ANGUSTIFOLIA Regulates Actin Filament Alignment for Nuclear Positioning in Leaves1 OPEN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
ANGUSTIFOLIA Regulates Actin Filament Alignment for
Nuclear Positioning in Leaves1[OPEN]
Kosei Iwabuchi,a,b Haruna Ohnishi,a Kentaro Tamura,a,c Yoichiro Fukao,d Tomoyuki Furuya,e Koro Hattori,e
Hirokazu Tsukaya,e,f and Ikuko Hara-Nishimura a,b,2,3
a
Graduate School of Science, Kyoto University, Kyoto 606-8502, Japan
b
Faculty of Science and Engineering, Konan University, Kobe 658-8501, Japan
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
c
School of Food and Nutritional Sciences, University of Shizuoka, Shizuoka 422-8526, Japan
d
College of Life Sciences, Ritsumeikan University, Shiga 525-8577, Japan
e
Graduate School of Science, University of Tokyo, Tokyo 113-0033, Japan
f
Okazaki Institute for Integrative Bioscience, Okazaki 444-8787, Japan
ORCID IDs: 0000-0002-3696-6806 (K.I.); 0000-0001-7387-2211 (K.T.); 0000-0002-0419-7520 (T.F.); 0000-0002-4430-4538 (H.T.);
0000-0001-8814-1593 (I.H.).
During dark adaptation, plant nuclei move centripetally toward the midplane of the leaf blade; thus, the nuclei on both the
adaxial and abaxial sides become positioned at the inner periclinal walls of cells. This centripetal nuclear positioning implies that
a characteristic cell polarity exists within a leaf, but little is known about the mechanism underlying this process. Here, we show
that ANGUSTIFOLIA (AN) and ACTIN7 regulate centripetal nuclear positioning in Arabidopsis (Arabidopsis thaliana) leaves.
Two mutants defective in the positioning of nuclei in the dark were isolated and designated as unusual nuclear positioning1 (unp1)
and unp2. In the dark, nuclei of unp1 were positioned at the anticlinal walls of adaxial and abaxial mesophyll cells and abaxial
pavement cells, whereas the nuclei of unp2 were positioned at the anticlinal walls of mesophyll and pavement cells on both the
adaxial and abaxial sides. unp1 was caused by a dominant-negative mutation in ACTIN7, and unp2 resulted from a recessive
mutation in AN. Actin filaments in unp1 were fragmented and reduced in number, which led to pleiotropic defects in nuclear
morphology, cytoplasmic streaming, and plant growth. The mutation in AN caused aberrant positioning of nuclei-associated
actin filaments at the anticlinal walls. AN was detected in the cytosol, where it interacted physically with plant-specific dual-
specificity tyrosine phosphorylation-regulated kinases (DYRKPs) and itself. The DYRK inhibitor (1Z)-1-(3-ethyl-5-hydroxy-2
(3H)-benzothiazolylidene)-2-propanone significantly inhibited dark-induced nuclear positioning. Collectively, these results
suggest that the AN-DYRKP complex regulates the alignment of actin filaments during centripetal nuclear positioning in
leaf cells.
The proper spatial arrangement of nuclei is essential Bergeijk et al., 2016). The proper nuclear positioning
for various cellular activities during cell division, also is required to help plants cope with environmental
growth, migration, and differentiation in eukaryotes stimuli, including pathogen infection, touch, tempera-
(Takagi et al., 2011; Gundersen and Worman, 2013; van ture, and light (Takagi et al., 2011; Griffis et al., 2014).
We recently reported that the nuclei of Arabidopsis
1
(Arabidopsis thaliana) leaf cells move to the anticlinal
This work was supported by Grants-in-Aid for Scientific Re- walls of cells in response to strong blue/ultraviolet A
search (22000014 and 15H05776 to I.H-.N., 23-1024 and 17K15145
light to reduce DNA damage and cell death caused by
to K.I., and 26711017 to K.T.) and a Grant-in-Aid for Scientific Research
on Innovative Areas (25113002 to H.T.) from the Japan Society for the
ultraviolet irradiation (Iwabuchi et al., 2016). Further-
Promotion of Science (JSPS). more, we found that nuclei in Arabidopsis leaf cells are
2
Author for contact: ihnishi@gr.bot.kyoto-u.ac.jp. positioned facing toward the body center of the leaf in
3
Senior Author. the dark (Iwabuchi et al., 2016). Specifically, the nuclei
The author responsible for distribution of materials integral to the of pavement and mesophyll cells on both the adaxial
findings presented in this article in accordance with the policy de- and abaxial sides of the leaf are positioned at inner
scribed in the Instructions for Authors (www.plantphysiol.org) is: periclinal walls. These findings indicate that nuclear
Ikuko Hara-Nishimura (ihnishi@gr.bot.kyoto-u.ac.jp). positioning is independent of gravity. This process is
K.I., K.T., and I.H.-N. conceived the research plan and designed induced repeatedly in the dark (Iwabuchi et al., 2007),
the experiments; K.I. and K.T. isolated the mutants; K.I., H.O., and
indicating that this nuclear positioning process is ac-
K.T. performed the molecular biological and cell biological studies;
Y.F. performed the mass spectrometry analysis; T.F., K.H., and H.T.
tive. This finding led to the observation that there is a
performed the imaging analysis of DYRKP2A; K.I., K.T., and I.H.-N. characteristic cell polarity in the adaxial-abaxial direc-
wrote the article. tion of a leaf.
[OPEN]
Articles can be viewed without a subscription. Light-induced nuclear positioning depends on chlo-
www.plantphysiol.org/cgi/doi/10.1104/pp.18.01150 roplast movement. Namely, nuclei cannot move by
Plant PhysiologyÒ, January 2019, Vol. 179, pp. 233–247, www.plantphysiol.org Ó 2019 American Society of Plant Biologists. All Rights Reserved. 233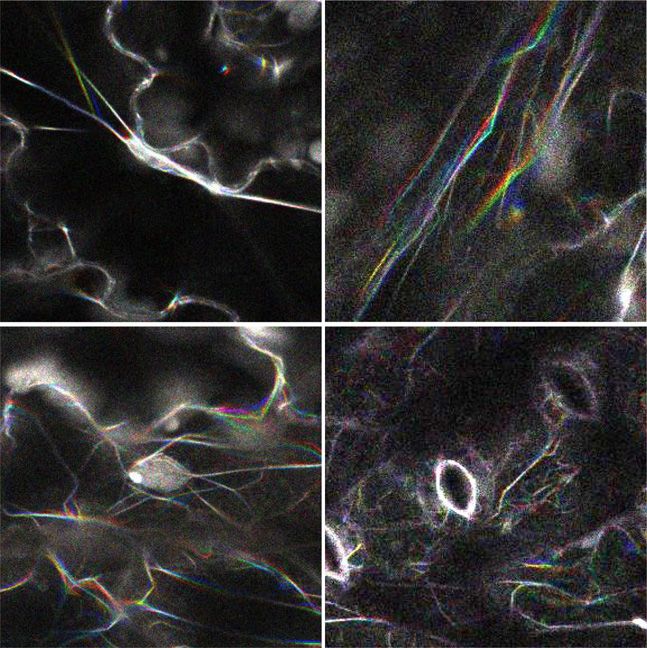
ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
themselves; instead, nucleus-attached chloroplasts Arabidopsis plants expressing the nuclear marker
carry the nuclei to anticlinal walls of cells (Higa et al., Nup50a-GFP (Tamura et al., 2013). In dark-adapted
2014). Light-induced chloroplast movement is a well- wild-type leaves, most nuclei in palisade mesophyll
known process that occurs in a wide variety of plants and pavement cells were positioned at the inner peri-
whose underlying mechanism has been investigated in clinal wall of the cell. In unp1 leaves, by contrast, 52% of
depth (Wada, 2013, 2016; Suetsugu et al., 2017). Chlo- nuclei were aberrantly positioned at the anticlinal walls
roplast movement is regulated by the blue light- of mesophyll cells, although most nuclei in pavement
receptor phototropins, actin filaments, and several cells were positioned at the inner periclinal walls, as in
regulatory proteins (Kong and Wada, 2016). Hence, in wild-type cells (Fig. 1). Leaf nuclei are lens shaped;
mutants lacking these proteins, this type of nuclear thus, the projection area of the nucleus correlates neg-
positioning does not occur. atively with the rate of nuclear positioning at the anti-
Unlike light-induced nuclear positioning, the mech- clinal wall (Iwabuchi et al., 2016). This was observed in
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
anism underlying dark-induced nuclear positioning is unp1 mesophyll cells (Supplemental Fig. S1).
not fully understood. The actin cytoskeleton is essential Nuclear positioning after exposure to 100 mmol m22 s21
for dark-induced nuclear positioning. The plant- blue light for 3 h also was investigated in unp1. In pave-
specific motor myosin XI-i, which is localized to the ment cells, 55% of wild-type nuclei and 35% of unp1 nuclei
nuclear envelope, is involved in dark-induced nuclear moved to the anticlinal walls, although in mesophyll cells,
positioning (Iwabuchi et al., 2010, 2016; Tamura et al., 87% of wild-type nuclei and 83% of unp1 nuclei moved to
2013). Some proteins that regulate chloroplast move- the anticlinal walls (Fig. 1B; Supplemental Fig. S2). These
ment also are involved in this response (Higa et al., results indicate that the unp1 mutation affected blue light-
2014; Suetsugu et al., 2015, 2017). In dark-adapted induced nuclear positioning in pavement cells. We also
pavement cells, thick, longitudinally aligned actin fila- observed the positions of chloroplasts in unp1 mesophyll
ments are associated with nuclei at inner periclinal cells and found no differences between the wild type and
walls (Iwabuchi et al., 2010; Iwabuchi and Takagi, 2010; unp1; the chloroplasts of both lines were distributed in the
Takagi et al., 2011). These actin filaments are thought to lower half of the cell in the dark and at the anticlinal walls
be required for the positioning of the nucleus to the in blue light (Fig. 1A; Supplemental Fig. S2). Thus, the
inner periclinal walls of the cell. Moreover, an analysis unp1 mutation did not appear to affect chloroplast
of actin8D and phototropin2 mutants indicates that dark- positioning.
