The Arabidopsis Locus RCB Mediates Upstream Regulation of Mitotic Gene Expression1
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Arabidopsis Locus RCB Mediates Upstream
Regulation of Mitotic Gene Expression1
Kristiina Himanen2, Christophe Reuzeau, Tom Beeckman, Siegbert Melzer, Olivier Grandjean,
Liz Corben3, and Dirk Inzé*
Department of Plant Systems Biology, Flanders Interuniversity Institute for Biotechnology, Ghent University,
Technologiepark 927, B–9052 Gent, Belgium (K.H., T.B., L.C., D.I.); CropDesign N.V., B–9052 Gent, Belgium
(C.R.); Université de Liège, Sart-Tilman, B–4000 Liège, Belgium (S.M.); and Institut National de la Recherche
Agronomique, Centre de Versailles, Route de Saint-Cyr, F–78026, France (O.G.)
Transcriptional regulation of cell cycle regulatory genes, such as B-type cyclins, is tightly linked with the mitotic activity in
the meristems. To study the regulation of a B-type cyclin gene, a targeted genetic approach was undertaken. An Arabidopsis
line containing a fusion construct between the CYCB1;1 promoter and a bacterial -glucuronidase marker gene (uidA) was
used in ethyl methanesulfonate mutagenesis. The mutants were screened for altered CYCB1;1::uidA expression patterns. In
a reduced CYCB1;1 expression mutant (rcb), the CYCB1;1::uidA expression was severely affected, being excluded from the
shoot and root apical meristems and leaf primordia and shifted to cells associated with root cap and stomata. In addition
to the overall reduction of the endogenous CYCB1;1 transcript levels, other G2-to-M phase-specific genes were also
down-regulated by the mutation. In the mutant plants, the inflorescence stem growth was reduced, indicating low meristem
activity. Based on the altered CYCB1;1::uidA expression patterns in rcb root meristem, a model is proposed for RCB that
mediates the tissue specificity of CYCB1;1 promoter activity.
The eukaryotic cell cycle is controlled at two major plant-specific CDK that is thought to interact with
checkpoints, one before DNA replication (G1 to S) the mitotic cyclin CYCB1;1 at the G2-to-M transition
and one before mitosis (G2 to M). In yeast (Saccharo- (Criqui et al., 2000) and also both genes show a
myces cerevisiae) and animals, progression through coordinated transcriptional up-regulation during the
these checkpoints is driven by cyclin-dependent ki- G2-to-M phase (Mironov et al., 1999). Three main
nases (CDKs), the activity of which depends on in- classes of cyclins (A, B, and D type) have been de-
teraction with their regulatory subunits, the cyclins. scribed in plants, and in the Arabidopsis genome, a
Different cyclins mediate the cell cycle phase-specific total of 30 cyclins have been identified (Vandepoele
activation of the CDKs and modify their substrate et al., 2002). D-type cyclins are preferentially induced
specificity in a spatial and temporal manner (Stals by mitogen stimuli at the G1 phase (Soni et al., 1995;
and Inzé, 2001). In Arabidopsis, five classes of CDKs Fuerst et al., 1996; Richard et al., 2002). Unlike the
have been identified (Vandepoele et al., 2002), but D-type cyclins, the levels of A- and B-type cyclins are
only two of them have been extensively character- tightly controlled by cell cycle progression. The pro-
ized: CDKA;1 (formerly known as Cdc2a), which is moter activities and transcript levels of A-type cyc-
active at the G1-to-S and G2-to-M checkpoints, and lins have been shown to increase at mid S phase
CDKB1;1 (formerly known as Cdc2b), whose activity (Fuerst et al., 1996; Shaul et al., 1996). For B-type
is highest at the G2-to-M-transition (Mironov et al., cyclins, G2 and M phase-specific peaks in expression
1999; Joubès et al., 2000; Porceddu et al., 2001; have been shown in many plant species, such as
Menges and Murray, 2002). CDKB1;1 represents a Arabidopsis (Ferreira et al., 1994a), Catharanthus ro-
seus (Ito et al., 1998), and Nicotiana sylvestris (Tréhin
1
This work was supported by the Interuniversity Poles of At- et al., 1999).
traction Program (Belgian State, Prime Minister’s Office, Federal As regulatory proteins, cyclins have a high turn-
Office for Scientific, Technical, and Cultural Affairs, grant no. over rate, and their cyclic appearance results from
P5/13), by the Academy of Finland (fellowship to K.H.), and by stringent regulation at the transcriptional level
the Finnish Cultural Foundation for fellowships (fellowship to (Shaul et al., 1996; Criqui et al., 2001). A well-
K.H.). characterized example is the B-type cyclin, CYCB1;1.
2
Present address: Laboratory of Plant Molecular Biology, Rock- Transcriptional regulation of CYCB1;1 is tightly re-
efeller University, 1230 York Avenue, New York, NY 10021.
3 lated to cell cycle activity and can be used as a
Present address: University of East Anglia, Norwich NR4 7TJ,
UK.
marker for detecting mitotically active tissues (Fer-
* Corresponding author; e-mail dirk.inze@psb.ugent.be; fax reira et al., 1994b; Shaul et al., 1996; Colón-Carmona
32–9 –3313809. et al., 1999). The expression of the CYCB1;1 promoter-
Article, publication date, and citation information can be found driven bacterial gus (-glucuronidase) gene (CYCB1;
at http://www.plantphysiol.org/cgi/doi/10.1104/pp.103.027128. 1;;uidA) has been shown to correlate well with the
1862 Plant Physiology, December 2003, Vol. 133, pp. 1862–1872, www.plantphysiol.org © 2003 American Society of Plant Biologists
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.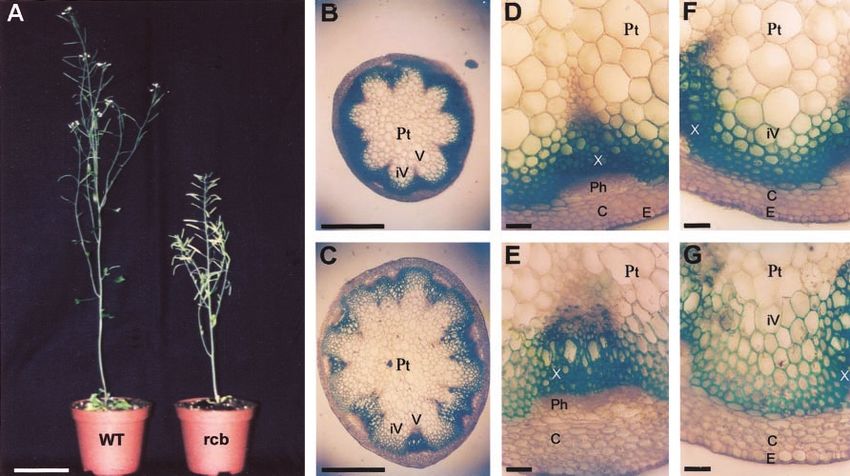
Arabidopsis Locus Affecting Mitotic Gene Expression
mRNA localization of the endogenous gene (de rcb mutation was semidominant. In the F2 popula-
Almeida Engler et al., 1999). The cell cycle-specific tion, three classes of expression patterns were ob-
gene activation of CYCB1;1 is mediated via cis-acting served. When compared with the homozygous wild-
elements in the promoter (Ito et al., 1998, 2001; Tréhin type (RCB/RCB; Fig. 1A) and homozygous mutant
et al., 1999; Planchais et al., 2002). plants (rcb/rcb; Fig. 1B), the heterozygous mutants
Despite extensive analysis of the transcriptional (rcb/RCB; Fig. 1C) had intermediate expression pat-
control of the mitotic cyclin CYCB1;1, little is known terns. As with the homozygous mutants, the het-
about the regulatory pathways that affect its pro- erozygous mutants (F1, 100%; and F2, 50%) showed a
moter activity. In an attempt to study the upstream gain of CYCB1;1::uidA expression in the lateral root
regulation of the CYCB1;1 promoter, a targeted ge- cap. In the root apical meristem, however, interme-
netic approach was undertaken. The CYCB1;1::uidA diate intensities of the CYCB1;1::uidA expression
line was used in chemical mutagenesis, thereby al- were observed, whereas in the homozygous mutants,
lowing visualization of mutations that specifically no expression was detected in that tissue. In the F2
affect the promoter activity. Mutants were screened population, the segregation pattern of ectopic
for altered patterns of CYCB1;1::uidA expression. We CYCB1;1::uidA expression in the root cap again sug-
report the identification and characterization of a gested the semidominant nature of the rcb mutation.
