteolysis of antigens produced by pathogens. It is novel CTSL cleavage sites of S, 259 T (named CTSL mainly detected in Golgi apparatus and traffic-related cleavage site 1 or CS-1) and 636Y (named CTSL cleavage organelles like endosomes and lysosome16. It is highly site 2 or CS-2) (Fig. 1c; Supplementary Fig. S1a–c), which expressed in most human tissues, including the respira- were further confirmed by liquid chromatography mass tory system, gastrointestinal tract, kidney and urinary spectrometry (LC-MS) analysis (Supplementary Fig. S1d, bladder, sexual tissues, bone marrow and lymphoid tis- e). CS-1 is located in the N-terminal domain (NTD) of sues, and endocrine tissues17. It was reported that SARS- the S1 subunit, while CS-2 is located in the C-terminal CoV-2 can directly infect and injure many extra- domain (CTD) of the S1 subunit (Fig. 1a). Similar to the pulmonary organs, like the heart, kidney, liver, pancreas, furin cleavage site at S1/S2, both CTSL cleavage sites are and gastrointestinal tract18–20, usually with high CTSL located in the exposed loops of the prefusion S protein, expression levels. Besides, the mRNA expression level of which are accessible to proteases (Fig. 1b). We noted that CTSL is higher than angiotensin-converting enzyme 2 a recent publication25 analyzed the endogenous proteo- (ACE2), FURIN, and TMPRSS2 in human lung tissues21. lysis sites of S protein during SARS-CoV-2 infection by Recently, we found that in SARS-CoV-2-infected patients, mass spectrometry and identified many cleavage sites, the circulating blood level of CTSL is highly correlated including 260 A and 637 S. However, they did not make with the severity and course of COVID-19 symptoms. We further assignments of the identified cleavage sites to also found that CTSL promotes SARS-CoV-2 pseudovirus specific proteases. Our work by discovering the CTSL (PsV) infection by cleaving the S protein and enhancing cleavage sites correlates precisely with their work and viral entry into cells12. However, the precise cleavage site provides the exact explanation of their data. As a result, and the mechanism by which CTSL activates the S protein we are more confident that the CTSL cleavage plays a remain uncharacterized. In addition, the effectiveness of very important role during the real SARS-CoV-2 infec- CTSL inhibitors in preventing or treating infections of tion process. SARS-CoV-2 virus, remains unknown. In this study, we To verify these two cleavage sites, we generated a identified two specific cleavage sites for CTSL in SARS- mutant with mutations at the sites of CS-1 and CS-2. It CoV-2 S protein, which are highly conserved among all has been reported that substrate recognition by CTSL is known SARS-CoV-2 variants. Mechanistic and functional determined mainly by the cleavage site residue (named studies revealed the mechanism by which CTSL cleavage the P1 residue) and its adjacent upstream residue (named is essential for efficient SARS-CoV-2 infection and inhi- the P2 residue)26. Therefore, we mutated both the P1 and bition of this process efficiently reduced virus infection. P2 residues in CS-1 and CS-2 to glycine to generate a 4- These results identify a new but promising target for Gly-mutant of S (Fig. 1e). As expected, treatment of the broad-spectrum antiviral therapy development. mutant with CTSL did not yield the two cleaved
Zhao et al. Cell Discovery (2022)8:53 Page 3 of 18
fragments, confirming the cleavage specificity at the sites experiments (Supplementary Fig. S3a). We also confirmed
of CS-1 and CS-2 (Fig. 1d). that the cleavage pattern of SR by CSTL is almost the
same as that of S2P (Supplementary Fig. S3b).
CS-1 and CS-2 sites are highly conserved among SARS- To ensure the comparability in the subsequent cryo-
CoV-2 variants electron microscopy (cryo-EM) structural analysis, the
To date, many SARS-CoV-2 variants have been found in samples in CTSL-treated group and control group were
different countries (Fig. 1f). These variants exhibit operated under the same condition, and the collected
enhanced transmission, pathogenicity, immune escape, or cryo-EM data were processed in the same workflow
a combination of all three, and different countries can (Supplementary Fig. S4). In the untreated control group,
have different dominant variants27. Variant viruses always all the particles fell into one population of the closed state
encoded spike proteins with substitutions in antibody (named S-closed) (Fig. 2a; Supplementary Figs. S4a and
binding hot spots. In contrast, our newly identified CTSL S8a), with a resolution of 3.2 Å in C3 symmetry according
cleavage sites in S protein are highly conserved in SARS- to the gold standard Fourier shell correlation coefficient at
CoV-2 variants (Fig. 1f), including the recently emerged 0.143 (Supplementary Figs. S5 and S6). In this closed
Omicron variant, suggesting that these CTSL cleavage state, the surface of the receptor-binding domain (RBD) is
sites may be essential for the SARS-CoV-2 life cycle. buried inside the trimer and is not accessible for the
It has been reported that SARS-CoV-2 may originate receptor ACE2 binding22,31. The NTD and fusion-peptide
from bat coronaviruses28, such as RaTG13. Further proximal region (FPPR) are highly ordered in this state,
sequence analysis of SARS-CoV-related coronaviruses resembling the “locked” closed conformation that was
(SARSr-CoVs) found in humans, bats, and pangolins previously reported in multiple studies using full-length
showed that the CS-2 site is highly conserved among most or ectodomain S proteins30–34.
SARSr-CoVs while the site of CS-1 is conserved only However, after CTSL treatment, the S particles fell into
among SARS-CoV-2 like CoVs (Supplementary Fig. S2a). four different populations, including one closed state (S-
It suggests that the two CTSL cleavage sites may evolve to closed, 26%) and three activated states, named S-1R-weak
fulfil different functional roles in different SARSr-CoVs. (25%), S-1R-open (17%), and S-2R-1N-weak (32%) (Sup-
The highly conserved CS-2 site seems to be more essential plementary Fig. S4b). Beside the S-2R1N-weak state
for the life cycle of SARSr-CoVs, while CS-1 is likely to (Supplementary Fig. S8b), the full pseudoatomic structural
play an auxiliary role in virus infection. Moreover, we models for the other three states were built at a resolution
compared the conservation of the two CTSL cleavage sites range of 3.5–4.5 Å (Supplementary Figs. S5 and S6).
among seven known human CoVs and found that these Except for the truncated NTD, the structure of S-closed
CTSL cleavage sites exist only in SARS-CoV-1 and SARS- state in CTSL-treated group is almost identical to that in
CoV-2 (Supplementary Fig. S2b), suggesting that CTSL the control group with a root mean square deviation
would play a unique role in the process of SARS-CoV-1/2 (RMSD) value of only 0.677 Å (Fig. 2b; Supplementary
infection. This observation is consistent with previous Table S1). For the S-1R-weak state (also known as inter-
studies showing that CTSL is also involved in SARS-CoV- mediate state), one of the three RBDs has a very weak
1 infection29. density, suggesting that this RBD exposes from the buried
surface and becomes dynamic for ACE2 binding (Fig.
