The Osteocyte: From "Prisoner" to "Orchestrator" - MDPI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of
Functional Morphology
and Kinesiology
Review
The Osteocyte: From “Prisoner” to “Orchestrator”
Carla Palumbo * and Marzia Ferretti
Section of Human Morphology, Department of Biomedical, Metabolic and Neural Sciences,
University of Modena and Reggio Emilia, 41124 Modena, Italy; marzia.ferretti@unimore.it
* Correspondence: carla.palumbo@unimore.it
Abstract: Osteocytes are the most abundant bone cells, entrapped inside the mineralized bone matrix.
They derive from osteoblasts through a complex series of morpho-functional modifications; such
modifications not only concern the cell shape (from prismatic to dendritic) and location (along the
vascular bone surfaces or enclosed inside the lacuno-canalicular cavities, respectively) but also their
role in bone processes (secretion/mineralization of preosseous matrix and/or regulation of bone
remodeling). Osteocytes are connected with each other by means of different types of junctions,
among which the gap junctions enable osteocytes inside the matrix to act in a neuronal-like manner,
as a functional syncytium together with the cells placed on the vascular bone surfaces (osteoblasts
or bone lining cells), the stromal cells and the endothelial cells, i.e., the bone basic cellular system
(BBCS). Within the BBCS, osteocytes can communicate in two ways: by means of volume transmission
and wiring transmission, depending on the type of signals (metabolic or mechanical, respectively)
received and/or to be forwarded. The capability of osteocytes in maintaining skeletal and mineral
homeostasis is due to the fact that it acts as a mechano-sensor, able to transduce mechanical strains
into biological signals and to trigger/modulate the bone remodeling, also because of the relevant
role of sclerostin secreted by osteocytes, thus regulating different bone cell signaling pathways. The
authors want to emphasize that the present review is centered on the morphological aspects of the
osteocytes that clearly explain their functional implications and their role as bone orchestrators.
Citation: Palumbo, C.; Ferretti, M.
The Osteocyte: From “Prisoner” to Keywords: osteocytes; bone mechano-sensor; skeletal homeostasis; mineral homeostasis; bone
“Orchestrator”. J. Funct. Morphol. remodeling
Kinesiol. 2021, 6, 28. https://doi.org/
10.3390/jfmk6010028
Academic Editor: Pietro Scicchitano 1. Osteoblast-to-Osteocyte Transformation
It has been known for more than a century [1–3] that the osteocyte originates from the
Received: 5 February 2021
Accepted: 11 March 2021
osteoblast. However, the process of osteoblast-to-osteocyte differentiation has been widely
Published: 17 March 2021
investigated only at a later time point with regard to both morphological and functional
aspects. The structural differences between osteoblasts and osteocytes were shown by vari-
Publisher’s Note: MDPI stays neutral
ous authors in the 1960s to1980s [4–8], but only afterwards was established the temporal
with regard to jurisdictional claims in
sequence of the events that allows the transformation of the prismatic osteoblast (generally
published maps and institutional affil- arranged in laminae facing the vascular bone surfaces) into the dendritic mature osteocyte
iations. (embedded in the mineralized matrix) [9–11]. Concerning the dynamic modification of the
cell body of preosteocyte (i.e., the differentiating osteocyte), the amount of the cytoplasmic
organelles decreases, whereas the nucleus-to-cytoplasm ratio increases depending on the
diminution of its secretive activity [9]. In parallel to both the cellular body reduction in size
Copyright: © 2021 by the authors.
and the modification in ultrastructure, the formation of the cytoplasmic processes proceeds
Licensee MDPI, Basel, Switzerland.
with an asynchronous and asymmetric pattern, considering that the cells in differentiation
This article is an open access article
are progressively further away from the vascular surface due to the osteoid secreted by
distributed under the terms and the osteoblastic lamina (Figure 1): firstly, the differentiating osteocytes maintain contacts
conditions of the Creative Commons with the mature osteocytes that recruited them from the osteoblastic lamina, forming short
Attribution (CC BY) license (https:// “mineral” processes; later, they establish contacts with the migrating osteoblastic lamina,
creativecommons.org/licenses/by/ elongating slender and long “vascular” processes, issued before the complete mineraliza-
4.0/). tion of the surrounding osteoid. The asynchronous and asymmetrical dendrogenesis is the
J. Funct. Morphol. Kinesiol. 2021, 6, 28. https://doi.org/10.3390/jfmk6010028 https://www.mdpi.com/journal/jfmk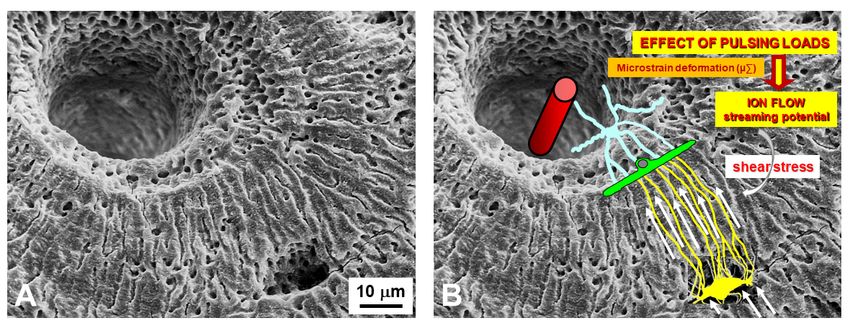
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 2 of 18
J. Funct. Morphol. Kinesiol. 2021, 6, 28 2 of 17
with the migrating osteoblastic lamina, elongating slender and long “vascular” processes,
issued before the complete mineralization of the surrounding osteoid. The asynchronous
and asymmetrical dendrogenesis is the expression of the fact that osteocytes (as all bone
expression of an
cells) live in theasymmetrical
fact that osteocytes (as all bone
environment, cells) live
between thein an asymmetrical
mature mineralized environment,
matrix and
between the mature
the vascular surface mineralized matrix and the
(covered by osteoblastic vascular
laminae surface
or bone (covered
lining by osteoblastic
cells); thus, it is logical
laminae
to expectorthat
bonenotlining
only cells); thus, it isbut
the osteocytes, logical
alsotothe
expect that not only
preosteocytes and the
the osteocytes,
osteoblasts,but
are
also the preosteocytes and the osteoblasts,
morpho‐functionally asymmetric cells. are morpho-functionally asymmetric cells.
Figure 1.
Figure 1. Schematic
Schematic drawing
drawingshowing
showingthetheasynchronous
asynchronousand andasymmetric
asymmetricpattern
patternofof
cytoplasmic
cytoplasmic
processes formation during osteocyte differentiation (preosteocytes = black; osteoblasts = white).
processes formation during osteocyte differentiation (preosteocytes = black; osteoblasts = white).
(A) Preosteocyte enlarges its secretory territory, thus reducing its appositional growth rate, and
(A) Preosteocyte enlarges its secretory territory, thus reducing its appositional growth rate, and
starts to radiate processes towards the osteoid. (B) Preosteocyte, located inside the osteoid seam
starts to in
but still radiate processes
contact with thetowards the osteoid.
osteoblastic (B) Preosteocyte,
lamina, continues located
to radiate inside
short and thecytoplasmic
thick osteoid seam
but
processes only from its mineral‐facing side. (C) Preosteocyte, before being completely cytoplasmic
still in contact with the osteoblastic lamina, continues to radiate short and thick buried by
processes only from
minerals, radiates its and
long mineral-facing side. (C)
slender processes fromPreosteocyte, before being
its vascular‐facing side tocompletely buried
remain in touch by
with
minerals, radiates
the osteoblastic long and slender processes from its vascular-facing side to remain in touch with
lamina.
the osteoblastic lamina.
At the end of the process, the osteocytes are confined to lacuno‐canalicular cavities,
At the end
“prisoners” of the
inside the process, the osteocytes
mineralized are confined
matrix. Despite this fact,tothey
lacuno-canalicular cavities,
are connected, thanks to
“prisoners” inside the
the dendrogenesis mineralized
process, to each matrix.
other Despite
and withthis fact,
the they
bone are on
cells connected, thanks
the vascular to the
surfaces
dendrogenesis
through a network process,
of to each otherrunning
dendrities, and withwithin
the bone
thecells on the vascular
canalicular network; surfaces through
this condition
aallows
network of dendrities, running within the canalicular network; this
osteocytes to act as “orchestrators” of bone processes [12,13]. Prerequisite forcondition allows
that
osteocytes to act as “orchestrators” of bone processes [12,13]. Prerequisite
is the existence of junctional complexes occurring among osteocyte cytoplasmic processes for that is the
existence
[7,14,15],ofsuggesting
junctional complexes
that the boneoccurring among
cells of osteocyte cytoplasmic
the osteogenic processes
lineage, arranged in [7,14,15],
network
suggesting that the bone cells of the osteogenic lineage, arranged in network
(Figure 2), might act as a functional syncytium, that includes also the cells covering (Figure the
2),
might act as a functional syncytium, that includes also the
vascular bone surfaces, bone lining cells [15] or osteoblasts [16]. cells covering the vascular bone
surfaces, bone lining cells [15] or osteoblasts [16].