induced nuclear positioning is differentially regulated The leaf petioles of the unp1 mutant were bent up-
between pavement cells and mesophyll cells (Iwabuchi ward (Supplemental Fig. S3A), and plant height, seed
et al., 2007, 2010, 2016). number per fruit, and fruit length were reduced sig-
Here, we screened for mutants defective in nuclear nificantly in unp1 compared with wild-type plants
position in the dark to identify additional regulatory (Supplemental Fig. S3, B–D). Furthermore, the nuclei of
proteins involved in dark-induced nuclear positioning. unp1 pavement cells were spherical, while those of
We obtained two independent mutants, which we wild-type cells were spindle shaped (Supplemental Fig.
designated unusual nuclear positioning1 (unp1) and unp2. S3E). The nuclei of unp1 and wild-type cells were al-
Whereas unp1 is a previously unreported dominant- most the same size (Supplemental Fig. S3F). These data
negative mutant of ACTIN7, unp2 is a recessive mu- show that, in addition to changes in nuclear position-
tant of the gene ANGUSTIFOLIA (AN). AN encodes a ing, the unp1 mutant exhibited pleiotropic phenotypes
plant homolog of C-terminal-binding protein/brefeldin in various organs.
A-ADP ribosylated substrate (CtBP/BARS; Folkers We isolated the unp2 mutant from an Arabidopsis
et al., 2002; Kim et al., 2002). AN is involved in deter- population carrying T-DNA insertions. Dark-induced
mining leaf and cell shapes, root formation, microtu- nuclear positioning was impaired in pavement and
bule organization, and abiotic stress responses in mesophyll cells of unp2 leaves; 65% of mesophyll nuclei
Arabidopsis (Tsuge et al., 1996; Folkers et al., 2002; Kim and 36% of pavement nuclei were positioned aberrantly
et al., 2002; Bai et al., 2013; Gachomo et al., 2013; Bhasin at the anticlinal walls (Fig. 1), indicating that the unp2
and Hülskamp, 2017). Our findings reveal the rela- mutation affected a gene required for dark-induced nu-
tionship between AN and the actin cytoskeleton in clear positioning. After irradiation with 100 mmol m22 s21
centripetal nuclear positioning in Arabidopsis leaves. blue light for 3 h, 89% of mesophyll nuclei and 69% of
pavement nuclei had moved to the anticlinal walls
(Fig. 1B; Supplemental Fig. S2), suggesting that the unp2
mutation did not affect blue light-induced nuclear posi-
RESULTS tioning. The positions of chloroplasts in unp2 under dark
Isolation of Two Arabidopsis Mutants with Defects in and blue light conditions were similar to those in the wild
Nuclear Positioning in the Dark type (Fig. 1A; Supplemental Fig. S2), suggesting that the
unp2 mutation did not affect chloroplast positioning.
To explore the mechanism of dark-induced nuclear We next investigated dark-induced nuclear position-
positioning, we employed a forward genetics approach. ing in spongy mesophyll cells and abaxial pavement
We isolated the unp1 mutant by screening an ethyl cells. Dark-induced nuclear positioning was impaired
methanesulfonate-mutagenized population of transgenic in these cells in unp1 and unp2 leaves: 64% of spongy
234 Plant Physiol. Vol. 179, 2019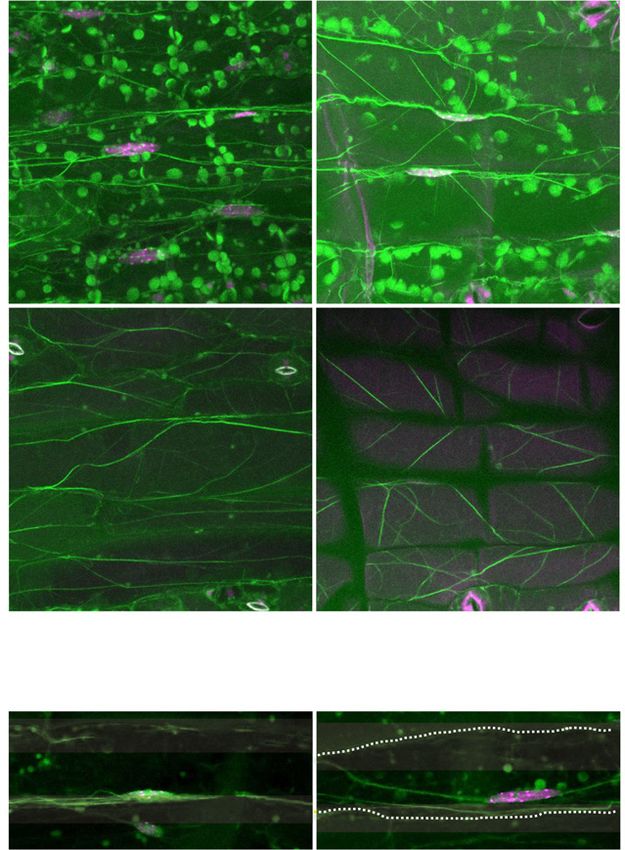
Iwabuchi et al.
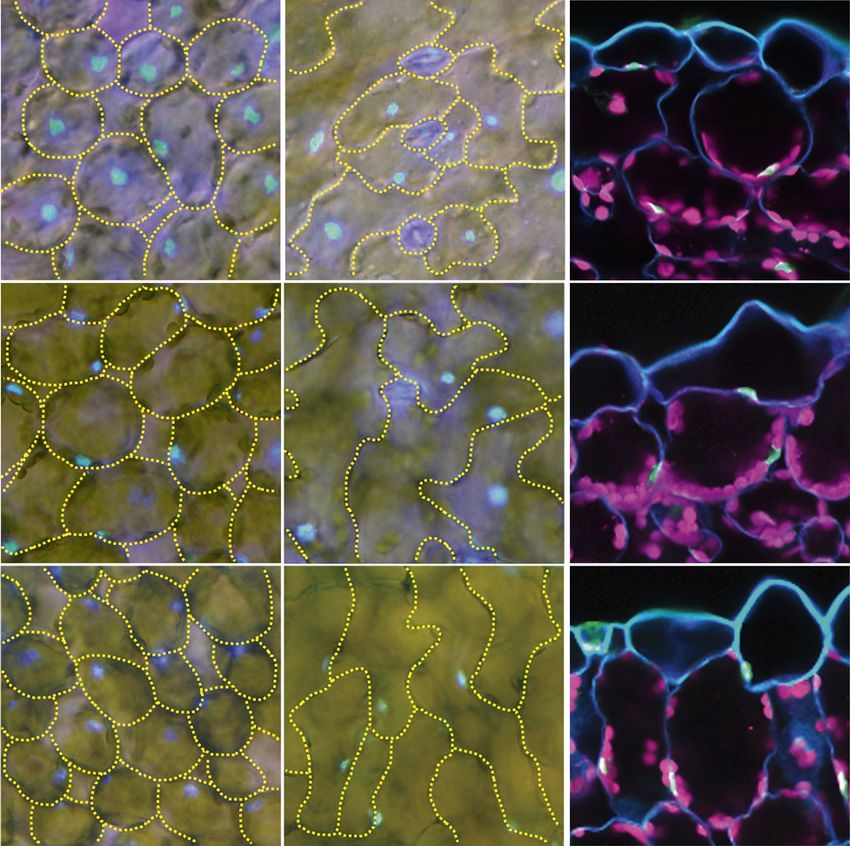
Figure 1. The unp1 and unp2
mutants exhibit aberrant nuclear
positioning in the dark. A, Distri-
bution patterns of nuclei in palisade
mesophyll cells and adaxial pave-
ment cells of wild-type, unp1, and
unp2 leaves in the dark. The left and
middle columns show horizontal
sections with nuclei (blue) stained
with Hoechst 33342. Cells are
outlined with yellow dotted lines.
The right column shows cross sec-
tions, including nuclei (green)
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
stained with Hoechst 33342, cell
walls (blue) stained with Calcofluor,
and chloroplasts (magenta). Bars =
20 mm. B, Percentage of nuclei
positioned on the anticlinal walls of
palisade mesophyll and adaxial
pavement cells of wild-type, unp1,
and unp2 leaves in the dark and
after illumination with blue light
(100 mmol m22 s21 for 3 h). Data
represent means 6 SE (n = 5 leaves;
**, P , 0.01 with Student’s t test).
Mesophyll and pavement cells
were observed in each of five leaves
from different plants; the mean
numbers of each cell type observed
per leaf were as follows: wild-type
leaves, 100 mesophyll and 67
pavement cells; unp1 leaves, 103
mesophyll and 49 pavement cells;
and unp2 leaves, 135 mesophyll
and 88 pavement cells.
mesophyll nuclei and 28% of abaxial pavement nuclei nuclei were positioned at the anticlinal walls in unp2
were positioned to anticlinal walls in unp1, while 40% of (Fig. 2). Thus, the unp1 and unp2 mutations were in-
spongy mesophyll nuclei and 20% of abaxial pavement volved in centripetal nuclear positioning in leaves.