mutant line with reduced CYCB1;1::uidA expression To further confirm that the rcb mutation was inde-
(designated rcb). The rcb mutant lost CYCB1;1::uidA pendent from the CYCB1;1::uidA transgene, test
expression in both shoot and root apical meristems crosses were performed between the rcb mutants and
and leaf primordia, whereas the expression was ec- the C24 wild-type plants. The CYCB1;1::uidA expres-
topically induced in cells associated with root cap and sion pattern in the F1 population resembled that ob-
stomata. In addition, the overall endogenous levels of tained from the backcrosses, although the GUS stain-
the CYCB1;1 transcripts and those of other mitotic ing was often much weaker because of the reduced
genes were reduced in the rcb mutant. In mature amount of CYCB1;1::uidA T-DNA in the progeny. In
plants, the total length of the inflorescence stem and the F2 population, CYCB1;1::uidA-driven GUS stain-
its diameter were significantly affected, indicating an ing patterns typical for wild-type (RCB/RCB, 24%),
effect on the meristem activity. Based on the altered heterozygous (rcb/RCB, 47%), rcb mutant (rcb/rcb,
CYCB1;1::uidA expression patterns in rcb root meris- 24%), and GUS-negative (5%) genotypes were en-
tems, we propose a model in which RCB mediates the countered. This result confirmed that the mutation
tissue specificity of the CYCB1;1 promoter activity.
was independent from the uidA promoter fusion it-
self because the wild-type (RCB/RCB) CYCB1;1::uidA
RESULTS expression patterns could be recovered without in-
troducing wild-type alleles of the uidA construct.
Isolation of the rcb Mutant and Genetic Analysis However, the segregation pattern of the GUS-
To study the regulation of the CYCB1;1 promoter negative plants was only 5% in the F2 population,
activity, 2,000 seeds of the transgenic Arabidopsis indicating that rcb contained two alleles of the uidA
line containing a CYCB1;1::uidA promoter fusion transgene. The segregation pattern of GUS-negative
(Ferreira et al., 1994b) were mutagenized with ethyl lines (5%) was also confirmed by kanamycin selec-
methanesulfonate. The M1 plants were self-fertilized, tion: Only 10 of 200 F2 plants were sensitive to the
and 7,850 of the M2 plants (314 M2 families) were selection. This result again underlines the fact that
screened for altered patterns of CYCB1;1::uidA pro- the rcb mutation resides outside the CYCB1;1::uidA
moter activity in the root tips. From the mutant fusion because the mutation was able to relocalize
screening, one line with rcb expression was chosen the expression of the two CYCB1;1::uidA transgenes.
for further analysis. In this line, the CYCB1;1::uidA To determine the genomic locus of the rcb muta-
expression appeared to be ectopically induced in the tion, genetic mapping was initiated with amplified
root cap cells, whereas it was absent from the root fragment length polymorphism (AFLP) markers (Pe-
apical meristem (Fig. 1, A and B). To confirm that the ters et al., 2001). To introduce mapped markers into
altered CYCB1;1::uidA expression in rcb was not the rcb mutant line, the M3 line of rcb (in Arabidopsis
caused by a mutation in the CYCB1;1::uidA promoter, C24 ecotype) was crossed with Columbia-0 (Col-0)
the wild-type and rcb promoters (1.27 kb) from the ecotype. From a segregating F2 population, 40 mu-
T-DNA constructs were isolated by PCR and se- tants, four wild-type plants, and the parental lines
quenced. The sequences did not differ, indicating were selected by GUS assay of 2-week-old root seg-
that the altered expression was caused by an inde- ments. The selected lines were processed for bulked
pendent mutation. segregant AFLP analysis according to Peters et al.
Before further phenotypic analysis, the mutant line (2001). From each of the 16 primer combinations used,
was subjected to three successive backcrosses with on average five polymorphic markers for each were
the CYCB1;1::uidA starter line. In the F1 population, identified for C24 and Col-0 ecotypes. The bulked
gain of CYCB1;1::uidA expression in the root cap segregant analysis showed linkage with eight markers
was detected in all of the progeny, indicating that the located in the lower arm of chromosome 2 for the rcb
Plant Physiol. Vol. 133, 2003 1863
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Himanen et al.
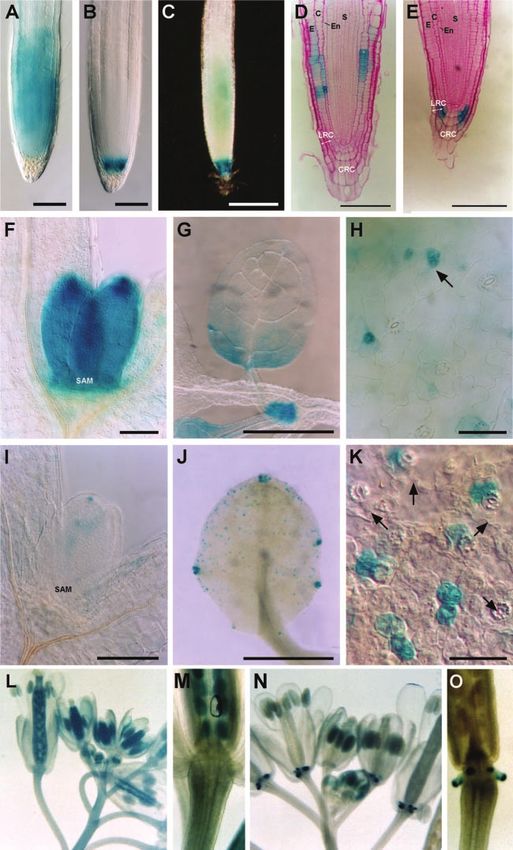
Figure 1. Bright- and dark-field images of whole-
mount GUS-stained plants. A, CYCB1;1::uidA in
wild type. B, CYCB1;1::uidA in rcb mutant. C,
CYCB1;1::uidA in a heterozygous (rcb/RCB) plant
with weak expression in the meristem combined
with root cap expression. Anatomical analysis of
histochemically stained Arabidopsis seedling
roots. D, Longitudinal section (5 m) of 9-d-old
wild-type seedling root in which CYCB1;1::uidA
staining is restricted to epidermal and cortical
cell files. E, Longitudinal section of rcb mutant
with CYCB1;1::uidA expression shifted to cells of
the lateral root cap and absence of expression in
the root apical meristem. Differential interference
contrast microscopy of histochemically stained
leaves at different developmental stages in 4-d-
old seedlings (F–K). F to H, CYCB1;1::uidA in
wild type. I to K, CYCB1;1::uidA in rcb. F and I,
Shoot apices showing the emerging first pair of
leaf primordia (SAM). G and J, Young developing
leaves. H and K, Detailed view of adaxial epider-
mis of developing leaves showing the stomatal
complexes. H, CYCB1;1::uidA expression re-
stricted to developing stomata at the meristemoid
stage in wild type. K, Localization of
CYCB1;1::uidA in palisade cells underneath ma-
ture guard cells; arrows indicate unstained sto-
mata. Expression of CYCB1;1::uidA during inflo-
rescence development (L and M). L, GUS staining
in wild-type inflorescences at different develop-
mental stages. CYCB1;1::uidA is expressed in the
entire pistil. M, Presence of GUS staining in wild-
type ovaries and absence from the nectaries. N,
CYCB1;1::uidA expression in rcb inflorescences.