CTSL cleavage activates SARS-CoV-2 S protein 2b)35. The RMSD value between the rest part of S-1R-
To investigate how CTSL cleavage affects the structure weak state and the S-closed state in CTSL-treated/control
of SARS-CoV-2 S, we performed cryo-EM studies of both group is 0.899/1.277 Å, respectively (Supplementary Table
the CTSL-treated and untreated S protein. The double- S1). For the S-1R-open state, one of the three RBDs
proline substitutions have been shown to affect S protein becomes upright and fully exposed for ACE2 binding (Fig.
conformation and dynamics. Therefore, to further study 2b)31, with RMSD values of 1.083/1.122 Å in comparison
the structural changes close to the natural state of S with the S-closed state in CTSL-treated/control group,
protein, we expressed and purified SARS-CoV-2 S protein respectively (Supplementary Table S1). We also noted
ectodomain named SR with minimum modifications30. that both NTDs in S-1R-weak and S-1R-open states are
Compared to the full-length S protein, SR protein contains also truncated in comparison with the S-closed state of
the ectodomain (residues M1–Q1208) of S with a foldon the control group, similar to the phenomenon observed
at the C-terminus, and the furin cleavage site (P681 to for the S-closed state after CTSL treatment. The FPPR
R685) at the S1/S2 junction is replaced with a single regions in S-1R-weak and S-1R-open states both have very
arginine (R) resembling that in SARS-CoV-1 S to avoid weak densities, also suggesting that the S proteins at these
the cleavage by furin during protein expression. The two states are activated32. Moreover, the biolayer inter-
expression tags of trimeric foldon were removed by ferometry (BLI) experiment was performed to examine
Tobacco Etch Virus protease (TEV) cleavage before our the interaction between S protein and human ACE2
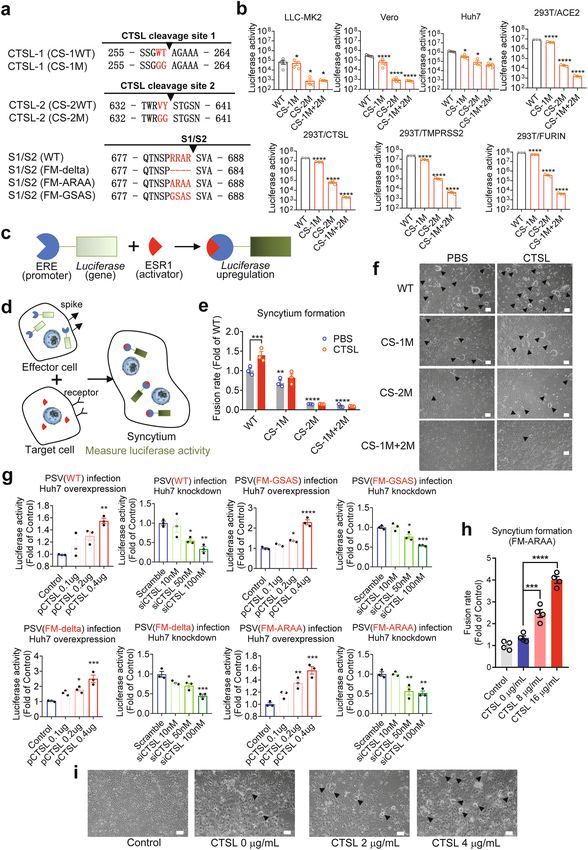
Zhao et al. Cell Discovery (2022)8:53 Page 4 of 18 Fig. 1 CTSL cleaves the SARS-CoV-2 S protein at two novel sites. a Schematic illustration of the SARS-CoV-2 S glycoprotein in which the functional domains and cleavage sites are highlighted (NTD N-terminal domain, RBD receptor-binding domain, CTD C-terminal domain, FPPR fusion-peptide proximal region, HR1 heptad repeat 1, HR2 heptad repeat 2, TMD transmembrane domain). CTSL cleaves at CTSL cleavage site 1 (CS-1) and CTSL cleavage site 2 (CS-2). Furin cleaves at the S1/S2 site. b Overall structure of the SARS-CoV-2 S ectodomain (PDB entry: 6VXX). The CTSL cleavage sites, CS-1 and CS-2, are colored in red, while the furin cleavage site at S1/S2 is colored orange. The three protomers of the S trimer are colored in pink, purple and green. c Schematic illustration and SDS-PAGE analysis for the cleavage of SARS-CoV-2 S glycoprotein. The purified SARS-CoV-2 S protein ectodomain was incubated with different concentrations of CTSL (2–8 μg/mL). The N-terminal sequencing results for band 1 (60 kDa), band 2 (100 kDa), and band 3 (170 kDa) are also shown. All samples were subjected to SDS-PAGE, and bands were detected by Coomassie blue staining. d 4-Gly-mutant SARS-CoV-2 S cannot be cleaved by CTSL into the 60 kDa (band 1) and 100 kDa (band 2) fragments. WT S2p protein (1 μg) and mutant S protein (1 μg) were incubated with CTSL (8 μg/mL). All samples were subjected to SDS-PAGE, and bands were detected by silver staining. e The P1 and P2 residues in CS-1 and CS-2 were mutated to glycine, and the mutant SARS-CoV-2-2-S glycoprotein was named 4-Gly-mutant S. f Amino acid sequence alignment of residues around CS-1 and CS-2 in SARS-CoV-2 variants. P1 and P2 residues that are the same as the WT residues are highlighted in red. The symbol * indicates amino acid residues that are conserved among all tested sequences. VOC variant of concern, VOI variant of interest, VUM variant under monitoring.

Zhao et al. Cell Discovery (2022)8:53 Page 5 of 18 Fig. 2 (See legend on next page.)
Zhao et al. Cell Discovery (2022)8:53 Page 6 of 18
(see figure on previous page)
Fig. 2 Cryo-EM structures of CTSL-untreated and -treated SARS-CoV-2 S proteins. a Side and top views of cryo-EM map of untreated S protein.