Plant Physiol. Vol. 179, 2019 235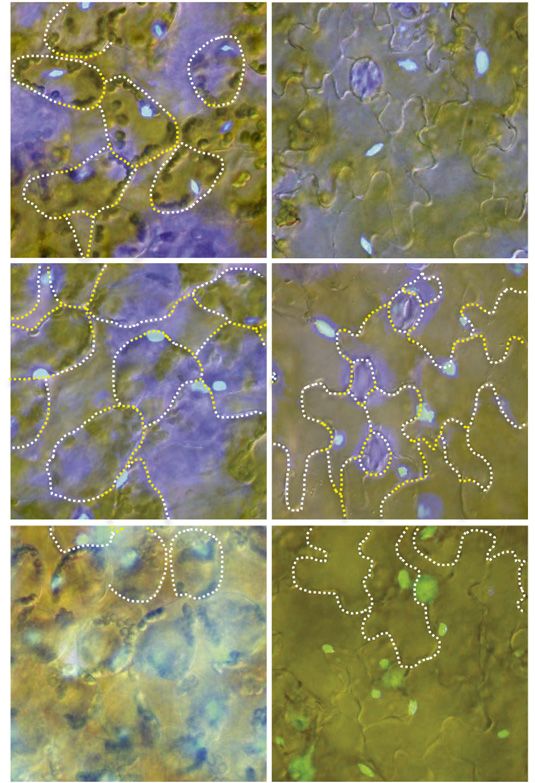
ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
ACTIN7 and AN Are Responsible for the unp1 and unp2
Phenotypes, Respectively
Backcrossing of unp1 with wild-type plants showed
that unp1 was a dominant-negative mutant. Map-based
cloning revealed a point mutation in the second exon of
ACTIN7 (At5g09810) in the unp1 mutant (Fig. 3A),
which resulted in the substitution of Gly-38 with Ser in
a region containing the DNase I-binding loop (D-loop)
of actin protein subdomain 2 (Fig. 3B). The Gly residue
of subdomain 2 is widely conserved in fungi, animals,
and plants (Supplemental Fig. S4). To confirm that this
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
mutation was responsible for the unp1 phenotype, we
introduced ACTIN7 harboring the unp1 mutation,
designated ACTIN7(G38S), into wild-type plants.
When these transgenic plants were placed in the dark,
94% of mesophyll nuclei were positioned at the anti-
clinal walls of cells (Fig. 3C), indicating that the unp1
phenotype resulted from the mutation in ACTIN7.
We also examined the effect of the unp1-type mutation
on other actin isoforms, including ACTIN2 (At3g18780)
and ACTIN8 (At1g49240); like ACTIN7, these genes are
strongly expressed in vegetative tissues of Arabidopsis
(Meagher et al., 1999). When either ACTIN2(G38S) or
ACTIN8(G38S) was expressed under the control of the
ACTIN7 promoter, the mesophyll nuclei of the trans-
genic plants were positioned at the anticlinal walls of
cells in the dark [Fig. 3C; 62% 6 6.6% for ACTIN2(G38S)
and 79% 6 4.6% for ACTIN8(G38S)]. This suggested that
all three actin isoforms were involved in dark-induced
nuclear positioning in mesophyll cells.
Genotyping revealed a T-DNA insertion in the first
intron of AN (At1g01510) in unp2 mutants (Fig. 3D).
Reverse transcription (RT)-PCR analysis did not detect
intact AN mRNA in unp2 (Fig. 3E). Another allele, an-1,
exhibited similar changes in nuclear positioning to
unp2: the percentages of nuclei positioned at the anti-
clinal walls were 73% 6 3.5% for mesophyll cells and
38% 6 6.4% for pavement cells (Fig. 3F). This indicated
that the unp2 phenotype resulted from the deficiency in
AN caused by the T-DNA insertion.
Pavement cells of wild-type leaves had jigsaw piece-
like, irregularly shaped anticlinal walls that interlocked
Figure 2. Dark-induced nuclear positioning on the abaxial side of
with those of neighboring cells. The pavement cells of wild-type, unp1, and unp2 leaves. A, Distribution patterns of nuclei in
unp2 leaves were smoother than those of wild-type cells, spongy mesophyll cells and abaxial pavement cells of wild-type, unp1,
as determined by the circularity index (Supplemental Fig. and unp2 leaves in the dark. Nuclei (blue) were stained with Hoechst
S5, A and B). We compared the circularity index of 33342. Cells are outlined with yellow dotted lines. Bars = 20 mm. B,
pavement cells having nuclei positioned on the anticlinal Percentage of nuclei positioned on the anticlinal walls of spongy me-
walls with that of cells having nuclei positioned on the sophyll and abaxial pavement cells of wild-type, unp1, and unp2 leaves
inner periclinal walls. There were no differences in cir- in the dark. Data represent means 6 SE (n = 5 leaves; *, P , 0.05 and **,
cularity indices between either wild-type or unp2 cells P , 0.01 with Student’s t test). Mesophyll and pavement cells were
(Supplemental Fig. S5C). Therefore, AN-dependent dark- observed in each of five leaves from different plants; the mean numbers
of each cell type observed per leaf were as follows: wild-type leaves,
induced nuclear positioning was not related to cell shape.
45 spongy mesophyll and 44 abaxial pavement cells; unp1 leaves,
30 spongy mesophyll and 45 abaxial pavement cells; and unp2 leaves, 43
spongy mesophyll and 57 abaxial pavement cells.
The unp1 and unp2 Mutations Affect Actin
Filament Organization
fluorescently labeled phalloidin staining and immuno-
We previously demonstrated the actin-dependent fluorescence staining (Iwabuchi et al., 2010). We exam-
regulation of dark-induced nuclear positioning by ined how the unp1 mutation affected actin organization
236 Plant Physiol. Vol. 179, 2019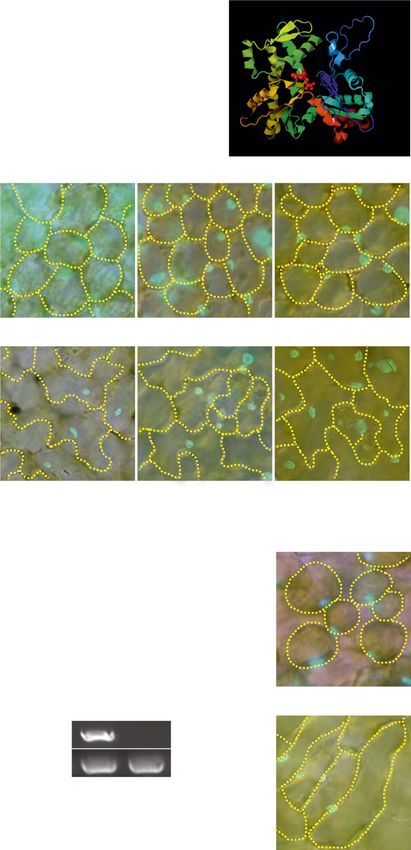
Iwabuchi et al.
by fluorescently labeled phalloidin staining. In the
pavement cells of wild-type leaves, nuclei were associ-
ated with thick, longitudinally aligned actin filaments at
inner periclinal walls in the dark (Fig. 4A). Actin fila-
ments also were well organized at the upper cell surface
(Fig. 4A). The number of actin filaments was reduced
drastically in unp1 pavement cells (Fig. 4, A and B), even
though the levels of actin protein (Fig. 4C) and actin
bundling (Fig. 4D) were comparable between the wild
type and unp1.
To investigate actin organization in living cells, we ex-
amined transgenic plants expressing Lifeact-Venus (Era
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
et al., 2009). The actin filaments were severely frag-
mented in the pavement cells of transgenic plants
(Fig. 5A), although their lateral movements were normal
in unp1 (Fig. 6; Supplemental Movies S1 and S2). The unp1
mutation also affected cytoplasmic streaming: unp1
cells showed a 36% decrease in the velocity of cytoplas-
mic streaming compared with the wild type (Fig. 5B;
Supplemental Movie S3). These results suggested that Gly-
38 in the D-loop of ACTIN7 was required for the polym-
erization of actin filaments and cytoplasmic streaming.