CYCB1;1::uidA is absent from pistils but strongly
expressed in the nectaries (O). C, cortex; CRC,
columella root cap; E, epidermis; En, endoder-
mis; LRC, lateral root cap; S, stele. Bars ⫽ 200
m (A–C), 100 m (D, E, F, and I), 1 mm (G and
J), and 50 m (H and K).
mutation (data not shown). Also, the non-linkage was patterns of 28:23:28 individuals for the C24 genotype,
analyzed, and the rcb mutation was not linked with 11:19:13 for the heterozygous mutant genotype, and
the polymorphic markers on chromosomes 1, 3, 4, or 5. 1:0:1 for the Col-0 genotype, respectively. These data
The CYCB1;1 gene itself is located in chromosome 4 suggest that the rcb mutation is located on the lower
(At4g37490). In addition, single simple length poly- arm of chromosome 2.
morphism markers (Bell and Ecker, 1994) for each of
the Arabidopsis chromosomes were tested to confirm The CYCB1;1::uidA Expression and CYCB1;1 mRNA
the putative loci. Analysis of 40 mutants with chromo- Patterns in the rcb Mutant
some 2-specific markers (nga1126 [chromosome 2, 51
cM], nga168 [chromosome 2, 74 cM], and an INDEL The rcb mutant was selected on the basis of a
marker in chromosome 2 [79 cM]) gave segregation remarkable change in the CYCB1;1 promoter-driven
1864 Plant Physiol. Vol. 133, 2003
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.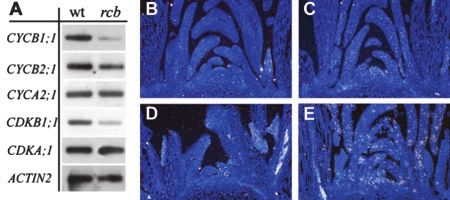
Arabidopsis Locus Affecting Mitotic Gene Expression
GUS staining patterns. To further characterize the changed during root cap development, whereas in
cell specificity of the CYCB1;1::uidA expression in rcb wild-type plants, it was high in the developing lateral
roots, the wild-type expression pattern was analyzed root primordia (Fig. 2A) until starch accumulated in
in anatomical sections. In the primary root apical the newly forming root cap cells (Fig. 2B). Upon dif-
meristem of the wild-type plant, the CYCB1;1::uidA ferentiation of statocyte layers in the columella, the
expression was detected in a cell type-specific man- CYCB1;1::uidA expression diminished (Fig. 2C), and in
ner in the epidermal and cortical cell files (Fig. 1D),
whereas in the newly developed meristems of lateral
root primordia, the expression was uniform (Ferreira
et al., 1994b; Beeckman et al., 2001). In rcb, this ex-
pression pattern was severely altered. The expression
was absent from the epidermal and cortical cell files
in the root apex but was ectopically induced in the
lateral root cap initials and their immediate daughter
cells (Fig. 1E), where no expression in the wild type
could be detected. Other meristematic tissues of
wild-type and rcb plants were analyzed to see
whether the effect of rcb mutation was root specific.
Wild-type shoot apical meristem and emerging leaf
primordia expressed CYCB1;1::uidA strongly (Fig.
1F). In young wild-type leaf primordia, the basipetal
gradient of cell division activity could be observed
(Fig. 1G; see also Donnelly et al., 1999). During the
wild-type stomata development, the expression was
restricted to the meristemoids (Fig. 1H; see also Serna
and Fenoll, 1997). In rcb, the meristematic cells in the
shoot apex, leaf primordia, and young leaves had no
CYCB1;1::uidA promoter activity (Fig. 1I). Instead, in
rcb, strong expression was present in the area of
hydathodes (Fig. 1J) and ectopically induced in the
palisade parenchyma cells beneath each developing
stomatum (Fig. 1K).
Also, in rcb flowers, an altered CYCB1;1::uidA ex-
pression was observed. In the flowers of wild-type
plants, CYCB1;1::uidA expression was strong in de-
veloping pistil, excluding the stigma and nectaries
(Fig. 1, L and M). In contrast, in the rcb mutant line,
no GUS staining was detected in the pistil, whereas
the nectaries were strongly stained (Fig. 1, N and O).
Thus, in different tissues and organs of the rcb
mutant, a shift in CYCB1;1::uidA localization takes
place in comparison with the wild type. Both in the
root and shoot apical meristems, the strong meris-
tematic CYCB1;1::uidA expression was lost in rcb,
whereas an ectopic expression was induced in tissues
that usually have no mitotic activity. In addition, the
semidominant nature of the rcb mutant indicates that
RCB could encode a dose-dependent activator of the
CYCB1;1 promoter in meristematic tissues and per-
haps a repressor outside the meristems. Figure 2. Comparative analysis of CYCB1;1::uidA expression during
lateral root development and de novo root cap formation under dark-
field stereoscopy. A to D, Young lateral root primordia, emerging
lateral roots, outgrowing lateral roots, and lateral root with fully de-
Root Cap Maturation in rcb veloped root caps in wild type, respectively. E to H, Young lateral root
primordia, emerging lateral roots, outgrowing lateral roots, and lateral
In the rcb roots, the CYCB1;1::uidA expression was root with fully developed root caps in rcb mutant, respectively. F,
affected in a cell type-specific manner, namely ec- Statoliths (arrow) become refractive under dark-field optics. G,
topically induced in the lateral root cap initial cells. CYCB1;1::uidA is only expressed (arrow) in rcb at the moment stato-
We analyzed the CYCB1;1::uidA expression in more cyte organization starts to appear. I and J, CYCB1;1::uidA expression in
detail in wild-type and rcb roots at different stages of 2,4-dichlorophenoxyacetic acid-treated seedling roots of wild type
root cap development: In rcb, it characteristically and rcb, respectively. Bars ⫽ 1 mm (A–H), 200 m (I and J).
Plant Physiol. Vol. 133, 2003 1865
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Himanen et al.
mature wild-type root caps, it was undetectable (Fig. were grown under short- (8 h/16 h) and long- (16 h/8
2D). In contrast, in rcb, the CYCB1;1::uidA expression h) day light conditions. Growth was analyzed by com-
followed an opposite pattern and appeared to be paring the number and size of rosette leaves, number
tightly linked with the development of the statocyte of rosette and lateral branches, plant height, flowering
tissues. In young lateral root primordia (before devel- time, and flower development. Observations were
opment of the statocytes), CYCB1;1::uidA was not ex- made on 40 plants in at least three replicates for the
pressed (Fig. 2, E and F). The CYCB1;1::uidA expres- growth conditions. Although under constant light
sion in lateral root caps of rcb appeared with the conditions no consistent growth phenotypes were de-
maturation of the statocytes in the columella (Fig. 2G). tected, under short- and long-day conditions, the
At the time the statocyte layers in the columella were growth of rcb plants was considerably altered when
fully developed, the lateral root cap cells showed compared with that of the wild-type controls (Table I).
strong GUS staining, whereas the columella remained The growth of rcb plants was repressed at the stage of
unstained (Fig. 2H). inflorescence stem development, which were approx-
In rcb mutants, the CYCB1;1::uidA expression was imately 25% shorter in height and more rigid than the
absent from the root apical meristem and was exclu- wild-type plants (Fig. 3A). Flowering time in rcb was
sively associated with the root cap-specified zone. delayed by approximately 6 d. Rosette leaf growth
We tested two conditions under which the size and was similar to that of wild-type controls, without any
cell patterning of root meristems were severely al- significant difference in leaf development and leaf
tered. Treatment with the auxin transport inhibitor number. Rosette branch development was reduced,
naphthylphthalamic acid is known to cause spanning whereas lateral branches developed as those of control
of tissue with root cap identity upwards from the plants. The growth of the CYCB1;1 knockout plants
root tip (Sabatini et al., 1999). In the rcb mutant, the was comparable with that of the wild-type plants,
treatment with 10⫺5 m naphthylphthalamic acid led with only a significantly reduced number of leaves
to expansion of the root cap with the characteristic and rosette branches in the mutant plants (Table I).
