JBC Papers in Press. Published on August 9, 2018 as Manuscript RA118.005006
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JBC Papers in Press. Published on August 9, 2018 as Manuscript RA118.005006
The latest version is at http://www.jbc.org/cgi/doi/10.1074/jbc.RA118.005006
Bacterially expressed HIV-1 gp120 outer-domain fragment immunogens with improved stability
and affinity for CD4 binding site neutralizing antibodies
Ujjwal Rathore1, Mansi Purwar1, Venkada Subramanian Vignesh1, Raksha Das1, Aditya Arun Kumar1,
Sanchari Bhattacharyya1, Heather Arendt2, Joanne DeStefano2, Aaron Wilson2, Christopher Parks2, Celia
C. La Branche3, David C. Montefiori3, Raghavan Varadarajan1,*.
From the 1 Molecular Biophysics Unit, Indian Institute of Science, Bangalore, India 560012, 2
International AIDS Vaccine Initiative, Brooklyn, NY 11226, USA, 3 Department of Surgery, Duke
University Medical Center, Durham, NC, USA.
Running Title: Design of HIV-1 outer domain immunogens
*To whom Correspondence should be addressed: Raghavan Varadarajan: Molecular Biophysics Unit,
Indian Institute of Science, Bangalore, India 560012; varadar@mbu.iisc.ernet.in
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
Keywords: protein design, mutagenesis, protein refolding, glycosylation, Yeast surface display,
Hydrogen deuterium exchange, vaccine development
given. Following two gp120 boosts, sera
ABSTRACT collected at week 22 showed cross-clade
Protein-minimization is an attractive approach neutralization of Tier 1 HIV-1 viruses. Using a
for designing vaccines against rapidly evolving number of different prime: boost combinations,
pathogens such as HIV-1 since it can help in we have identified a cyclically permuted OD
focussing the immune response towards fragment as the best priming immunogen, and a
conserved conformational epitopes present on trimeric, cyclically permuted gp120 as the most
complex targets. The outer domain (OD) of suitable boosting molecule amongst the tested
HIV-1 gp120 contains epitopes for a large immunogens. This study also provides insights
number of neutralizing antibodies and therefore into some of the biophysical correlates of
is a primary target for structure-based vaccine improved immunogenicity.
design. We have previously designed a
bacterially expressed outer domain immunogen
(ODEC) that bound CD4 binding site (CD4bs) Introduction
ligands with 3-12µM affinity and elicited a Designing an effective vaccine against Human
modest neutralizing antibody response in rabbits. Immunodeficiency Virus (HIV-1) is one of the
In this study, we have optimized ODEC using most challenging scientific problems of the
consensus sequence design, cyclic permutation current century. Elicitation of broadly
and structure-guided mutations to generate a neutralizing antibodies (bNAbs) is a desirable
number of variants with improved yields, trait for any anti-HIV-1 vaccine. The HIV-1
biophysical properties, stabilities and affinities surface envelope (Env) glycoprotein gp120 is
(KD of 10-50 nM) for various CD4bs targeting, the major target for vaccine design, since it
broadly neutralizing antibodies, including the contains sites for cellular receptor and co-