We also examined the effect of the unp2 mutation on
the actin cytoskeleton in dark-adapted pavement cells
using phalloidin staining but not Lifeact-Venus, which
caused artifactual defects in actin organization (for an
explanation of our choice of protocol, see “Materials
and Methods”). We observed longitudinally aligned
actin filaments associated with the nuclei in the wild
type and unp2 (Fig. 4A). However, the actin filaments in
the wild type were aligned along inner periclinal walls,
while those in unp2 were aligned aberrantly along the
anticlinal walls (Fig. 4A). The number of actin filaments
was slightly but significantly reduced at the cell surface
in unp2 cells compared with wild-type cells (Fig. 4B). By
contrast, the levels of actin protein (Fig. 4C), actin
bundling (Fig. 4D), and the velocity of cytoplasmic
streaming (Fig. 5B) were normal in the mutant. In ad-
dition, to eliminate the possibility that the aberrant cell
shape affected actin filament alignment in unp2, we
examined pavement cells positioned just above the leaf
midvein. The shapes of these pavement cells in wild-
Figure 3. ACTIN7 and AN are required for nuclear positioning in the type and unp2 plants were similar: the cells were long,
dark. A, Schematic representation of ACTIN7 (At5g09810) showing the narrow, and less wavy than pavement cells in other
position of the unp1 mutation. Gray boxes, Untranslated regions; black areas of the leaf (Fig. 7A). Even in such pavement cells
boxes, exons; solid lines, introns. B, Front view of a model of actin of unp2, nucleus-associated actin filaments were
protein (Protein Data Bank code, 3MFP; Fujii et al., 2010). C, Adaxial aligned aberrantly along the anticlinal walls (Fig. 7A).
pavement and palisade mesophyll cells of plants expressing ACTIN7, As shown in Figure 1B, some pavement nuclei were
ACTIN2, or ACTIN8 containing unp1-type mutations in the dark. Blue, positioned at the anticlinal and inner periclinal walls in
Nuclei stained with Hoechst 33342. Cells are outlined with yellow
wild-type and unp2 cells, respectively, despite dark
dotted lines. Values below the images indicate the percentages of nuclei
positioned on the anticlinal walls. Data represent means 6 SE (n = 5
adaptation. At such times, actin filaments were aligned
leaves). Mesophyll and pavement cells were observed in each of five with the nuclei along the anticlinal walls in the wild-type
leaves from different plants; the mean numbers of each cell type ob-
served per leaf were as follows: ACTIN2(G38S), 102 mesophyll and 43
pavement cells; ACTIN7(G38S), 101 mesophyll and 40 pavement mesophyll and adaxial pavement cells of an-1 plants in the dark. Blue,
cells; and ACTIN8(G38S), 92 mesophyll and 35 pavement cells. Nuclei stained with Hoechst 33342. Cells are outlined with yellow
Bars = 20 mm. D, Schematic representation of AN (At1g01510) showing dotted lines. Values below the images indicate the percentages of nuclei
the positions of the unp2 and an-1 mutations. Gray boxes, Untranslated positioned on anticlinal walls. Data represent means 6 SE (n = 5 leaves).
regions; black boxes, exons; solid lines, introns. E, RT-PCR of AN and Mean numbers of 101 mesophyll cells and 50 pavement cells were
ACTIN2 (control) transcripts in wild-type and unp2 leaves. F, Palisade observed in each of five leaves from different an-1 plants. Bars = 20 mm.
Plant Physiol. Vol. 179, 2019 237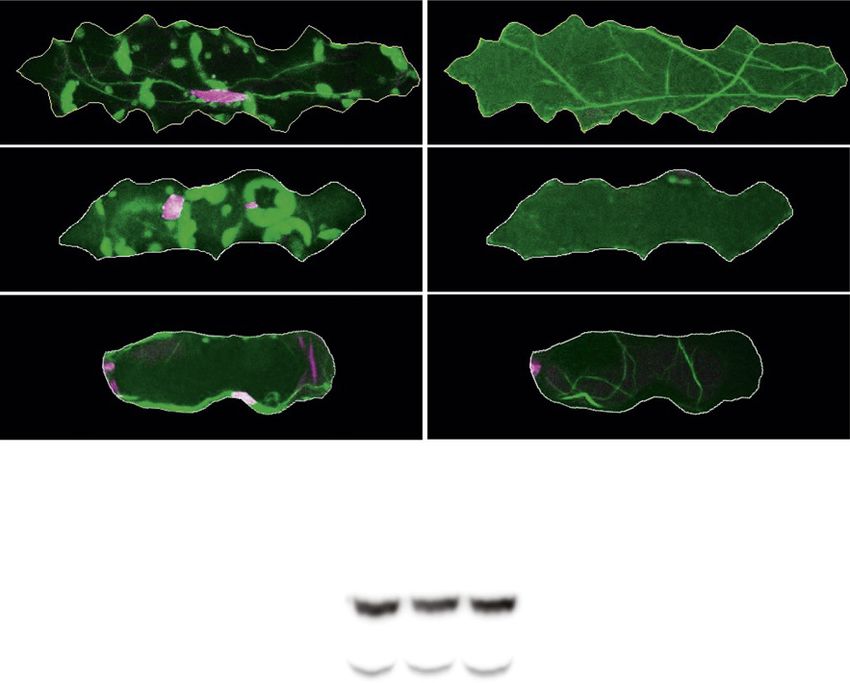
ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
Figure 4. Organization of actin filaments in pavement cells of wild-type, unp1, and unp2 leaves in the dark. A, Maximum in-
tensity projections of actin filaments in the vicinity of the nuclei (left) and at the surface (right) of adaxial pavement cells of wild-
type, unp1, and unp2 leaves in the dark. Magenta (arrows), Nuclei stained with Hoechst 33342; green (arrowheads), actin
filaments stained with phalloidin. Cells are outlined with sold white lines. Sequential images were taken from the top to bottom of
cells along the optical z axis at 0.5-mm intervals. Round, green structures approximately 5 mm in diameter (indicated with black
asterisks) are chloroplasts in the underlying mesophyll cells. Bars = 20 mm. B, Occupancy rates of actin filaments at the cell
surfaces of adaxial pavement cells in wild-type, unp1, and unp2 leaves (n = 34 cells in wild-type leaves, 33 cells in unp1 leaves,
and 48 cells in unp2 leaves; *, P , 0.05 and **, P , 0.01 with Student’s t test). Data represent means 6 SE. C, Immunoblot analysis
of wild-type, unp1, and unp2 leaf proteins using anti-actin and anti-tubulin antibodies. Tubulin was used as a loading control. D,
Fluorescence levels of actin filaments at the cell surface of adaxial pavement cells in wild-type, unp1, and unp2 leaves (n = 34
cells in wild-type leaves, 33 cells in unp1 leaves, and 48 cells in unp2 leaves). Data represent means 6 SE.
cells and on the inner periclinal walls in unp2 cells (Fig. 7B), rather than in the nucleus in both pavement and me-
producing a strong relationship between the positions of sophyll cells (Supplemental Fig. S6). The cytosolic lo-
nuclei and the alignment of actin filaments. Taken to- calization of AN-GFP was not affected by irradiation
gether, these results indicated that AN was required for the with 100 mmol m22 s21 blue light for 1 h, which nor-
proper alignment of nucleus-associated actin filaments. mally causes nuclei to begin to move to the anticlinal
walls of cells (Supplemental Fig. S6). These results in-
dicated that AN was constitutively present in the cy-
AN Constitutively Localizes to the Cytosol under Dark and tosol under both dark and blue light conditions.
Blue Light Conditions
We also investigated the intracellular localization of AN Associates with Dual-Specificity Tyr
AN in pavement and mesophyll cells of transgenic Phosphorylation-Regulated Kinases
plants expressing both AN-GFP and the nuclear mem-
brane marker SUN2-tRFP. Under dark conditions, To identify proteins that interacted with AN, we
AN-GFP was detected predominantly in the cytosol performed an interactome analysis of transgenic plants
238 Plant Physiol. Vol. 179, 2019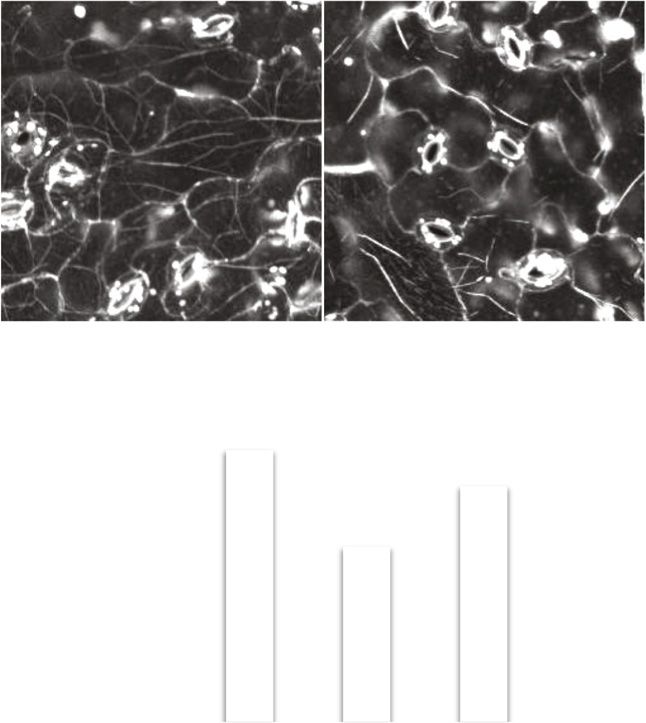
Iwabuchi et al.
(3H)-benzothiazolylidene)-2-propanone], a benzothiazole
compound that inhibits animal Dyrk1A (Ogawa et al.,
2010), on dark-induced nuclear positioning. INDY binds
to the ATP pocket of Dyrk1A and is a highly selective
inhibitor of its kinase activity (Ogawa et al., 2010). After
exposure to 100 mmol m22 s21 blue light for 3 h, leaves
were treated with INDY at concentrations of 10 mM,
100 mM, or 1 mM for 5 h in the dark. Concentrations of
100 mM and 1 mM INDY significantly inhibited the
movement of nuclei from the anticlinal walls to the inner
periclinal walls in both pavement cells and mesophyll
cells (Fig. 9, A and B). By contrast, 100 mM INDY did not
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
inhibit blue light-induced nuclear positioning (Fig. 9, C
and D). Therefore, DYRKPs appear to be involved in
dark-induced nuclear positioning but not in blue light-
induced nuclear positioning.
