Titanium dioxide nanoparticles induce emphysema-like
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The FASEB Journal • FJ Express Full-Length Article
Titanium dioxide nanoparticles induce emphysema-like
lung injury in mice
Huei-Wen Chen,* Sheng-Fang Su,*,† Chiang-Ting Chien,‡ Wei-Hsiang Lin,§
Sung-Liang Yu,† Cheng-Chung Chou,† Jeremy J. W. Chen,†,储,1 and Pan-Chyr Yang†,††,1,2
*Department and Institute of Pharmacology, School of Medicine, National Yang-Ming University,
Taipei, Taiwan; †NTU Center for Genomic Medicine, National Taiwan University, Taipei, Taiwan;
‡
Department of Medical Research, National Taiwan University Hospital, Taipei, Taiwan; §Institute of
Life Sciences, College of Life Sciences, National Chung-Hsing University, Taichung, Taiwan;
储
Institutes of Biomedical Sciences and Molecular Biology, College of Life Sciences, National Chung-
Hsing University, Taichung, Taiwan; and ††Department of Internal Medicine, National Taiwan
University Hospital and National Taiwan University College of Medicine, Taipei, Taiwan
ABSTRACT Titanium dioxide nanoparticles (nano- which may cause pulmonary toxicity (2). Chronic inha-
TiO2) have been widely used as a photocatalyst in air lation studies in rats have shown that nanoparticles can
and water cleaning. However, these nanoparticles inha- induce impaired lung clearance, chronic pulmonary
lation can induce pulmonary toxicity and its mechanism inflammation, pulmonary fibrosis, and lung tumors
is not fully understood. In this study we investigated the (reviewed in ref. 3). Previous studies (reviewed in ref.
pulmonary toxicity of nanoTiO2 and its molecular 4) suggested that attention should be paid to nanopar-
pathogenesis. The adult male ICR mice were exposed ticle-induced toxicity, including the possibility that
to intratracheal single dose of 0.1 or 0.5 mg nanoTiO2 some of the nanoparticles are deposited by diffusional
(19 –21 nm) and lung tissues were collected at 3rd day, mechanisms in all regions of the respiratory tract when
1st wk, and 2nd wk for morphometric, microarray gene inhaled, then may undergo transcytosis across epithe-
expression, and pathway analyses. NanoTiO2 can in- lial and endothelial cells into the blood and lymph
duce pulmonary emphysema, macrophages accumula- circulation, and could induce various biological re-
tion, extensive disruption of alveolar septa, type II sponses such as inflammation and free radical modula-
pneumocyte hyperplasia, and epithelial cell apoptosis. tion. It is therefore important to clarify the effects of
NanoTiO2 induced differential expression of hundreds various nanoparticles on pulmonary health as well as
of genes include activation of pathways involved in cell the pathogenic mechanisms and signaling pathways
cycle, apoptosis, chemokines, and complement cas- involved.
cades. In particular, nanoTiO2 up-regulates placenta Titanium dioxide nanoparticles (nanoTiO2) (⬍100
growth factor (PlGF) and other chemokines (CXCL1, nm) are widely used as photocatalysts in air and water
CXCL5, and CCL3) expressions that may cause pulmo- cleaning (5). The potential pulmonary toxicity is not
nary emphysema and alveolar epithelial cell apoptosis. yet clear. Earlier studies indicated that a single intratra-
Cultured human THP-1 cell-derived macrophages cheal exposure to nanoTiO2 nanoparticles (2 mg per
treated with nanoTiO2 in vitro also resulted in up- rat) is cytotoxic for pulmonary alveolar macrophages
regulations of PlGF, CXCL1, CXCL5, and CCL3. These (6). The toxic effects of TiO2 particles are dose- and
results indicated that nanoTiO2 can induce severe size-dependent. Smaller nanoTiO2 (20 nm) cause a
pulmonary emphysema, which may be caused by acti-
greater pulmonary inflammatory response in rats and
vation of PlGF and related inflammatory pathways.—
mice than larger TiO2 particles (250 nm). The toxicity
Chen, H-W., Su, S-F., Chien, C-T., Lin, W-H., Yu, S-L.,
of nanoTiO2 correlates well with their surface area per
Chou, C-C., Chen, J. J. W., Yang, P. C. Titanium
unit mass (7). Cocultures of human A549 epithelial
dioxide nanoparticles induce emphysema-like lung in-
cells and macrophages (differentiated THP-1 cells)
jury in mice. FASEB J. 20, E1732–E1741 (2006)
show increased sensitivity to nanoparticles and in-
creased cytokine release [interleukin-6 and interleu-
Key Words: nanotechnology 䡠 chemokines 䡠 placenta growth
factor 䡠 microarray 䡠 pulmonary emphysema kin-8 (IL-8)], as compared with mono-cultures of each
cell type (8). However, exposure of THP-1 cell-derived
Nanotechnology, not only is widely used in indus- 1
try, but also has been extensively explored for possible These authors contributed equally to this work.
2
Correspondence: Department of Internal Medicine Na-
applications in medicine. However, the potential toxic- tional Taiwan University Hospital and National Taiwan Uni-
ity issues regarding these powerful nanoparticles are versity College of Medicine, No. 7 Chung-Shan South Rd.,
often ignored (reviewed in ref. 1). Nanoparticles are Taipei 100, Taiwan. E-mail: pcyang@ha.mc.ntu.edu.tw
defined as particles with a diameter less than 100 nm, doi: 10.1096/fj.06-6485fje
E1732 0892-6638/06/0020-1732 © FASEB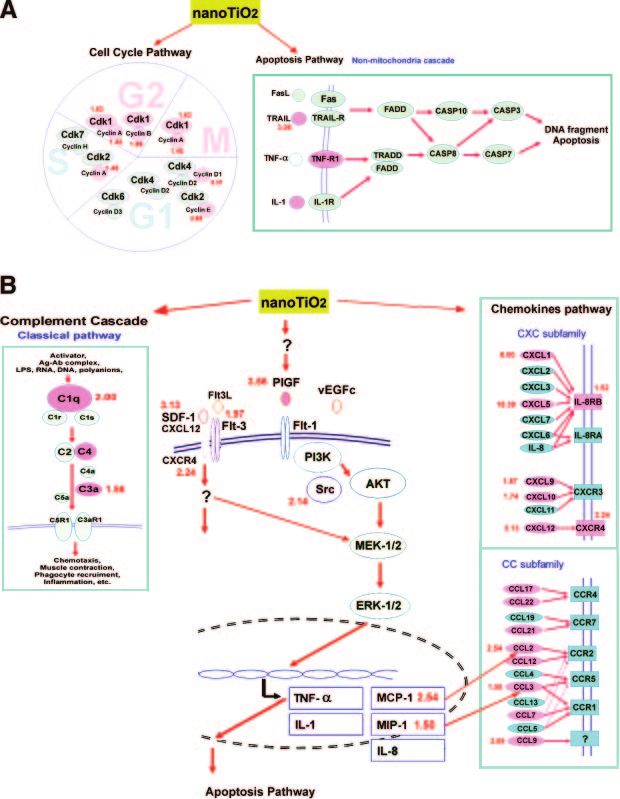
macrophages or endothelial cells to chitosan-DNA three times with PBS, and incubated for another 24 h to
nanoparticles or other nanoparticles (PVC, TiO2, SiO2, eliminate the effect of PMA. The conditioned medium was
Co, Ni) does not induce the release of proinflammatory then used to stimulate fresh THP-1 cells to differentiate into
macrophages (THP-1-derived macrophages), which were
cytokines or have cytotoxic effects (9, 10). grown in normal culture medium for 24 h, then treated with
A host inflammatory or immune response to inhaled or without nanoTiO2 for 24 h.
toxic gases and particles might lead to pulmonary
emphysema and chronic obstructive pulmonary disease Intratracheal instillation of nanoparticles
(COPD) (11), which is a widespread illness with an
increasing prevalence and mortality rate (12). Emphy- The intratracheal instillation procedure was modified from
sema is characterized by an increased number of alve- that in a previous study (16). In brief, after being anesthetized
olar macrophages, neutrophils, and cytotoxic T lym- with 3 to 5% isoflurane in a small chamber, individual mice
phocytes, and the release of inflammatory mediators were secured on an inclined plastic platform and anesthesia
(lipids, chemokines, cytokines, and growth factors) continued via a small nose cone. The trachea was exposed by
a 1 cm incision in the ventral neck skin for instillation of
(13). Overexpression of placenta growth factor (PlGF) normal saline (NS, control) or the nanoTiO2 suspension. The
may contribute to the pathogenesis of pulmonary em- instillation procedure for nanoTiO2 [0.1 mg per mouse (low
physema in transgenic mice (14). Although it is known dose) or 0.5 mg per mouse (high dose) in a 50 l aliquot] was
that nanoTiO2 or other nanoparticles can induce seri- modified from previous reports to ensure that the instilled
ous pulmonary toxicities, the mechanisms and the material was delivered into the lungs of mice with good
molecular pathogenesis are still unclear. distribution (16, 17). The NS group underwent the same
surgical procedure and intratracheal instillation with normal
This study we investigate the effect of nanoTiO2 on
saline.
the induction of pulmonary toxicity and emphysema, The mice recovered and were active within 10 min after
its mechanisms, and the molecular pathogenesis. removal of the inhalation anesthetic. The incision healed
within two days, and then the animals were observed daily
until their scheduled termination, including 3 days for hyper-
acute response, 1 wk for acute-phase, and 2 wk for chronic
MATERIALS AND METHODS phase. After instillation for 3 days, 1 wk, or 2 wk, the mouse
was injected intraperitoneally (i.p.) with a lethal dose (0.1 ml)
Animals and nano materials of pentobarbital sodium solution (Nembutal, Abbott, North
Chicago, IL, USA) and the three right lobes of the lung
Adult male ICR mice (2 months old, 30 g; Harlan Sprague- tissues were collected and frozen in liquid nitrogen for RNA
Dawley, Indianapolis, IN, USA), free of known rodent patho- or protein extraction. For the histological study, the lung
gens, were obtained from the National Taiwan University tissues were inflated with air at constant pressure (25 cmH2O)
(Taipei, Taiwan). They were cared and used humanely ac- and then fixed with 10% buffered formalin by tracheal
cording to the Guide for the Care and Use of Laboratory instillation.