germline reverted version of the broadly receptor binding, and is the primary target of the
neutralizing antibody, VRC01. In contrast to humoral immune response. The primary reason
ODEC, the optimized immunogens elicited high for the difficulty in generating an effective
anti-gp120 titers in rabbits as early as 6 weeks vaccine against HIV-1 lies in its extensive
post-immunization, before any gp120 boost was sequence variability. Moreover, the conserved
1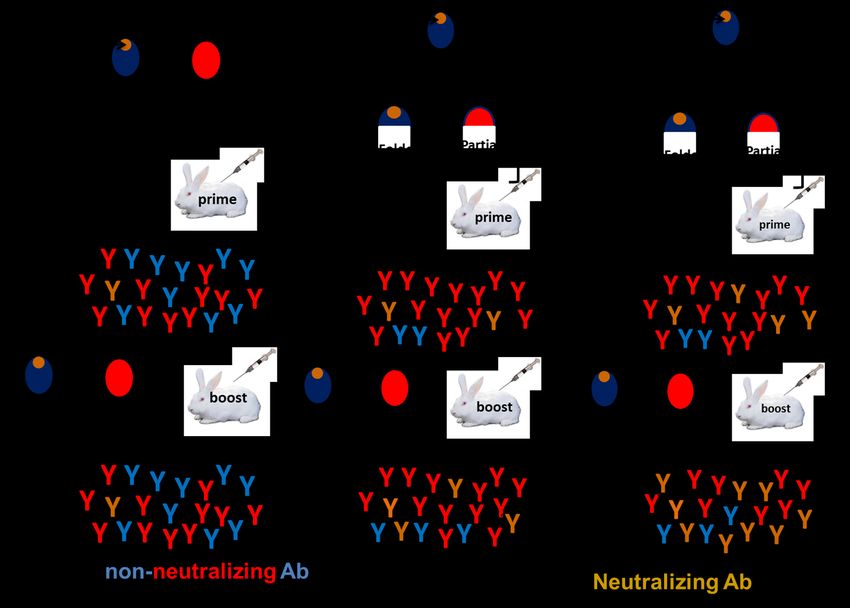
Design of HIV-1 outer domain immunogens
epitopes targeted by broadly neutralizing Results
antibodies are often discontinuous in sequence
and heavily glycosylated. Approaches to focus The initial OD design. The goal of this work
the immune response towards specific was to generate fragment immunogens targeting
conformational epitopes targeted by known conserved sites on the gp120 outer domain (OD)
broadly neutralizing antibodies (bNAbs) against with high-affinity. All outer domain
HIV-1 are desirable. Protein-minimization is an immunogens described in this study are based on
attractive approach for designing vaccines a previously designed construct known as ODEC
against rapidly evolving pathogens since it can (7). ODEC design was focused on the CD4
help in focussing the immune response towards binding site (CD4bs) of gp120 glycoprotein
conserved, conformational epitopes present on (Figure 1A-B; Table 1). In summary, CD4
complex protein targets. The outer domain (OD) (primary cellular receptor) interacting residues
of HIV-1 gp120 contains epitopes for a large of gp120 were identified by calculating amino
number of bNAbs (1-6). OD is therefore acid accessibilities in the presence and absence
considered to be an important candidate for of CD4 using coordinates from the gp120-CD4
structure-based vaccine design. We previously crystal structure (PDBID 1G9M (8)). Most CD4
reported the design of a non-glycosylated, interacting residues were found to be present in
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
E. coli expressed outer domain fragment the OD and bridging sheet regions (Figure 1A).
immunogen (ODEC) that bound CD4 with Thus, an OD fragment (ODEC) from residue 255-
micromolar affinity and elicited a modest 474 having most of the OD and a part of the
neutralizing antibody response in rabbits (7). bridging sheet was selected for bacterial
ODEC bound CD4 and bNAb b12 with ~200-250 expression (Figure 1B). ODEC retained ~ 70 % of
fold weaker affinity (3-12µM) as compared to the CD4 epitope. It was devoid of the inner
full-length gp120 (10-30nM). ODEC also showed domain, part of the bridging sheet, V1V2 and V3
a high tendency to aggregate during purification variable loops and had 11 computationally
and storage. Sera elicited in ODEC immunized guided and rationally designed hydrophobic to
animals without boosting with gp120 showed hydrophilic mutations at the interface of inner
very weak anti-gp120 titers (7). and outer domain to prevent aggregation and
In the current study, we rationally modified the increase solubility, to minimize the chances of
initial ODEC design to generate several aggregation (7).