We next focused on the phosphorylation of actin
protein. To examine the level of actin phosphorylation,
we subjected wild-type leaf extracts to immunoblotting
on a SuperSep Phos-tag gel with anti-actin antibody.
Pretreatment of the leaf extract with alkaline phos-
phatase affected the migration of actin protein (Fig. 10).
Unphosphorylated actin protein was not detected in the
absence of alkaline phosphatase treatment (Fig. 10),
clearly showing that most actin proteins were phos-
phorylated in planta. The phosphorylation status of
actin proteins did not differ between unp2 and wild-
Figure 5. Fragmentation of actin filaments affects the velocities of type plants (Fig. 10), suggesting that the AN-DYRKP
particles in pavement cells of unp1 leaves. A, Maximum intensity pro- complex was not involved directly in the phosphoryl-
jections of actin filaments at the surfaces of adaxial pavement cells in ation of actin.
Lifeact-Venus/wild-type and Lifeact-Venus/unp1 leaves. Cells are out-
lined with dotted yellow lines. Sequential images were taken from the
top to bottom of cells along the optical z axis at 0.5-mm intervals.
Bars = 20 mm. B, Maximal velocities of particles in adaxial pavement DISCUSSION
cells in wild-type, unp1, and unp2 leaves. Particles in pavement cells
located above the midveins of the leaves were imaged at 0.13-s intervals A Model for Centripetal Nuclear Positioning in Plants
for 30 s (see Supplemental Movie S3). The distances traveled by the
particles in 5 s were determined. Data represent means 6 SE (n = 21 cells
Our results indicated that centripetal nuclear posi-
in wild-type leaves, 17 cells in unp1 leaves, and 23 cells in unp2 leaves) tioning required the presence of actin filaments aligned
from three to four leaves (**, P , 0.01 with Student’s t test). at the inner periclinal walls in an AN-dependent man-
ner. We propose a model for dark-induced nuclear
positioning in inner periclinal walls in palisade meso-
expressing AN-GFP. Mass spectrometry analysis of the phyll and pavement cells of plants (Fig. 11). In this
fraction pulled down by the anti-GFP antibody identi- model, the AN-DYRKP complex regulates the phos-
fied several forms of plant-specific dual-specificity phorylation of an unknown protein X in the cytosol.
tyrosine phosphorylation-regulated kinases (DYRKPs; Protein X regulates the alignment of actin filaments at
DYRKP-1, DYRKP-2A, DYRKP-2B, and DYRKP-3; Fig. 8, the inner periclinal walls. The actin filaments are com-
A and B; Supplemental Fig. S7). posed of ACTIN2, ACTIN7, and ACTIN8, all of which
To confirm a physical interaction between AN and are phosphorylated. Further studies should focus on
DYRKPs, we performed a yeast two-hybrid assay. AN clarifying the detailed functions of AN and the DYRKPs
interacted physically with DYRKP-2B but not DYRKP-3 and on identifying component(s) downstream from the
(Fig. 8C). In addition, AN self-assembled in this assay AN-DYRKP complex. We showed previously that, in
(Fig. 8C). We also investigated the intracellular locali- the dark, nuclei move to the centers of the inner peri-
zation of DYRKP-2A in transgenic plants expressing clinal walls via a process involving the myosin XI-i and
GFP-DYRKP-2A. Distribution was similar to that of WITs. These proteins, together with the nuclear mem-
AN, being localized predominantly to the cytosol in brane proteins WIP and SUN and the lamina-like pro-
both pavement and mesophyll cells (Fig. 8D). Thus, the teins CROWDED NUCLEI and KAKU4 (Tamura et al.,
AN-DYRKP complex appears to function in the cytosol. 2013, 2015; Goto et al., 2014), form the LINC complex. It
Several inhibitors have been used to investigate the would be useful to investigate whether centripetal nu-
function of DYRK in animal cells (Pozo et al., 2013). We clear positioning involves a relationship between the
examined the effect of INDY [(1Z)-1-(3-ethyl-5-hydroxy-2 AN-DYRKP and LINC complexes.
Plant Physiol. Vol. 179, 2019 239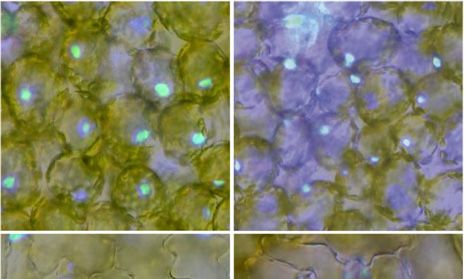
ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
Figure 6. Lateral movements of actin filaments
in leaf pavement cells of Lifeact-Venus/wild-type
and Lifeact-Venus/unp1 plants. A, Movement of
actin filaments. Actin filaments were imaged at 5-s
intervals for 60 s (see Supplemental Movies S1 and
S2). Images at the 0-, 5-, and 10-s time points were
merged. Arrows indicate nuclei. Bars = 20 mm. B,
Cross-correlation coefficients between the first
image and the subsequent time-sequential images
of actin filaments in the vicinity of nuclei (left) and
at the cell surface (right) in Lifeact-Venus/wild-
type and Lifeact-Venus/unp1 plants. Data repre-
sent means 6 SE (n = 4–6 leaves). A mean of four
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
pavement cells were observed in each of four to
six leaves from different plants.
unp1 Is a Dominant-Negative Mutant of ACTIN7 actins) also are incorporated into the filaments with
some frequency. These mutated actins inhibit the fur-
The D-loop is required for the actin-actin interactions ther incorporation of normal actins, as they are unable
in mammal skeletal muscles (Khaitlina and Strzelecka-
to undergo actin-actin interactions; thus, short actin
Gołaszewska, 2002; Oda et al., 2009; Fujii et al., 2010).
filaments are formed. Therefore, short actin filaments in
Additionally, in the slime mold Dictyostelium dis-
coideum, phosphorylation of Tyr-53 in the actin D-loop unp1 may be able to move laterally via myosin (Ueda
disrupts associations between actin monomers, result- et al., 2010).
ing in a decrease in polymerization (Liu et al., 2006; There are two other dominant-negative mutants of
Baek et al., 2008). In unp1, Gly-38 of the D-loop of actin: act2-2D, in which Arg-149 is substituted by Cys in
subdomain 2 of ACTIN7 was substituted with Ser, subdomain 3 of ACTIN2 (Nishimura et al., 2003), and
suggesting that the D-loop has an important role in actin8D/fryzzy1, in which Glu-272 is replaced with Lys
actin polymerization in plants. The D-loop also is im- in the hydrophobic loop between subdomains 3 and 4
portant for actin-myosin interactions in D. discoideum in ACTIN8 (Kato et al., 2010). unp1 exhibited similar
(Kubota et al., 2009). In plants, the actomyosin system pleiotropic phenotypes to these mutants (Supplemental
drives cytoplasmic streaming (Ueda et al., 2010), pos- Fig. S1), although, in contrast to the severe growth de-
sibly due to the actin-myosin interactions through the fects of act2-2D and actin8D, the growth of unp1 plants
actin D-loop. was relatively normal. This dissimilarity might result
As discussed previously (Nishimura et al., 2003; Kato from the positions of the mutations, as there were no
et al., 2010), many normal actins are polymerized into obvious growth differences between ACTIN2(G38S),
actin filaments in unp1, but mutated actins (unp1-type ACTIN7(G38S), and ACTIN8(G38S) plants.
240 Plant Physiol. Vol. 179, 2019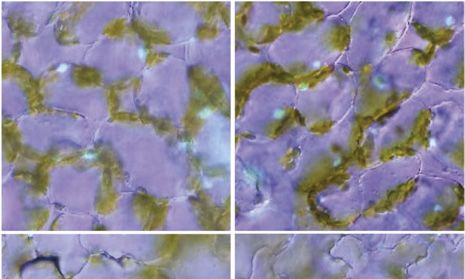
Iwabuchi et al.
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
Figure 7. Organization of actin filaments in pavement cells of wild-type
and unp2 leaves in the dark. Maximum intensity projections are shown
for actin filaments in the vicinity of nuclei and at the surfaces of adaxial
pavement cells of wild-type and unp2 leaves (A) and actin filaments
associated with nuclei occasionally positioned at the anticlinal wall of
wild-type pavement cells and at the inner periclinal wall of unp2
pavement cells (B). Magenta (arrows), Nuclei stained with Hoechst
33342; green (arrowheads), actin filaments stained with phalloidin.
Cells are outlined with dotted yellow lines. Sequential images were
taken from the top to bottom of cells along the optical z axis at 0.5-mm
intervals. Round, green structures approximately 5 mm in diameter
(indicated with black asterisks) are chloroplasts of the underlying me-
sophyll cells. Bars = 20 mm.