Animals as adopted and promulgated by the National Health
Research Institutes (NHRI, Taiwan). This study was approved Morphometric analysis of mice lungs after nano
by the Institutional Review Board and the Animal Care and TiO2 exposure
Use Committee of the National Chung-Hsing University
(Taichung, Taiwan). Morphometric measurements mice lungs were performed by
NanoTiO2 (Rutile crystal phase, ultra-fine TiO2 nanopar- an investigator who was unaware of the identity of the
ticles), a highly dispersed and hydrophilic fumed TiO2 with a samples. The mice lung sections (5 m) were prepared and
diameter of 19 –21 nm (average primary particle size 21 nm), viewed with a 20⫻ objective and the images digitized, con-
a specific surface area of 50 ⫾ 15 m2/g, and a purity ⱖ 99.5%, verted to tagged image format file, and analyzed using
purchased from Degussa (Frankfurt, Germany). NanoTiO2 MetaMorph® Imaging System software (Universal Imaging
readily aggregate to form microparticles in normal saline or Corp., Downingtown, PA, USA). To evaluate the pathological
culture medium. To avoid aggregation, the nanoTiO2 suspen- changes following treatment, enlarged alveoli, disrupted
sion was ultrasonicated before it was used to treat animals or septa, and thickened epithelia were measured as in previous
cells. Each sample was vortexed just before an aliquot was studies (14, 18, 19). Three parameters were measured in each
drawn for instillation. TiO2 microparticles have a diameter of section. The airspace area was measured and compared
180 –250 nm and a specific surface area of 6.5 m2/g (Fisher between the test groups and the NS group. A second param-
Scientific, Springfield, NJ, USA). eter, which we call septal chord length, was measured as an
indication of the thickness of the septa; this is identical to the
Cell lines parameter called airspace wall thickness in an earlier report
(18). The third parameter, the mean linear intercept (MLI),
was used as a measure of the interalveolar wall distance (19).
The human monocyte cell line THP-1 (American Type Cul-
Five areas selected randomly of each section slide were
ture Collection TIB 202; ATCC, Manassas, VA, USA) and the
counted and six sections were examined in each animal.
human lung carcinoma A549 cells (American Type Culture
Collection CCL-185) were grown in normal culture medium
[RPMI 1640 medium (GIBCO-Life Technologies, Inc., Gaith- Apoptosis assay by terminal deoxynucleotidyltransferase-
ersburg, MD, USA), supplemented with 1.5 g/liter of mediated dUTP nick-end labeling (TUNEL)
Na2HCO3, 4.5 g/liter of glucose (Glc), and 10% FBS (FBS;
GIBCO-Life Technologies)] (15). To collect conditioned Apoptotic cells in lung tissue sections were detected with the
medium to stimulate other THP-1 cells, THP-1 cells were ApopTag in situ apoptosis detection kit (Roche Diagnostics,
treated for 24 h with 3.2 ⫻ 10⫺7 M phorbol myristate acetate Mannheim, Germany) as in our previous study (20) accord-
(PMA; Sigma Chemical Co., St. Louis, MO, USA), washed ing to the protocol provided by the manufacturer. The cells
NANOTIO2-INDUCED EMPHYSEMA E1733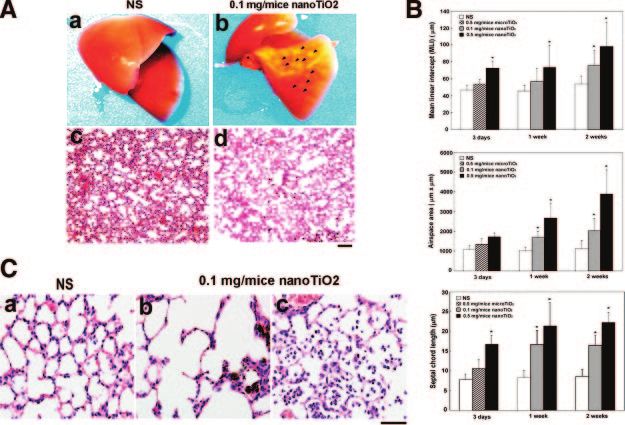
were counterstained with methyl green and those that were biochemical pathway database using the genes selected from
intact and exhibited dark brown-stained nuclei (TUNEL- cDNA microarray analysis as described previously (25).
positive cells) were considered positive for apoptosis (21).
The number of positively stained cells was measured in five Real-time quantitative RT-polymerase chain reaction
randomly selected high-power fields (⫻400) per slide, and (RT-PCR)
the significance of differences between the nanoTiO2-treated
and NS groups was examined using Student’s t test. To validate the microarray data, real-time quantitative RT-
PCR was used in a 96-well format according to our previous
Immunohistochemical staining described method (26). Total RNA from lung tissues or cell
cultures with or without nanoTiO2 treatment was prepared.
Immunohistochemistry was performed as in previous reports Primers were designed using Primer Express v2.0 Software
(20, 22). Lung tissue sections were incubated with a 1:100 (Applied Biosystems Inc., Foster City, CA, USA) (Table 1). All
dilution of rabbit polyclonal anti-F4/80 antibody (Ab) reactions were carried in 50 l volumes containing 25 l of
(marker for mouse macrophages) or anti-PCNA Ab (marker SYBR Green PCR Master Mix (Applied Biosystems Inc., Foster
for proliferative cells) (both from Santa Cruz Biotechnology, City, CA, USA). The amount of test gene cDNA relative to the
Santa Cruz, CA, USA) or goat polyclonal anti-PlGF2 Ab (R&D amount of glyceraldehyde-3-phosphate dehydrogenase
Systems, Minneapolis, MN, USA). The slides were then incu- (GAPDH) cDNA (mouse tissues) or TATA box binding
bated with biotinylated secondary Ab and peroxidase-labeled protein (TBP) cDNA (human cells) (housekeeping controls)
streptavidin (avidin-biotin complex kit; Vector Laboratories, was measured as –⌬ computed tomography (CT) ⫽ – [com-
Burlingame, CA, USA). Negative control slides that were not puted tomographyTested gene - CTGAPDH or TBP]. The ratio of
treated with primary Ab were included for each staining the tested gene mRNA copies relative to those for GAPDH or
procedure. Finally, 3,3⬘-diamino-benzidine (DAB) was used to TBP was defined as 2 – ⌬ CT ⫻ K (K: constant).
develop the signals (brown color), while methyl green or
hematoxylin was used for counterstaining. The observers Western blotting
randomly selected five areas and counted positive cells
(brown staining) on a ⫻200 field (i.e., ⫻20 objective lens and The detailed protocol has been previously reported (23).
⫻10 ocular lens, 1.227 mm2 per field). All counts were PlGF expression was detected using anti-mouse PlGF2 poly-
performed by two investigators blinded to the type of sample clonal antibody (pAb) (1:1,000 dilution; R&D Systems) or
and both had to agree as to what constituted a positive cell anti-human PlGF pAb (1:500 dilution; Santa Cruz Biotechnol-
before any cell was included in the count. ogy). Western blot data was analyzed by densitometry (Image-
master V-DS; Amersham Biosciences, Piscataway, NJ, USA).
cDNA microarray analysis
Quantification of cytokine levels
The detailed protocol for the mouse cDNA microarray anal-
Serum from mice with or without nanoTiO2 treatment or cell
ysis has been reported in our previous studies (20, 23). Mouse
culture medium from A549 or THP-1-derived macrophages
expressed sequence tag (EST) clones were obtained from the
with or without nanoTiO2 treatment was collected and stored
IMAGE consortium libraries through its distributor (ResGen
at – 80°C until analysis. Levels of PlGF, MCP-1, and MIP-1
Invitrogen, Huntsville, AL, USA) (24). The cDNA microarray
were measured using ELISA assay (R&D Systems) as de-
carrying 6,144 polymerase chain reaction (PCR) -amplified
scribed (14).
cDNA fragments was prepared using an arraying machine.