In conclusion, throughout the whole differentiation process, preosteocytes are always
in close relationship with the neighboring cells (osteoblasts, osteocytes) by means of
variously-shaped intercellular contacts (invaginated finger-like, side-to-side, and end to-
end) and two types of specialized junctions: gap and adherens [14]. The pivotal role played
by these contacts and junctions in osteocyte differentiation and activity will be discussed in
the context of their distinct functional significance.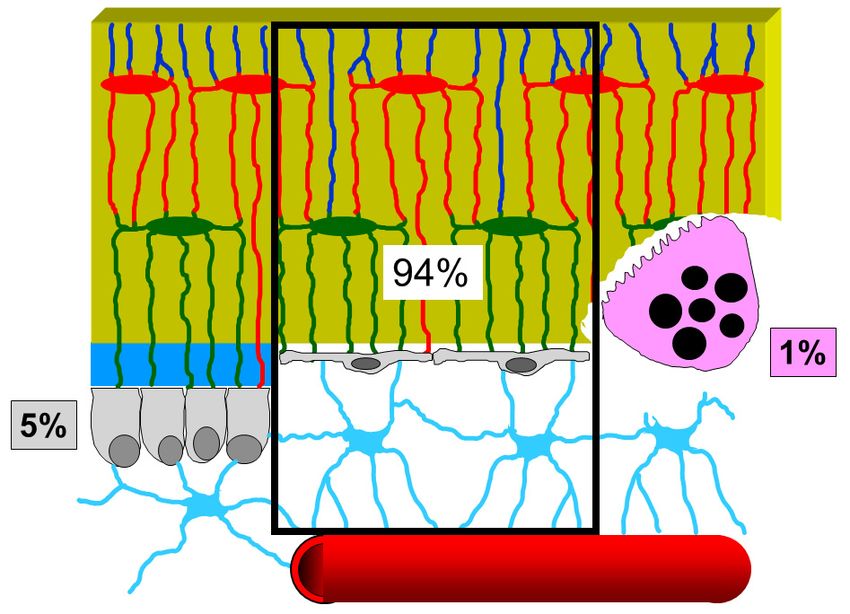
J. Funct. Morphol. Kinesiol. 2021, 6, 28 3 of 17
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 3 of 18
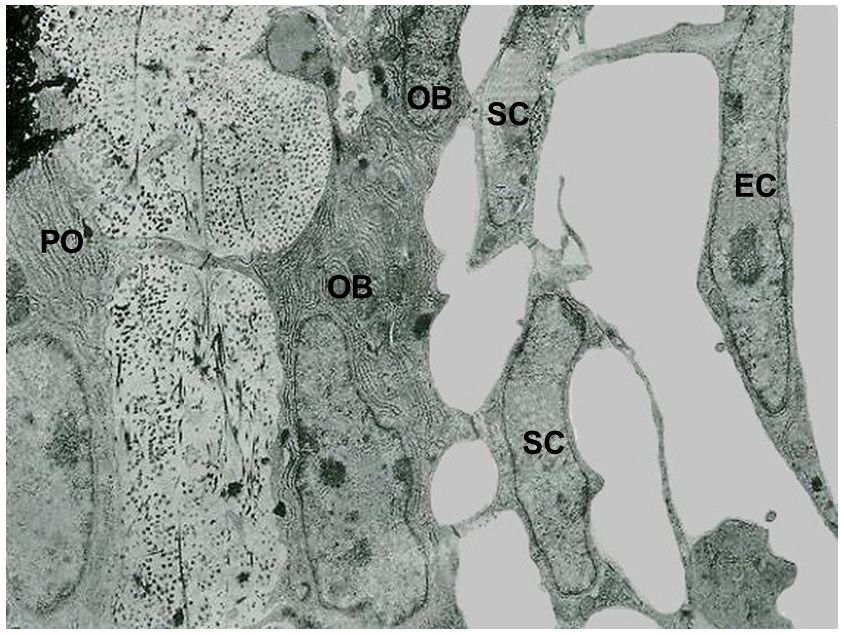
Figure 2.
Figure 2. Transmission
Transmission electron
electron microscope
microscope (TEM)
(TEM) micrograph
micrograph showing
showing the
the continuous
continuous cytoplasmic
cytoplas‐
mic network of the cells of the osteogenic lineage, extending from osteocytes to endothelial cells.
network of the cells of the osteogenic lineage, extending from osteocytes to endothelial cells. (PO
(PO preosteocyte, OB osteoblast, SC stromal cell, EC endothelial cell). ×22,500.
preosteocyte, OB osteoblast, SC stromal cell, EC endothelial cell). ×22,500.
In conclusion,
2. Osteogenesis throughout
Processes the wholeMorphology
and Osteocyte differentiation process, preosteocytes are al‐
ways in close relationship with the neighboring cells (osteoblasts, osteocytes) by means of
To fully understand the osteocyte morphology in the different contexts in which it
variously‐shaped intercellular contacts (invaginated finger‐like, side‐to‐side, and end to‐
originates, it is important to mention that morphological studies have shown that osteocyte
end) and two types of specialized junctions: gap and adherens [14]. The pivotal role
differentiation is quite different in two types of osteogenesis. We clearly demonstrated,
played by these contacts and junctions in osteocyte differentiation and activity will be
for the first time in 2002 in our labs, that not all osteoblasts are arranged in the well-
discussed in the context of their distinct functional significance.
known movable laminae: the two types of osteogenesis were named, during our studies
on intramembranous ossification, “static bone formation” or static osteogenesis (SO) and
2. Osteogenesis Processes and Osteocyte Morphology
“dynamic bone formation” or dynamic osteogenesis (DO) [17–21]. SO and DO occur in
temporalTo fully
sequenceunderstand
and dependthe osteocyte
on whether morphology in the different
or not osteoblast movement contexts
occurs.inWewhich
pointedit
originates, it is important to mention that morphological studies
out that the well-known dynamic osteogenesis (DO), performed by migrating osteoblast have shown that osteo‐
cyte differentiation
laminae, is precededis by quite different
static in two (SO)
osteogenesis typesinofwhich,
osteogenesis. We clearly
at the onset demon‐
of ossification,
strated, for the first time in 2002 in our labs, that not all osteoblasts
cords of stationary osteoblasts (instead of laminae of movable osteoblasts) transform are arranged in the
well‐known movable laminae: the two types of osteogenesis
into osteocytes in the same site where they differentiated from mesenchymal progenitorwere named, during our
studies on intramembranous ossification, “static bone formation”
elements. In SO (Figure 3A,B), stationary osteoblasts (acting in sites where no bone pre- or static osteogenesis
(SO) and
exists) are “dynamic bone formation”
in very peculiar and unusualorconditions
dynamic osteogenesis
with respect to (DO)
the [17–21].
well-known SO and DO
dynamic
occur in temporal sequence and depend on whether or not osteoblast
osteoblasts, arranged in laminae and secreting bone close to the preexistent one: i.e., movement occurs.
We pointed
they are in aout that the well‐known
symmetrical environment dynamic
sinceosteogenesis
the osteoblast (DO), performed
cords by migrating
differentiate at about
osteoblast laminae, is preceded by
half distance between two vessels; in this extent, several factors are involved,ofsuch
static osteogenesis (SO) in which, at the onset ossifi‐as
endothelial-cell-derived cytokines (Endothelin-1) and growth factors (EDGF, PDGF).trans‐
cation, cords of stationary osteoblasts (instead of laminae of movable osteoblasts) In fact,
formgive
they intoriseosteocytes in the same
to big globous site where
osteocytes, oftenthey differentiated
located from mesenchymal
inside confluent lacunae, with pro‐short
genitor
and elements.dendrites
symmetrical In SO (Figure
that 3A,B), stationary
can radiate osteoblastsfrom
simultaneously (acting
allin sites where
around no bone
their cell body
pre‐exists)
because theyare areincompletely
very peculiar and unusual
surrounded by theconditions
unmineralizedwith matrix.