derivatives with improved yield, biophysical
properties, immunogenicity and nanomolar Purification and characterization of ODEC
binding affinity to broadly neutralizing refolded in the presence and absence of
antibodies targeting the primary receptor (CD4) ArgHCl. In the original study (7), ODEC was
binding site. All OD immunogens described in purified by performing a 10 fold rapid dilution
this study could also bind to the germline- of denatured protein into PBS containing 1mM
reverted version of the CD4bs directed VRC01 EDTA, in order to reduce the GdnCl
antibody with nM high-affinity. Immunization of concentration from 6M to 0.6M. This sudden
rabbits with ODEC followed by boosting with refolding protocol was developed because ODEC
gp120 resulted in no detectable neutralization at exhibits a very high tendency to aggregate upon
week 22. However, the improved OD slow removal of denaturant via dialysis. Sudden
immunogens designed in this study could refolding of protein resulted in decreased but
rapidly elicit sera with heterologous cross-clade still significant precipitation during the refolding
neutralization of Tier 1 viruses after 22 weeks. step and also during storage. Moreover, the
This study also provides insights into some of refolded protein bound weakly with CD4 and
the biophysical correlates of improved bNAb b12, as compared to full-length gp120,
immunogenicity. indicating that it is not well folded (7). In order
to obtain a better-folded protein, we explored a
number of variations in the refolding protocol
and found that the inclusion of 0.5M ArgHCl
2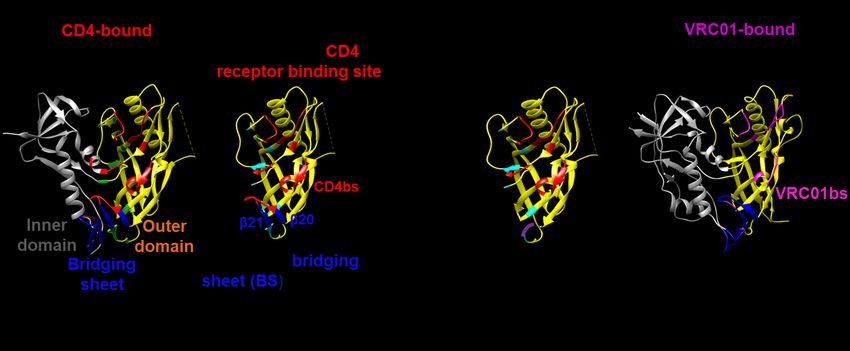
Design of HIV-1 outer domain immunogens
resulted in reduced precipitation. In comparison causes the antibody to reduce its contact with the
to the ODEC protein that was refolded in the bridging sheet and the CD4 binding loop, while
absence of ArgHCl, the CD spectrum of the contacting the V5 loop in the proximal barrel of
ArgHCl refolded protein showed a spectrum the outer domain (Figure 1D) (1). As VRC01
characteristic of a beta-sheet (Figure 3A). This is makes minimal contacts with the bridging sheet,
expected since, with the exception of one helix we removed the two bridging sheet strands
(A2), the outer domain is largely composed of present in the ODEC molecule. Bridging sheet
beta-sheet structure. residues from 423 to 434 were deleted and
residue 422 was connected to residue 435 using
SPR binding studies with ArgHCl refolded a Gly-Ala-Gly (GAG) linker in the ∆BS-ODEC
ODEC. We characterized the ArgHCl refolded construct (∆BS stands for bridging sheet deleted)
ODEC molecule for binding to 4-domain soluble (Figure 1C). We hypothesized that apart from
CD4 (sCD4) and to the broadly neutralizing focussing the immune response to the VRC01
antibodies b12 and VRC01. VRC01 is a potent, epitope, removal of this possibly unstructured
broadly neutralizing antibody (1,2) that binds to bridging sheet region in ∆BS-ODEC construct
the CD4 binding site and shows cross-clade might also result in improved stability and other
neutralization. As shown in Table 2, ArgHCl biophysical properties.
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
refolded ODEC bound sCD4 with ~15 fold, and Methods for engineering proteins with improved
b12 with ~ 80 fold improved affinity as stability can be classified as rational structure-
compared to the protein refolded in the absence based methods, directed evolution based
of ArgHCl (7). ODEC (ArgHCl refolded) binds methods and semi-rational sequence frequency-
only ~5-15 times weaker to these ligands than based methods (9). It is often difficult to predict
core gp120 lacking V1/V2 and V3 variable stabilizing mutations solely using structure-
loops. Interestingly, ArgHCl refolded ODEC also based methods. Frequency-based methods make
bound the antibody VRC01 with a KD of use of amino acid conservation amongst
359±55nM, which was about 15 fold weaker homologous proteins to identify possible
when compared to gp120 (Table 2). beneficial mutations. Consensus protein design
is based on the assumption that the frequency of
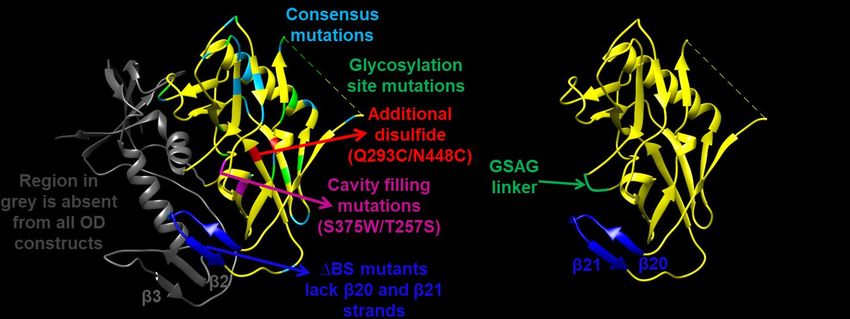
Designing next generation OD fragment a given amino acid at a particular position in a