CYCB1;1::uidA expression for rcb (data not shown). Fur- In rcb, the silique length and diameter were signif-
thermore, synthetic auxin 2,4-dichlorophenoxyacetic icantly shorter (40%) and larger than those of wild-
acid was used to induce ectopic cell division in the type plants, respectively (Table I). Furthermore, un-
meristem. In both wild-type and rcb plants, treatment der both long- and short-day conditions, the stem
with 2,4-dichlorophenoxyacetic acid increased the mer- diameter increased significantly in rcb plants from
istematic tissue in both plant types and induced a the basis of the rosette when the flowering stem was
strong expression of CYCB1;1::uidA in the expanded 2 d old (1-cm total height) until the end of the devel-
meristem of wild-type plants (Fig. 2I) but not in the rcb opment. An increase in diameter of the lateral
root. In the mutant, the typically restricted pattern of branches of rcb was similarly observed.
CYCB1;1::uidA expression was observed (Fig. 2J). To identify the origin of the stem diameter in-
crease, the stems were sectioned at different posi-
Shoot Phenotype tions, starting from 1 cm above the rosette (below any
lateral branches) and at approximately 10 cm from
To analyze the shoot-specific growth phenotypes of the rosette. The rcb stem sections were larger but had
rcb, the mutant and CYCB1;1::uidA wild-type plants a normal radial symmetry consisting of an epidermal
(C24 ecotype) and the CYCB1;1 knockout line from layer, four to five layers of chlorenchyme, a layer of
the Versailles T-DNA collection and the correspond- alternating vascular bundles and intervascular fibers,
ing wild-type plants (Wassilewskija [Ws] ecotype) and a central zone. In control plants, the average
Table I. Phenotypic measurements of wild-type C24 and rcb mutants under short-day conditions 11 weeks after sowing
Values were averaged from five sections for each sample. Observations were made on 40 plants in at least three different
replicates.
Phenotypic trait CYCB1;1::uidA (C24) rcb (C24) Wild Type (Ws) CYCB1;1 Mutant (Ws)
Longest rosette leaf (cm) 81.6 80.5 79.3 80.4
No. of rosette leaves 18 16 59.6 28.5
Plant height (cm) 22.2 16.5 35.5 30.6
No. of rosette branches 6.4 4.1 11.1 2.6
No. of lateral branches 7.1 7.3 7.2 10.1
Time to anthesis (d) 78 83 73 72
Silique length (mm) 14.3 8.6 14.5 14.4
Cells in inflorescence stem
a
No. of cells in pith 24.1 29.8 ND ND
No of cells in cortex 5.0 5.2 ND ND
No. of interfascicular cells 5.2 9.1 ND ND
a
ND, Not determined.
1866 Plant Physiol. Vol. 133, 2003
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Arabidopsis Locus Affecting Mitotic Gene Expression
Figure 3. Shoot phenotype. A, Wild-type and rcb plants grown under long-day conditions. B and C, Hand sections of 5- to
6-week-old wild-type C24 and rcb plants, respectively. D and E, Transverse section of the basal part of the stem showing
a vascular bundle of wild type and rcb, respectively. F and G, Transverse section of the basal part of the stem showing the
interfascicular region of wild type and rcb, respectively. C, cortex; E, epidermis; iV, interfascicular region; P, phloem; Pt,
pith; V, vascular bundle; X, xylem. Bars ⫽ 0.5 mm (B), 1 mm (C), and 10 m (D–G).
number of vascular strands in the stem sections was Himanen et al. (2002). One week after germination,
nine, instead of an average of 11 in rcb plants (Fig. 3, seedlings were used for RNA extraction. The mutant
B and C). The structure of the vascular strands in rcb had reduced CYCB1;1 transcript levels when com-
differed from that of control plants by larger vascular pared with those of wild-type plants, in agreement
bundles at the base of the stem and at one-third of the with the reduced GUS activity. In addition to down-
overall stem height. The difference in shape of the regulation of CYCB1;1, other G2-to-M and M phase-
vascular bundles appears to be due to an increase in specific genes, such as CYCB2;1 and CDKB1;1, also
cell size both in the phloem and in the xylem regions showed similarly reduced transcript levels (Fig. 4A).
(Fig. 3, D and E). Also, the cells between the vascular Markers of the G1-to-S cell cycle phase, such as
bundles were enlarged, contributing to the overall CYCA2;1 and CDKA;1, were not affected by the mu-
expansion of the stem diameter (Fig. 3, F and G). The tation. To further confirm the reduced endogenous
number of cells in the inflorescence stem was esti- transcript levels of CYCB1;1, in situ hybridization was
mated by calculating the number of cells from 10 performed on shoot apices from wild-type and mutant
images taken of sections cut 1 cm above the rosette.
Although the cell numbers did not change in the
cortex, in the central stele, and especially in the in-
terfascicular zone, considerably more cells were
present in rcb than in the wild type (Table I).
Kinematic analysis of root growth from germina-
tion until 2 weeks of age, quantification of the num-
ber of lateral roots, and flow cytometric analysis of
root and leaf tissues did not reveal any differences
between rcb and wild-type plants (data not shown).
Figure 4. Reverse transcription (RT)-PCR gel blot. A, Transcript level
rcb Down-Regulates a Set of Mitotic of the cell cycle regulatory genes (CYCB1;1, CYCB2;1, CYCA2;1,
Genes in Arabidopsis CDKA;1, CDKB1;1, and ACTIN-2) in 1-week-old wild-type and rcb
seedlings. B to E, In situ hybridization of CYCB1;1 mRNA on shoot
To test whether the rcb mutation affected other cell apical meristem tissue of rcb mutant and wild-type plants. B, rcb
cycle genes, transcript levels of CYCA2;1, CYCB1;1, CYCB1;1 sense probe. C, rcb hybridized with CYCB1;1 antisense
CYCB2;1, CDKA;1, and CDKB1;1 were analyzed by probe. D, Wild type hybridized with CYCB1;1 sense probe. D, Wild
semiquantitative RT-PCR as described previously by type hybridized with CYCB1;1 antisense probe.
Plant Physiol. Vol. 133, 2003 1867
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Himanen et al.
plants grown under conditions described by Corbesier ity, CYCB1;1:uidA, is ectopically induced in the lat-
et al. (1996). In the rcb shoot apical meristem, no signal eral root cap cells. However, no differences in the
of CYCB1;1 mRNA could be detected with antisense or root cap size or structure are observed in the mu-
sense probes (Fig. 4, B and C). In wild-type plants, the tants, indicating that the ectopic CYCB1;1::uidA ex-
localization of CYCB1;1 mRNA was patchy when hy- pression alone is not adequate to drive additional cell
bridized with antisense probes, whereas the sense divisions in these cells.
probes gave no signal (Fig. 4, D and E).