respect In to DO
the (Figure
well‐known3C,D),
dynamicosteoblasts
instead, osteoblasts,transform
arrangedintoin laminae and secreting bone close
small ovoidal/ellipsoidal spideryto the preexistent
osteocytes insideone:
an
i.e., they are in a symmetrical environment since the osteoblast
asymmetrical environment, whose dendrites form in an asynchronous and asymmetrical cords differentiate at about
half distance
manner between two
in concomitance vessels;
with, and in this extent,
depending on,several factors are
the advancing involved, such
mineralizing as en‐
surface and
dothelial‐cell‐derived
the migrating osteogenic laminae. cytokines (Endothelin‐1) and growth factors (EDGF, PDGF). In fact,
they give rise to big globous osteocytes, often located inside confluent lacunae, with short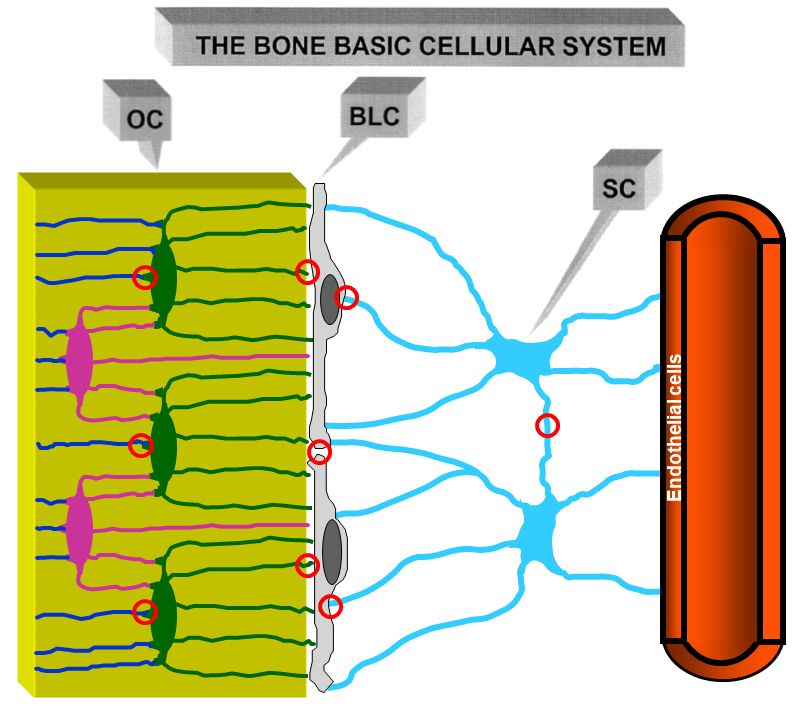
and symmetrical dendrites that can radiate simultaneously from all around their cell body
because they are completely surrounded by the unmineralized matrix. In DO (Figure
3C,D), instead, osteoblasts transform into small ovoidal/ellipsoidal spidery osteocytes in‐
J. Funct. Morphol. Kinesiol. 2021, 6, 28 side an asymmetrical environment, whose dendrites form in an asynchronous and asym‐ 4 of 17
metrical manner in concomitance with, and depending on, the advancing mineralizing
surface and the migrating osteogenic laminae.
Figure 3. Schematic drawing showing the occurrence, in sequence, of static (SO) and dynamic
Figure 3. Schematic drawing showing the occurrence, in sequence, of static (SO) and dynamic (DO)
(DO) osteogenesis. In SO, stationary osteoblast cords (A) differentiate at about half distance be‐
osteogenesis. In SO, stationary osteoblast cords (A) differentiate at about half distance between two
tween two vessels, giving rise to big globous osteocytes (B), often located inside confluent lacunae,
vessels, giving
with short andrise to big globous
symmetrical osteocytes
dendrites. In DO,(B), often located
osteoblastic inside
laminae confluent lacunae,
differentiate with short
on the surface of
and symmetrical
SO‐derived dendrites.
trabeculae In DO,
(C) and osteoblastic
transform laminae
into small differentiate
spidery osteocyteson(D)
thewhose
surface of SO-derived
asymmetrical
trabeculae (C) and
dendrites form transform intowith
in concomitance smallthe
spidery osteocytes
migration (D) whose lamina.
of the osteogenic asymmetrical dendrites
See text form
for explana‐
in concomitance
tion. with the migration
(EDGF: endothelial‐derived of the
growth osteogenic
factor; PDGF: lamina. See textgrowth
platelet‐derived for explanation.
factor). (EDGF:
endothelial-derived growth factor; PDGF: platelet-derived growth factor).
Among all osteocytes (both SO‐ and DO‐derived) and between osteocytes and oste‐
Among
oblasts, all contacts
simple osteocytes
and(both SO- and
specialized gapDO-derived)
junctions were and observed,
between osteocytes
that allow and os-
all bone
teoblasts, simple contacts and specialized gap junctions were observed, that
cells to remain functionally connected; therefore, a continuous osteocyte network extends allow all bone
cells to remain
throughout thefunctionally connected;
bone, regardless of its therefore, a continuous
static or dynamic origin.osteocyte network
This cellular extends
network has
throughout the bone, regardless of its static or dynamic origin. This cellular
the characteristics of a functional syncytium, potentially capable of modulating, by wiring network has
the characteristics
transmission (see of a functional
below), syncytium,
the cells potentially
of the osteogenic capable
lineage of modulating,
covering the bonebysurfaces.
wiring
transmission (see below), the cells of the osteogenic lineage covering the
Morphology, as well as density and distribution of osteocytes, is not only related to the bone surfaces.
Morphology, as well as
type of osteogenesis density
from whichand distribution
they derive butof osteocytes,
also is not only
to the collagen related
texture whereto they
the
type of osteogenesis from which they derive but also to the collagen texture
are located, on whose spatial organization the osteocytes play a pivotal role [22]. Unlike where they
are
whatlocated, on whose spatial
was traditionally organization
reported in textbooks the
inosteocytes
relation to play
boneahistology,
pivotal role [22].
about Unlike
3 decades
what was traditionally reported in textbooks in relation to bone histology, about 3 decades
ago Marotti and collaborators reworked the classification of bone tissues, demonstrating,
ago Marotti and collaborators reworked the classification of bone tissues, demonstrating,
by means of both SEM (scanning electron microscopy) and TEM (transmission electron
by means of both SEM (scanning electron microscopy) and TEM (transmission electron
microscopy) analyses, that woven bone can be arranged in two different tissues [10,22–
microscopy) analyses, that woven bone can be arranged in two different tissues [10,22–24]:
24]: “not lamellar woven bone” mostly formed by SO and “lamellar woven bone” mostly
“not lamellar woven bone” mostly formed by SO and “lamellar woven bone” mostly
formed by DO. Comparing the two type of tissue (Figure 4): in not‐lamellar‐woven‐bone
formed by DO. Comparing the two type of tissue (Figure 4): in not-lamellar-woven-bone
(Figure 4A), osteocytes are symmetric, randomly distributed in clusters, with a large and
(Figure 4A), osteocytes are symmetric, randomly distributed in clusters, with a large and
irregularly globous shape of the cell body; in lamellar‐woven‐bone (Figure 4B) (made up
irregularly globous shape of the cell body; in lamellar-woven-bone (Figure 4B) (made
of alternate dense and loose lamellae, both with an interwoven fibrous texture), osteocytes
up of alternate dense and loose lamellae, both with an interwoven fibrous texture), os-
are asymmetric, distributed in planes exclusively located inside loose lamellae, with an
teocytes are asymmetric, distributed in planes exclusively located inside loose lamellae,
with an almond-like shape of the cell body (i.e., a triaxial ellipsoid) [10,25–28]. It is to be
underlined that, generally, morphology of osteocytes is indirectly inferred from the shape
of the lacuno-canalicular network in which they are enclosed rather than directly from
their protoplasm.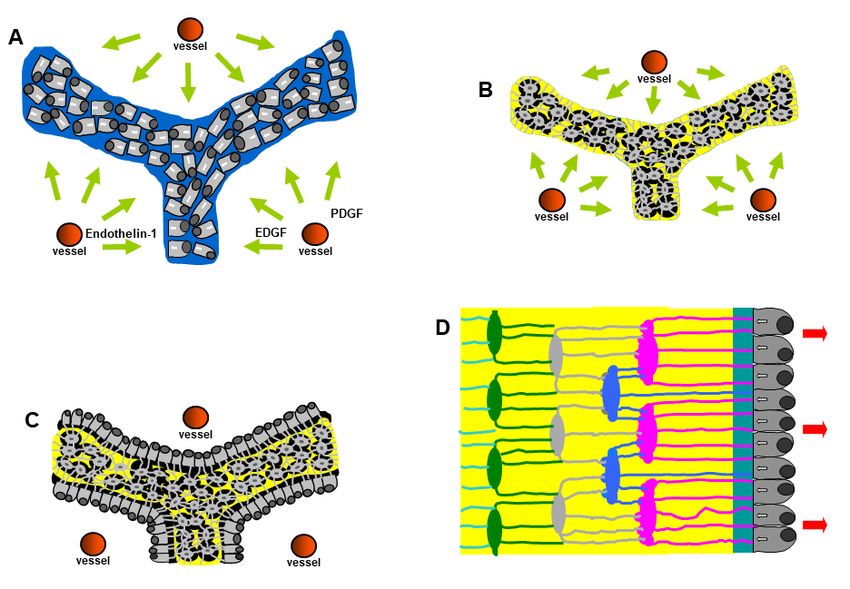
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 5 of 18
almond‐like shape of the cell body (i.e., a triaxial ellipsoid) [10,25–28]. It is to be under‐
J. Funct. Morphol. Kinesiol. 2021, 6, 28 lined that, generally, morphology of osteocytes is indirectly inferred from the shape 5ofofthe
17
lacuno‐canalicular network in which they are enclosed rather than directly from their pro‐
toplasm.