Figure 8. AN associates with the protein kinase DYRKP. A, Silver
actin8D, like unp1, exhibits normal nuclear position- staining of products from transgenic plants expressing either free GFP or
ing in pavement cells in the dark (Iwabuchi et al., 2016), AN-GFP pulled down by anti-GFP antibody. AGI, Arabidopsis Genome
which suggests that nuclear position in pavement cells Initiative. B, Identification of putative AN-associating protein kinases by
requires an actin-independent nuclear centering sys- mass spectrometry. C, Yeast two-hybrid assay showing physical inter-
tem. Alternatively, the seemingly normal positioning of actions of AN with DYRKP-2B and AN. D, GFP signals in the adaxial
nuclei in unp1 and actin8D may result from an absence pavement and palisade mesophyll cells of 35S:GFP-DYRKP-2A trans-
genic plants. Bars = 20 mm.
of light-induced nuclear positioning, in which case the
nuclei would always keep a position at the inner peri-
clinal wall. Furthermore, dark-induced nuclear posi-
tioning was affected by the unp1 mutation in abaxial unp1 is a new dominant-negative mutant of ACTIN7
pavement cells but not in adaxial pavement cells, pos- and also is a valuable tool for manipulating actin fila-
sibly due to a difference in the response to the unp1 ments in plants. As actin polymerization can be
mutation between adaxial and abaxial pavement cells. inhibited genetically by the expression of the dominant-
Plant Physiol. Vol. 179, 2019 241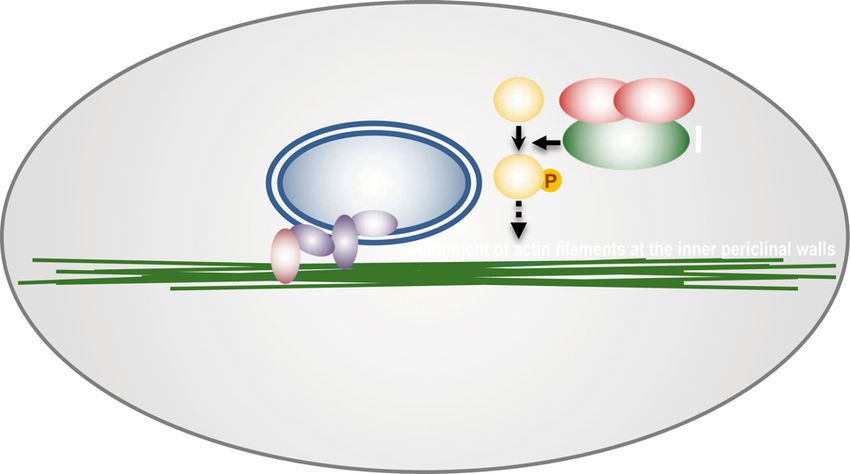
ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
Figure 9. INDY inhibits dark-induced nu-
clear positioning but not blue light-induced
nuclear positioning. A, Distribution patterns
of nuclei in palisade mesophyll and adaxial
pavement cells of leaves treated with either
100 mM INDY or dimethyl sulfoxide
(DMSO; Mock) for 5 h in the dark following
exposure to 100 mmol m22 s21 blue light
for 3 h. Nuclei (blue) were stained with
Hoechst 33342. Cells are outlined with
yellow dotted lines. Bars = 20 mm. B, Effects
of INDY on dark-induced nuclear position-
ing. Data represent means 6 SE (n = 5–6
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
leaves; *, P , 0.05 and **, P , 0.01 with
Student’s t test). Mean numbers of 121
palisade mesophyll cells and 78 adaxial
pavement cells were observed in each of
five to six leaves from different plants. C,
Distribution patterns of nuclei in palisade
mesophyll and adaxial pavement cells of
leaves exposed to 100 mmol m22 s21 blue
light for 3 h in the presence of either 100 mM
INDY or DMSO (Mock). The INDY or
DMSO treatments were performed 1 h be-
fore the end of the dark treatment. Nuclei
(blue) were stained with Hoechst 33342.
Cells are outlined with yellow dotted lines.
Bars = 20 mm. D, Percentage of nuclei po-
sitioned on the anticlinal walls of palisade
mesophyll and adaxial pavement cells in
blue light. Data represent means 6 SE (n = 5
leaves). Mean numbers of 106 mesophyll
cells and 72 pavement cells were observed
in each of five leaves from different plants.
negative ACTIN2, ACTIN7, and ACTIN8 proteins, the gene under a tissue-specific or inducible promoter. This
role of the actin cytoskeleton in cellular processes could tissue specificity cannot be obtained using actin
be investigated in tissues of interest by generating inhibitors.
transgenic plants expressing a dominant-negative actin
AN Regulates the Alignment of Actin Filaments for
Centripetal Nuclear Positioning
AN controls the arrangement of cortical microtubules
in Arabidopsis (Folkers et al., 2002; Kim et al., 2002). On
the other hand, the arrangement of microtubules affects
the alignment of actin filaments and vice versa
(Mineyuki and Palevitz, 1990; Seagull, 1990; Eleftheriou
and Palevitz, 1992; Tominaga et al., 1997; Era et al.,
Figure 10. Representative phosphorylation status of actin in wild-type,
unp1, and unp2 leaves. Top, Immunoblot using a SuperSep Phos-tag gel 2013). Actin filaments in unp2 pavement cells were
showing the shift in the actin band toward a higher mass in samples not aligned aberrantly to anticlinal walls; thus, AN might
treated with alkaline phosphatase (2) compared with those treated with regulate the actin filament alignment directly or indi-
alkaline phosphatase (+). Bottom, Immunoblot using a standard gel as rectly. The shape of unp2 pavement cells did not affect
the loading reference. the nuclear position in the dark; thus, it is also possible
242 Plant Physiol. Vol. 179, 2019Iwabuchi et al.
Figure 11. Model of dark-induced nu-
clear positioning. AN associates with
DYRKPs to regulate the alignment of actin
filaments at the inner periclinal walls
through the phosphorylation of the putative
target, X. Actin filaments are composed of
ACTIN2, ACTIN7, and ACTIN8, and the
actin proteins are highly phosphorylated.
Actin filaments associate with the linker of
nucleoskeleton and cytoskeleton (LINC)
complex, which is composed of myosin XI-
i and the nuclear membrane proteins WPP
domain-interacting tail-anchored proteins
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
(WITs), WPP domain-interacting protein
(WIP), and Sad1/UNC-84 (SUN).
that AN regulates nuclear positioning independently of conserved in animals, plants, fungi, and protists, and is
cell shape formation. involved in developmental processes, cell homeostasis,
Several studies suggest diverse roles for AN. For and cellular stresses (Aranda et al., 2011). DYRK3 is in-
example, deficiency of AN results in various pheno- volved in stress granule formation in HeLa cells (Wippich
types, including narrow leaves (Tsukaya et al., 1994; et al., 2013), and AN is involved in this process in Arabi-
Tsuge et al., 1996), trichomes with reduced branching dopsis leaf cells (Bhasin and Hülskamp, 2017), suggesting
(Cho et al., 2005), twisted roots and seed pods (Bai et al., that DYRK3 and AN function in the same pathway. Given
2013; Kwak et al., 2015), and increased tolerance to that DYRKPs had a high Mascot score in mass spec-
drought and pathogen attack (Gachomo et al., 2013). trometry analysis (Fig. 8), it is possible that AN and
The molecular roles of AN in these phenotypes remain DYRKPs constitutively associate with each other to func-
to be clarified. tion in the cytosol. The DYRK inhibitor, INDY, also
AN encodes a protein related to CtBP/BARS (Folkers inhibited dark-induced nuclear positioning, suggesting
et al., 2002; Kim et al., 2002). CtBP/BARS has an NAD- that DYRKP, together with AN, was involved in regulat-
binding motif, the NAD(H)-dependent D-hydroxyacid ing dark-induced nuclear positioning. The inhibitory effect
dehydrogenase catalytic triad, and PXDLS-binding of INDY on dark-induced nuclear positioning was not a
domains, required for functioning as a transcriptional side effect because blue light-induced nuclear positioning
corepressor (Chinnadurai, 2002, 2007). AN, however, was statistically normal in the presence of INDY. Never-
has mutations in all these regions (Stern et al., 2007), has theless, further analyses using DYRKP-deficient or kinase-
a plant-specific region at the C terminus (Kim et al., dead mutants are required to clarify the molecular mech-
2002; Stern et al., 2007), and functions outside the nu- anisms through which the DYRKPs affect centripetal nu-
cleus (Minamisawa et al., 2011), suggesting that its ac- clear positioning in plants.
tion differs from that of CtBP.
MATERIALS AND METHODS
AN Associates with the Protein Kinases DYRKPs Plant Materials and Growth Conditions
AN-GFP was detected previously not only in the Arabidopsis (Arabidopsis thaliana) accession Columbia was used as the wild
cytosol of Arabidopsis cells but also in the close vicinity type. The unp1, unp2 (SALK_026489), an-1 (Tsukaya et al., 1994), and Lifeact-
of the trans-Golgi, where it formed several small particles Venus (Era et al., 2009) plants were all in the Columbia background. The unp1
plants expressing Lifeact-Venus (Lifeact-Venus/unp1) were generated by
(Folkers et al., 2002; Minamisawa et al., 2011). These small crossing unp1 with transgenic plants expressing Lifeact-Venus. The plants
particles, however, were not observed under our experi- were grown on compost for 2 to 5 weeks at 22°C under a 16-h-white light
mental conditions, even though we used the same trans- (30–50 mmol m22 s21)/8-h-dark cycle. Alternatively, the plants were grown
genic line as a previous study (Minamisawa et al., 2011); aseptically on germination medium plates (one-half-strength Murashige and
thus, we conclude that cytosolic AN is required for dark- Skoog salts, 0.025% [w/v] MES-KOH, pH 5.7, and 0.5% [w/v] Gellan gum)
under continuous white light.
induced nuclear positioning.