Potential interindividual variability was minimized by pooling
the mRNA samples from two mice from each group to yield a Statistical analysis
representative sample for analysis. Total RNA was extracted
from the pooled lung tissues for each group using RNAzolTM Detailed descriptions and an excellent discussion of the issues
B solution (Life Tech, Gaithersburg, MD, USA) and mRNAs involved in generating the microarray data, data normaliza-
were extracted using an mRNA isolation kit (Qiagen, Hilden, tion, statistical analysis, and its interpretation are given in our
Germany), in accordance with the manufacturer’s protocol. previous studies (20, 27). Genes which up-regulated or down-
Five micrograms of mRNA from each sample was used in each regulated in response to nanoTiO2 treatment were identified
array. The microarray images were scanned, digitized, and and were used for pathway analysis. An up-regulated gene had
analyzed using a flatbed scanner (PowerLook 3000; UMAX, to show a 1.5-fold increase in the cDNA microarray. These
Taipei, Taiwan) and GenePix 3.0 software (Axon Instru- genes were further analyzed by our in-house data mining tool
ments, Union City, CA, USA). In designing experiments based on KEGG and BIOCARTA pathway databases
involving microarrays, we adhered to the guidelines of the (http://biochip.nchu.edu.tw/SpecificDB/mouse.html).
Microarray Gene Expression Data (MGED) Society (www. ANOVA (Excel, Microsoft, Taipei, Taiwan) or Student’s t
mged.org/Workgroups/MIAME/miame_checklist.html). test was used to determine if significant differences were seen
in replicate experiments between the NS and nanoTiO2-
treated groups for the numbers of F4-positive, PCNA-positive,
Identification of pathways using the KEGG and or TUNEL-positive cells in lung tissue sections and for PlGF
BioCata database expression on Western blots or by ELISA analysis.
Gene identification was performed to determine which
biochemical pathways were altered during the nanoTiO2- RESULTS
induced pulmonary inflammatory response. Having identi-
fied genes on the basis of the cDNA microarray data, it was
of interest to determine whether any of these genes were NanoTiO2 can induce emphysema-like lung injury
part of the same pathway. The approach taken was to in mice
search the Kyoto Encyclopedia of Genes and Genomes
(KEGG) (http://www.genome.ad.jp/kegg/pathway.html) and the We found that 1 wk after single intratracheal instillation
BIOCARTA (http://www.biocarta.com/genes/allpathways.asp) with nanoTiO2 in mice, the lungs showed significant
E1734 Vol. 20 November 2006 The FASEB Journal CHEN ET AL.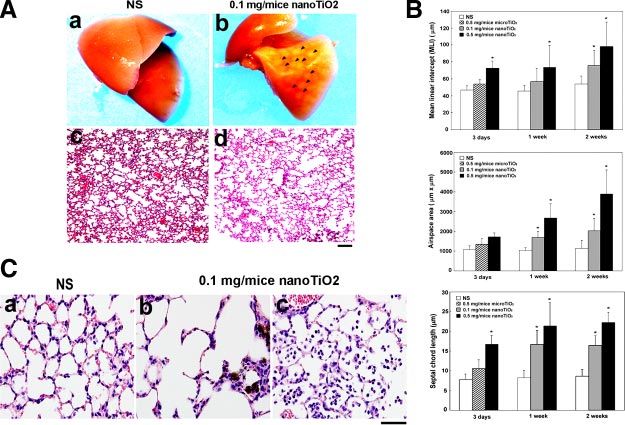
TABLE 1. NanoTiO2-induced genes detected using the “homemade” cDNA microarray (n⫽3)
cDNA microarray
Public ID Symbol (nanoTiO2/N) Categories
NM_007659 Cdc2a 1.63 Cell cycle/cell growth regulator
AU015121 Ccnb1 1.89 Cell cycle/cell growth regulator
NM_007631 Ccnd1 3.31 Cell cycle/cell growth regulator
NM_007633 Ccne1 2.62 Cell cycle/cell growth regulator
BC037601 Fgf1 2.03 Cell cycle/angiogenesis
BC010200 Fgfr1 2.14 Cell cycle/angiogenesis
NM_008827 Plgfa 3.56 VEGF signaling pathway
AF470537 Traila 3.06 Chemokine/apoptosis
NM_010229 Flt3a 1.97 Chemokine
AF065933 Ccl2 2.54 Chemokine/chemotaxis
NM_011337 Ccl3a 1.50 Chemokine/chemotaxis
AF128196 Ccl9 2.89 Chemokine/chemotaxis
NM_008176 Cxcl1a 2.50 Chemokine/chemotaxis
NM_009141 Cxcl5a 3.12 Chemokine/chemotaxis
NM_009911 Cxcr4 2.24 Chemokine/chemotaxis
U05264 Gp49b 3.92 Chemokine/chemotaxis
BB333624 C3ar1 1.56 Complement activation/chemotaxis
NM_008198 H2-Bf 2.68 Complement activation/chemotaxis
NM_008127 Gjb4 2.12 Gap junction proteins-connexins
NM_008965 Ptger4 (EP4)a 2.09 GPCRs class A rhodopsin-like/TGF- signaling pathway
NM_030701 Gpr109b 2.01 GPCRs class A rhodopsin-like/TGF- signaling pathway
BC057952 Mmp15 1.98 Matrix metalloproteinases
BC070430 Mmp2 2.00 Matrix metalloproteinases
a
Confirmed by real-time quantitative RT-PCR. Abbreviations: Cdc2a, cell division cycle 2 homolog A (S. pombe); Ccnb1, cyclin B1; Ccnb2,
cyclin B2; Ccnd1, cyclin D1; Ccne1, cyclin E1; Fgf1, fibroblast growth factor-1; Fgfr1, fibroblast growth factor receptor type 1; Trail, TNF␣-related
apoptosis-inducing ligand (TRAIL); Flt3, MS-like tyrosine kinase 3; Plgf, placental growth factor; Ccl2, chemokine (C-C motif) ligand 2,
monocyte chemoattractant protein-1 (MCP-1); Ccl3, chemokine (C-C motif) ligand 3, macrophage inflammatory protein-1alfa (MIP-1␣); Ccl9,
chemokine (C-C motif) ligand 9, macrophage inflammatory protein-1gamma (MIP-1␥); Cxcl1, chemokine (C-X-C motif) ligand 1, growth-
related oncogene-alpha (GRO-␣); Cxcl5, chemokine (C-X-C motif) ligand 5, epithelial cell-derived neutrophil-activating peptide-78 (ENA-78);
Cxcr4, chemokine (C-X-C motif) receptor 4; Gp49b, glycoprotein 49 B; C3ar1 complement component 3a receptor 1; H2-Bf, histocompatibility
2, complement component factor B; Gjb4, gap junction membrane channel protein beta 4; Ptger4 prostaglandin E receptor 4 (subtype EP4);
Gpr109b G-protein-coupled receptor 109B; Mmp15, matrix metalloproteinase 15; Mmp2, matrix metalloproteinase 2.
changes in morphology and histology (Fig. 1A), matous change), type II pneumocyte proliferation,
whereas no obvious morphological changes were seen increased alveolar epithelial thickness, and accumula-
in the NS-treated control group (Fig. 1A). Disruption of tion of particle-laden macrophages (Fig. 1B) were
the alveolar septa and alveolar enlargement (emphyse- observed in the low dose (0.1 mg/mouse) nanoTiO2-
Figure 1. NanoTiO2-induced pulmonary mor-
phological and histological changes. A) Morpho-
logical (a, b) and histological (c, d) changes (H/E,
hematoxylin and eosin staining) in the mouse
lung at 1 wk after intratracheal instillation with NS
(normal saline, a, c), or 0.1 mg/mouse nanoTiO2
(b, d). Arrowheads indicate the nodule-like le-
sions caused by chronic inflammation. Original
magnification ⫻100, bar ⫽ 100 m for H/E
histological image (c, d). B) Histological changes
in the mouse lung after intratracheal instillation
with nanoTiO2 for 1 wk. Lung tissues were col-
lected from NS-treated control mice (a) and
nanoTiO2 (0.1 mg/mouse) -treated mice (b, c).
Original magnification, ⫻400, bar ⫽ 50 m.
Similar results were obtained in six dependent ex-
periments. C) Morphometric measurements from
lungs at 3 days, 1 wk, and 2 wk after installation with
NS or nanoTiO2 (0.1 or 0.5 mg/mouse). The
mean linear intercept (MLI), mean airspace area,
and septal chord length were measured (n⫽6) as
described in Materials and Methods. The data are
the means ⫾sd *P ⬍ 0.05 in Student’s t test
compared with the control (NS) group.
NANOTIO2-INDUCED EMPHYSEMA E1735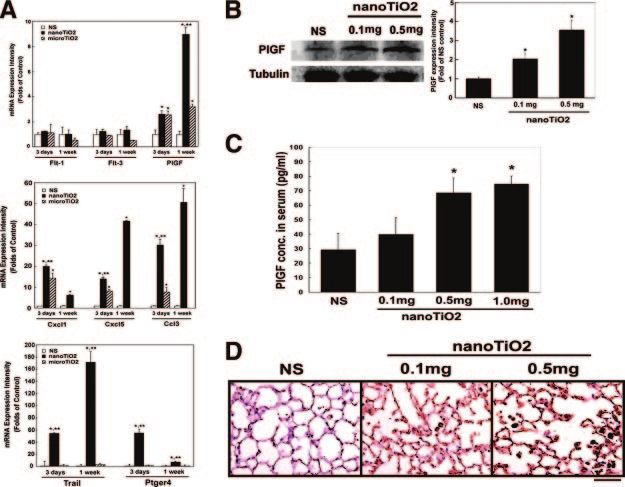
treated group and were more severe in mouse lung
treated with higher dose of NanoTiO2 (0.5 mg/
mouse). These pathological changes diffusely involved
the entire both lungs and were considerably more
severe in areas in which nanoTiO2 accumulated. The
NS control group showed no significant morphological
or histological changes.