The three protomers of S protein are shown in light green, yellow, and sky blue. The three RBD domains are highlighted in Sea Green, gold, and deep
sky blue, respectively. Glycosylation modifications are colored in tomato. b Side and top views of cryo-EM map of CTSL-treated S protein in different
states. The map colors are as same as a. c The four structures of S proteins are superposed together in different colors. CS-1 and CS-2 sites are colored
in red and as indicated. The CS-1 and CS-2 regions are zoomed in to show the details. The cleavage sites are indicated by the red lighting-shaped
symbol. For untreated S protein, the electron density around the cleavage site is represented as black grid. For the CTSL-treated S, the cleavage site
cannot be traced and is represented as dotted line.
receptors before and after CTSL cleavage (Supplementary CTSL cleavage is essential for SARS-CoV-2 infection and
Fig. S7). The results showed that CTSL treatment cell fusion
increased the binding affinity between S protein and Our structural studies above suggested the important
ACE2 receptor, suggesting the activation of S protein role of CTSL cleavage in the activation of S protein. To
by CTSL. verify these observations, we investigated the functions of
For the S-2R1N-weak state, although the resolution is CTSL during viral infection. We generated several
too low to build the pseudoatomic model (Supplementary mutants of PsVs based on the two cleavage sites (CS
Fig. S8b), its functional domains, including NTD, mutant), including CS-1 mutant (CS-1M), CS-2 mutant
C-terminal domain (CTD), and RBD, could be clearly (CS-2M), and combination mutant (CS-1M + 2 M)
distinguished from the cryo-EM map. In this state, two (Fig. 3a). The binding affinity for hACE2 was not reduced
RBDs and one NTD all have very weak densities. We in the CS mutant spike (CS-1M + 2 M) compared with
postulated that this state might represent a state in which the control S2p (Supplementary Fig. S9a). As the proper
S1 is dissociating to expose S2 subunit for membrane control, we generated three PsV mutants that have been
fusion. proven to escape furin cleavage: furin mutant (FM)-delta
Compared with the control group, the proportion of the (deletion of residues 682–685), FM-ARAA (682ARAA685),
inactive state population (S-closed) was significantly and FM-GSAS (682GSAS685). The expression level of S
reduced from 100 to 26% after the cleavage by CTSL protein in different PsVs was comparable (Supplementary
(Fig. 2b), suggesting that CTSL cleavage induces sig- Fig. S9b).
nificantly increased dynamics of the RBDs and NTDs with Then, we compared the infectivity of wild-type (WT)
an increased proportion of activated S. and mutant PsVs in four human or monkey cell lines (Fig.
3b), which we have previously validated to have the
Structural evidence for CTSL cleavage site highest susceptibility to SARS-CoV-2 infection6,12,36. It
Although the overall structures of S trimer before and has been reported previously that entry into most
after CTSL treatment are basically the same (Fig. 2c), they (including Huh7, A549, and Vero E6) if not all of the cell
have significant structural differences in the local regions lines tested depends on CTSL activity12,37. All these cell
around CS-1 and CS-2 sites. For the CS-1 region, residues lines can be infected with WT PsV. However, after the
from S255 to G261 can be clearly traced in the density CTSL cleavage sites were mutated to glycine in CS
map and the cleavage site of CS-1 between T259 and mutant, these PsVs gradually lost most of their infection
A260 can be modeled with high confidence before CTSL ability in all four cell lines. For CS-1M + 2 M PsV with
treatment (Fig. 2c). However, after CTSL treatment, all both CTSL cleavage sites mutated, the viral infectivity was
the densities around CS-1 site disappeared, and we can reduced 100–700-fold compared with that of WT PsV
only build as far as G261 in the models of S-closed state, (Fig. 3b), showing that the two CTSL cleavage sites,
and A263/A264 in the S-1R-weak/S-1R-open state (Fig. especially CS-2, are essential for SARS-CoV-2 infection.
2c). These results confirmed the occurrence of CTSL Moreover, we also examined the infectivity in three
cleavage around the CS-1 site. 293 T/ACE2 cell lines with stable overexpression of CTSL,
For the CS-2 region, residues from W633 to N641 can TMPRSS2, or FURIN (Fig. 3b). We found that the
be clearly traced in the density map and the cleavage site reduction in infectivity of CS mutant PsVs could not be
of CS-2 between Y636 and S637 can be modeled with high fully rescued by overexpression of CTSL, TMPRSS2, or
confidence before CTSL treatment (Fig. 2c). However, FURIN, indicating that the function of CTSL could not be
after CTSL treatment, all the densities around CS-2 site fully substituted by TMPRSS2 or furin. Similar results
disappeared, and we can only build as far as N641 in the were also observed with different PsV concentrations
models of S-closed, S-1R-weak and S-1R-open states. (Supplementary Fig. S9c).
These results also confirmed the occurrence of CTSL The fusogenic activity of SARS-CoV-2 S is a character
cleavage around the CS-2 site. of SARS-CoV-2 infection, and the presence of syncytia inZhao et al. Cell Discovery (2022)8:53 Page 7 of 18 Fig. 3 (See legend on next page.)
Zhao et al. Cell Discovery (2022)8:53 Page 8 of 18
(see figure on previous page)
Fig. 3 CTSL cleavage sites are essential for SARS-CoV-2 infection and efficient cell–cell fusion. a Overview of SARS-CoV-2 S proteins with
mutations in CS-1, CS-2, and the S1/S2 cleavage site. b Infectivity of PsVs with different point mutations in CS-1 and CS-2 was assessed in LLC-MK2,
Vero, Huh7, 293 T/ACE2 cells and in 293 T/ACE2 cells with CTSL (293 T/CTSL), TMPRSS2 (293 T/TMPRSS2), and FURIN (293 T/FURIN) genes
overexpression. PsV infectivity was measured by a luciferase assay and is shown as the raw luciferase activity (n = 3–4). Statistical significance was
assessed by one-way ANOVA with Tukey’s post-hoc test. c–e Quantitative analysis of syncytium formation induced by CS mutant SARS-CoV-2 S
proteins (see also Supplementary Fig. S9). c Luciferase gene expression was driven by the ERE promoter, and ESR1 (activator) bound and activated the
ERE promoter to upregulate luciferase expression. d, e Effector Huh7 cells were cotransfected with plasmids expressing ERE-luciferase and different S
proteins as indicated, and another plate of target Huh7 cells was transfected with plasmid expressing ESR1. After 24 h, the effector cells were
detached and added to the target cells for 30–60 min. Then, the supernatant was removed and treated with PBS or CTSL (8 μg/mL) for 20 min. The
reaction was stopped by adding 500 μL of medium, and culture was continued for another 24 h to allow cell–cell fusion. When a target cell and
effector cell fused to form a syncytium, ESR1 bound and activated the ERE promoter to upregulate luciferase expression. Luciferase activity was then
measured as a proxy for the fusion rate. The data were normalized to the WT-PBS group (n = 3). Statistical significance was assessed between the
indicated group and the WT-PBS group by two-way ANOVA with Dunnett’s post-hoc test. f Images of syncytium formation induced by CS mutant
SARS-CoV-2 S proteins. Huh7 cells were transfected with plasmids to express the WT, CS-1M, CS-2M, or CS-1M + 2 M S protein. Cells were treated in
the absence (PBS, pH = 5.8) or presence of CTSL (4 μg/mL, pH = 5.8). Images were acquired after an additional 10–16 h of incubation in medium
(scale bars, 50 μm). The black arrowheads indicate syncytia. Representative data from three independent experiments are shown. g Overexpression or
knockdown of the CTSL gene dose-dependently promoted or inhibited, respectively, infection with WT (Wuhan-1) and three mutant SARS-CoV-2 PsVs
with different point mutations in the furin cleavage site (FM-delta, FM-ARAA, and FM-GSAS). PsV infectivity in Huh7 cells was measured by a luciferase
assay and is shown as the relative luciferase activity (n = 3). Statistical significance was assessed by one-way ANOVA with Tukey’s post-hoc test.