immunogens. The ODEC construct was further multiple sequence alignment (MSA) of related
modified with an aim to improve its biophysical proteins is directly correlated with that amino
and binding properties. A part of the bridging acid’s contribution to protein stability or
sheet was included in the original ODEC design, function. In other words, more conserved
along with most of the outer domain of gp120 residues contribute more to the stability of a
(Figure 1B). In the CD4 bound conformation of protein, and thus, replacing non-consensus
gp120, the bridging sheet is made up of four residues with the consensus (most frequent) ones
anti-parallel β strands (β3, β2, β21, β20). β21 could result in stabilization (9-12). Using this
and β20 strands are close to the outer domain consensus-based protein design approach, we
and are unlikely to be folded in the absence of made an ODEC variant named as
the remaining two strands (8). However, these ODECConsensus (Figure 2). Briefly, the most
two strands were included in the original ODEC frequent amino acid at each position in a
design to retain certain critical CD4 contacts multiple sequence alignment of 814 HIV-1 B-
(Figure 1B). The broadly neutralizing antibody subtype sequences was identified. Each residue
VRC01 neutralizes around 91% of 190 viruses in ODEC was mutated to the most frequent
tested, with a geometric mean IC50 of about residue at that position in the MSA. Residues
0.33μg/ml VRC01 (2). VRC01 targets the involved in interaction with CD4, bNAb b12 and
CD4bs in the outer domain, it rotates about 40 bNAb VRC01 were identified using accessible
degrees with respect to CD4 and translates 6Å surface calculations as described previously (13)
away from the bridging sheet region, making the and these were not mutated. In addition, eleven
approach angle different from that of CD4. This hydrophobic to hydrophilic mutations introduced
3Design of HIV-1 outer domain immunogens
in the original ODEC molecule to prevent native protein, re-confirming that native proteins
aggregation were retained (Figure 2). Finally, a are well folded and oxidized (Figure S3A,B).
total of 27 consensus mutations were introduced ∆BS-ODEC and ODECConsensus were examined
in the ODEC background (Figure S1). Models for for binding to sCD4 and to the broadly
ODEC and ODECConsensus constructs were made neutralizing antibodies b12 and VRC01 on a
using the program MODELLER 9v1 (14,15). BIACORE 2000 machine. ODECConsensus
TKBK electrostatic calculations confirmed that bound VRC01 with about 5-6 times better
the consensus mutations have not introduced any affinity than WT ODEC. Its binding to b12 was
unfavourable electrostatic interaction in OD about 2-3 times better, while binding to CD4
(Figure S2) (16,17). was similar to ODEC (Table 2). ∆BS-ODEC bound
VRC01 about 3-4 times tighter and to b12 and
Purification, biophysical characterization and CD4 ~4-5 fold and ~2-3 fold weaker than
binding studies of new OD constructs (∆BS- corresponding values for WT ODEC. This
ODEC and ODECConsensus). Both proteins reduction in the binding affinity for b12 and
were expressed and purified from E.coli for CD4 is expected for ∆BS-ODEC, as the bridging
further characterization using ArgHCl assisted sheet region contains important residues for
sudden refolding. The yield was about 3- interaction with b12 and CD4. The bridging
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
4mg/litre of the culture for ODECConsensus sheet was deleted in this construct in an attempt
(similar to ODEC) and ~20 mg/litre for ∆BS- to stabilize the molecule and to focus the design
ODEC. This drastic improvement in the yield for on the VRC01 epitope. A comparison of all
∆BS-ODEC was attributed to the significant kinetic parameters for binding is shown in Table
reduction in precipitation during the purification 2. In an alternative attempt to improve the
process, which could be due to the removal of biophysical and binding properties of OD
the potentially unstructured bridging sheet immunogens, we also designed OD constructs
region. SDS-PAGE studies confirmed that both with additional disulfides or cavity-filling
the proteins were at least 90% pure. The mutations as described below.