Serial morphometric measurements of injured mice
lungs were made at 3rd day, 1st wk, and 2nd wk after
intratracheal installation of nanoTiO2 (Fig. 1C). The
mean linear intercept (MLI) (a measure of the interal-
veolar wall distance), the airspace area, and the septal
chord length (a parameter that increases with septal
thickness) were usually used as the pathological mark-
ers of pulmonary emphysema and granuloma. All three
parameters were slightly increased at 3rd day (hyper-
acute phase) and significantly increased at 1st wk
(acute-phase) after instillation of nanoTiO2, and the
pathological changes persisted until 2nd wk (chronic
phase). The nanoTiO2 therefore can induce time- and Figure 2. Effects of nanoTiO2 on macrophage accumulation,
cell proliferation, and apoptosis. The left panels show typical
dose-dependent pulmonary emphysema-like changes results, the right panels the quantified results expressed as the
in mice. mean⫾sd (n⫽6); *P ⬍ 0.05 vs. controls (NS). A) Immuno-
staining with mouse macrophage-specific anti-F4 Ab. The red
arrows show the brown colored DAB F4/80-positive cells,
Alveolar macrophage infiltration and pulmonary cell
Original magnification ⫻400, bar ⫽ 50 m. B) Immunostain-
apoptosis and proliferation ing with Ab against PCNA (proliferative cell nuclear antigen)
for proliferating cells. The red arrows indicate PCNA-positive
The nanoTiO2 treated mouse lungs showed significant cells. Original magnification ⫻ 400, bar ⫽ 50 m. C) TUNEL
staining for apoptotic cells. The red arrows indicate apoptotic
increase in alveolar macrophage infiltration, alveolar macrophages, the blue arrows apoptotic epithelial cells. Orig-
epithelial septal thickness, and alveolar enlargement. inal magnification ⫻ 400, bar ⫽ 50 m.
Figure 2A shows that the number of alveolar macro-
phage was significantly increased at wk 1 in the group
treated with 0.1 mg of nanoTiO2 (4.16-fold increase), mice, 318 genes being up-regulated and 188 genes
indicating that a severe inflammatory response oc- down-regulated (Supplemental data, Fig. S1).
curred. These infiltrated particle-laden macrophages Some of these nanoTiO2-regulated genes are listed
accumulated in the alveolus and could be identified by and categorized by their putative functions (Table 1).
mouse macrophage-specific anti-F4/80 receptor anti- Several vascular endothelial growth factor (VEGF) -re-
bodies. Using PCNA as the marker for cell prolifera- lated factors, G protein-coupled receptors (GPCR), cell
tion, nanoTiO2-stimulated abnormal proliferation of growth regulators, and chemotaxis and immune re-
type II pneumocytes could be identified in pulmonary sponse factors were significantly up-regulated by nano-
tissues (Fig. 2B). TiO2-treatment. These gene expression changes re-
TUNEL staining showed that the number of apopto- flect, at the molecular level, the observed nanoTiO2-
tic cells was significantly increased in the nanoTiO2- induced inflammatory response.
treated group; these TUNEL-positive cells were both
macrophages and alveolar type II pneumocytes (Fig. Pathway analysis of nanoTiO2-induced transcriptomic
2C). The nanoparticle-induced alveolar epithelial cell changes
apoptosis might cause abnormal airspace enlargement,
which is a major pathological change in pulmonary The differentially expressed profiles of nanoTiO2-
emphysema. induced genes were categorized and integrated to fit
the transduction signaling map using the KEGG and
BIOCARTA pathway database. This provided us with
Gene expression profiles of the mice lung after new information by giving a biological interpretation of
nanoTiO2 treatment the voluminous data generated by microarray experi-
ments. According to the pathway analysis, four major
Messenger RNAs from mice lung tissues with or without pathways were up-regulated by nanoTiO2: the cell cycle
1 wk of nanoTiO2 treatment (0, 0.1, or 0.5 mg per regulatory pathway and apoptosis pathway (Fig. 3A)
mouse) were analyzed using the mouse cDNA microar- and the chemokines pathway and complement cascade
ray (19, 24). There were 506 genes out of the 6,144 (classical pathway) (Fig. 3B).
putative genes showed a statistically significant differ- As shown in Fig. 3A, the cell cycle pathway analysis
ence (1.5-fold difference) in expression at wk 1 in showed that nanoTiO2 regulated key factors for G2/M
nanoTiO2-treated mice compared with NS-treated progression by increasing the expression of cdc2a,
E1736 Vol. 20 November 2006 The FASEB Journal CHEN ET AL.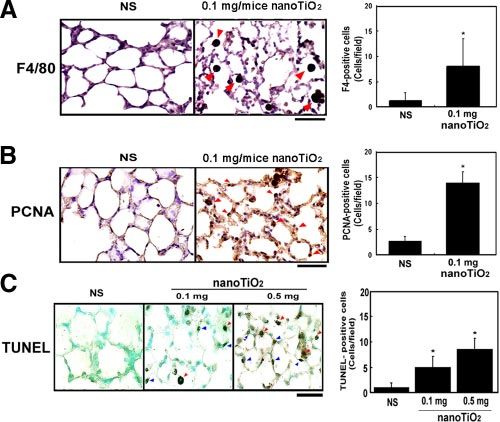
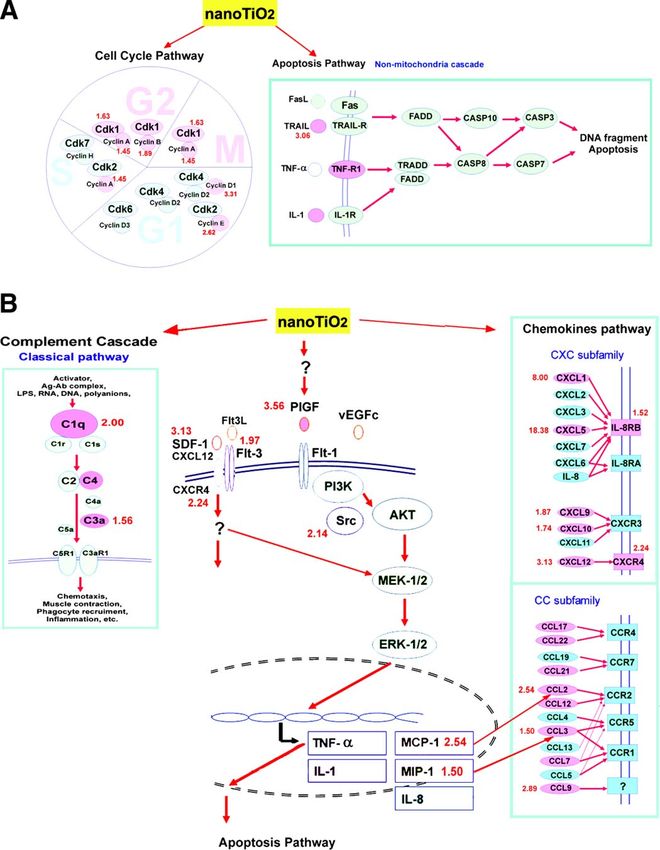
Figure 3. NanoTiO2-induced genes in different pathways according to the KEGG pathway database and BIOCARTA. The pink color indicates nanoTiO2-induced genes, the red numbers close to the genes the fold increase. A) The cell cycle and apoptosis pathways. B) The PlGF/chemokines pathway and the classic complement pathway. The hypothetical nanoTiO2-regulated signaling pathways were modified from the KEGG and BIOCARTA database, while the PlGF pathway was modified from Selvaraj SK, 2003 (28). NANOTIO2-INDUCED EMPHYSEMA E1737
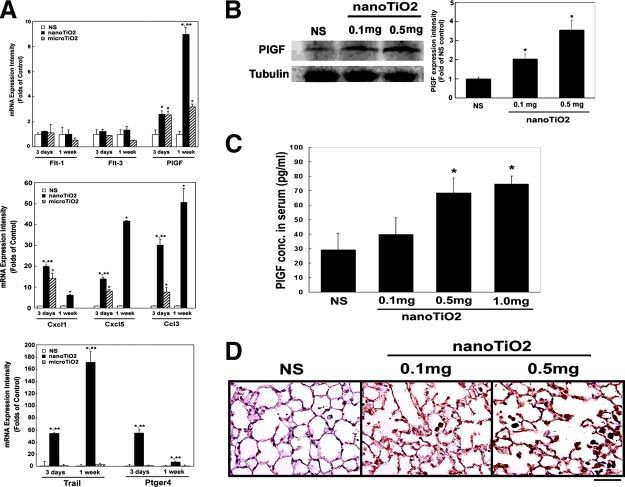
Figure 4. PlGF, chemokines and related factors
expression in mice after single intratracheal
instillation with nanoTiO2. A) Real-time quan-
titative RT-PCR for flt-1 (PlGF receptor), flt-3,
plgf, chemokines (cxcl1, cxcl5, ccl3), and apo-
ptosis-related factors (trail and ptger4). The
data are expressed as the fold increase com-
pared with the NS control ⫾sd *P ⬍ 0.05 vs. the
NS control; **P ⬍ 0.05 vs. microTiO2 (n⫽4). B)
Western blotting for the effect of nanoTiO2 on
PlGF protein expression. The right panel shows
the expression of PlGF relative to that for
␣-tubulin expressed as a fold increase com-
pared with the NS control ⫾sd *P ⬍ 0.05 vs. NS
control (n⫽4). C) Serum PlGF protein levels
measured by ELISA. The data are expressed as
the mean ⫾sd. *P ⬍ 0.05 vs. controls (n⫽4). D)
Immunohistochemical staining with anti-PlGF2
Ab showing overexpression of PlGF (brown
color, DAB staining) in mice lung tissues after
intratracheal instillation with nanoTiO2 (0.1
mg and 0.5 mg per mice). Similar results were
obtained in 4 experiments.