Figure4.4.(A,B):
Figure (A, B):Schematic
Schematicdrawing
drawingshowing
showingarrangement
arrangementofofosteocytes
osteocytesand
andthe
thesurrounding
surroundingcolla-
col‐
lagen fibers in not‐lamellar‐woven‐bone (A) and in lamellar‐woven‐bone (B). In both types of bone
gen fibers in not-lamellar-woven-bone (A) and in lamellar-woven-bone (B). In both types of bone
tissue, osteocyte lacunae (blue ovals) are surrounded by perilacunar matrices of loosely arranged
tissue, osteocyte lacunae (blue ovals) are surrounded by perilacunar matrices of loosely arranged
collagen fibers. In lamellar‐woven‐bone, the loose lamellae result as a consequence of alignment
collagen
and fusion fibers. Inperilacunar
of the lamellar-woven-bone, theofloose
loose matrices lamellae result
the osteocytes as a in
arranged consequence
planes; the of alignment
dense bun‐
and
dlesfusion of thefibers
of collagen perilacunar
do notloose matrices
contain of the(C,
osteocytes. osteocytes
D): SEMarranged in planes;
micrographs the dense bundles
of not‐lamellar‐woven‐
of collagen
bone (C) andfibers do not contain osteocytes.
lamellar‐woven‐bone (C,D): SEM
(D); the osteocyte micrographs
lacunae of not-lamellar-woven-bone
are larger, more numerous, and ir‐
regularly
(C) distributed in not‐lamellar‐woven‐bone
and lamellar-woven-bone with respect
(D); the osteocyte lacunae to lamellar‐woven‐bone,
are larger, where
more numerous, and irregularly
they are only
distributed located inside loose lamellae.
in not-lamellar-woven-bone withNote thatto
respect dense lamellae, alternating
lamellar-woven-bone, withthey
where the loose
are only
ones, are
located thinner
inside and
loose do not contain
lamellae. osteocyte
Note that lacunae. alternating with the loose ones, are thinner
dense lamellae,
and do not contain osteocyte lacunae.
But we can ask: “was the egg or the hen born first?” that is to wonder if osteocyte
affects
Butthewecollagen
can ask:texture“was orthevice‐versa
egg or thethe hencollagen texturethat
born first?” modifies the cell morphology
is to wonder if osteocyte
and arrangement?
affects The factorthat
the collagen texture lamellarthe
vice-versa bone appears
collagen to bemodifies
texture a varietythe of cell
woven bone and
morphology
thatarrangement?
and osteocytes areThe located in loose
fact that lamellae
lamellar only hastosuggested
bone appears be a variety that
of the
wovendifferences
bone and in
that osteocytes
texture betweenare located in loose
lamellar‐woven andlamellae only has suggested
not‐lamellar‐woven bone depend that the differences
on the in
distribution
texture between
of osteocytes lamellar-woven
throughout the bone andmatrix,
not-lamellar-woven
that is to say, bone
on the depend
manner onofthe distribution
recruitment of
of
theosteocytes throughout the osteoblasts
osteocyte‐differentiating bone matrix, thatthe
from is to say, on the
osteogenic manner
laminae of recruitment
[22,23,29]. of
In not‐la‐
the osteocyte-differentiating
mellar‐woven‐bone (generallyosteoblasts
laid downfrom verythe osteogenic
rapidly) laminae [22,23,29].
the osteoblasts In not-
that differentiate
lamellar-woven-bone (generally laid down very rapidly) the osteoblasts
into osteocytes are recruited haphazardly and “enter” the bone in a random fashion. As a that differentiate
into osteocytes
result, such type areofrecruited haphazardly
not‐lamellar‐woven andconsists
bone “enter” oftheanbone in a random
irregular fashion.
distribution As
of oste‐
aocyte‐rich
result, such type of not-lamellar-woven bone consists of an irregular
areas where the collagen is loosely arranged (since it corresponds to that of the distribution of
osteocyte-rich areas where
perilacunar matrices) the 4A,C).
(Figure collagen is loosely arranged (since it corresponds to that of
the perilacunar matrices) (Figure
In lamellar‐woven‐bone, 4A,C).
whose matrix is usually laid down at a lower rate than that
In lamellar-woven-bone, whose
of not‐lamellar‐woven‐bone, the recruitment matrix is ofusually laid down
the osteoblasts thatatdifferentiate
a lower rate than
into os‐
that of not-lamellar-woven-bone, the recruitment of the osteoblasts
teocytes occurs in an orderly manner. Since the cellular lamellae are only those formed that differentiate into
by
osteocytes occurs
loose texture, thisin suggests
an orderlythatmanner. Since the cellular
the osteoblasts committedlamellae are only those formed
to differentiating by
into osteo‐
loose
cytestexture, this suggests
are recruited that thegroups
in successive osteoblasts committed
and that to differentiating
those pertaining to eachinto
grouposteocytes
are dis‐
are recruited in successive groups and that those pertaining to each
tributed in a single plane, namely that corresponding to a loose lamella. Thus, loose la‐ group are distributed
in a single
mellae plane,
could namely
simply formthat
as corresponding to a loose lamella.
a result of the alignment and fusion Thus,of loose lamellae could
the loosely‐arranged
simply form as a result of the alignment and fusion of the loosely-arranged fibers pertaining
to the loose perilacunar matrices of the osteocytes they contain (Figure 4B,D).
Moreover, van Oers and coworkers [28] showed interactions between osteocyte shape
and mechanical loading. In particular, their correlation seems to be related to the orientation
of collagen fibers during osteoblast bone deposition: collagen gives bone its tensile strength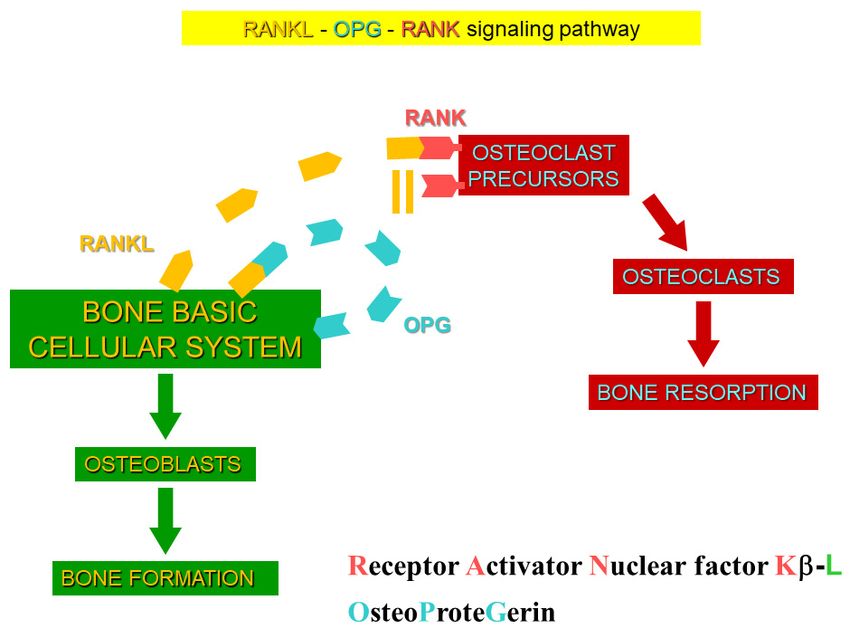
J. Funct. Morphol. Kinesiol. 2021, 6, 28 6 of 17
which, in turn, conditions the elongation of osteocyte axes in relation to the direction
of stress lines. These interactions are twofold: firstly, as regards the mechanical loading
affecting the shape of osteocyte lacunae; secondly, as regards the shape of osteocytes
influencing their mechano-sensibility and subsequent control of bone remodeling (see
below). Shape variations of the lacuna hosting the osteocyte will also alter the direct cell
strain, as well as the fluid flow and, in case, the microdamage inputs to the osteocyte.
Furthermore, the shape of the osteocyte cell body affects its sensitivity to these inputs [30].