expected masses for ODEC, ∆BS-ODEC and
ODECConsensus were 23033.0, 21820.7 and Design of disulfide and cavity filling mutants
23378.4 Daltons respectively, whereas observed of ODEC. Though naturally occurring disulfides
masses using ESI-MS were found to be 23035.0, are known to typically stabilize proteins, all
21821.1 and 23380.0 Daltons respectively, designed disulfides do not necessarily result in
confirming the identities of these proteins. The stabilization. Certain stereochemical criteria
expected and observed masses correspond to the need to be satisfied in order for a disulfide bond
disulfide-formed oxidized mass of proteins. to form. We have previously reported the effect
CD-spectra indicated that both the proteins are of the introduction of disulfides in different
well folded and predominantly in a β-sheet secondary structural elements in diverse proteins
conformation as expected (Figure 3A, B). The (19,20). It was found that almost all naturally
fluorescence spectrum of these proteins showed occurring cross-strand disulfides in anti-parallel
an expected red-shift and a change in emission β strands are located at non-hydrogen bonded
intensity upon denaturation, confirming that the (NHB) positions (19,21). Subsequently,
proteins are likely to be folded with the burial of experiments on model proteins such as Top7,
some tryptophan residues in the native state MBP and other periplasmic proteins showed that
(Figure 3C). Native proteins run on a C5 cysteines introduced at exposed NHB positions
analytical reverse–phase column elute as a single form disulfides spontaneously, while those
peak, thereby showing that they exist as a engineered at HB positions either remain in the
homogeneous species in solution and not as a reduced form or if they form a disulfide, result in
mixture of different disulfide bonded isomers destabilization of the protein (19). Based on the
(18). The denatured, reduced proteins eluted at a above observations, we attempted to reduce the
different acetonitrile concentration than the conformational flexibility of ODEC by
engineering disulfides at an exposed NHB
4Design of HIV-1 outer domain immunogens
position (293-448) in an anti-parallel β strand. CD4bs (18) were also mutated in this construct
Residues 293 and 448 are present at the distal to generate the ODECC2 construct (Table 1, #8).
barrel of the outer domain, have high surface
accessibility (59% and 42%, respectively) and a Yeast surface display (YSD) of the next-
4.61Ǻ distance between their Cα- Cα atoms. generation OD immunogens. Yeast surface
Both these residues were mutated to cysteines in display (YSD) was used as an initial screening
order to introduce an additional disulfide bond in system to characterize the newly designed OD
the ODEC molecule, which generated the immunogens. YSD allows rapid screening of a
ODECSS construct (Figure 4A; Table 1, #5). large number of mutants for binding with
Analysis of the crystal structure of gp120 bound various ligands, thereby alleviating the need to
to CD4 (PDBID 1G9M) shows that two express and purify the individual clones (Figure
prominent cavities exist in the binding region S4) (24). Expression on the yeast surface has
(22). A surface cavity of gp120 remains filled previously been shown to be directly correlated
with water molecules and is surrounded on all with the folding efficiency of the protein (25). In
sides by residues from gp120 and CD4. Another the YSD system, the protein of interest (OD
much deeper cavity extends into the interior of fragment immunogens in this case) is displayed
gp120. Phe43 from CD4 plugs the opening of as a fusion to the adhesion subunit of the yeast
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
this cavity, and thus it is known as the Phe43 agglutinin protein Aga2p, which attaches to the
cavity. Previously, a double mutant yeast cell wall through disulfide bonds to Aga1p
S375W/T257S of gp120 was shown to improve (Figure S4A). Expression of the Aga2p fusion is
the binding affinity to CD4 without affecting under the control of a galactose-inducible
b12 binding (23). In the current study, we promoter on the yeast display plasmid pPNLS.
attempted to reduce the conformational entropy Each yeast cell typically displays 1X104 to
of ODEC and stabilize it in a CD4 bound 1X105 copies of the protein. Protein surface
conformation by incorporating both these well- expression can be detected by using
characterized cavity-filling mutations fluorescently labelled antibodies against c-Myc
(T257S+S375W). This ODEC mutant was named and HA-epitope tags followed by FACS.