cyclin A2 (2.30-fold), cyclin B1 and B2, cyclin D1, cyclin levels of PlGF protein (Fig. 4C), which might be pro-
E1 while the apoptosis pathway analysis showed they duced mainly by infiltrating macrophages and some
increased expression of tumor necrosis factor-related pulmonary epithelial cells, as shown by immunostain-
apoptosis-inducing ligand (TRAIL) and tumor necrosis ing with PlGF-specific antibodies (Fig. 4D).
factor-receptor1 (TNF-R1) expression, respectively. As
shown in Fig. 3B, nanoTiO2 stimulated the expression
of several cytokines and chemokines, including PlGF PlGF and other chemokines expression in
(3.56-fold), a prochemokine known to regulate the macrophagse and lung epithelial cells exposed to
expression of MCP-1, IL-1, and TNF-␣ (45, 46), as seen nanoTiO2 in vitro
in our system. Other C-C and C-X-C chemokines were
also up-regulated (Ccl2, Ccl3, Ccl9, Cxcl1, Cxcl5, To study the responses of different human pulmonary
Cxcr4, and Gp49b). cells to nanoTiO2, THP-1-derived macrophages and the
lung epithelial cell line, A549, were used as in vitro models.
PlGF and chemokines in nanoTiO2-treated mice Incubation of cultured cells with nanoTiO2 showed that
lung tissues they caused significant dose-dependent induction of PlGF
expression at both the mRNA and protein levels in
To confirm the role of the PlGF/chemokine pathway in THP-1-derived macrophages, as shown by Western blot-
nanoTiO2-induced pulmonary injury, real-time quanti-
ting (Fig. 5A), real-time quantitative RT-PCR (Fig. 5B),
tative RT-PCR, Western blotting, ELISA, and immuno-
and ELISA (Fig. 5C), but had less effect on human
staining were used to demonstrate PlGF and related
A549 pulmonary epithelial cells (Fig. 5B).
cytokines induction in nanoTiO2-treated mice.
To examine whether these effects on PlGF/chemo-
Real-time quantitative RT-PCR analysis showed that
plgf, chemokines (cxcl1, cxcl5, and ccl3), TRAIL, and kines induction pathways were specific to nanoTiO2,
prostaglandin E receptor 4 (ptger4, EP4) were signifi- the macrophages THP-1 and lung epithelial cells A549
cantly up-regulated in the lung tissues of mice treated were treated with nanoTiO2 or microTiO2. The results
with nanoTiO2 for 1 wk, while expressing of flt-1 and showed that nanoTiO2 caused significant induction of
flt-3 (PlGF receptors), were not significantly affected PlGF expression in macrophages and lung epithelial
(Fig. 4A), the results are comparable to those obtained cells at the protein level (Fig. 5A) and mRNA level (Fig.
in the microarray and pathway analyses (Fig. 3). Fine 5B) in cells and increased levels of secreted protein
TiO2 microparticles (microTiO2) had less effect on (Fig. 5C), whereas microTiO2 had no, or only a slight,
induction of these genes (Fig. 4A). effect. Figure 5D, E shows that cxcl5 mRNA levels were
The nanoTiO2-induced PlGF expression was also increased by nanoTiO2 in both types of the cells, while
examined at the protein level after single intratracheal microTiO2 had little effect. To study the downstream
instillation with nanoTiO2 (0.1 mg or 0.5 mg per mice) effectors of PlGF, we examined protein levels of MCP-1
for 1 wk. Western blotting showed that nanoTiO2 (CCL2) in the culture medium and found that these
caused significant induction of PlGF expression in a chemokines were increased by nanoTiO2 in THP-1
dose-dependent manner (Fig. 4B), while ELISA analysis cell-derived macrophages, but not in A549 cells, and
showed that nanoTiO2-treated mice had higher serum that microTiO2 had no effect (Fig. 5F).
E1738 Vol. 20 November 2006 The FASEB Journal CHEN ET AL.Figure 5. A) Western blotting of PlGF protein
expression in human THP-1 cell-derived mac-
rophages treated for 24 h with nanoTiO2 (0.1,
0.2, 0.5 g/ml) or microTiO2 (0.2 g/ml). The
right panel shows the expression of PlGF rela-
tive to that for ␣-tubulin expressed as a fold
increase compared with the controls (n⫽3).
The data are expressed as the mean⫾sd *, P ⬍
0.05 vs. controls. B) Real-time quantitative RT-
PCR analysis showing effects of nanoTiO2 or
microTiO2 on PlGF expression in THP-1 cell-
derived macrophages and A549 cells. C) ELISA
measurements of PlGF levels in THP-1 cell-
derived macrophage or A549 cells culture me-
dium following nanoTiO2 or microTiO2 treat-
ment. (D, E) Real-time quantitative RT-PCR
analysis for cxcl5 mRNA in THP-1 cell-derived
macrophages (D) and A549 cells (E). F) Expres-
sion of MCP-1 (ccl2) in THP-1 cell-derived
macrophages and A549 cells following nano-
TiO2 or microTiO2 treatment. The data are the
mean ⫾sd (n⫽3).
DISCUSSION increase in septal thickness seen in this study. The activa-
tion of apoptosis pathway indicate that nanoTiO2 can
The results of this study indicate that single intratracheal increase TRAIL expression, which may account for the
instillation of 0.1 mg nanoTiO2 can induce severe pulmo- increased number of TUNEL-positive cells in nanoTiO2-
nary inflammation and emphysema in the mouse lung. treated samples, explaining the alveolar type II cell apo-
The finding of pulmonary inflammation is consistent with ptosis, abnormal airspace enlargement, and pulmonary
another report showing that nanoparticle inhalation can emphysema.
induce pulmonary inflammation (2), However, the obser- The pathway analysis also shows that nanoTiO2 can
vation that nanoTiO2 can induce pulmonary emphysema stimulate the expression of several cytokines and chemo-
after single intratracheal expose of 0.1 mg nanoTiO2 is kines, including PlGF, a prochemokine that can regulate
novel. Our results indicate that the pulmonary emphy- the expression of MCP-1, IL-1, and TNF-␣ (13, 45, 46).
sema is triggered by nanoTiO2 activation of macrophages,
These chemokines may also affect the expression of other
up-regulations of PlGF and other inflammatory cytokines
C-C and C-X-C chemokines (Ccl3, Cxcl1, and Cxcl5) that
that resulted in disruption of alveolar septa, alveolar
modulate chemotaxis, neutrophil infiltration, macro-
epithelial injury, type II cell proliferation and apoptosis.
phage accumulation, epithelial cell proliferation, and
This information may have important clinical implica-
tions regarding the safety issue, as nanoTiO2 are widely apoptosis to generate the inflammatory cascade, which,
used as a photocatalyst in air and water cleaning (4) and may lead to the pathogenesis of pulmonary emphysema.