3. Osteocyte Network and Communications
Morphological investigations [31,32] showed that an osteogenic cellular system is
present inside the bone, formed by various cell types from depth to the vascular surfaces:
osteocytes, osteoblasts, or bone lining cells (according to whether the bone surfaces are in
growing or resting phase, respectively), stromal cells, and endothelial cells. They constitute
a functional syncytium whose cells play different roles and have different relationships
with the surrounding environment. Osteocytes are enclosed inside bone microcavities
filled with the bone fluid compartment (BFC); the stromal cells, extending from vascular
endothelia to the cells carpeting the bone surfaces, are bathed by the perivascular interstitial
fluid (PIF); osteoblasts and/or bone lining cells, located at the interface “mineralized ma-
trix/vascular surfaces”, separate the two different fluid compartments and are in contacts
both with the “deep” and “superficial” cell populations. Intercellular contacts (simple
contacts and/or specialized junctions) were observed throughout all cells of the osteogenic
system [11,14]. Thus, these cells form a continuous protoplasmic network that extends
from the osteocytes to the vascular endothelia [33]. The presence of gap junctions (allowing
electric coupling) suggests that the bone osteogenic cells could be considered a functional
syncytium regulated not only by diffusion through the intercellular fluids (volume trans-
mission) but also by signals issued through the cytoplasmic network and driven through
cytoplasmic extentions (wiring transmission) (see below).
In the resting phase (namely, when no bone formation/erosion occur massively)
osteocytes, bone lining cells, and stromal cells were named, as a whole, the bone basic
cellular system (BBCS) (Figure 5) [31,32], because they represent the only permanent
cellular background capable of perceiving mechanical strains and biochemical factors and
then of triggering and driving both processes of bone formation and bone resorption.
It is appropriate to underline, in this context, that the majority of researchers consider
osteoblasts and osteoclasts as the only relevant bony cells. Here, the authors want to
emphasize that such a view is incorrect and misleading: indeed, osteoblasts and osteoclasts
(the bone forming and bone reabsorbing cells, respectively) are transient cells, thus they
cannot be the first to be involved, in the resting phase, in sensing skeletal requirements
which modulate bone processes. Briefly, according to our view, osteoblasts and osteoclasts
represent the “arms” of a worker; the actual “operation center” (i.e., the “brain”) is con-
stituted by the cells of the osteogenic lineage in the resting phase (BBCS), particularly the
osteocytes. (Figure 6) [34]. As shown by our previous studies, signaling throughout BBCS
can occur by volume transmission (VT) and/or wiring transmission (WT) [31,32,35,36]. VT
corresponds to the routes followed by soluble substances (hormones, cytokines, growth
factors), whereas WT represents the diffusion of ionic currents along cytoplasmic processes
in a neuron-like manner. It is likely that biochemical signals first affect stromal cells em-
bedded in PIF and diffuse by VT to reach the other cells of BBCS, whereas mechanical
agents are firstly sensed by osteocytes bathed by BFC inside the mineralized matrix and
then issued throughout BBCS by WT (Figure 7).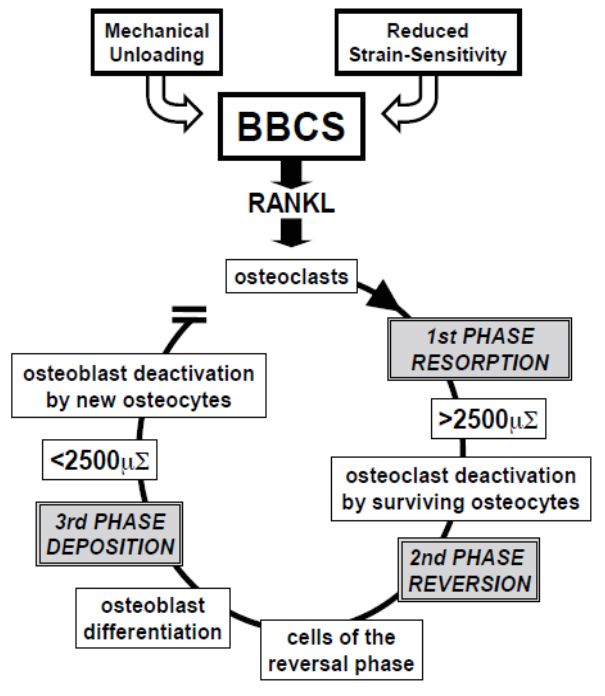
J. Funct. Morphol. Kinesiol. 2021, 6, 28 7 of 17
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 7 of 18
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 7 of 18
Figure5.5.Schematic
Figure Schematicdrawing
drawingofofthethecells
cellsofofthe
theosteogenic
osteogenic lineage
lineage inin
thethe resting
resting phase,
phase, thethe Bone
Bone Basic
Basic Cellular System.
Figure 5.System.
Schematic From
drawing left to right: osteocytes
of theosteocytes
cells of the(OC), (OC),
osteogenic bone lining
lineage in thecells (BLC),
resting stromal
phase, cells
the(SC)
Boneand
Cellular From left to right: bone lining cells (BLC), stromal cells
(SC)
Basicand one vascular
Cellular System. capillary.
From left This cell osteocytes
to right: network forms
(OC),a bone
functional
liningsyncytium
cells (BLC),since cellscells
stromal are
one vascular capillary. This cell network forms a functional syncytium since cells are joined by gap
joined by one
(SC) and gapvascular (red circles).
junctionscapillary. This cell network forms a functional syncytium since cells are
junctions (red circles).
joined by gap junctions (red circles).
Figure 6. Schematic drawing showing the percentage of the various bone cells present in the bone
resting phase:
Figure 6. 94% BBCS,
Schematic 5% showing
drawing osteoblasts,
theand 1% osteoclasts
percentage (data from
of the various boneParfitt [34]). See
cells present intext
the for
bone
Figure 6. Schematic drawing showing the percentage of the various bone cells present in the bone
explanation.
resting phase: 94% BBCS, 5% osteoblasts, and 1% osteoclasts (data from Parfitt [34]). See text for
resting phase: 94% BBCS, 5% osteoblasts, and 1% osteoclasts (data from Parfitt [34]). See text
explanation.
for explanation.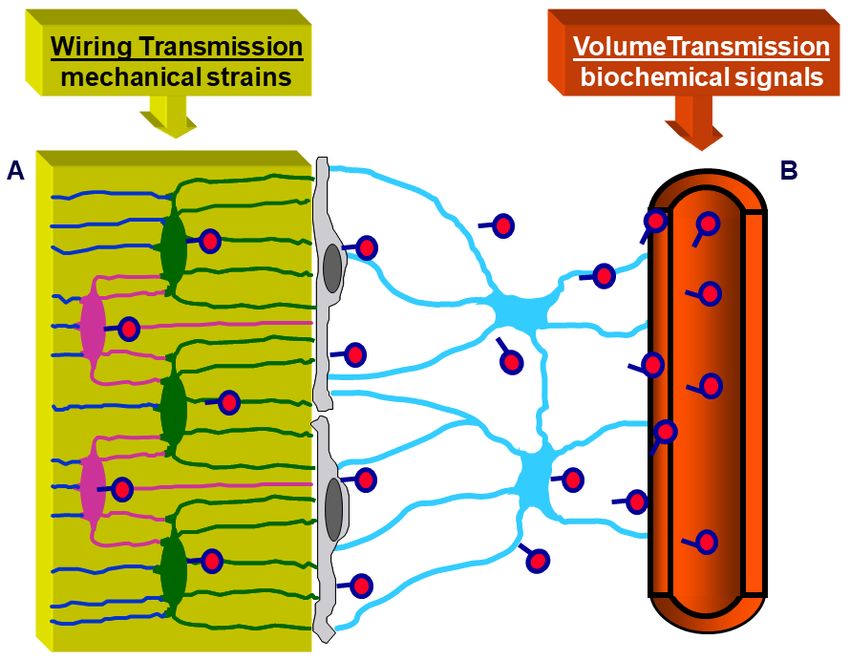
J. Funct. Morphol. Kinesiol. 2021, 6, 28 8 of 17
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 8 of 18
Figure 7. Schematic picture of the BBCS, in which signal transmissions can occur by volume trans‐
Figure 7. Schematic picture of the BBCS, in which signal transmissions can occur by volume trans-
mission (VT) and/or wiring transmission (WT): (A) VT corresponds to the routes followed by solu‐
mission (VT) and/or wiring transmission (WT): (A) VT corresponds to the routes followed by soluble
ble substances; (B) WT represents the diffusion of ionic currents along cytoplasmic processes. Bio‐
substances; (B) WT
chemical signals representsdiffusing
(hormones the diffusion
fromof ionic
the currents
blood, alongetc.)
cytokines, cytoplasmic processes.
first affect Biochemi-
stromal cells and
cal
diffuse by VT to reach the other cells of BBCS, whereas mechanical agents are firstly senseddiffuse
signals (hormones diffusing from the blood, cytokines, etc.) first affect stromal cells and by
by VT to reach
osteocytes andthe
thenother cells
issued of BBCS, whereas
throughout BBCS bymechanical
WT. agents are firstly sensed by osteocytes
and then issued throughout BBCS by WT.