In rcb Inflorescence, Meristem Activity Is Affected But
DISCUSSION Not Organ Initiation
rcb Shows an “Inverse” Development of the Based on the phenotypic analysis of the rcb mutant,
CYCB1;1::uidA Expression Pattern during Root Cap we conclude that in addition to the regulation of
Maturation Compared with the Wild Type CYCB1;1 tissue specificity in roots, RCB is necessary
for inflorescence meristem activity. Plant develop-
The most striking phenotype of the rcb mutant
ment is generally due to a balance between cell divi-
was the ectopic expression of CYCB1;1::uidA ob-
sion in meristematic tissues to produce organs and
served in the root cap where wild-type plants do not
cell growth and differentiation to allow organ growth
show any expression. Our results indicate that the
and maturation. The stem phenotype appeared
CYCB1;1::uidA expression pattern is strictly localized
rather late, during rosette and inflorescence growth,
in specific cell types, depending on the developmen-
namely at the onset of inflorescence stem growth.
tal stage of the particular organ. In the wild-type root
Because the number of lateral organs remained un-
apical meristem, the CYCB1;1::uidA is expressed in
altered, the size and the early functioning of the
the dividing cortical and epidermal cell files, whereas
shoot apical meristem were probably not strongly
in the rcb, no expression is detected. However, dur-
affected. However, the meristem function may be
ing the lateral root development in rcb, the
modified at a later stage, perhaps by affecting the cell
CYCB1;1::uidA expression is only induced upon root
production rate in the meristem responsible for in-
cap maturation and becomes restricted to the lateral
florescence stem growth. Furthermore, increased
root cap cells, whereas it is not expressed in the
number of cells in the transverse sections of the in-
mature root cap of the wild type. In pea (Pisum
florescence stems was detected, suggesting that these
sativum), the root cap meristem is regulated indepen-
growth activities are regulated independently from
dently from the primary root apical meristem and is
each other.
programmed to produce a species-specific amount of
In the flower organs of wild-type plants,
root cap cells (Hawes et al., 1998). When the organ
CYCB1;1::uidA expression was found in the young tip
reaches a certain size, cell production ceases. The
of the pistil where actively dividing cells are present
cells differentiate progressively through a series of
(Liu et al., 1997; Broadhvest et al., 2000). In rcb, the
developmental stages until the cells at the periphery
CYCB1;1::uidA expression pattern is different because
of the root cap separate as border cells. The separa-
it is shifted from the pistil to the stamen. The siliques
tion of these metabolically active cells depends on the
of rcb were also significantly shorter and wider than
environmental conditions, such as water potential.
those of the wild-type lines, indicating that RCB was
When incubated in water with gentle agitation, the
necessary to maintain CYCB1;1 gene expression in
border cells respond immediately by expansion and
pistils of the wild-type plants, and lack of CYCB1;1
release from the root cap, whereas under dry condi-
expression in rcb pistils repressed the mature silique
tions, they remain attached to the root. When the
length.
border cells are still attached to a mature root cap, the
root cap meristem arrests in the G1 phase of the cell
cycle (Brigham et al., 1998). As a sign of the G1 arrest, Redundancy in Plant Cell Cycle
the mature root cap cells fail to express the histone
H2 gene (Tanimoto et al., 1993). However, upon re- The rcb mutation affected various organs. In roots
moval of the border cells, a cell division marker gene and leaves, where changes in CYCB1;1::uidA expres-
is induced within 15 min (Woo and Hawes, 1997). sion were detected, no growth phenotypes were ob-
In Arabidopsis, the border cells appear to be tightly served. On the other hand, the inflorescence stem
associated with the root cap and are not released had a strong growth phenotype probably because of
during water agitation treatment (Hawes et al., 1998). the effect of the rcb mutation on the expression of
The slow growth rate of the root cap indicates that CYCB1;1 in the shoot apex. In addition, a similar
the cell division activity is generally low. This con- response was observed in the siliques, which re-
clusion is supported by lack of CYCB1;1::uidA (our mained shorter but broader than in wild type. Inter-
observations) and CYCA2;1 expression, as well as by estingly, the phenotype of a CYCB1;1 knockout mu-
thymidine labeling in wild-type root caps (Burssens tant differed from that observed in rcb, showing a
et al., 2000a). In rcb, an opposite development is decrease in number of rosette leaves and rosette
observed, because the marker gene for mitotic activ- branches. These observations indicate that the tissue
1868 Plant Physiol. Vol. 133, 2003
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Arabidopsis Locus Affecting Mitotic Gene Expression
specificity of CYCB1;1 expression and CYCB1;1 CYCB1;1::uidA in the cortical and epidermal cells in
knockout had a different effect on the shoot apical wild-type root apical meristems. However, based on
meristem activity. database information of Arabidopsis, at least 30 cyc-
In addition to the down-regulation of CYCB1;1, the lins are predicted (Vandepoele et al., 2002). Such a
rcb mutation also down-regulated the M phase- high number of cyclins is more than that reported for
specific genes (CYCB2;1, and CDKB1;1) at the tran- any other organism to date. The actual function of all
scriptional level. The fact that the rcb mutant was still the cyclins still remains to be studied, but plant-
able to grow indicated that the functions of these specific features of growth (no cell migration) and
genes have probably been taken over by other genes. development (postembryonic organ development)
These data suggest a high level of redundancy for cell may require special regulation of the cell cycle. To
cycle genes in plants. Similar situations are found in what extent these numerous plant cyclins can confer
yeast with three G1 cyclins (Cln1, Cln2, and Cln3). functional redundancy between each other still re-
Only loss-of-function of the Cln3 delays slightly en- mains to be elucidated.
try into S phase; however, cells that are double mu-
tants for certain combinations of these genes have a
more severe cell cycle delay than the single mutants RCB May Have a Dual Function in Regulating the
(Thomas, 1993). Also, the four mitotic B-type cyclins Tissue Specificity of the CYCB1;1 Expression
of yeast (Clb1, Clb2, Clb3, and Clb4) are highly re-
dundant (Tjandra et al., 1998). Only deletion of Clb2 We aimed at identifying direct regulators of
retards mitosis, whereas any of the other three mi- CYCB1;1 promoter activities. In the rcb mutant, iden-
totic cyclins can be deleted without noticeable phe- tified from the screen of ethyl methanesulfonate-
notypes (Amon et al., 1993). Nevertheless, only dele- mutagenized CYCB1;1::uidA plants, a shift in tissue
tion of the three other genes can create a Clb2- specificity of GUS activity was encountered, indicat-
ing that RCB itself may be situated upstream of
dependent genetic background. In addition, the yeast
the CYCB1;1 promoter regulation. Based on the
cyclins appear to be capable of complementing each
CYCB1;1::uidA expression patterns in wild-type and
other’s functions, despite the cell cycle phase in
rcb mutants, we propose a model for the function of
which they normally function. In yeast, two addi-
RCB. In this model, RCB plays a dual role both as a
tional B-type cyclins, Clb5 and Clb6, are produced at
positive regulator for CYCB1;1 promoter activation in
the onset of the S phase, but they still display func-
the outer layers of the root apical meristem and as a
tional redundancy with the other four B-type cyclins repressor in lateral root cap cells (Fig. 5A). This
(Segal et al., 1998). conclusion derives from the observed loss of
In mammals, such flexibility does not seem to exist. CYCB1;1::uidA expression in the root apical meristem
In mice (Mus musculus), deletion of one of the two and, on the other hand, on the gain-of-expression in
B-type cyclins (CycB1) is lethal, although the function the lateral root cap in rcb (Fig. 5B). Further support
of the other, CycB2, can be compensated (Brandeis et for the model comes from the analysis of the het-
al., 1998). In animal systems, transcriptional up- erozygous mutants in which the meristematic expres-
regulation of the B-type cyclins have been shown to sion has been reduced to an intermediate level in
play a central role in regulating the entry into mitosis combination with a gain-of-expression in the root cap
(Murray and Kirschner, 1989; Nurse, 1990). In plants, (Fig. 5C). It is interesting that a similar change in
CYCB1;1 has been suggested to be the main mitotic CYCB1;1::uidA expression patterns has been ob-
cyclin (Doerner et al., 1996). Ectopic expression of served in the shoot of rcb. The typically strong ex-
CYCB1;1 stimulates cell division activity in root api- pression pattern of wild-type plants in the shoot
cal meristems, indicating that the level of CYCB1;1 is apical meristem is absent in rcb; instead, a shift to
a limiting factor for the entry into mitosis. However, parenchyma cells beneath the developing stomata is
the absence of severe growth phenotypes in rcb ar- observed. It is tempting to speculate that RCB en-
gues against this conclusion. Clearly, reduced levels codes a trans-acting factor that would act both as an
of CYCB1;1, CYCB2;1, and CDKB1;1 had no dramatic activator and a repressor, depending on the interac-
effects on cell cycle progression. Other B-type, or tion with other proteins. The existence of trans-acting
even A-type, cyclins probably take over the role of factors, which act both positively and negatively on
cyclins with defective function. the expression of cell cycle genes, is not unprece-
Not much is known about the redundancy between dented. In animals, the heterotrimeric transcription
plant cyclins. Immunolocalization studies in maize factor NFY has been shown to transcriptionally in-
(Zea mays) have revealed different subcellular local- hibit CycB1, CycB2, and Cdc25 promoters upon DNA
izations for several closely related plant cyclins damage-induced G2 arrest (Manni et al., 2001). In
(Mews et al., 1997, 2000), and strictly localized pat- addition to negative regulation, CCAAT-binding
terns also have been reported for the D-type cyclins transcription factors also mediate positive regulation
in the floral meristems of snapdragon (Antirrhinum of cell cycle phase-specific CycB1 expression (Katula
majus; Gaudin et al., 2000). In addition, we have et al., 1997; Sciortino et al., 2001). Together with other
shown strictly tissue-specific expression patterns for transcription factors, NFYs appear to mediate balanc-
Plant Physiol. Vol. 133, 2003 1869
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Himanen et al.