TiO2 is used as a pigment in the paint industry. Extra The nanoTiO2-induced expression of many cytokines
caution should therefore be taken in the handling of and chemokines (Fig. 3) may play an important role in
higher dose nanoTiO2. the macrophage accumulation, neutrophil infiltration,
Pulmonary toxicity caused by nanoTiO2 inhalation has cell apoptosis, lung destruction, and pulmonary emphy-
been reported (5, 6), but its molecular pathogenesis is not sema seen in this study. The induction of chemokines
known. In this study, the microarray gene expressions and by nanoTiO2 has some similarities with the gene ex-
pathway analysis indicated that the cell cycle, apoptosis, pression profiles seen in a lipopolysaccharide (LPS)
chemokine, and complement pathways may be involved (lipopolysaccharide) -induced acute lung injury model
in nanoTiO2-induced pulmonary toxicity (Fig. 3). The (28). CCL22 (macrophage-derived chemokine, MDC),
activation of cell cycle pathway suggests that nanoTiO2 CCL3 (MIP-1 alpha), CCL2 (MCP-1 alpha), CXCL2/3
can regulate key factors for G2/M progression by increas- (MIP-2), and CXCL1 (keratinocyte cell-derived chemo-
ing the expression of cdc2a, cyclin A2, and cyclin B1, kine, KC) were induced in both lung injury models.
which may explain the increase in the number of prolif- Among these factors, expression of MIP-1 alpha and
erating (PCNA-positive cells) type II pneumocytes and the MIP-2, has been shown to be induced by TiO2 in A549
NANOTIO2-INDUCED EMPHYSEMA E1739epithelial cells and macrophages in previous studies studies will be done to confirm the central role of PlGF
(29, 30). In our study, CXCL1, CXCL5, and CCL3 were in nanoTiO2-induced pulmonary inflammation and
significantly induced and this was confirmed in in vivo emphysema via using PlGF knockout mice.
and/or in vitro studies. CXCL1 (KC) is a potent neu- In this study, we found that a single intratracheal
trophil chemoattractant involved in several lung injury exposure to nanoTiO2 could induce pulmonary emphy-
processes, and its up-regulation has been correlated sema and severe lung injury in mice. However, no signif-
with neutrophil infiltration and the development of icantly pathological changes were seen using the same
granulomas (31). CXCL5 (epithelial cell-derived neu- dose of microTiO2 (180 –250 nm). This finding is consis-
trophil-activating peptide-78, ENA-78) is also a neutro- tent with other reports that nanoTiO2 (20 nm) cause a
phil chemoattractant involved in pulmonary inflamma- significantly greater pulmonary inflammatory response
tion (32), while CCL3 (MIP-1 alpha) is an important than microTiO2 (250 nm) in rats and mice (6, 46). The
chemokine involved in pulmonary host defense during greater toxicity of nanoTiO2 might correlate with their
infections (33, 34). Several of these factors were also greater surface area per unit mass. A significantly greater
found to be increased in patients with emphysema and increase in PlGF was induced by nanoTiO2 than micro-
␣1-antitrysin deficiency-related emphysema in a mi- TiO2 in this study, suggesting that nanoTiO2-induced
croarray study (35). These key chemokines might play pulmonary toxicity may be mediated by PlGF.
important roles in nanoTiO2-induced inflammatory The results of this study add our understanding of
responses and could also be involved in the pathogen- nanoTiO2-induced pulmonary toxicity and pulmonary
esis of pulmonary granuloma and emphysema. emphysema. Both are complicated multifactorial dis-
NanoTiO2 caused increased expression of the classi- ease processes. We suggest that PlGF, chemokines, and
cal complement pathway components, C1q, C3a, and the complement cascade may cause inflammatory cell
C4, which could lead to complement activation, includ- chemotaxis, cell proliferation and apoptosis, resulting
ing C5 and its receptor, C5R1, leading to more phago- in serious lung injury. Further investigations are
cyte recruitment and chemotaxis and an inflammatory needed to elucidate the potential pulmonary toxicity of
response in the lungs (36, 37) and the destruction of different nanoparticles and their pathogenesis.
pulmonary tissues. A recent study demonstrated that C5
and the C5a receptor are involved in the mycobacterial The authors thank Drs. Gene Alzona Nisperos, W. K. Chan,
and Tom Barkas for their excellent editing. This investigation
glycolipid trehalose 6,6⬘-dimycolate-induced pulmo- was supported by grants from the National Science Council,
nary granulomatous response (38). These findings sug- Taiwan (NSC94 –2314-B-005– 004 and NSC95–2314-B-005– 003).
gest that the classical complement pathway might be
involved in nanoTiO2-induced lung injury.
In this study, nanoTiO2 increased mRNA and protein
levels of PlGF (a chemokine inducer) both in vivo REFERENCES
(mice) and in vitro (human THP-1 cell-derived macro- 1. Moghimi, S. M., Hunter, A. C., and Murray, J. C. (2005)
phages) and up-regulated factors downstream of PlGF Nanomedicine: current status and future prospects. FASEB J. 19,
[MIP-1 (CCL3), MCP-1 (CCL2), and IL-1] in a concen- 311–330
tration-dependent manner. PlGF has been highlighted 2. Borm, P. J., and Kreyling, W. (2004) Toxicological hazards of
inhaled nanoparticles—potential implications for drug delivery.
to be involved in pathophysiologic monocyte recruit- J. Nanosci. Nanotechnol. 4, 521–531.
ment previously (39 – 42). It and its receptor, Flt-1, can 3. Oberdorster, G. (1996) Significance of particle parameters in
activate the PI3 kinase/AKT and ERK-1/2 pathways the evaluation of exposure-dose-response relationships of in-
haled particles. Inhal. Toxicol. 8, 73– 89
through monocytes and cause increased expression of 4. Oberdorster, G., Oberdorster, E., and Oberdorster, J. (2005)
cytokines (TNF-alpha and IL-1beta) and chemokines Nanotoxicology: an emerging discipline evolving from studies of
(monocyte chemotactic protein-1 [MCP-1, CCL2], IL-8, ultrafine particles. Environ. Health Perspect. 113, 823– 839
and macrophage inflammatory protein-1 [MIP-1, 5. Sun, D., Meng, T. T., Loong, H., and Hwa, T. J. (2004) Removal
of natural organic matter from water using a nano-structured
CCL3]) in both normal monocytes and the THP-1 photocatalyst coupled with filtration membrane. Water Sci. Tech-
monocytic cell line (43). Recently, PlGF has been nol. 49, 103–110
correlated with the pathogenesis of pulmonary emphy- 6. Afaq, F., Abidi, P., Matin, R., and Rahman, Q. (1998) Cytotox-
icity, pro-oxidant effects and antioxidant depletion in rat lung
sema, as PlGF transgenic mice show lung epithelial cell alveolar macrophages exposed to ultrafine titanium dioxide.
apoptosis and spontaneous pulmonary emphysema J. Appl. Toxicol. 18, 307–312
(13). We suggest that PlGF may be involved in nano- 7. Oberdorster, G., Finkelstein, J. N., Johnston, C., Gelein, R., Cox,
TiO2-induced pulmonary emphysema by regulating C., Baggs, R., and Elder, A. C. (2000) Acute pulmonary effects of
ultrafine particles in rats and mice. Res. Rep. Health Eff. Inst. 96,
certain inflammatory responses, including chemokines 5–74
induction, macrophage infiltration, cell proliferation 8. Wottrich, R., Diabate, S., and Krug, H. F. (2004) Biological
and apoptosis. Previous PlGF knockout mice studies effects of ultrafine model particles in human macrophages and
epithelial cells in mono- and co-culture. Int. J. Hyg. Environ.
have shown that the absence of PlGF could reduce Health 207, 353–361
vascular leakage induced by skin wounding, allergens, 9. Peters, K., Unger, R. E., Kirkpatrick, C. J., Gatti, A. M., and
and neurogenic inflammation in many diseases, also Monari, E. (2004) Effects of nano-scaled particles on endothe-
the inflammatory angiogenesis and edema formation lial cell function in vitro: studies on viability, proliferation and
inflammation. J. Mater. Sci. Mater. Med. 15, 321–325
were inhibited (44, 45). These results indicated that the 10. Chellat, F., Grandjean-Laquerriere, A., Le Naour, R., Fernandes,
important role of PlGF in immuno-regulation. More J., Yahia, L., Guenounou, M., and Laurent-Maquin, D. (2005)
E1740 Vol. 20 November 2006 The FASEB Journal CHEN ET AL.Metalloproteinase and cytokine production by THP-1 macro- (1993) Macrophage inflammatory proteins 1 and 2: expression by
phages following exposure to chitosan-DNA nanoparticles. Bio- rat alveolar macrophages, fibroblasts, and epithelial cells and in rat
materials 26, 961–970 lung after mineral dust exposure. Am. J. Respir. Cell Mol. Biol. 8,
11. Hogg, J. C. (2004) Pathophysiology of airflow limitation in 311–318
chronic obstructive pulmonary disease. Lancet 364, 709 –721 31. Londhe, V. A., Belperio, J. A, Keane, M. P., Burdick, M. D., Xue,
12. American Thoracic Society Patient information series. (2005) Y. Y., and Strieter, R. M. (2005) CXCR2 is critical for dsRNA-
Chronic obstructive pulmonary disease (COPD). Am. J. Respir. induced lung injury: relevance to viral lung infection. J. Inflamm.
Crit. Care Med. 171, 3– 4 (London) 2, 4
13. Barnes, P. J. (2003) New concepts in chronic obstructive pul- 32. Jeyaseelan, S., Manzer, R., Young, S. K., Yamamoto, M., Akira, S.,
monary disease. Annu. Rev. Med. 54, 113–129 Mason, R. J., and Worthen, G. S. (2005) Induction of CXCL5
14. Tsao, P. N., Su, Y. N., Li, H., Huang, P. H., Chien, C. T., Lai, during inflammation in the rodent lung involves activation of
Y. L., Lee, C. N., Chen, C. A., Cheng, W. F., and Wei, S. C. alveolar epithelium. Am. J. Respir. Cell Mol. Biol. 32, 531–539
(2004) Overexpression of placenta growth factor contributes to 33. Walrath, J., Zukowski, L., Krywiak, A., and Silver, R. F. (2005)
the pathogenesis of pulmonary emphysema. Am. J. Respir. Crit. Resident Th1-like effector memory cells in pulmonary recall
Care Med. 169, 505–511 responses to Mycobacterium tuberculosis. Am. J. Respir. Cell Mol.