It should be noted that besides WT, other similarities do exist between osteocytes and
It should
neurons. be noted
The short and that
thickbesides
mineralWT, other similarities
cytoplasmic processesdoofexist between
osteocytes osteocytes
resemble neu‐
and neurons. The short and thick mineral cytoplasmic processes
ronal dendrites, whereas the long and slender vascular cytoplasmic processes are of osteocytes resemble
similar
neuronal
to neuronal dendrites, whereas theoflong
axons. Transmission andthrough
signals slenderosteocytes
vascular cytoplasmic
seems to occur processes are
by gap junc‐
similar to neuronal axons. Transmission of signals through osteocytes
tions instead of synapses, though it has been recently shown that osteocytes produce typ‐ seems to occur
by
icalgap junctions insteadsuch
neurotransmitters, of synapses,
as nitric though it has been
oxide [37–40] recently shownE2
and prostaglandin that osteocytes
[38,41]. Addi‐
produce typical neurotransmitters, such as nitric oxide [37–40] and prostaglandin E2 [38,41].
tionally, about two decades ago we provided evidence that WT occurs along osteocytes
Additionally, about two decades ago we provided evidence that WT occurs along osteocytes
in amphibian and murine cortical bone depending on loading [42,43], demonstrating that
in amphibian and murine cortical bone depending on loading [42,43], demonstrating that
osteocytes and bone lining cells are at the origin of ionic currents, by operating as a cellular
osteocytes and bone lining cells are at the origin of ionic currents, by operating as a cellular
membrane partition which regulates the ionic flows between bone (BFC) and plasma
membrane partition which regulates the ionic flows between bone (BFC) and plasma (PIF).
(PIF). Such osteocyte ionic currents are constantly directed to the bone lining cells and
Such osteocyte ionic currents are constantly directed to the bone lining cells and stromal
stromal cells during the resting phase.
cells during the resting phase.
As regards the transmission of mechanical signals, both recent and less recent litera‐
As regards the transmission of mechanical signals, both recent and less recent lit-
ture indicates the osteocyte as the main strain‐sensitive bone cell [44–58]. We have shown
erature indicates the osteocyte as the main strain-sensitive bone cell [44–58]. We have
[42,43] that shear stress‐activated osteocytes are capable of steadily increasing and main‐
shown [42,43] that shear stress-activated osteocytes are capable of steadily increasing and
taining the basal current produced by the ionic fluxes (streaming potentials), which occur
maintaining the basal current produced by the ionic fluxes (streaming potentials), which
inside the lacuno‐canalicular
occur inside the lacuno-canalicular microcavities
microcavitiesin in
response
responsetotopulsing
pulsing mechanical loading
mechanical loading
(Figure 8). Briefly, the fact that all osteocytes take part in the formation
(Figure 8). Briefly, the fact that all osteocytes take part in the formation of a potential of a potential os‐
teocyte syncytium
osteocyte syncytiumsupports
supportsthetheview
viewthat
that mechanical
mechanical signals throughout bone
signals throughout bone cells
cellsare
are
mainly issued by wiring transmission since volume transmission does
mainly issued by wiring transmission since volume transmission does not need cell con- not need cell con‐
tactsin
tacts inorder
orderto tooccur.
occur.AAfunctional
functionalsyncytium
syncytiumas assuch
suchisisable
ableto
toinitiate,
initiate,perform
performand andstop
stop
bone remodeling in order to meet current metabolic and mechanical
bone remodeling in order to meet current metabolic and mechanical bone requirements. bone requirements.
In this
In this context,
context, the theosteocyte
osteocyterole,
role,played
playedininbone
boneadaptation
adaptation to mechanical
to mechanical stimuli, is well
stimuli, is
explained
well explained by theby“Mechanostat
the “MechanostatSetpoint Theory”
Setpoint (Figure
Theory” 9), according
(Figure to which
9), according the oste‐
to which the
ocytes are are
osteocytes likely capable
likely of of
capable modulating
modulating bone
bone resorption
resorption andandbone
boneformation
formationwithin
withina
range of physiological mechanical strains whose upper and lower
a range of physiological mechanical strains whose upper and lower limits are named limits are named set‐
points. When the lower (about 50 μ∑) and upper (about 2500 μ∑) setpoint values are ex‐
ceeded bone resorption and bone formation, respectively, are triggered. [44,59–61].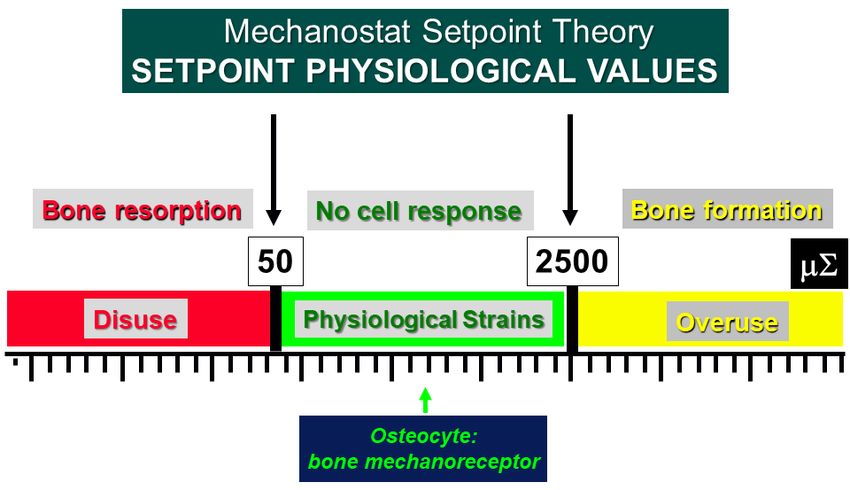
J. Funct. Morphol. Kinesiol. 2021, 6, 28 9 of 17
unct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 9o
setpoints. When the lower (about 50 µ∑) and upper (about 2500 µ∑) setpoint values are
exceeded bone resorption and bone formation, respectively, are triggered [44,59–61].
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 9 of 18
Figure 8. SEM micrograph of a portion of an osteon (A) showing the lacuno-canalicular cavities, on which the bone cellular
Figure 8. SEM micrograph
Figure 8. SEM of a portion of anofosteon (A)(A)
showing the lacuno‐canalicular cavities, on the
which the bone cellular
system has beenmicrograph
superimposed of a(in
portion an osteon
(B)), to explain showing
the events the lacuno‐canalicular
occurring during mechanical cavities, onresponse
stress: in which tobone cellular
mechanical
system hassystem
been superimposed (in B),(in
toB),
explain thethe
events occurring during mechanical stress: in response to mechanical
loading, has
bonebeen superimposed
micro-deformations to explain
(induced by loading events occurring
on bone) during
produce mechanical
ionic stress:
fluxes (streaming in potentials)
response toinmechanical
the bone
loading, bone micro‐deformations
loading, bone micro‐deformations (induced
(inducedby loading
by loadingon onbone) produce
bone) produce ionic
ionic fluxes
fluxes (streaming
(streaming potentials)
potentials)
fluid compartment that, in turn, activate the osteocytes via shear stress. Osteocytes, by means of wiring transmission, issue
in the bone
in the bone
fluid compartment
fluid compartment that, in that, inactivate
turn, turn, activate
the the osteocytes
osteocytes viaviashear
shear stress.
stress. Osteocytes,
Osteocytes, by means
by of wiring
means of transmission,
wiring issue
transmission, issue
signals to the cells covering the bone surface, thus activating osteoblast recruitment and bone formation. Vessel (red cylinder
signals to the cells covering the bone surface, thus activating osteoblast recruitment and bone formation. Vessel (red cyl‐
signals to inside
the cells coveringcanal);
the bone surface, thus activating osteoblast recruitment and bone formation. Vessel (red cyl‐
inder the Haversian
inside stromal
the Haversian canal); cell (light
stromal cellblue);
(lightbone
blue);lining
bonecell (green);
lining osteocyte
cell (green); (yellow).
osteocyte (yellow).
inder inside the Haversian canal); stromal cell (light blue); bone lining cell (green); osteocyte (yellow).
Figure9.9. Schematic
Figure Schematic illustration
illustration of
ofFrost’s
Frost’sMechanostat
MechanostatSetpoint
SetpointTheory.