as ODEC-cavity filling (ODECCF) (Figure 4A; Binding of the surface displayed molecule with a
Table 1). ligand can be detected by the addition of a
fluorescently labelled secondary antibody
Design of ODEC constructs with multiple followed by FACS (Figure S4A). This technique
designed mutations. Introducing various allows binding affinity determination in a
stabilizing mutations individually in the ODEC surface display format without the need for
background did not result in achieving gp120 purification and expression of each individual
like affinity for bNAbs, so we decided to clone (Figure S4B). YSD can also be used for
combine the various stabilizing mutations rapid screening of mutant libraries (24,26). All
(described above) with the glycosylation site ODEC variants expressed well on the yeast
mutations earlier identified by our group (18). surface. In order to assess the conformational
The first combined construct (ODECC1) was integrity of surface displayed OD molecules,
made by deleting the bridging sheet region their binding affinities with the conformation
coupled with the introduction of an additional specific VRC01 bNAb were determined using
disulfide (294-448) and cavity filling mutations FACS (Table 3). The original ODEC molecule
in the ΔG14-ODEC background (all 14 bound VRC01 with a KD of ~400nM. ∆BS-ODEC
glycosylation sites present in the outer domain and ODECConsensus showed ~3-6 fold higher
were mutated) (18) (Table 1, #7). In another affinity for VRC01 than ODEC (Table 3).
combined construct, the additional disulfide and Importantly, these constructs did not show
cavity filling mutations were introduced in the measurable binding with the non-neutralizing
bridging sheet deleted ODECConsensus protein MAb b6 as they lack regions of the b6 epitope.
background. Four glycosylation sites close to the This improvement in affinity for the VRC01
bNAb shows that both these new constructs
5Design of HIV-1 outer domain immunogens
mimic the VRC01 binding epitope of gp120 upon unfolding (27). ODECC1 and ODECC2 also
better than the original ODEC construct, even on showed a co-operative transition in urea
the yeast surface. The VRC01 binding affinities mediated denaturation with much higher
for ODEC, ∆BS-ODEC and ODECConsensus were apparent Cm’s of 4.2M and 3.3M urea
already available from the SPR studies with the respectively (Figure 5). The free energy of
purified proteins (Table 2) and the fact that they unfolding at zero denaturant concentration
match closely with the values obtained from (ΔG°u) was ~2.8 kcal/mol for ODECC1 (m value
YSD (Table 3) demonstrates the utility of YSD. = -0.67 kcal/mol/M) and ~2.53 kcal/mol for
Both ODECC1 (Figure S5) and ODECC2 (Figure ODECC2 (m value = -0.81 kcal/mol/M). A
S6) showed substantial improvement in the comparison of Cm and ΔG°u values coupled with
binding affinity for bNAb VRC01 (Table 3) with YSD VRC01 binding measurements indicates
KD’s of about 3-4nM. In comparison, yeast that both the combined constructs ODECC1 and
displayed core gp120 bound VRC01 with a KD ODECC2 are more stable and better folded
of ~24nM (Table 3, #1). compared to the previous ODEC mutants.