h CTSL promoted syncytium formation induced by the FM-ARAA mutant SARS-CoV-2 S protein. Effector cells were cotransfected with ERE-luciferase
plasmids and either FM-ARAA S or scramble vectors (Control). Target cells were transfected with ESR1 expression plasmid. After the effector cells and
target cells were mixed, the supernatant was removed and treated with PBS or CTSL (8 μg/mL and 16 μg/mL). Luciferase activity was then measured
and normalized to that in the control group (n = 4). Statistical significance was assessed by one-way ANOVA with Tukey’s post-hoc test. i Huh7 cells
were transfected with scramble vector (Control) or FM-ARAA S protein expression plasmid. Cells were treated in the absence or presence of CTSL (2
or 4 μg/mL). Images were acquired after an additional 10–16 h of incubation in medium (scale bars, 50 μm). The black arrowheads indicate syncytia.
Representative data from four independent experiments are shown. The data are presented as the means ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001,
***P < 0.0001.
COVID-19 patient lung tissue indicates that this activity markedly increased its infection efficiency, while
plays a role in the pathological process38. Syncytium for- knockdown of this gene significantly reduced the infec-
mation was quantified with an ESR1-ERE transactivation tion efficiency (Fig. 3g), similar to our previous finding12.
system (Fig. 3c–e; Supplementary Fig. S10a, b) and This result indicated that CTSL plays a crucial role in the
observed by bright-field microscopy in our study (Fig. 3f). SARS-CoV-2 infection process. Furthermore, for the
With the similar expression levels of S protein in trans- FM-mutants PsVs, overexpression or knockdown of the
fected cells (Supplementary Fig. S10c), compared to the CTSL gene also significantly affected the infection levels
control group, the CS-1M group exhibited significantly in a dose-dependent manner (Fig. 3g), similar to the
reduced syncytium formation (Fig. 3e, f), while syncytium effects on WT PsV. These results suggested that CTSL-
formation was inhibited even more severely in the CS-2M enhanced SARS-CoV-2 viral entry is independent of
group, and almost no syncytia were observed in the CS- furin cleavage.
1M + 2 M group (Fig. 3e, f). More importantly, the inhi- The furin cleavage site has been reported required for
bitory effects of these mutations were not rescued by the SARS-CoV-2 S driven cell–cell fusion13. Here, we used
addition of CTSL into the system (Fig. 3e, f), confirming the same cell fusion system (Fig. 3d) to evaluate the
the promotive effect of CTSL on syncytium formation, as effect of CTSL on FM-mutants-mediated syncytium
previously shown12, was due to S protein cleavage at CS-1 formation. FM-ARAA PsV induced a low level of syn-
and CS-2 sites. Taken together, our findings proved that cytium formation similar to that of the control group
CTSL cleavage at CS-1 and CS-2 sites is essential for (Fig. 3h, i). However, after CTSL was added to the
SARS-CoV-2 infection. mutant group, syncytium formation increased sig-
nificantly, and this rescue effect exhibited a dose
CTSL cleavage promotes SARS-CoV-2 infection dependency with the concentration of CTSL (Fig. 3h, i).
independent of furin This result suggested that CTSL cleavage is an important
As the CS-2 region is very close to the furin cleavage furin-independent factor to induce SARS-CoV-2 S dri-
site, we next investigated the relationship between CTSL ven cell–cell fusion.
cleavage and furin cleavage by using PsV infection assays
in human Huh7 cell line6,36,39 under CTSL over- CTSL inhibitors prevent SARS-CoV-2 infection
expression or knockdown conditions (Supplementary Since CTSL plays a key role in mediating SARS-CoV-2
Fig. S11). For WT PsV, overexpression of the CTSL gene infection, we selected six compounds (K777, cathepsinZhao et al. Cell Discovery (2022)8:53 Page 9 of 18 Fig. 4 (See legend on next page.)
Zhao et al. Cell Discovery (2022)8:53 Page 10 of 18 (see figure on previous page) Fig. 4 CTSL inhibitors prevent infection with SARS-CoV-2 and mutant variant PsVs in vitro. a Vero E6 cells were pretreated with increasing concentrations of each compound for 16 h and were then infected with different SARS-CoV-2 variant PsVs as indicted. At 24 hpi, infectivity was measured by a luciferase assay. The data were normalized to the average value in vehicle-treated cells and are shown as inhibition rates (n = 3). Statistical significance was assessed between the indicated variant and WT PsV by two-way ANOVA with Dunnett’s post-hoc test. The data are presented as the means ± SEM. *P < 0.05, **P < 0.01. b Vero E6 cells were pretreated with increasing concentrations of each compound for 16 h and were then infected with SARS-CoV-2 at an MOI of 0.01. At 24 h post-infection, viral RNA copies in supernatants were quantified by RT-qPCR. The data were normalized to the average value in vehicle-treated cells and are shown as relative infection percentages. The EC50 values for each compound are indicated. Cell viability was evaluated with a CCK kit (TransGen Biotech) (n = 3). c Ex vivo lung tissues from hACE2 mice or a human donor were infected with SARS-CoV-2 with an inoculum of 1 × 106 PFU/mL for 2 h. Then the inoculum was removed and changed with medium with indicated compounds (10 μM for molnupiravir, 4 μM for E64d, 5 μM for Z-FY-CHO, and 0.4 μM for K777) for another 48 h. Tissues were harvested (without adding compounds) at 2 hpi or 48 hpi to determine the viral growth ability. Tissues were harvested at 48 hpi for quantification of viral RNA (n = 3). Statistical significance was assessed between 2 hpi and 48 hpi by unpaired two-tailed Student’s t-test. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Statistical significance was assessed between the indicated drug and 48 hpi by one-way ANOVA with Tukey’s post-hoc test. #P < 0.05, ##P < 0.01, ###P < 0.001, ####P < 0.0001. The data are presented as the means ± SEM. inhibitor 1, E64d, Z-FY-CHO, MDL-28170, and oxo- Thirdly, we further validated the antiviral effects of the carbazate) that can inhibit CTSL activity by over 90% with compounds in an ex vivo lung culture system. Lung tis- a concentration ranging from 30 nM to 50 μM, respec- sues from human ACE2 (hACE2)-transgenic mice or a tively (Supplementary Fig. S12), and evaluated their effects human donor were infected with the live SARS-CoV-2 on SARS-CoV-2 infection. virus. First, tissues were harvested (without adding drugs) Firstly, we examined the antiviral effects of the six at 2 h post-infection (hpi) and 48 hpi to examine the compounds against variants of SARS-CoV-2 (including system. The viral RNA copies from tissues harvested at 48 the recently emerged Omicron variant) using the PsV hpi were significantly higher than those from 2 hpi, infection assays (Fig. 4a). As we expected, all six indicating that the tissues were successfully infected compounds can efficiently inhibit infection of all (Fig. 4c). Then, lung tissues were infected and treated with mutant PsVs variants. Among them, E64d and MDL- the selected compounds, with molnupiravir as a control. 28170 showed higher inhibitory efficiency against Three CTSL inhibitors (E64d, Z-FY-CHO, and K777) Omicron variant, while oxocarbazate was more effec- were chosen for their lower EC50 and safety for in vivo tive against Epsilon variant. It seems that CTSL inhi- studies12. All compounds were used in the dose of double bitors are most likely resistant to mutational escape of EC90 concentrations assessed from Vero E6 cells41 (10 μM SARS-CoV-2, including Omicron and Beta variants, for molnupiravir42, 4 μM for E64d, 5 μM for Z-FY-CHO, against which the effects of most current vaccines are and 0.4 μM for K777). After 48 h, tissues were harvested reduced9,10,40. Neither of the compounds inhibited and the viral RNA copies were quantified. All CTSL control vesicle stomatitis virus (VSV) PsV virus infec- inhibitors potently antagonized viral replication in human tion (Supplementary Fig. S13), indicating that the and mouse lung tissues, even with stronger effects than inhibitory effects of CTSL inhibitors depend on SARS- molnupiravir (Fig. 4c). CoV-2 S protein. Finally, we investigated the in vivo antiviral effects of Secondly, in the cellular experiments using live SARS- CTSL inhibitors in hACE2 mice, which are susceptible to CoV-2 virus, all six compounds efficiently blocked SARS-CoV-2 infection43. We selected two compounds, SARS-CoV-2 infection at non-toxic doses, with half- E64d and Z-FY-CHO, which have been validated to be maximal effective concentration (EC50) values ranging safe for in vivo studies12,44. E64d and Z-FY-CHO were from 35 nM to 10.9 μM (Fig. 4b). All compounds administered in prophylactic and therapeutic regimens, decreased the viral RNA load in Vero E6 cells by ~103- and mice were treated at appropriate doses according to to 104-fold at the maximal non-toxic dose. This finding previous studies12,44. Each mouse was infected with 106 further confirmed the critical role of CTSL cleavage in plaque-forming units (PFU) SARS-CoV-2 at 0 days post- SARS-CoV-2 infection. To be noted, we confirmed that infection (dpi) by intranasal instillation. Tissue samples all drugs lack potent inhibitory activity on the viral were collected at 4 dpi (Fig. 5a), when the viral load PLpro and 3CLpro (Supplementary Fig. S14), and also on peaked, and exhibited obvious histopathological changes. human furin and TMPRSS2 (Supplementary Fig. S15) Both compounds significantly decreased the number of under the highest tested concentrations used in this viral RNA copies in lung tissues by ~1–4 log10, with infection study, indicating that the antiviral activities of undetectable (below the lower limit of detection, LOD) these compounds are most likely due to the CTSL- viral loads in lung tissues from two mice treated with Z- specific inhibition. FY-CHO and four mice treated with E64d (Fig. 5b). Both
Zhao et al. Cell Discovery (2022)8:53 Page 11 of 18 Fig. 5 CTSL inhibitors prevent SARS-CoV-2 infection in vivo. a E64d and Z-FY-CHO were administered intraperitoneally at 2 days before infection to 3 dpi as prophylactic treatment, and mice were challenged with 106 PFU at 0 dpi; the two drugs were administered therapeutically at 0–3 dpi. Tissue samples were collected at 4 dpi. b Viral RNA copies in mouse lung and nasal turbinate tissues (n = 5 mice/group). The dotted line indicates the lower limit of detection (LOD). Statistical significance was assessed between the indicated group and control group by one-way ANOVA with Tukey’s post-hoc test. c Representative images from histological analysis of lungs from SARS-CoV-2-infected hACE2 mice at 4 dpi. Magnified views of the boxed regions in each image are shown below the corresponding image. The black arrows indicate inflammatory cell infiltration, the black arrowheads indicate bronchiolar epithelial cell degeneration, and the red arrows indicate alveolar septal thickening. The scale bars are indicated in the figures. d Semiquantitative histological scoring of each lung tissue was performed by grading the severity of bronchiolar epithelial cell damage (0–10), alveolar damage (0–10) and inflammatory cell infiltration in blood vessels and bronchioles (0–10) and summing these scores to calculate the total score. Normal = 0, indeterminate = 1–2, mild = 3–4, moderate = 5–7, severe = 8–10. (n = 3) Statistical significance was assessed between the indicated group and control group by one-way ANOVA with Tukey’s post-hoc test. drugs also significantly decreased the viral load in the pathological changes, as evidenced by the large areas of nasal turbinate (Fig. 5b). Histological analysis of the lungs alveolar septal thickening, inflammatory cell infiltration showed that vehicle-treated mice exhibited moderate and bronchiolar epithelial cell degeneration. In contrast,
Zhao et al. Cell Discovery (2022)8:53 Page 12 of 18
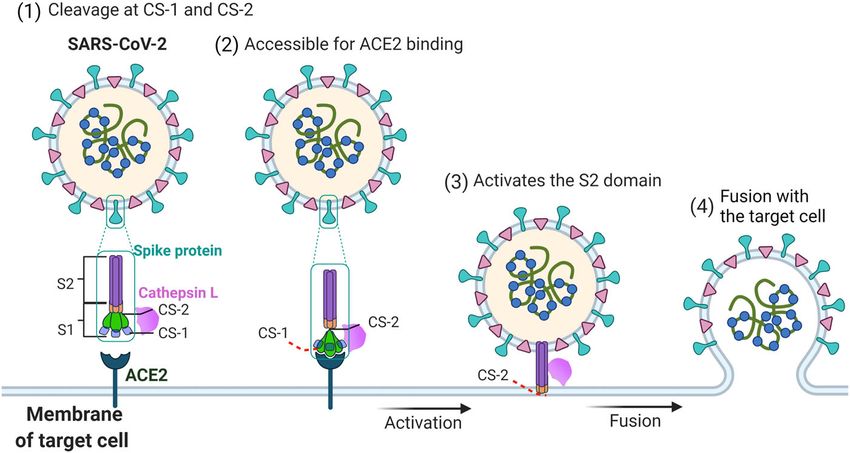
Fig. 6 Proposed mechanism by which CTSL promotes SARS-CoV-2 infection. SARS-CoV-2 S contains S1 and S2 subunits. CS-1 is located in the
NTD of the S1 subunit, and CS-2 is located near the S1/S2 site. CTSL cleaves the SARS-CoV-2 S protein: (1) By binding with the S protein on the surface
of SARS-CoV-2, CTSL cleaves S at CS-1 and CS-2 sites. (2) The cleavage increases the dynamics of the RBD and makes it accessible to ACE2 for binding.