Figure 5. Model for the putative role of the
wild-type RCB and the effects of homozygous
and heterozygous rcb mutations. A, RCB activa-
tor for CYCB1;1 genes in the root apical meris-
tem and repressor for the same gene in the
lateral root cap tissues in wild type. B, In the
homozygous rcb mutant, lack of CYCB1;1 ex-
pression in the root apical meristem and ectopi-
cal induction in the lateral root cap. C, In the
heterozygous mutant, induction of intermediate
CYCB1;1 expression in the root apical meristem
and in the root cap cells.
ing between activation and repression of genes in a compared with the promoter from the mutant starter line in multiple align-
ments (GCG Wisconsin Package; Accelrys, San Diego, CA).
tissue-specific manner (Gilthorpe et al., 2002).
MATERIALS AND METHODS Genetic Analysis
Plant Material For mapping of the rcb mutant, an AFLP mapping system was used (Vos
et al., 1995). Homozygous mutant plants from the M3 population were
The transgenic Arabidopsis line CYCB1;1::uidA (Ferreira et al., 1994b) was crossed with the parental line of the wild-type Col-0 ecotype, for which
in the genetic background of the C24 ecotype. For in vitro culture, seeds AFLP markers were available. The F1 plants were self-fertilized, and the
were sterilized and sown on K1 germination media (Valvekens et al., 1988) segregating F2 progenies were screened by GUS assay to select mutant
without vitamins. Plants were grown in a growth chamber with continuous individuals for mapping. AFLP reactions were performed on total genomic
light (110 E m⫺1 s⫺1 photosynthetically active radiation supplied by DNA isolated from 40 mutant plants, four wild-type individuals, and the
cool-white fluorescent tubes) held at 22°C. Plants were grown on square two parental lines with the DNeasy protocol (Qiagen). The mutant DNA
plates (Greiner Labortechnik, Frickenhausen, Germany) in a vertical posi- samples were pooled in bulks of eight and were subjected to digestion with
tion to facilitate the accessibility to the root system. Material was collected SacI and MseI enzymes, together with the wild-type and parental samples.
at various time points depending on the requirements of each experiment. Pre-amplification reactions were done using SacI and MseI primers without
For phenotypic characterization, seedlings were used 36 h, 4 d, 1 week, or 2 selective nucleotides, and the AFLP fingerprints were generated with two
weeks after germination. After 3 weeks of sterile culture, plants were selective nucleotides added to each primer. Adaptors and primers were
transferred to a mixture of soil and vermiculite (1:3 [w/v]) for self- obtained from Genset (Paris). The SacI primers were 33P-labeled for visual-
fertilization under the same growth conditions. ization of the fragments by autoradiography. The AFLP banding patterns
from 16 primer combinations were scored for presence or absence of bands
representing polymorphic markers between Col-0 and C24 ecotypes. The
Mutagenesis map position was deduced based on linkage and non-linkage of the markers
for the mutation. The chromosomal location of the uidA T-DNA was deter-
The Arabidopsis CYCB1;1::uidA promoter fusion line was chemically mined from markers that were present in the mutant pools but absent from
mutagenized with ethyl methanesulfonate. Treatment with 0.3% (w/v) the four wild-type individuals. The marker analysis was performed by
ethyl methanesulfonate was performed for 12 h, after which the seeds were standard PCR as described by Bell and Ecker (1994).
extensively washed with water. M1 plants were germinated on K1 medium
and transferred to soil after 2 weeks of growth for self-fertilization. M2
progenies were analyzed in a GUS assay-based mutant screening. From each In Situ Hybridization
line, 25 seeds were germinated together with wild-type controls. Root
cuttings were collected for the GUS assay after 1 week of growth. The lines For in situ hybridization, the plant material was grown according to the
were screened for alterations in the pattern or intensity of the CYCB1;1::uidA method of Corbesier et al. (1996). Longitudinal sections of shoot apices from
expression. The selected putative mutant lines were self-fertilized, and the 2-month-old plants were hybridized as described by Segers et al. (1996). The
M3 plants were subjected to three successive backcrosses with the starter CYCB1;1 [35S]UTP-labeled antisense and sense probes were prepared from
line before phenotypic analysis. Intactness of the CYCB1;1::uidA promoter the linearized pGEMCYC1At vector using SP6 and T7 RNA polymerase,
fusion in the mutants was controlled by sequencing. The promoters from respectively.