Purification and characterization of ODECC1 Design of a cyclic-permutant of ODEC. Protein
and ODECC2. As both the combined constructs design largely relies on amino acid mutations to
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
when displayed on yeast surface showed alter stability or function. Circular permutation
significant improvement in VRC01 binding, we of proteins is an attractive approach for protein
expressed and purified these two proteins from design, as it allows manipulation of stability and
E.coli using ArgHCl assisted sudden refolding folding with minimal perturbation of the three
as has already been described for ODEC. The dimensional structure (28-30). In cyclically
yield was about ~15-20 mg/litre of the culture permuted proteins, the original N and C-termini
for both the proteins. are connected through a linker whereas new N
Far-UV and near-UV CD-spectra indicated that and C-termini are generated by introducing a
both the proteins are well-folded and break elsewhere in the sequence (29). The ODEC
predominantly in a β-sheet conformation (Figure molecule is an artificially created fragment
S8A, B). The fluorescence spectra of these immunogen, and it is possible that the N and C-
proteins showed an expected red-shift and a terminal regions generated due to the removal of
change in emission intensity upon denaturation, the inner domain as well as their neighbouring
indicating that the proteins are likely to be regions are not correctly folded. The original
folded with burial of some tryptophan residues ODEC design consists of residues 255 to 474
in the native state (Figure S7A,B). from gp120 and thus the residues 255 and 474
Isothermal urea denaturation studies were respectively form the N and C-termini of the
carried out to estimate stabilities of all the ODEC molecule (Figure 4B). The ODEC fragment
purified ODEC variants (ODEC, ∆BS-ODEC, lacks V1V2 and V3 loops but retains V4 loop
ODECConsensus, ODECC1 and ODECC2). The residues (7). The V4 loop is highly flexible, its
chemical unfolding of ODEC was not co- function is not well established, has a high
operative whereas both ∆BS-ODEC and tolerance for insertion, and reasonable tolerance
ODECConsensus show a co-operative isothermal for deletions in the context of native, trimeric
urea denaturation profile with apparent Cm’s of Env (31,32) and is therefore not considered to be
2.4M and 2.15M urea respectively (Figure 5). a good vaccine target (33). Due to its high
The free energy of unfolding at zero denaturant flexibility, V4 loop region electron density is not
concentration (ΔG°u) was found to be ~1.6 visible in the gp120 crystal structure (PDBID
kcal/mol for ∆BS-ODEC (m value = -0.85 1G9M) (Figure 4B) (8). To cyclically permute
kcal/mol/M) and ~2.23 kcal/mol for ODEC, new N- and C-termini were created at the
ODECConsensus (m value = -0.95 kcal/mol/M). V4 loop region via deletion of residues 398 to
Cm is the concentration of denaturant at which 409. The old N- and C-termini of ODEC were ~
50% protein is unfolded, whereas the m value is 8Ǻ apart and connected via a GSAG (Gly-Ser-
proportional to the amount of surface area buried Ala-Gly) amino acid linker (Figure 4B). The
6Design of HIV-1 outer domain immunogens
overall idea was to stabilize the regions close to equilibrium between the folded and partially
the old N and C-termini of ODEC as they are folded forms shifts towards the unfolded state
important for the binding of various CD4bs (Figure 6B). Hence, on immunization with less
ligands like CD4 and bNAb VRC01, by making stable fragment immunogens, non-neutralizing
new N and C-termini within the flexible V4 antibodies against the unfolded state can still
loop. We also hypothesized that removal of the predominate. Protein stabilization can minimize
immunodominant and flexible V4 variable loop the generation of non-neutralizing antibodies
might also result in an improved immune against the unfolded state and thus focus the
response towards the conserved CD4 binding immune response against conserved epitopes.
site. After immunization with fragment immunogens,
a full-length better-folded protein (for e.g.
Purification, characterization and SPR gp120) could be used as a boost to selectively
binding studies of ODECCycV4. The protein amplify the cross-reactive antibodies, resulting
was expressed and purified from E. coli using in increased amounts of CD4 binding site
ArgHCl assisted sudden refolding as has been antibodies (Figure 6C). Therefore, we used a
described for ODEC. The yield was about 1- prime: boost strategy to study the potential of
2mg/litre. ESI-MS analysis was performed to OD immunogens to elicit neutralizing
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
confirm the identity of the protein. It appeared antibodies. In each group, an ODEC variant was
folded from far-UV CD (Figure S8A), near-UV used as the priming immunogen at weeks 0 and
CD (Figure S8B) and fluorescence spectra 4 followed by an Env variant as the boosting
(Figure S7C). ODECCycV4 was examined for immunogen at weeks 12 and 20 (Table 4). In
binding to sCD4 and to the broadly neutralizing addition to WT JRFL gp120, two other Env
antibodies b12 and VRC01 on a BIACORE derivatives were used. iVSV-JRFL is inactivated
2000 machine. ODECCycV4 showed ~5 fold vesicular stomatitis virus displaying Env
higher on-rates and ~5 fold slower off-rates for ectodomain and MPER joined to the VSV G
binding with bNAb VRC01, which resulted in transmembrane (TM) and cytoplasmic (CT)
~20-30 fold improvement in the VRC01 binding domains on its surface (34). V1cycP gp120
affinity (KD, 16nM). ODECCycV4 also showed molecule is a trimeric cyclic-permutant of
~4-5 fold improved binding affinity for both gp120, made by introducing new N and C-
CD4 and bNAb b12. The binding affinities of termini at the V1 loop (at amino acids 144-142)
ODECCycV4 for CD4, bNAb b12 and bNAb while connecting the original N and C-termini
VRC01 are comparable with that of full-length, with a short linker. A h-CMP (human cartilage
glycosylated gp120 (Table 2). Bacterially matrix protein) trimerization domain was also
expressed, glycan free ODECCycV4 also bound added at the newly created N and C-termini to
germline-reverted VRC01 (GL-VRC01) with a generate this cyclically permuted trimeric gp120
KD of ~10-20nM whereas, as expected, full- variant (hCMP 144-142 V1cyc-JRCSF gp120)
length glycosylated gp120 did not show any hereafter referred to as V1cycP gp120 (30,35).