(3) CTSL cleaves CS-2 site to separate S1 and S2 subunits to expose the S2 subunit for membrane fusion. (4) The virus fuses with the membrane of
target cell, and the viral genetic material is released into the host cell.
the lungs of E64d- and Z-FY-CHO-treated mice exhibited CS-1 and CS-2 sites on the S protein. Then dynamics of
improved morphology and less infiltration (Fig. 5c, d). the RBD and NTD are increased to facilitate the activation
Taken together, these results indicated that CTSL inhi- of S protein and promote ACE2 binding. Next, the clea-
bitors efficiently block SARS-CoV-2 infection both vage at CS-2 site separates S1 and S2 domains to expose
in vitro and in vivo and are resistant to viral mutational S2 ready for membrane fusion and then complete the
escape. infection process. Considering the localization and activ-
ity characteristics of CTSL, these cleavage reactions are
Discussion most likely to occur in the endosomes.
In this study, we identified two specific cleavage sites Previous studies indicated that SARS-CoV-1, MERS-
of CTSL in the SARS-CoV-2 S protein, named CS-1 and CoV, and SARS-CoV-2 were all capable of using both
CS-2. Both sites are located in the flexible loop regions endosomal and membrane fusion routes to enter the
within S1 subunit, while CS-1 is in the NTD and CS-2 is cell13,45. TMPRSS2 appears to be one of the major pro-
in the CTD near the S1/S2 furin cleavage site. Both sites teases for priming S for entry via the plasma membrane,
are highly conserved among all SARS-CoV-2 variants, whereas CTSL performs the priming function during
including the most recently emerging variants. Our entry through the endosome in vitro13,45. In support of
structural studies proved that CTSL cleavage can greatly this view, endosomal acidification inhibitors have been
enhance S protein dynamics with an increased propor- shown to effectively block the entry of SARS-CoV-1 and
tion of activated states ready for receptor-binding and SARS-CoV-2 by preventing the activation of CTSL29,45.
membrane fusion. The viral infection assay using the Cell culture studies showed that TMPRSS2 was more
PsV system proved that CTSL cleavage, especially significantly associated with the entry of SARS-CoV-2
cleavage at CS-2, is an essential step in SARS-CoV-2 into Calu-3 cells13, while the endosomal entry route might
infection. Furthermore, CTSL-specific inhibitors not be dominant for SARS-CoV-2 into Vero cells46. There-
only blocked the infection of PsV/live virus in cells but fore, the flexibility of protease usage and entry pathway
also reduced the infection rate of live virus significantly seems to be a consistent strategy for coronavirus use, and
ex vivo and in vivo. Our results suggested CTSL clea- in some cases, its role depends on cell type. In patients
vage is an important process during SARS-CoV-2 with SARS-CoV-2 infection, the most susceptible organ is
infection. the lung, which contains many kinds of cell type. In this
Based on our structural and functional study, we pro- study, we further validated the antiviral effects of the
posed a possible model for the CTSL-mediated SARS- CTSL inhibitors in human lung tissue ex vivo.
CoV-2 infection process (Fig. 6). When the viral S protein In addition, SARS-CoV-2 attachment cofactors can
encounters activated CTSL, the cleavage occurs at both facilitate SARS-CoV-2 S binding to ACE2. Subsequently,Zhao et al. Cell Discovery (2022)8:53 Page 13 of 18 the cleavage of cell surface proteases can lead to cell mutations. Our study shows that CTSL and its new membrane fusion23. Our study indicates that CTSL can cleavage sites at the SARS-CoV-2 S protein are the crucial attach S protein at CS-1 site, implying that may CTSL new targets for the future development of anti-SARS- serve as SARS-CoV-2 attachment cofactor to facilitate CoV-2 therapy. SARS-CoV-2 S binding to ACE2. It is well known that furin and TMPRSS2 are important Materials and methods human enzymes that are employed by SARS-CoV-2 Protein expression and purification during the infection process. The expression of The S2P protein was purified as previously reported24. TMPRSS2 has been confirmed within COVID-19 relevant Briefly, the prefusion S ectodomain, the mammalian lung cell types19 and its role in mediating SARS-CoV-2 codon-optimized gene coding SARS-CoV-2 (Wuhan-Hu- infection has been proved13. TMPRSS2 also has been 1 strain, GenBank ID: MN908947.3) S glycoprotein reported to mediate ACE2 activation in SARS-CoV-1 ectodomain (residues M1–Q1208) with proline sub- infection47. Here, we further identified that SARS-CoV-2 stitutions at K986 and V987, a “GSAS” substitution at the can utilize human CTSL to assist infection independent of furin cleavage site (R682 to R685), a C-terminal thrombin furin. Therefore, all these human enzymes, including tag, a T4 fibritin trimerization motif and a 6× HisTag was CTSL, TMPRSS2, and furin, could be involved in the viral synthesized and cloned. To express 4-Gly-mutant S infection process, and are most likely to play com- protein, residues W258, T259, V635, and Y636 were plementary or compensatory roles in different human substituted into glycine. These expression vectors were tissues. Drugs targeting these human enzymes should be used to transiently transfect Expi293 cells and purified by paid more attention to. using cOmpleteHis-Tag Purification Resin to capture the Moreover, for the drugs that do not target the virus target protein. The protein was then subjected to addi- directly, they would be less easily escaped by the SARS- tional purification by size-exclusion chromatography CoV-2 variants. As expected, we found that the CTSL- using Superdex 200 10/300 GL column (GE Healthcare) specific inhibitors can nearly completely prevent infection in PBS. of SARS-CoV-2 variant PsVs and reduce infection of live The SR protein with only an arginine left at the furin SARS-CoV-2 by 103–104-fold in Vero E6 cells. More cleavage site (P681–R685) was constructed, expressed, importantly, CTSL-specific inhibitors can effectively pre- and purified as previously described30. Briefly, 300 μg of vent SARS-CoV-2 infection in hACE2-transgenic mice. plasmid was incubated with 810 μg of polyethylenimine These results have provided important evidence for the for 10 min, and then transfected into 300 mL of 293 F cells development of next-generation broad-spectrum drugs at 2.7 million/mL. The transfected cells were cultured at against SARS-CoV-2. 