We identified DYRKP-2B as an AN-interacting pro-
tein (Fig. 8); its homolog DYRKP-1 also interacts physically Isolation of the unp1 Mutant and Map-Based Cloning
with AN (Bhasin and Hülskamp, 2017). DYRKP-1 and of UNP1
DYRKP-2B are members of a plant-specific subgroup in Seeds of a transgenic plant expressing Nup50a-GFP were mutagenized by
the DYRK family (Kajikawa et al., 2015), which is widely ethyl methanesulfonate treatment as described previously (Tamura et al., 2013).
Plant Physiol. Vol. 179, 2019 243ANGUSTIFOLIA-Dependent Dark Positioning of the Nucleus
M2 seeds were collected from individual M1 plants to generate M2 lines. Each aberrant nuclear positioning in the dark was selected. Nuclei were visualized by
seedling was examined with a fluorescence microscope (Axioskop 2 plus; Hoechst 33342 nuclear staining as described below.
Zeiss), and a mutant line that exhibited aberrant nuclear positioning was se- Total RNA was isolated from leaves with an RNeasy Plant Mini Kit (Qiagen)
lected. Map-based cloning was performed as described previously (Tamura according to the manufacturer’s instructions. Total RNA was subjected to first-strand
et al., 2005). The unp1 mutant expressing Nup50a-GFP was crossed with a cDNA synthesis using Ready-To-Go RT-PCR Beads (GE Healthcare), and the cDNA
wild-type plant to remove Nup50a-GFP for nuclear positioning analysis. was amplified by PCR under the following conditions: 30 cycles of 95°C for 15 s,
60°C for 15 s, and 72°C for 60 s. The primer sets were as follows: 59-GCCTCA
CGTCGTTACACTC-39 (AN forward) and 59-CCTGTTGCCTACTGGTGGAT-39
Generation of Transgenic Plants Expressing ACTIN2 (AN reverse); and 59-AGAGATTCAGATGCCCAGAAGTCTTGTTCC-39 (ACTIN2
(G38S), ACTIN7(G38S), and ACTIN8(G38S) forward) and 59-AACGATTCCTGGACCTGCCTCATCATACTC-39 (ACTIN2
reverse).
The Gateway cloning system (Invitrogen) was used to construct ProACTIN7:
ACTIN7(G38S). The ProACTIN7:ACTIN7(G38S) fragment from unp1 DNA was
amplified by PCR using the following primers: 59-AACCAATTCAGTCGA Dark and Light Treatments
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
CGATATATCACGAAAACCGATC-39 (ACTIN7 forward) and 59-AA
For dark treatment, detached leaves were placed on GM plates and incubated
GCTGGGTCTAGATATCCGTTTCGCATCAGAATGGTAATACAC-39 (ACTIN7
in the dark for at least 16 h. For light treatment, detached leaves were irradiated
reverse). The amplified fragments were subcloned into the pENTR1A entry
with 100 mmol m22 s21 blue light (470 nm) using a light-emitting diode light
vector (Invitrogen) that had been digested with SalI and EcoRV using an In-
source system (IS-mini; CCS). Light intensity was measured using a quantum
Fusion HD Cloning Kit (Takara). The ProACTIN7:ACTIN7(G38S) fragment was
sensor (LI-190SA; LI-COR).
introduced into the pFAST-G01 plant expression vector via the LR recombi-
nation reaction (Shimada et al., 2010).
To construct ProACTIN7:ACTIN2(G38S), the promoter region of ACTIN7 Visualization of Nuclei, Cell Walls, and Chloroplasts
and the genomic regions of ACTIN2 and ACTIN8 were amplified from DNA
from wild-type plants via PCR using the following primer sets: 59-AACCAA Samples were fixed in buffer solution (50 mM PIPES, 10 mM EGTA, and 5 mM
TTCAGTCGACGATATATCACGAAAACCGATC-39 (ACTIN7-promoter for- MgSO4, pH 7) containing 2% (w/v) formaldehyde (Wako) and 0.3% (w/v)
ward) and 59-TTTTCACTAAAAAAAAAG-39 (ACTIN7-promoter reverse); 59- glutaraldehyde (Wako) for 1 h. The fixed samples were stained with 5 mg mL21
TTTTTTTAGTGAAAAATGGCTGAGGCTGATGAT-39 (ACTIN2-genomic Hoechst 33342 (CalBiochem) diluted in buffer solution supplemented with
forward) and 59-TAACAACACTGGGAAAAACA-39 (ACTIN2-G38S reverse); 0.03% (v/v) Triton X-100 for at least 1.5 h.
59-TTTCCCAGTGTTGTTAGTAGGCCAAGACATCATGGT-39 (ACTIN2-G38S
forward) and 59-TAAGACAAGACACACTTAGAAACATTTTCTGTGA-39
(ACTIN2-genomic reverse); and 59-GTGTGTCTTGTCTTATCTGG-39 (AC- Measurement of Nuclear Area
TIN7-terminator forward) and 59-AAGCTGGGTCTAGATATCCGTTTCGCA
TCAGAATGGTAATACAC-39 (ACTIN7-terminator reverse). The projection area of the nucleus was measured as reported previously
ProACTIN7:ACTIN8(G38S) was constructed as described for ProACTIN7: (Iwabuchi et al., 2016).
ACTIN2(G38S) using the following primer sets: 59-AACCAATTCAGTCGA
CGATATATCACGAAAACCGATC-39 (ACTIN7-promoter forward) and 59-
TTTTCACTAAAAAAAAAG-39 (ACTIN7-promoter reverse); 59-TTTTTTTA
Visualization of Actin Filaments
GTGAAAAATGGCCGATGCTGATGACATTC-39 (ACTIN8-genomic forward) Actin filaments were visualized in leaf samples that had been fixed as de-
and 59-TAACAACACTGGGGAAAACC-39 (ACTIN8-G38S reverse); 59- scribed for the visualization of nuclei (Iwabuchi et al., 2010). Samples were
TTCCCCAGTGTTGTTAGTCGACCTAGACATCATGGT-39 (ACTIN8-G38S stained with 200 nM Alexa Fluor 488 phalloidin (Invitrogen) and 5 mg mL21
forward) and 59-TAAGACAAGACACACTTAGAAGCATTTTCTGTGGAC-39 Hoechst 33342, diluted in buffer solution supplemented with 0.03% (v/v) Tri-
(ACTIN8-genomic reverse); and 59-GTGTGTCTTGTCTTATCTGG-39 (AC- ton X-100, for at least 1.5 h.
TIN7-terminator forward) and 59-AAGCTGGGTCTAGATATCCGTTTCGCA Lifeact-Venus/unp1 (F3) and Lifeact-Venus/unp2 (F3) lines were generated
TCAGAATGGTAATACAC-39 (ACTIN7-terminator reverse). by crossing the Lifeact-Venus line and unp1 and unp2, respectively, and actin
The fragments were fused and subcloned into the pENTR1A entry vector filaments were inspected in living cells. The Lifeact-Venus/unp2 line exhibited
using an In-Fusion HD Cloning Kit (Takara). The ProACTIN7:ACTIN2(G38S) abnormally and significantly thicker actin bundles than the Lifeact-Venus line.
and ProACTIN7:ACTIN8(G38S) fragments were introduced into the pFAST- There was also an overaccumulation of Lifeact-Venus protein in the Lifeact-
G01 plant expression vector. Plants were stably transformed with these vec- Venus/unp2 line; therefore, the possibility that the thicker actin-bundle for-
tors by Agrobacterium tumefaciens-mediated transformation. mation was caused by an overaccumulation of Lifeact-Venus rather than by the
unp2 mutation could not be excluded. Given these facts, only phalloidin was
applied for staining unp2.
Generation of Transgenic Plants To visualize actin filaments in transgenic plants expressing Lifeact-Venus,
Expressing GFP-DYRKP-2A leaves were fixed in buffer solution (50 mM PIPES, 10 mM EGTA, and 5 mM
MgSO4, pH 7) containing 0.02% (w/v) paraformaldehyde (Wako) and 0.003%
The DYRKP-2A cDNA sequence was amplified from cDNA of Columbia (w/v) glutaraldehyde (Wako) for 5 min to stop the movement of actin
plants by PCR using KOD Plus Neo (Toyobo). The cDNA fragment was cloned filaments.
into the entry vector, pENTR/D-TOPO (Life Technologies). The sequence of Cells were imaged using a confocal scanning microscope (LSM800; Zeiss),
DYRKP-2A on the entry vector was inserted into a binary vector, pH35GW, and sequential images were taken from the outer periclinal walls to the inner
with the LR reaction using LR clonase II (Life Technologies) to construct the periclinal walls of the cell along the optical z axis at 0.5-mm intervals. Maximum
binary vectors for pro35S:GFP-DYRKP-2A. The transformation was performed intensity projections were generated using the Fiji image-processing package.