15. Yao, P. L., Tsai, M. F., Lin, Y. C., Wang, C. H., Liao, W. Y., Chen, Biol. 33, 48 –55
J. J., and Yang, P. C. (2005) Global expression profiling of 34. Lindell, D. M., Standiford, T. J., Mancuso, P., Leshen, Z. J., and
theophylline response genes in macrophages: evidence of air- Huffnagle, G. B. (2001) Macrophage inflammatory protein
way anti-inflammatory regulation. Respir. Res. 8, 89 –95 1alpha/CCL3 is required for clearance of an acute Klebsiella
16. Lam, C. W., James, J. T., McCluskey, R., and Hunter, R. L. pneumoniae pulmonary infection. Infect. Immun. 69, 6364 – 6369
(2004) Pulmonary toxicity of single-wall carbon nanotubes in 35. Golpon, H. A., Coldren, C. D., Zamora, M. R., Cosgrove, G. P.,
mice 7 and 90 days after intratracheal instillation. Toxicol. Sci. Moore, M. D., Tuder, R. M., Geraci, M. W., and Voelkel, N. F.
77, 126 –134 (2004) Emphysema lung tissue gene expression profiling. Am. J.
17. Leong, B. K., Coombs, J. K., Sabaitis, C. P., Rop, D. A., and Respir. Cell Mol. Biol. 31, 595– 600
Aaron, C. S. (1998) Quantitative morphometric analysis of 36. Van Buchem, M. A., Levelt, C. N., Hogendoorn, P. C., Colly,
pulmonary deposition of aerosol particles inhaled via intratra- V. P., Kluin, P. M., Willemze, R., and Daha, M. R. (1993)
cheal nebulization, intratracheal instillation or nose-only inha- Involvement of the complement system in the pathogenesis of
lation in rats. J. Appl. Toxicol. 18, 149 –160 pulmonary leukostasis in experimental myelocytic leukemia.
18. Hoyle, G. W., Li, J., Finkelstein, J. B., Eisenberg, T., Liu, J. Y., Leukemia. 7, 1608 –1614
Lasky, J. A., Athas, G., Morris, G. F., and Brody, A. R. (1999) 37. Morgan, B. P. (1995) Physiology and pathophysiology of comple-
Emphysematous lesions, inflammation, and fibrosis in the lungs ment: progress and trends. Crit. Rev. Clin. Lab. Sci. 32, 265–298
of transgenic mice overexpressing platelet-derived growth fac- 38. Borders, C. W., Courtney, A., Ronen, K., Pilar Laborde-Lahoz,
tor. Am. J. Pathol. 154, 1763–1775 M., Guidry, T. V., Hwang, S. A., Olsen, M., Hunter, R. L., Jr.,
19. Thurlbeck, W. M. (1967) Measurement of pulmonary emphy- Hollmann, T. J., Wetsel, R. A., and Actor, J. K. (2005) Requisite
sema. Am. Rev. Respir. Dis. 95, 752–764 role for complement C5 and the C5a receptor in granulomatous
20. Yu, S. L., Chen, H. W., Yang, P. C., Peck, K., Tsai, M. H., Chen, response to mycobacterial glycolipid trehalose 6,6⬘-dimycolate.
J. J., and Lin, F. Y. (2004) Differential Gene Expression in Scand. J. Immunol. 62, 123–130
Gram-Negative and Gram-Positive Sepsis. Am. J. Resp. Crit. Care 39. Kader, H. A., Tchernev, V. T., Satyaraj, E., Lejnine, S., Kotler, G.,
Med. 169, 1135–1143 Kingsmore, S. F., and Patel, D. D. (2005) Protein microarray
21. Negoescu, A., Lorimier, P., Labat-Moleur, F., Drouet, C., Rob- analysis of disease activity in pediatric inflammatory bowel disease
ert, C., Guillermet, C., Brambilla, C., and Brambilla, E. (1996) demonstrates elevated serum PLGF, IL-7, TGF-beta1, and IL-12p40
In situ apoptotic cell labelling by the TUNEL method: improve- levels in Crohn’s disease and ulcerative colitis patients in remission
ment and evaluation on cell preparation. J. Histochem. Cytochem. versus active disease. Am. J. Gastroenterol. 100, 414 – 423
44, 959 –968 40. Levine, R. J., Thadhani, R., Qian, C., Lam, C., Lim, K. H., Yu,
22. Mutasa, H. C., and Pearson, E. C. (1988) Use of light micro- K. F., Blink, A. L., Sachs, B. P., Epstein, F. H., and Sibai, B. M.
scopic immunotechniques in selecting preparation conditions (2005) Urinary placental growth factor and risk of preeclamp-
and immunoprobes for ultrastructural immunolabelling of lac- sia. J. Am. Med. Assoc. 293, 77– 85
toferrin. Histochem. J. 20, 558 –566 41. Perelman, N., Selvaraj, S. K., Batra, S., Luck, L. R., Erdreich-
23. Chen, H. W., Yu, S. L., Chen, W. J., Yang, P. C., Chien, C. T., Epstein, A., Coates, T. D., Kalra, V. K., and Malik, P. (2003)
Chou, H. Y., Li, H. N., Peck, K., Huang, C. H., Lin, F. Y., and Placenta growth factor activates monocytes and correlates with
Lee, Y. T. (2004) Profiling of gene expression pattern during sickle cell disease severity. Blood 102, 1506 –1514
postnatal myocardial development by cDNA microarray. Heart 42. Luttun, A., Tjwa, M., Moons, L., Wu, Y., Angelillo-Scherrer, A.,
90, 927–934 Liao, F., Nagy, J. A., Hooper, A., Priller, J., and De Klerck, B.
24. Lennon, G., Auffray, C., Polymeropoulos, M., and Soares, (2002) Revascularization of ischemic tissues by PlGF treatment,
M. B. (1996) The I.M.A.G.E. Consortium: an integrated and inhibition of tumor angiogenesis, arthritis and atheroscle-
molecular analysis of genomes and their expression. Genomics rosis by anti-Flt1. Nat. Med. 8, 831– 840
33, 151–152 43. Selvaraj, S. K., Giri, R. K., Perelman, N., Johnson, C., Malik, P.,
25. Kanehisa, M., Goto, S., Kawashima, S., and Nakaya, A. (2002) The and Kalra, V. K. (2003) Mechanism of monocyte activation and
KEGG databases at Genome Net. Nucleic Acids Res. 30, 42– 46 expression of proinflammatory cytochemokines by placenta
26. Chen, H. W., Chen, J. J., Yu, S. L., Li, H. N., Yang, P. C., Su, growth factor. Blood 102, 1515–1524
C. M., Au, H. K., Chang, C. W., Chien, L. W., and Chen, C. S. 44. Luttun, A., Brusselmans, K., Fukao, H., Tjwa, M., Ueshima, S.,
(2005) Transcriptome analysis in blastocyst hatching by cDNA Herbert, J. M., Matsuo, O., Collen, D., Carmeliet, P., and Moons,
microarray. Hum. Reprod. 20, 2492–2501 L. (2002) Loss of placental growth factor protects mice against
27. Chen, J. J., Lin, Y. C., Yao, P. L., Yuan, A., Chen, H. Y., Shun, vascular permeability in pathological conditions. Biochem. Bio-
C. T., Tsai, M. F., Chen, C. H., and Yang, P. C. (2005) phys. Res. Commun. 295, 428 – 434
Tumor-associated macrophages: the double-edged sword in 45. Oura, H., Bertoncini, J., Velasco, P., Brown, L. F., Carmeliet, P.,
cancer progression. J. Clin. Oncol. 23, 953–964 and Detmar, M. (2003) A critical role of placental growth factor
28. Jeyaseelan, S., Chu, H. W., Young, S. K., and Worthen, G. S. in the induction of inflammation and edema formation. Blood
(2004) Transcriptional profiling of lipopolysaccharide-induced 101, 560 –567
acute lung injury. Infect. Immun. 72, 7247–7256 46. Hohr, D., Steinfartz, Y., Schins, R. P., Knaapen, A. M., Martra,
29. Steerenberg, P. A., Zonnenberg, J. A., Dormans, J. A., Joon, G., Fubini, B., and Borm, P. J. (2002) The surface area rather
P. N., Wouters, I. M., van Bree, L., Scheepers, P. T., and Van than the surface coating determines the acute inflammatory
Loveren, H. (1998) Diesel exhaust particles induced release of response after instillation of fine and ultrafine TiO2 in the rat.