33 °C. After 4 days, cell culture supernatant was collected In our study, we found that both CTSL cleavage sites and then loaded onto 5 mL Hitrap Talon Cobalt column. (CS-1 and CS-2) possess extra high sequence conserva- The column was washed with 100 mL buffer A (25 mM tions among SARS-CoV-2 variants, indicating their phosphate pH 8.0, 300 mM NaCl, 5 mM imidazole) and importance for the viral life cycle. It may be inferred that eluted with 100 mL 0–100% linear gradient of buffer B those two cleavage sites cannot be easily mutated in the (25 mM phosphate pH 8.0, 300 mM NaCl, 200 mM imi- live SARS-CoV-2 variants, or that the variant with dazole). After that, the protein was concentrated and mutations in those two sites might have weak infectivity, buffer exchanged into PBS by a 100 kDa MWCO ultra which is indicated in our experiments that the mutations centrifugal filter (Millipore). of these two sites, especially CS-2 site, largely impaired the infectivity of the virus. Therefore, the development of CTSL treatment specific drugs or vaccines targeting CS-1 and CS-2 sites Recombinant CTSL (Novoprotein) was used to cleave would be a new strategy to prevent SARS-CoV-2. If these SARS-CoV-2 S2P protein. Purified SARS-CoV-2 S protein sites are occupied by drugs or antibodies which can block was incubated with CTSL (2–8 μg/mL as indicated and CTSL cleavage, the viral infection could be effectively 8 μg/mL for cryo-EM sample preparation) in the presence inhibited. This approach would be much safer than the of 100 mM NaAC, 1 mM EDTA and 1 mM dithiothreitol strategy of using compounds to inhibit the activity of (DTT), pH 5.5 for 24 h at 25 °C. CTSL was preactivated at multiple human proteases. 30 °C for 1 min before use. The resulting protein was then In conclusion, the high mutation rate of SARS-CoV-2 subjected to sodium dodecyl sulfate–polyacrylamide gel often leads to viral escape from neutralizing antibodies or electrophoresis (SDS-PAGE) and stained by coomassie vaccines. An understanding of the precise mechanisms blue or silver staining methods as indicated. For cryo-EM underlying SARS-CoV-2 infection is urgently needed to sample preparation, the SR protein was firstly cleaved by facilitate the development of broad-spectrum antiviral TEV enzyme in a mass ratio at 10:1 for 16 h at 25 °C, then drugs or vaccines with increased resistance to viral treated with CTSL in pH 5.5 for 24 h at 25 °C.
Zhao et al. Cell Discovery (2022)8:53 Page 14 of 18
Sequence analysis equipped with a GIF Quantum energy filter, with a silt
Sequence alignments were performed using the width of 20 eV, operated in super-resolution mode,
CLUSTALW online tool (https://www.genome.jp/tools- resulting in a pixel size of 0.68 Å for output super-
bin/clustalw). The representative variant sequence of each resolution movies. SerialEM49 with in-house scripts were
SARS-CoV-2 PANGO lineage was obtained from Global used for data collection50, defocus values for either sam-
Initiative on Sharing All Influenza Data (GISAID) data- ples were set between −1.0 and −2.0 μm.
base: B.1.1.529 (GISAID: EPI_ISL_6900143), B.1.617
(GISAID: EPI_ISL_1544002), P.1 (GISAID: Image processing
EPI_ISL_1164984), B.1.351 (GISAID: EPI_ISL_935042), Image processing steps were performed using
B.1.1.7 (GISAID: EPI_ISL_1257795), B.1.621 (GISAID: RELION51,52 and cryoSPARC53. The output super-
EPI_ISL_6864938), C.37 (GISAID: EPI_ISL_6794513), resolution movies were first subjected to motion correc-
AY.4.2 (GISAID: EPI_ISL_6905104), C.36 (GISAID: tion using MotionCor254, with the binning level of 2 in
EPI_ISL_6866297), C.1.2 (GISAID: EPI_ISL_6906407), Fourier space, and dose weighting was also performed
B.1.427 (GISAID: EPI_ISL_1221570), B.1.1.318 (GISAID: during the process. Contrast transfer function (CTF)
EPI_ISL_6661876), B.1.429 (GISAID: EPI_ISL_1160035), parameters estimation were performed using Gctf55.
B.1.640 (GISAID: EPI_ISL_6887009), B.1.525 (GISAID: Gautomatch (https://www.mrc-lmb.cam.ac.uk/kzhang/
EPI_ISL_6903493), P.3 (GISAID: EPI_ISL_6756120), Gautomatch/) was used to pick all particles from raw
B.1.617.1 (GISAID: EPI_ISL_6877952). In addition, the micrographs. Then good particles were extracted and
following sequences information were obtained from the sorted by 2D classification. After 2D classification, one
National Center for Biotechnology Information (NCBI) round of 3D classification was performed to investigate
database: SARS-CoV BJ01 (GenBank: AAP30030.1), different populations of S proteins. To prevent any model
MERS-CoV (GenBank: QBM11748.1), HCoV NL63 bias, the previously reported cryo-EM structure of SARS-
(GenBank: APF29063.1), HcoV 229E (GenBank: CoV-2 S (Protein Data Bank, PDB entry 6VXX) lowpass
AWH62679.1), HcoV OC43 (GenBank: AIX10763.1), filtered to 60 Å was used as the initial reference and no
HcoV HKU1 (GenBank: AMN88694.1). additional symmetry was imposed throughout the initial
image processing pipeline. Then the well-aligned classes
N-terminal Edman sequencing and LC-MS/MS analysis with clear secondary structure features were selected
CTSL-treated SARS-CoV-2 S protein was resolved on separately for subsequent image processing, including
an SDS-PAGE gel. The protein was transferred to PVDF auto-refinement, Bayesian polishing, CTF refinement, and
membrane before Edman sequencing. Each protein band no-alignment 3D classification. All these steps were per-
was excised individually. The sequence of the first 10 formed in RELION. For the final round of refinement, the
amino acids for each band was determined by Edman non-uniform (NU) refinement in cryoSPARC was per-
sequencing using ABI Procise-cLC machine. LC-MS/MS formed to refine the final maps to a higher resolution.
analysis of excised bands was performed at Laboratory of
Proteomics, Institute of Biophysics, Chinese Academy of Model building and refinement
Sciences. All the cryo-EM maps are improved by density mod-
ification procedures before model building56. To build the
Cryo-EM sample preparation and data collection atomic model of CTSL-treated or untreated SARS-CoV-2
For each grid, 3 μL of purified protein solution of CTSL- S structures, the reported cryo-EM structure of SARS-
treated or untreated SARS-CoV-2 S proteins were applied CoV-2 S (PDB entry 6VXX & 6VYB)22 was used as an
to newly glow-discharged holy carbon film grids (R1.2/1.3, initial model. We were able to trace most regions with
200 meshes, Au, Quantifoil, Germany) or holy Ni-Ti film side chains using COOT57. The built model was further
grids48 (R1.2/1.3, 300 meshes, Au, Zhenjiang Lehua refined in real space using Phenix58. All figures were
Electronic Technology Co., LTD, China). Then the grid created by Pymol59, UCSF Chimera60, and UCSF Chi-
was blotted and vitrified by plunge freezing into liquid meraX61. The parameters for data collection and structure
ethane using Vitrobot Mark IV (Thermo Fisher Scientific, determination are summarized in Supplementary Table
USA) at 4 °C and 100% humidity. All movies were col- S2. The overall model building quality is shown in Sup-
lected on a Titan Krios G2 TEM (Thermo Fisher Scien- plementary Videos S1–S4.