by the floral dip method (Clough and Bent, 1998). To select transformants, ster-
ilized seeds were sown on Murashige and Skoog medium (Murashige and Skoog,
1962) containing Gamborg’s B5 vitamins (Gamborg et al., 1968), 3% (w/v) Suc, Measuring the Movements, Occupancy, and Fluorescence
0.8% (w/v) agar, and 10 mg L21 hygromycin (Wako Pure Chemical Industries). Levels of Actin Filaments
The fluorescence of GFP was detected using a confocal laser scanning microscope
(FV1000; Olympus). GFP signal was excited with a 473-nm laser and detected with To analyze lateral movements of the actin filament, pavement cells of
a 490- to 540-nm window. transgenic plants expressing Lifeact-Venus were examined. Actin filaments
were imaged at 5-s intervals for 60 s with a confocal laser scanning microscope
(LSM800). The sequential images were aligned to correct experimental drift
Isolation of the unp2 Mutant and RT-PCR using the Fiji plug-in Align Slices in Stack (https://sites.google.com/site/
qingzongtseng/template-matching-ij-plugin/tuto2#updates), as reported pre-
Mature leaves of Arabidopsis T-DNA-tagged plants were examined with a viously (Tseng et al., 2011). To evaluate the lateral movements of actin fila-
fluorescence microscope (Axioskop 2 plus), and a mutant line that exhibited ments, cross-correlation coefficients between the first image and the subsequent
244 Plant Physiol. Vol. 179, 2019Iwabuchi et al.
time-sequential images were determined with the Fiji plug-in Image Correla- After washing, the gel was treated with 50% (v/v) acetonitrile in 50 mM am-
tionJ (http://www.gcsca.net/IJ/ImageCorrelationJ.html), as reported previ- monium bicarbonate and dried in a vacuum concentrator. The gel was treated
ously (Chinga and Syverud, 2007). with 10 mg mL21 trypsin in 50 mM ammonium bicarbonate and incubated
To evaluate actin filament occupancy in a cell, each pavement cell was overnight at 37°C. The digested peptides were recovered twice with 50 mL of 5%
outlined using Fiji. Actin filaments visualized by phalloidin staining were then (v/v) formic acid in 50% to 70% (v/v) acetonitrile. The extracted peptides were
extracted as skeletonized images using the Fiji plug-in LPX Filter2d (https:// combined and evaporated to 10 mL in a vacuum concentrator.
lpixel.net/products/lpixel-imagej-plugins/), as reported previously (Ueda
et al., 2010). Actin filament occupancy was calculated as the percentage of
pixel numbers of the skeletonized actin filaments versus those of cells using Fiji Mass Spectrometric Analysis and Database Searching
and Excel. Fluorescence levels of the skeletonized actin filaments were deter-
mined using Fiji. Liquid chromatography-tandem mass spectrometry analysis was per-
formed using the LTQ-Orbitrap XL-HTC-PAL system and the Mascot server as
described previously (Tamura et al., 2010).
Measuring the Velocity of Cytoplasmic Streaming
Downloaded from https://academic.oup.com/plphys/article/179/1/233/6116404 by guest on 16 December 2021
Cytoplasmic streaming was examined in pavement cells located above the Yeast Two-Hybrid Assay
midvein of the leaf. To estimate the velocity, particles present in the cytosol were
imaged at 0.13-s intervals for 30 s using a fluorescence microscope (Axio Scope. Gene constructs were prepared as reported previously (Takagi et al., 2013).
A1; Zeiss) equipped with a CCD camera (VB-7010; Keyence). The greatest Briefly, the coding regions of AN, DYRKP-2B, and DYRKP-3 were amplified by
distance traveled by particles in a 5-s interval was determined using Fiji. PCR from wild-type cDNA using the following primer sets: 59-ATGAGCAAGA
TCCGTTCGTCTG-39 (AN forward) and 59-ATCGATCCAACGTGTGATACC-
39 (AN reverse); 59-ATGGCAGACCAAAGCTCTGTTG-39 (DYRKP-2B for-
Measurement of the Circularity Index of Pavement Cells ward) and 59-AGCAGAGATTGGCTCGTATGGG-39 (DYRKP-2B reverse); and
59-ATGGCGGTTGATGTTAAATC-39 (DYRKP-3 forward) and 59-ATTGTAA
Pavement cells were traced using Fiji to determine A and P, where A is the GATGAAGAAGAAG-39 (DYRKP-3 reverse). The amplified fragments were
area and P is the perimeter of the cell. The circularity index of pavement cells subcloned into the pENTR1A entry vector (Invitrogen) that had been digested
was calculated as (4pA)/P2, as reported previously (Iwabuchi et al., 2010). The with SalI and EcoRV using an In-Fusion HD Cloning Kit (Takara). The frag-
circularity index reflects the extent to which a shape approaches a circle (a ments were introduced into destination vector pDEST-GBKT7 or pDEST-
perfect circle has a circularity index of 1). GADT7 via the LR reaction. The Saccharomyces cerevisiae Y2H gold strain
(Clontech) was transformed with these vectors. The transformed yeasts
were first selected on synthetic defined (SD)/-Leu/-Trp plates. The se-
SDS-PAGE and Immunoblot Analysis lected yeasts were examined subsequently on either SD/-Leu/-Trp plates or
SD/-Leu/-Trp/-Ade/-His/+X-a-gal/+aureobasidin A plates at 30°C. As neg-
Protein extracts from mature leaves were subjected to SDS-PAGE followed ative controls, pGBKT7 and pGADT7 vectors were used. The yeast failed to
by immunoblot analysis. Immunoreactive signals were detected with the ECL proliferate when expressing either DYRKP-2A or DYRKP-1.
detection system (GE Healthcare), using an anti-actin antibody (clone 10-B3;
Sigma-Aldrich) at a dilution of 1:2,000, an anti-microtubule antibody (clone B-5-
1-2; Sigma-Aldrich) at a dilution of 1:2,000, and ECL anti-mouse IgG horse- Inhibitor Treatment
radish peroxidase-linked species-specific whole antibody (GE Healthcare) at a
dilution of 1:1,000. Sample leaves were vacuum infiltrated using a syringe containing INDY
SDS-PAGE was performed using a SuperSep Phos-tag gel (Wako) or a (Sigma-Aldrich) solution. For the analysis of blue light-induced nuclear posi-
conventional precast gel (Criterion XT; Bio-Rad). For dephosphorylation, pro- tioning, INDY was applied to the leaves for 1 h before the end of the dark
tein extracts were treated with alkaline phosphatase (Sigma-Aldrich) at a di- treatment, and subsequent light irradiation was performed in the presence of
lution of 1:100 for 60 min at 30°C. Silver staining was performed using a Silver the inhibitor. For the analysis of dark-induced nuclear positioning, sample
Staining Kit (GE Healthcare) according to the manufacturer’s instructions. leaves were irradiated with 100 mmol m22 s21 blue light for 3 h, and subsequent
dark treatment was performed for 5 h in the presence of the inhibitor.
Immunoprecipitation
Statistical Analysis
Immunoprecipitation was performed using a mMACS Epitope Tag Protein
Isolation Kit (Miltenyi Biotec) as reported previously (Tamura et al., 2010). Values are represented as means 6 SE, as determined using StatPlus. P values
Seven-day-old transgenic plants expressing AN-GFP or free GFP were ho- were determined using an unpaired Student’s t test.
mogenized in buffer containing 50 mM Tris-HCl, pH 8, 150 mM NaCl, and 1%
(v/v) Triton X-100. Homogenates were centrifuged at 20,400g at 4°C to remove
cellular debris. The supernatants were mixed with magnetic beads conjugated Accession Numbers
to an anti-GFP antibody (Miltenyi Biotec) and incubated on ice for 15 min. The
mixtures were applied to m Columns (Miltenyi Biotec) in a magnetic field to Sequence data from this article can be found in the GenBank/EMBL
capture the magnetic antigen-antibody complex. After extensive washing with data libraries under the following accession numbers: At5g09810 (ACTIN7),
buffer containing 50 mM Tris-HCl, pH 8, 150 mM NaCl, 1% (v/v) Igepal CA-630 At3g18780 (ACTIN2), At1g49240 (ACTIN8), At1g01510 (AN), At3g17750
(formerly Nonidet P-40), 0.5% (w/v) sodium deoxycholate, and 0.1% (w/v) (DYRKP-1), At1g73460 (DYRKP-2A), At1g73450 (DYRKP-2B), and At2g40120
SDS, immunoaffinity complexes were eluted with buffer containing 50 mM Tris- (DYRKP-3).
HCl, pH 6.8, 50 mM DTT, 1% (w/v) SDS, 1 mM EDTA, 0.005% (w/v) Bromo-
phenol Blue, and 10% (v/v) glycerol. Supplemental Data
The following supplemental materials are available.
Preparation of Peptides for Tandem Mass
Supplemental Figure S1. Nuclear projection areas of mesophyll and pave-
Spectrometry Analysis
ment cells of wild-type, unp1, and unp2 leaves in the dark.
Peptides were prepared for analysis as reported previously (Tamura et al., Supplemental Figure S2. Blue light-induced nuclear positioning in the
2010). For in-gel digestion, the protein components of the immunoprecipitates wild type and unp1 and unp2 mutants.
were separated on a 2.5-cm-long SDS gel. The gel was cut into three fractions.
Each excised gel fraction was treated with 100% (v/v) acetonitrile for 15 min Supplemental Figure S3. Pleiotropic phenotypes of unp1 plants.
and dried in a vacuum concentrator. The dried gel was treated with 10 mM DTT
Supplemental Figure S4. Alignment of actin proteins.
in 50 mM ammonium bicarbonate for reduction, followed by 55 mM
2-iodoacetamide in 50 mM ammonium bicarbonate for 30 min for alkylation. Supplemental Figure S5. Nuclear positioning is not related to cell shape.
Plant Physiol. Vol. 179, 2019 245You can also read

















