interleukin 6 and 8 by (primed) human bronchial epithelial Int. J. Hyg. Environ. Health 205, 239 –244
cells (BEAS 2B) in vitro. Exp. Lung Res. 24, 85–100
30. Driscoll, K. E., Hassenbein, D. G., Carter, J., Poynter, J., Asquith, Received for publication May 17, 2006.
T. N., Grant, R. A., Whitten, J., Purdon, M. P., and Takigiku, R. Accepted for publication June 19, 2006.
NANOTIO2-INDUCED EMPHYSEMA E1741The FASEB Journal • FJ Express Summary
Titanium dioxide nanoparticles induce emphysema-like
lung injury in mice
Huei-Wen Chen,* Sheng-Fang Su,*,† Chiang-Ting Chien,‡ Wei-Hsiang Lin,§
Sung-Liang Yu,† Cheng-Chung Chou,† Jeremy J. W. Chen,†,储,1 and Pan-Chyr Yang†,††,1,2
*Department and Institute of Pharmacology, School of Medicine, National Yang-Ming University,
Taipei, Taiwan; †NTU Center for Genomic Medicine, National Taiwan University, Taipei, Taiwan;
‡
Department of Medical Research, National Taiwan University Hospital, Taipei, Taiwan; §Institute of
Life Sciences, College of Life Sciences, National Chung-Hsing University, Taichung, Taiwan;
储
Institutes of Biomedical Sciences and Molecular Biology, College of Life Sciences, National Chung-
Hsing University, Taichung, Taiwan; and ††Department of Internal Medicine, National Taiwan
University Hospital and National Taiwan University College of Medicine, Taipei, Taiwan
To read the full text of this article, go to http://www.fasebj.org/cgi/doi/10.1096/fj.06-6485fje
SPECIFIC AIMS genes (1.5-fold difference) of nanoTiO2-induced genes
were categorized and integrated to fit the transduction
Titanium dioxide nanoparticles (nanoTiO2) have been
signaling map using the KEGG and BIOCARTA path-
widely used as a photocatalyst in air and water cleaning.
way database. According to cDNA microarray analysis,
However, inhalation of these nanoparticles may cause
318 was up-regulated and 188 down-regulated at wk 1 in
pulmonary toxicity that is often ignored, and its mech-
nanoTiO2-treated mice compared with normal saline-
anism is not fully understood. The specific aims of this
treated mice, including cell growth regulators (cdc2a,
study were to 1) investigate the pulmonary toxicity of
cyclin B, D, and E), vascular endothelial growth factor
nanoTiO2 in mice model and 2) study the pathological,
(VEGF) -related factors (PlGF), G-protein-coupled re-
cellular, and molecular mechanisms of nanoTiO2-in-
ceptors (GPCR), chemokines (CXCL1, CXCL5, and
duced pulmonary diseases in vivo and in vitro.
CCL3), matrix metalloproteinases (MMP2 and 15), and
other immune response factors. PlGF and these che-
PRINCIPAL FINDINGS mokines have been reported to be involved in the
pathogenesis of pulmonary emphysema. According to
1. NanoTiO2 can induce emphysema-like lung injury the pathway analysis, four major pathways were up-
in mice (Fig. 1) regulated by nanoTiO2: the cell cycle regulatory path-
We found that 1 wk after single intratracheal instillation way, apoptosis pathway, the chemokines pathway, and
with 0.1 mg nanoTiO2 in mice, the lungs showed signifi- complement cascade (classical pathway).
cant changes in morphology and histology, including
disruption of the alveolar septa and alveolar enlargement 3. The PlGF/chemokine pathway may involve in
(emphysematous change), type II pneumocyte prolifera- nanoTiO2-induced pulmonary injury (Fig. 2)
tion, increased alveolar epithelial thickness, and accumu-
lation of particle-laden macrophages (Fig. 1A, B). The
Real-time quantitative RT-polymerase chain reaction
mean linear intercept (MLI) of interalveolar wall dis-
(RT-PCR) analysis showed that plgf, chemokines
tance, the air space area, and the septal chord length (a
(cxcl1, cxcl5, and ccl3), TRAIL, and prostaglandin E
parameter that measures alveolar septal thickness) were
receptor 4 (ptger4, EP4) were significantly up-regu-
significantly increased the first week (acute-phase) after
lated in lung tissues of mice treated with nanoTiO2 for
instillation of nanoTiO2, and the pathological changes
1 wk (Fig. 2A). Western blotting also showed that nano-
persisted until the second week (chronic phase) (Fig. 1C).
TiO2 caused significant induction of PlGF expression in a
Immunostaining of the nanoTiO2-induced lung injury
dose-dependent manner (Fig. 2B), while ELISA analysis
revealed a significant increase in macrophage accumula-
showed that nanoTiO2-treated mice had higher serum
tion, alveolar type II cell proliferation, and alveolar epi-
thelial cell apoptosis 1 wk after nanoTiO2 treatment.
1
These authors contributed equally to this work.
2. Microarray gene expression profiles of the nanoTiO2- 2
Correspondence: Department of Internal Medicine Na-
induced injured lung and signaling pathway analysis tional Taiwan University Hospital and National Taiwan Uni-
versity of Medicine College No. 7, Chung-Shan South Rd.,
The nanoTiO2-induced injured mice lungs were exam- Taipei, 100, Taiwan. E-mail: pcyang@ha.mc.ntu.edu.tw
ined by cDNA microarray. The differentially expressed doi: 10.1096/fj.06-6485fje
0892-6638/06/0020-2393 © FASEB 2393Figure 1. NanoTiO2 nanoparticle-induced
pulmonary morphological and histological
changes. A) Morphological (a, b) and histo-
logical (c, d ) changes (H/E, hematoxylin and
eosin staining) in the mouse lung 1 wk after
intratracheal instillation with NS (normal sa-
line, a, c) or 0.1 mg/mouse nanoTiO2 (b, d ).
Arrowheads indicate the nodule-like lesions
caused by chronic inflammation. Original
magnification ⫻100, bar ⫽ 100 m for H/E
histological image (c, d ). B) Histological
changes in the mouse lung after intratracheal
instillation with nanoTiO2 for 1 wk. Lung
tissues were collected from NS-treated control
mice (a) and nanoTiO2 (0.1 mg/mouse)
-treated mice (b, c). Original magnification,
⫻400, bar ⫽ 50 m. Similar results were
obtained in 6 independent experiments. C)
Morphometric measurements from lungs at 3
days, 1 wk, and 2 wk after installation with NS
or nanoTiO2 (0.1 or 0.5 mg/mouse). The
mean linear intercept (MLI), mean air space
area, and septal chord length were measured (n⫽6) as described in Materials and Methods. The data are the means ⫾sd.
*P ⬍ 0.05 in Student’s t test compared with the control (NS) group.
levels of PlGF protein (Fig. 2C), which might be produced is triggered by nanoTiO2 activation of macrophages,
mainly by infiltrating macrophages and some pulmonary up-regulations of PlGF, and other inflammatory cyto-
epithelial cells, as shown by immunostaining with PlGF- kines that resulted in disruption of alveolar septa,
specific antibodies (Fig. 2D). alveolar epithelial injury, alveolar epithelial cell prolif-
eration, and apoptosis. This information may have
4. In vitro exposures of nanoTiO2 in macrophage cell important clinical implications regarding safety, as
line THP-1 dose-dependently induce significant nanoTiO2 are widely used as a photocatalyst in air and
increase of PlGF, Cxcl5, and Ccl2 (MCP1) expression water cleaning, and TiO2 is used as a pigment in the
paint industry. Extra caution therefore should be taken
CONCLUSIONS AND SIGNIFICANCE in handling higher doses of nanoTiO2.
In this study, the microarray gene expressions and
The results of this study indicate that single intratra- pathway analysis indicated that the cell cycle, apoptosis,
cheal instillation of 0.1 mg nanoTiO2 can induce severe chemokine, and complement pathways may be involved
pulmonary inflammation and emphysema in the mouse in nanoTiO2-induced pulmonary toxicity (Fig. 3). Acti-
lung. Our results indicate that pulmonary emphysema vation of the cell cycle pathway suggests that nanoTiO2
Figure 2. Expression of PlGF, chemokines, and
related factors in mice after a single intratra-
cheal instillation with nanoTiO2. A) Real-time
quantitative RT-PCR for flt-1 (PlGF receptor),
flt-3, plgf, chemokines (cxcl1, cxcl5, ccl3), and
apoptosis-related factors (trail and ptger4). The
data are expressed as the fold increase com-
pared with the NS control ⫾sd. *P ⬍ 0.05 vs.
the NS control; **P ⬍ 0.05 vs. microTiO2
(n⫽4). B) Western blotting for the effect of
nanoTiO2 on PlGF protein expression. The
right panel shows the expression of PlGF rela-
tive to that for ␣-tubulin expressed as a fold
increase compared with the NS control ⫾sd.
*P ⬍ 0.05 vs. NS control (n⫽4). C) Serum PlGF
protein levels measured by ELISA. Data are
expressed as the mean ⫾sd. *P ⬍ 0.05 vs.
controls (n⫽4). D) Immunohistochemical stain-
ing with anti-PlGF2 antibody (Ab) showing
overexpression of PlGF (brown color, DAB
staining) in mice lung tissues after intratracheal
instillation with nanoTiO2 (0.1 mg and 0.5 mg
per mice). Similar results were obtained in 4
experiments.
2394 Vol. 20 November 2006 The FASEB Journal CHEN ET AL.You can also read

















