Theory.The
Thesetpoint values
setpoint areare
values
expressed in microstrains (μ∑) and the bone mechanoreceptors (osteocytes) are activated only
expressed in microstrains (µ∑) and the bone mechanoreceptors (osteocytes) are activated only when
when the physiological setpoints are exceeded. In the “disuse” window, bone is lost owing to in‐
the physiological setpoints are exceeded. In the “disuse” window, bone is lost owing to increased
creased resorption. In the “overuse” window, bone is gained owing to increased bone formation.
resorption. In the “overuse” window, bone is gained owing to increased bone formation.
Figure 9. Schematic illustration of Frost’s Mechanostat Setpoint Theory. The setpoint values are
4.4. Osteocytes
Osteocytesas
asBone
BoneMechanical
MechanicalSensors
Sensorsand
andTransducers
TransducersofofMechanical
MechanicalStrains
Strainsinto
into
expressed in microstrains
Biological Signals (μ∑) and the bone mechanoreceptors (osteocytes) are activated only
Biological Signals
when the physiological setpoints are exceeded. In the “disuse” window, bone is lost owing to in
Lozupone
Lozupone and
and coworkers
coworkers [62]
[62] showed
showed in
in vitro
vitrothat
thatthe
thenumber
number of
ofgap
gapjunctions
junctions be-
be‐
creased resorption. In the “overuse” window, bone is gained owing to increased bone formation
tween osteocytes increases in answer to the mechanical load applied to bone segments
tween osteocytes increases in answer to the mechanical load applied to bone segments
placed
placedin in cultures. This is
cultures. This isin
inline
linewith
withthethesuggestion
suggestionthat that gap
gap junctions
junctions play
play anan impor-
important
4. Osteocytes
tant
rolerole
bothbothascell‐to‐cell
in Bone Mechanical
in cell-to-cell Sensors
communications
communications and
and and
in inTransducers
cell ofenabling
cell synchronization,
synchronization, Mechanical
enabling Strains in
smallsmall
mol‐
molecules
Biological to be
ecules Signals
to be exchanged
exchanged between
between thecoupled
the coupledcells
cells[11,14–16,63];
[11,14–16,63];itit is
is also
also in
in line
line with
withthe
the
fact that osteocytes are considered as mechanoreceptors, able to give rise
fact that osteocytes are considered as mechanoreceptors, able to give rise to the transduc‐to the transduction
Lozupone
mechanical and coworkers
strains into biological [62] showed
stimuli. in vitro
LM data that the number ofnumber
gap junctions
tion mechanical strains into biological stimuli. LMalsodatademonstrated
also demonstrated that the
that of
the num‐
tweenberosteocytes
of viable osteocytes is higher in loaded bones than in the control unloaded ones; TEMsegme
increases in answer to the mechanical load applied to bone
placedanalyses
in cultures.
showedThisthat is
theinintermittent
line with compressive
the suggestion loadingthat
notgap
onlyjunctions playfunc‐
exerts a trophic an importa
role both in cell‐to‐cell communications and in cell synchronization, enabling
tion on osteocytes, probably improving the fluid flows inside the canalicular network small m
[64],
eculesbut
toalso stimulates their
be exchanged metabolic
between theactivity,
coupled particularly their protein and/or
cells [11,14–16,63]; glycoprotein
it is also in line with tJ. Funct. Morphol. Kinesiol. 2021, 6, 28 10 of 17
viable osteocytes is higher in loaded bones than in the control unloaded ones; TEM analyses
showed that the intermittent compressive loading not only exerts a trophic function on
osteocytes, probably improving the fluid flows inside the canalicular network [64], but also
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 10 of 18
stimulates their metabolic activity, particularly their protein and/or glycoprotein synthesis,
as shown by the increment of rough endoplasmic reticulum, thus suggesting that these
molecules are secreted in answer to mechanical stimuli.
Moreover,Rubinacci
Moreover, Rubinacciand andcoworkers
coworkers [42]
[42] showed
showed thatthat
the the damage‐generated
damage-generated ionicionic
cur-
currents
rents theythey observed
observed in cortex
in the the cortex
of frogof frog metatarsal
metatarsal bonesbones
had ahad a cellular
cellular origin. origin. Meta‐
Metatarsal
tarsal bones
bones of adult of amphibian
adult amphibian were purposely
were purposely selected
selected for thefor the study
study becausebecause
their their
shaftsshafts
(un-
(unlike
like the the mammalian
mammalian one)one) contain
contain onlyonly
the the osteocyte‐bone
osteocyte-bone lining
lining cellcell system,
system, thusthus giv‐
giving
the
ing first convincing
the first evidence
convincing that that
evidence damage-generated
damage‐generated ionicionic
currents are generated
currents are generatedby theby
osteocyte-bone lining cell system, i.e., the actual responsible “entity” of bone
the osteocyte‐bone lining cell system, i.e., the actual responsible “entity” of bone adapta‐ adaptation to
both mechanical
tion to loading and
both mechanical mineral
loading andhomeostasis. Later, the Later,
mineral homeostasis. same authors
the same [43]authors
confirmed[43]
the observations
confirmed also in murine
the observations alsomodels.
in murine models.
Recently
Recently [58],
[58], as
as far
far as
as the
the bone
boneadaptation-related
adaptation‐related cell-signaling
cell‐signaling is is concerned,
concerned, bothboth
Erk1/2
Erk1/2 andand Akt
Akt were showed to be hyperphosphorylated
hyperphosphorylated in frog long long bones
bones of of stressed
stressed
samples
samples (forced
(forcedswimming)
swimming)suggesting
suggestingthat among
that among thethe
putative osteocyte
putative osteocyte signal transduc-
signal trans‐
tion mechanisms, Akt signaling is boosted by increased mechanical
duction mechanisms, Akt signaling is boosted by increased mechanical stresses. Moreo‐ stresses. Moreover, the
authors confirmed again that the increase of osteocyte gap junction
ver, the authors confirmed again that the increase of osteocyte gap junction number is number is dependent
on mechanical
dependent loading (Figure
on mechanical 10). (Figure 10).
loading
Figure 10.
Figure 10. TEM
TEMmicrograph
micrographpanel
panelrepresentative
representativeofof
the abundance
the abundance of of
gapgap
junctions (A–E)
junctions with
(A–E) with
respect to
respect to simple
simple contacts
contacts(F)
(F)ininoverloaded
overloadedfrogs. The
frogs. head‐arrow
The in (A)
head-arrow indicates
in (A) the tip
indicates theoftip
a cy‐
of a
toplasmic process protruding in a cell body of an adjacent osteocyte showing an “invaginated fin‐
cytoplasmic process protruding in a cell body of an adjacent osteocyte showing an “invaginated
ger‐like” junction. Samples in (B) and (F) are decalcified.
finger-like” junction. Samples in (B) and (F) are decalcified.
5. Bone
5. Bone Remodeling
Remodeling and and Sclerostin
Sclerostin Involvement
Involvement
The bone
The bone remodeling
remodeling process
process is characterized
characterized by distinct phases [65,66] (Figure 11).
Duringunloading,
During unloading,or orwhen
whensensitivity
sensitivitytoto strain
strain is altered
is altered by by hormones
hormones (such
(such as parathy‐
as parathyroid
roid hormone,
hormone, estrogens,
estrogens, etc.), etc.), osteocytes
osteocytes stop stop producing
producing the ionic
the ionic currents
currents maintaining
maintaining the
the resting
resting phase,
phase, thusthus allowing
allowing thethe bone
bone lining
lining cells,
cells, the
the stromalcells
stromal cellsand,
and,above
aboveall,
all,the
the
osteocytes (the
osteocytes (the most
most sensitive
sensitive ones
ones to
to loading
loading changes)
changes) to to produce
produce RANKL,
RANKL, that
that activates
activates
the osteoclastogenesis
the osteoclastogenesis (first
(first phase
phase resorption).
resorption). During
During bone
bone resorption,
resorption, BBCS
BBCS is destroyed,
but it is
but is likely
likelythat
thatsurviving
survivingoverstrained
overstrained osteocytes
osteocytes areare
involved in crucial
involved signaling
in crucial that
signaling
that
stopsstops osteoclast
osteoclast activity
activity and triggers
and triggers the reversal
the reversal phase (second
phase (second phase reversion).
phase reversion). Various
authors believe that the cells of the reversal phase could be involved in sending or receiv‐
ing these signals [67–69]. Other signaling pathways may include matrix‐derived factors
such as bone morphogenic protein (BMP)‐2, transforming growth factor (TGF)‐β, and in‐
sulin‐like growth factor (IGF) [66,70,71]. The cells of the reversal phase (probably of stro‐J. Funct. Morphol. Kinesiol. 2021, 6, 28 11 of 17
Various authors believe that the cells of the reversal phase could be involved in sending
or receiving these signals [67–69]. Other signaling pathways may include matrix-derived
factors such as bone morphogenic protein (BMP)-2, transforming growth factor (TGF)-β,
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR
andPEER REVIEW growth factor (IGF) [66,70,71]. The cells of the reversal phase (probably
insulin-like 11 ofof
18
stromal-fibroblast origin) differentiate into osteoblasts, thus bone formation occurs (third
phase deposition). During bone deposition, BBCS is progressively renewed and osteoblast
activity
activitystops
stopswhen
whenlocal
localphysiological
physiologicalloading
loadingconditions
conditionsarearerestored;
restored;the
theosteocytes
osteocytesin in
the
thenewly-laid-down
newly‐laid‐downbone bonematrix
matrixproduce
produceagain
againthe
thesteady
steadyionic
ioniccurrents
currentsthat
thatreturn
returnthe
the
bone
bonetotothe
theresting
restingphase,
phase,therefore
thereforehalting
haltingosteoblast
osteoblastactivity.
activity.