tific, USA) operated at 300 KV on EF-TEM mode with
nominal magnification set to be ×105,000, resulting in a BLI
calibrated physical pixel size of 1.36 Å on a specimen The BLI assay was performed at 30 °C on the Octet
level. The total dose was set to be 60 e−/Å2, with an RED96 System (ForteBio) with 100 mM NaAC, 1 mM
exposure time of 15 s. The movies were acquired by Gatan EDTA, and 1 mM DTT, pH 5.5 and 0.02% Tween-20 as
K2 Summit DDD detector (Gatan Company, USA) running buffer. Purified human IgG Fc-hACE2Zhao et al. Cell Discovery (2022)8:53 Page 15 of 18
(Novoprotein) was loaded for 300 s onto Anti-Human IgG procedure described by us previously. For this VSV-based
Fc Capture (AHC) biosensors (ForteBio). To measure the PsV system, the backbone was provided by VSV-G pseu-
binding affinity between ACE2 and S variants, the bio- dotyped virus (G*ΔG-VSV) that packages expression cas-
sensors were then incubated with various concentrations settes for firefly luciferase instead of VSV-G in the VSV
of the purified WT S2P and CS mutant (CS-1M + 2 M) S2P genome. For quantification of PsV, viral RNA was extracted
proteins for 90 s, followed by 180 s of dissociation. Both by using the QIAamp Viral RNA Mini Kit (Cat. No. 52906,
WT and CS mutant S2P proteins have proline substitu- QIAGEN), and the reverse transcription was performed with
tions at K986 and V987 and a “GSAS” substitution at the RevertAidTM First Strand cDNA Synthesis Kit (Fermentas
furin cleavage site. For the CTSL cleavage activation assay, K1622) according to the manufacturer. The real-time qPCR
the biosensors were incubated for 180 s with various was then performed on the LightCycler® 96 Real-Time PCR
concentrations of SR before and after CTSL treatment, System (Roche) using SYBR Green I Master Mix reagent
followed by 200 s of dissociation. The data were analyzed (Roche). The P protein gene of VSV virus was quantified and
using the software supplied by the manufacturer. the viral copy number calculated accordingly. The primers
were: forward-TCTCGTCTGGATCAGGCGG; reverse-
Cell culture TGCTCTTCCACTCCATCCTCTTGG. All PsVs were
The Huh7 (Homo sapiens, liver), 293T-hACE2 normalized to the same amount as previously described6.
(293 T cells stably expressed, Homo sapiens, embryonic
kidney), Vero (Cercopithecus aethiops, kidney), VeroE6 Western blot analysis
(Cercopithecus aethiops, kidney), and LLC-MK2 (Macaca 7 mL of SARS-CoV-2 PsVs with a titre of 1.86 × 105
mulatta, kidney) cells were maintained in high glucose 50% tissue culture infectious dose/mL (TCID50/mL) were
Dulbecco’s modified Eagle’s medium (DMEM) pelleted through a 25% sucrose cushion by ultra-
(Sigma–Aldrich, St. Louis, MO, USA) supplemented with centrifugation at 100,000 × g for 3 h. The layer of super-
10% fetal bovine serum (FBS, Gibco, Carlsbad, CA), 100 natant was discarded. The pellet was collected and sub-
units/mL Penicillin-Streptomycin (Gibco). All the cells jected to western blot analysis as previously described62,
were maintained at 37 °C in a humidified atmosphere and detected by anti-S1 mice serum, VSV-M antibody
containing 95% air and 5% CO2. (KeraFast, Cat. No. EB0011), anti-GAPDH antibody
(Sigma), and anti-His antibody (Sigma).
Experimental mice
The study used human ACE2-transgenic mice, a mouse PsV infection assay
model expressing hACE2 generated by using clustered Before infection, the 96-well plates were seeded with
regularly interspaced short palindromic repeats/CRISPR cells adjusted to 2 × 105 cells/mL. Then, 100 μL of the
associated protein 9 (CRISPR/Cas9) knock-in technology normalized PsV with indicated dilution fold was added to
as previously reported43. The hACE2 mice used in this wells in 96-well cell culture plate. After 24 h incubation at
manuscript were 17-week-old female C57BL/6 mice, with 37 °C, the activities of firefly luciferase were measured on
the body weight between 22 g. All animal protocols were cell lysates using luciferase substrate (PerkinElmer,
approved by the Ethical Review Committee at the Insti- BRITELITE PLUS 100 mL KIT, Cat. No. 6066761) fol-
tute of Zoology, Capital Medical University, China. lowing the manufacturer’s instructions. Luciferase activity
was measured using a luminometer (Promega).
SARS-CoV-2-S plasmids and site-directed mutagenesis
SARS-CoV-2-S plasmid was constructed by inserted the Overexpression and knockdown of CTSL
codon-optimized S gene of SARS-CoV-2 (GenBank: Overexpression and knockdown of CTSL gene in Huh7
MN_908947) into pcDNA3.1 as previously described6. For cells were described previously12. Briefly, for CTSL knock-
the site-directed mutagenesis, 15–20 nucleotides before down, Huh7 cells were plated in 48-well plates, and trans-
and after the target mutation site were selected as forward fected with indicated concentrations of siRNAs against
primers, while the reverse complementary sequences were homo CTSL mRNA (si-CTSL) or 50 nM negative control
selected as reverse primers. Following site-directed siRNA (scramble) using Lipofectamine 3000 reagent
mutagenesis PCR, the template chain was digested using (Thermo Fisher Scientific). For CTSL overexpression, Huh7
DpnI restriction endonuclease (NEB, USA). The PCR cells were plated in 48-well plates and transfected with
product was transformed into E. coli DH5α competent indicated concentrations of human CTSL expression plas-
cells; single clones were selected and then sequenced. mid (GenBank: NM_001912, Vigenebio) or 0.2 μg control
plasmid. The overexpression and knockdown efficiencies
Production and quantification of PsVs were validated in both mRNA and protein levels by us
PsVs incorporated with S protein from either SARS-CoV- previously12. Twenty-four hours post-transfection, the
1, SARS-CoV-2, or mutants were constructed using a medium was replaced with a fresh medium. Then the cellsYou can also read

















