both wild type and rcb were isolated by standard PCR amplification with Pfu
(Promega, Madison, WI). Primers were designed to hybridize with the flank-
ing sequences on each side of the actual CYCB1;1 promoter in the Histochemical GUS Assays
CYCB1;1::uidA construct and inside the promoter (5⬘CGCGATCCAGACT-
GAATGCCCACAGGCCG with 5⬘CGTGCCACGCGCTACAGACCACGCCC For histochemical GUS assays, complete seedlings or root cuttings were
and 5⬘GGGCGTGGTCTGTAGCGCGTGGCACG with 5⬘GCCTGGGGTGC- stained in multiwell plates (Falcon, Becton-Dickinson, Bedford, MA). GUS
CTAATGAGTGAGAATTGACGG). PCR products were run on 0.9% (w/v) assays were performed as described by Beeckman and Engler (1994). Sam-
agarose gels, purified with Qiaquick Gel Extraction Kit (Qiagen, Hilden, ples mounted in Tris-saline buffer or lactic acid were observed and photo-
Germany), and sequenced. Sequences from three replicate samples were graphed with a stereoscope (Stemi SV11, Zeiss, Jena, Germany) or with
1870 Plant Physiol. Vol. 133, 2003
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Arabidopsis Locus Affecting Mitotic Gene Expression
differential contrast optics on a standard light microscope (Leica, Wien, fertile whereas cyclin B1-null mice die in utero. Proc Natl Acad Sci USA
Austria). 95: 4344–4349
Brigham LA, Woo H-H, Wen F, Hawes MC (1998) Meristem-specific sup-
pression of mitosis and a global switch in gene expression in the root cap
Microscopy of pea by endogenous signals. Plant Physiol 118: 1223–1231
Broadhvest J, Baker SC, Gasser CS (2000) SHORT INTEGUMENTS 2 pro-
For anatomical sections, samples from the GUS assays were fixed over- motes growth during Arabidopsis reproductive development. Genetics
night in 1% (v/v) glutaraldehyde and 4% (w/v) paraformaldehyde in 50 155: 899–907
mm phosphate buffer. Samples were dehydrated and embedded in Techno- Burssens S, de Almeida Engler J, Beeckman T, Richard C, Shaul O,
vit 7100 resin (Heraeus Kulzer, Wehrheim, Germany) according to the Ferreira P, Van Montagu M, Inzé D (2000a) Developmental expression of
manufacturer’s instructions. For proper orientation of the samples, trans- the Arabidopsis thaliana CycA2;1 gene. Planta 211: 623–631
parent strips were used to facilitate alignment of the tissues (Beeckman and Burssens S, Himanen K, van de Cotte B, Beeckman T, Van Montagu M,
Viane, 2000). Sections of 5-m were cut with a microtome (Minot 1212, Leitz, Inzé D, Verbruggen N (2000b) Expression of cell cycle regulatory genes
Wetzlar, Germany), dried on object glasses, counterstained for cell walls and morphological alterations in response to salt stress in Arabidopsis
with 0.05% (w/v) ruthenium red (Fluka Chemical, Buchs, Switzerland) in thaliana. Planta 211: 632–640
tap water for 30 s, mounted in DePex, and covered with coverslips for
Colón-Carmona A, You R, Haimovitch-Gal T, Doerner P (1999) Spatio-
analysis and photography.
temporal analysis of mitotic activity with a labile cyclin-GUS fusion
protein. Plant J 20: 503–508
Corbesier L, Gadisseur I, Silvestre G, Jacqmard A, Bernier G (1996) Design
Whole-Plant Analysis, Anatomical
in Arabidopsis thaliana of a synchronous system of floral induction by one
Sections, and Microscopy long day. Plant J 9: 947–952
Criqui MC, Parmentier Y, Derevier A, Shen W-H, Dong A, Genschik P
Plants were grown at 20°C in small pots on soil in the greenhouse under
(2000) Cell cycle-dependent proteolysis and ectopic overexpression of
long-day conditions (16 h of light/8 h of darkness) with sodium lamps and
cyclin B1 in tobacco BY2 cells. Plant J 24: 763–773
at 15°C under short-day conditions (8 h of light/16 h of darkness) conditions
Criqui MC, Weingartner M, Capron A, Parmentier Y, Shen W-H, Heberle-
with cool-white lights (75–100 E m⫺2, Philips, Eindhoven, The Nether-
Bors E, Bögre L, Genschik P (2001) Sub-cellular localisation of GFP-
lands). Stem samples taken at 1 and 10 cm above the rosette were sectioned
tagged tobacco mitotic cyclins during the cell cycle and after spindle
manually. Tissues samples were treated 20 min in 50% (v/v) sodium hypo-
checkpoint activation. Plant J 28: 569–581
chlorite, washed in water for 30 min, fixed for 5 min in acetic acid, stained
de Almeida Engler J, De Vleesschauwer V, Burssens S, Celenza JL Jr, Inzé
for 5 min in 0.1% (w/v) toluidine blue, and washed in tap water. Samples
D, Van Montagu M, Engler G, Gheysen G (1999) Molecular markers and
were examined through a Wild M3 Microscope (Leica) and photographed
cell cycle inhibitors show the importance of cell cycle progression in
under bright field. Data were collected and analyzed with Optimas 6.0
nematode-induced galls and syncytia. Plant Cell 11: 793–808
(MediaCybernetics, http://www.optimas.com/optimas.htm).
Doerner P, Jørgensen J-E, You R, Steppuhn J, Lamb C (1996) Control of
root growth and development by cyclin expression. Nature 380: 520–523
RT-PCR Donnelly PM, Bonetta D, Tsukaya H, Dengler RE, Dengler NG (1999) Cell
cycling and cell enlargement in developing leaves of Arabidopsis. Dev Biol
Endogenous transcript levels of a set of cell cycle regulatory genes were 215: 407–419
analyzed by semiquantitative RT-PCR as described by Burssens et al. Ferreira P, Hemerly A, de Almeida Engler J, Bergounioux C, Burssens S,
(2000b) and Himanen et al. (2002). For the cDNA synthesis, total RNA was Van Montagu M, Engler G, Inzé D (1994a) Three discrete classes of
prepared with RNeasy reagents (Qiagen) from 1-week-old wild-type and rcb Arabidopsis cyclins are expressed during different intervals of the cell
seedlings. The cDNA was prepared from three independent RNA samples, cycle. Proc Natl Acad Sci USA 91: 11313–11317
and 15, 20, and 25 cycles of PCR were tested to verify the exponential phase Ferreira PCG, Hemerly AS, de Almeida Engler J, Van Montagu M, Engler
of the amplification. G, Inzé D (1994b) Developmental expression of the Arabidopsis cyclin
gene cyc1At. Plant Cell 6: 1763–1774
Fuerst R, Soni R, Murray J, Lindsey K (1996) Modulation of cyclin tran-
ACKNOWLEDGMENTS script levels in cultured cells of Arabidopsis thaliana. Plant Physiol 112:
1023–1233
The authors acknowledge Christiane Genetello for the effort in maintain- Gaudin V, Lunness PA, Fobert PR, Towers M, Riou-Khamlichi C, Murray
ing the mutant lines during the project, Janny Peters and Tom Gerats for JAH, Coen E, Doonan J (2000) The expression of D-cyclin genes defines
help in determining the map position of the RCB locus, Katia Belcram for her distinct developmental zones in snapdragon apical meristems and is
technical contribution in the identification and analysis of shoot phenotypes, locally regulated by the cycloidea gene. Plant Physiol 122: 1137–1148
Nathalie Detry for in situ hybridizations, Rebecca Verbanck for artwork, Jan Gilthorpe J, Vandromme M, Brend T, Gutman A, Summerbell D, Totty N,
Zethof for INDEL primers, Meeta Mistry for critical reading of the manu- Rigby PWJ (2002) Spatially specific expression of Hoxb4 is dependent on
script, and Martine De Cock for help in preparing it. the ubiquitous transcription factor NFY. Development 129: 3887–3899
Received May 21, 2003; returned for revision June 25, 2003; accepted Sep- Hawes M, Brigham L, Wen F, Woo H, Zhu Y (1998) Function of root border
tember 9, 2003. cells in plant health: pioneers in the rhizosphere. Annu Rev Phytopathol
36: 311–327
Himanen K, Boucheron E, Vanneste S, de Almeida Engler J, Inzé D,
LITERATURE CITED Beeckman T (2002) Auxin-mediated cell cycle activation during early
lateral root initiation. Plant Cell 14: 2339–2351
Amon A, Tyers M, Futcher B, Nasmyth K (1993) Mechanisms that help the Ito M, Araki S, Matsunaga S, Itoh T, Nishihama R, Machida Y, Doonan
yeast cell cycle clock tick: G2 cyclins transcriptionally activate G2 cyclins JH, Watanabe A (2001) G2/M-phase-specific transcription during the
and repress G1 cyclins. Cell 74: 993–1007 plant cell cycle is mediated by c-Myb-like transcription factors. Plant Cell
Beeckman T, Burssens S, Inzé D (2001) The peri-cell-cycle in Arabidopsis. J 13: 1891–1905
Exp Bot 52: 403–411 Ito M, Iwase M, Kodama H, Lavisse P, Komamine A, Nishihama R,
Beeckman T, Engler G (1994) An easy technique for the clearing of histo- Machida Y, Watanabe A (1998) A novel cis-acting element in promoters
chemically stained plant tissue. Plant Mol Biol Rep 12: 37–42 of plant B-type cyclin genes activates M phase-specific transcription.