Figure 11. Schematic drawing of postulated BBCS role during bone remodeling cycle. See text for
Figure 11. Schematic drawing of postulated BBCS role during bone remodeling cycle. See text
explanation.
for explanation.
Asfar
As farasasthe
theregulation
regulationof ofbone
boneremodeling
remodelingprocess
processisisconcerned,
concerned,itithas
hasrecently
recentlybeen
been
shown that sclerostin has a relevant role, displaying both autocrine and paracrine
shown that sclerostin has a relevant role, displaying both autocrine and paracrine effects [66]. effects
[66]. Sclerostin
Sclerostin (a 22‐kDa
(a 22-kDa glycoprotein),
glycoprotein), due todue
SOST to promoter
SOST promoter hypomethylation
hypomethylation [72,73], [72,73],
is cur-
is currently considered the major mediator of the molecular osteocyte
rently considered the major mediator of the molecular osteocyte mechanisms involved mechanisms in‐
volved
in in the of
the process process
adaptiveof adaptive bone responses.
bone responses. In the skeleton,
In the mature mature skeleton,
sclerostinsclerostin
is mainlyis
synthesized by differentiated mature osteocytes, while preosteocytes, bone lining cells,lining
mainly synthesized by differentiated mature osteocytes, while preosteocytes, bone and
cells, and osteoblasts
osteoblasts express very express veryof
low levels low levels ofThe
sclerostin. sclerostin.
central The
role central role of
of sclerostin is sclerostin
performedis
performed
through thethrough
interplaythe interplay
between twobetween
opposing two opposing mechanisms:
mechanisms: (1) unloading‐in‐
(1) unloading-induced-high
duced‐high sclerostin levels, that antagonize canonical Wnt in
sclerostin levels, that antagonize canonical Wnt in osteocytes and osteoblasts osteocytes and
and osteoblasts
promote
and promote non‐canonical
non-canonical Wnt and/or other Wnt and/or other
pathways in pathways
osteocytesinandosteocytes and[74–76];
osteoclasts osteoclasts [74–
(2) me-
76]; (2) mechanical
chanical loading‐induced‐low
loading-induced-low sclerostin
sclerostin levels, that levels,
activatethat activate Wnt‐canonical
Wnt-canonical signaling and sig‐
naling
bone and bone Thus,
formation. formation. Thus,bone
adaptive adaptive bone remodeling,
remodeling, occurring in occurring
differentinbone
different bone
compart-
compartments, is driven by altered sclerostin levels, which regulate the expression of the
other osteocyte‐specific proteins, such as RANKL, its decoy receptor osteoprotegerin
(OPG), and other proteins (Figure 12). Under the regulation of differential RANKL and
OPG production, due to specific conditions, sclerostin creates a dynamic RANKL/OPG
ratio [77–79], leading to either bone resorption or formation. Such opposite up‐ or down‐J. Funct. Morphol. Kinesiol. 2021, 6, 28 12 of 17
ments, is driven by altered sclerostin levels, which regulate the expression of the other
osteocyte-specific proteins, such as RANKL, its decoy receptor osteoprotegerin (OPG), and
other proteins (Figure 12). Under the regulation of differential RANKL and OPG produc-
tion, due to specific conditions, sclerostin creates a dynamic RANKL/OPG ratio [77–79],
J. Funct. Morphol. Kinesiol. 2021, 6, x FOR PEER REVIEW 12 of 18
leading to either bone resorption or formation. Such opposite up- or down-regulation of
the remodeling phases allows osteocytes (i.e., the permanent bone cells) to function as
“the orchestrators” of osteoclasts and osteoblasts (i.e., the transient operating bone cells)
ensuring theensuring
bone cells) transition
thefrom bone resorption
transition from bone toresorption
bone formation.
to boneInformation.
this context,
Inthe
thisputative
context,
inhibition of sclerostin
the putative inhibitionrepresents a strategy
of sclerostin to target
represents bone remodeling
a strategy to target boneunbalance in skeletal
remodeling unbal‐
disorders, as osteoporosis,
ance in skeletal disorders, asrheumatoid arthritis,
osteoporosis, bone related-genetic
rheumatoid arthritis, bonedisorders, notwith-
related‐genetic dis‐
standing further studies are needed to better clarify how sclerostin-Ab modulates
orders, notwithstanding further studies are needed to better clarify how sclerostin‐Ab bone
resorption
modulatessincebonesome authors reported
resorption since somethe lack of modulation
authors reported theof RANKL,
lack ofOPG, and other
modulation of
regulators
RANKL, OPG, of osteoclastogenesis during
and other regulators of sclerostin-Ab treatment
osteoclastogenesis during[80–82]. Recently,treatment
sclerostin‐Ab various
papers
[80–82].were published
Recently, on the
various topicwere
papers wherepublished
new therapeutic strategies
on the topic wherewere
newproposed, as
therapeutic
denosumab, rosomozumab,
strategies were proposed, asetc. [83–86]. rosomozumab, etc. [83–86].
denosumab,
Figure 12. Scheme of RANK/RANKL/OPG system. Under specific conditions, differential RANKL
Figure 12. Scheme of RANK/RANKL/OPG system. Under specific conditions, differential RANKL
and OPG production are induced, and the dynamic RANKL/OPG ratio leads to either bone for‐
and OPG production are induced, and the dynamic RANKL/OPG ratio leads to either bone formation
mation or bone resorption.
or bone resorption.
6.6. Interplay
Interplaybetween
betweenMineral
Mineraland
andSkeletal
SkeletalHomeostasis
Homeostasisand
andOsteocyte
OsteocyteRole
RoleMediated
Mediated
bySclerostin
by Sclerostin
Boneremodeling
Bone remodelingisisthe themain
main tool
tool byby which
which thethe skeleton
skeleton answers
answers to both
to both the meta‐
the metabolic
bolicmechanical
and and mechanical demands,
demands, thus thus regulating
regulating the the mineral
mineral andand skeletal
skeletal homeostasis.The
homeostasis. The
mineralhomeostasis
mineral homeostasis keeps
keeps in inrelatively
relativelystable
stablebalance
balancethe theconcentration
concentrationof ofmineral
mineralions,
ions,
as
as calcium
calcium and phosphate,
phosphate, in inthe
theorganic
organicliquids;
liquids;the the skeletal
skeletal homeostasis
homeostasis allows
allows thethe
ad‐
adjustment of shape, mass and bone structure
justment of shape, mass and bone structure following following the actual mechanical needs of
mechanical needs of the the
skeletal
skeletalsegments.
segments. The
The two
two processes
processes are arenot
notindependent
independentof ofeach
eachother;
other;in infact,
fact,various
various
investigations
investigationsshowed
showedthatthatthey
theyarearefunctionally
functionallycorrelated
correlatedin indriving
drivingthe
thebone
bone responses,
responses,
as
asaawhole,
whole,totodifferent
differentexperimental
experimentalconditions
conditions[87–91].
[87–91].
Here,
Here,the
theauthors
authorswant
wantto tostress
stressthe
theimportance
importanceof ofthe
theinterplay
interplaybetween
betweenmineral
mineralandand
skeletal homeostasis (i.e., a dynamic balance of the two homeostases) in
skeletal homeostasis (i.e., a dynamic balance of the two homeostases) in modulating and modulating and
guiding bone’s response to dietary/metabolic alterations and/or unbalance of loading con‐
ditions. In particular, it is important to be emphasized that mineral homeostasis involves
bone response to a variation of mineral serum levels (in consequence of various condi‐
tions, such as hormonal alterations or dietary regimen), while skeletal homeostasis im‐
plies bone answers to loading modifications.You can also read

















