Beeckman T, Viane R (2000) Embedding of thin plant specimens for ori- Plant Cell 10: 331–341
ented sectioning. Biotechnol Histochem 75: 23–26 Joubès J, Chevalier C, Dudits D, Heberle-Bors E, Inzé D, Umeda M,
Bell CJ, Ecker JR (1994) Assignment of 30 microsatellite loci to the linkage Renaudin J-P (2000) CDK-related protein kinases in plants. Plant Mol
map of Arabidopsis. Genomics 19: 137–144 Biol 43: 607–620
Brandeis M, Rosewell I, Carrington M, Crompton T, Jacobs MA, Kirk J, Katula KS, Wright KL, Paul H, Surman DR, Nuckolls FJ, Smith JW, Ting
Gannon J, Hunt T (1998) Cyclin B2-null mice develop normally and are JP, Yates J, Cogswell JP (1997) Cyclin-dependent kinase activation and
Plant Physiol. Vol. 133, 2003 1871
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Himanen et al.
S-phase induction of the B1 cyclin gene are linked through the CCAAT Sciortino S, Gurtner A, Manni I, Fontemaggi G, Dey A, Sacchi A, Ozato K,
elements. Cell Growth Diff 8: 811–820 Piaggio G (2001) The cyclin B1 gene is actively transcribed during mitosis
Liu J-q, Seul U, Thompson R (1997) Cloning and characterization of a in HeLa cells. EMBO Rep 2: 1018–1023
pollen-specific cDNA encoding a glutamic-acid-rich protein (GARP) Segal M, Clarke DJ, Reed SI (1998) Clb5-associated kinase activity is
from potato Solanum berthaultii. Plant Mol Biol 33: 291–300 required early in the spindle pathway for correct preanaphase nuclear
Manni I, Mazzaro G, Gurtner A, Mantovani R, Haugwitz U, Krause K, positioning in Saccharomyces cerevisiae. J Cell Biol 143: 135–145
Engeland K, Sacchi A, Soddu S, Piaggio G (2001) NF-Y mediates the Segers G, Gadisseur I, Bergounioux C, de Almeida Engler J, Jacqmard A,
transcriptional inhibition of the cyclin B1, cyclin B2 and cdc25C promoters Van Montagu M, Inzé D (1996) The Arabidopsis cyclin-dependent kinase
upon induced G2 arrest. J Biol Chem 276: 5570–5576 gene cdc2bAt is preferentially expressed during S and G2 phases of the
Menges M, Murray JAH (2002) Synchronous Arabidopsis suspension cul- cell cycle. Plant J 10: 601–612
tures for analysis of cell-cycle gene activity. Plant J 20: 203–212 Serna L, Fenoll C (1997) Tracing the ontogeny of stomatal clusters in
Mews M, Sek FJ, Moore R, Volkmann D, Gunning BES, John PCL (1997) Arabidopsis with molecular markers. Plant J 12: 747–755
Mitotic cyclin distribution during maize cell division: implications for the Shaul O, Mironov V, Burssens S, Van Montagu M, Inzé D (1996) Two
sequence diversity and function of cyclins in plants. Protoplasma 200: Arabidopsis cyclin promoters mediate distinctive transcriptional oscilla-
128–145 tion in synchronized tobacco BY-2 cells. Proc Natl Acad Sci USA 93:
Mews M, Sek FJ, Volkmann D, John PCL (2000) Immunodetection of four 4868–4872
mitotic cyclins and the Cdc2a protein kinase in the maize root: their Soni R, Carmichael J, Shah Z, Murray J (1995) A family of cyclin D
distribution in cell development and dedifferentiation. Protoplasma 212: homologs from plants differentially controlled by growth regulators and
236–249 containing the conserved retinoblastoma protein interaction motif. Plant
Mironov V, De Veylder L, Van Montagu M, Inzé D (1999) Cyclin- Cell 7: 85–103
dependent kinases and cell division in higher plants: the nexus. Plant Cell Stals H, Inzé D (2001) When plant cells decide to divide. Trends Plant Sci
11: 509–521 6: 359–364
Murray AW, Kirschner MW (1989) Cyclin synthesis drives the early em- Tanimoto EY, Rost TL, Comai L (1993) DNA replication-dependent histone
bryonic cell cycle. Nature 339: 275–280 H2A mRNA expression in pea root tips. Plant Physiol 103: 1291–1297
Nurse P (1990) Universal control mechanism regulating onset of M-phase. Thomas JH (1993) Thinking about genetic redundancy. Trends Genet 9:
Nature 344: 503–508 395–398
Peters JL, Constandt H, Neyt P, Cnops G, Zethof J, Zabeau M, Gerats T Tjandra H, Compton J, Kellogg D (1998) Control of mitotic events by the
(2001) A physical amplified-fragment length polymorphism map of Ara- Cdc42 GTPase, the Clb2 cyclin and a member of the PAK kinase family.
bidopsis. Plant Physiol 127: 1579–1589 Curr Biol 8: 991–1000
Planchais S, Perennes C, Glab N, Mironov V, Inzé D, Bergounioux C Tréhin C, Glab N, Perennes C, Planchais S, Bergounioux C (1999) M
(2002) Characterisation of cis-acting element involved in cell cycle phase- phase-specific activation of the Nicotiana sylvestris Cyclin B1 promoter
independent activation of Arath;CycB1;1 transcription and identification involves multiple regulatory elements. Plant J 17: 263–273
of putative regulatory proteins. Plant Mol Biol 50: 109–125 Valvekens D, Van Montagu M, Van Lijsebettens M (1988) Agrobacterium
Porceddu A, Stals H, Reichheld J-P, Segers G, De Veylder L, De Pinho tumefaciens-mediated transformation of Arabidopsis thaliana root explants
Barrôco R, Casteels P, Van Montagu M, Inzé D, Mironov V (2001) A by using kanamycin selection. Proc Natl Acad Sci USA 85: 5536–5540
plant-specific cyclin-dependent kinase is involved in the control of G2/M Vandepoele K, Raes J, De Veylder L, Rouzé P, Rombauts S, Inzé D (2002)
progression in plants. J Biol Chem 276: 36354–36360 Genome-wide analysis of core cell cycle genes in Arabidopsis. Plant Cell
Richard C, Lescot M, Inzé D, De Veylder L (2002) Effect of auxin, cytokinin, 14: 903–916
and sucrose on cell cycle gene expression in Arabidopsis thaliana cell Vos P, Hogers R, Bleeker M, Reijans M, van de Lee T, Hornes M, Frijters
suspension cultures. Plant Tissue Organ Cult 69: 167–176 A, Pot J, Peleman J, Kuiper M et al. (1995) AFLP: a new technique for
Sabatini S, Beis D, Wolkenfelt H, Murfett J, Guilfoyle T, Malamy J, DNA fingerprinting. Nucleic Acids Res 23: 4407–4414
Benfey P, Leyser O, Bechtold N, Weisbeek P et al. (1999) An auxin- Woo H-H, Hawes MC (1997) Cloning of genes whose expression is corre-
dependent distal organizer of pattern and polarity in the Arabidopsis root. lated with mitosis and localized in dividing cells in root caps of Pisum
Cell 99: 463–472 sativum L. Plant Mol Biol 35: 1045–1051
1872 Plant Physiol. Vol. 133, 2003
Downloaded on April 25, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