binding to GL-VRC01 (Table 2). All other OD All four rabbits in group 1 showed undetectable
constructs also show very high-affinity binding gp120 specific titers after two primes with ODEC
with GL-VRC01. protein (at week 0 and 4). The gp120 specific
titers rose to 105 only after two boosts with full-
Prime: boost immunization studies with OD length JRFL gp120 at week 20. In contrast
fragment immunogens. Immunization with significant anti-gp120 titers were seen as early
gp120 primarily elicits non-neutralizing as week 6, when improved ODEC designs were
antibodies, in part because the immune response used as the prime (Table 4; groups 2-8). The
is not focused on conserved neutralization average week 6 anti-gp120 titers for groups
epitopes (Figure 6A). Fragment immunogen primed with the second generation OD designs
design enables removal of non-neutralizing range from ~3103 to ~104 (Table 4) whereas for
epitopes. However, it is possible that in such ODEC immunized animals, anti-gp120 titers were
artificially designed fragment immunogens, the
7Design of HIV-1 outer domain immunogens undetectable (
Design of HIV-1 outer domain immunogens
SPR binding studies with ODECC1 and buried, amide functional groups exchange on
ODECC2. Both ODECC1 and ODECC2 were time scales that can be measured and hence can
selected for the immunization studies since they provide information on protein structure and
bound VRC01 with very high-affinity when dynamics. The exchange of protons with
displayed on yeast and demonstrated an deuteriums is probed by analysing the increase
increased resistance to urea denaturation. in mass of the protein as a function of time. The
However, sera from animals primed with rate of exchange depends on various factors such
ODECC1 failed to show any neutralization in the as pH, temperature, and the accessibility of the
TZM-bl cell assays (Table 5). This was exchangeable hydrogen to solvent (37). Buried
surprising since the week 22 anti-gp120 titers hydrogens within a protein can exchange
elicited by ODECC1 were comparable to the through local/global unfolding of the protein. As
other groups (Table 4). In order to understand shown in Figure 7A, ODEC showed a very fast
the reason for this discrepancy, we did SPR rate of exchange in the first couple of minutes,
binding studies with these two proteins (Table with about 130 backbone amide hydrogens
6). In agreement with the yeast surface display getting deuterated in that time. From 1-20
results (KD ~4nM), purified ODECC2 protein minutes, there was a slow phase, during which
bound VRC01 with a very high-affinity (KD = another 5-7 deuteriums are added. However, the
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
16±8nM). However, purified ODECC1 bound mass obtained after 20 minutes of exchange was
VRC01 with a much lower affinity (KD = identical to that obtained after 1-2 hours of
200±70nM) than it did on the yeast surface (KD exchange at 37°C (saturation). This indicated
~3.4nM) (Table 3). It is important to note that that all the backbone amide hydrogens were
the ODEC protein purified without ArgHCl exchanged within 20 minutes (Figure 7A). It is
refolding bound CD4bs ligands with 3-12 µM expected that a conformationally stable
affinity (7). The VRC01 binding affinity for the domain/subdomain in a protein should show
yeast displayed ODEC protein was ~400nM. The slower hydrogen exchange kinetics, with some
inclusion of ArgHCl in the refolding buffer protons remaining protected even after hours of
resulted in purified ODEC achieving a VRC01 exchange. These experiments show that although
affinity comparable to that on the yeast surface ODEC appears to be well-structured by CD and
(Tables 2 and 3). These data suggest that the shows a prominent thermal transition, it is labile
protein purification protocol used in the present to hydrogen-deuterium exchange and therefore
study does not result in properly refolded dynamically flexible (Figure 7A). Both ΔBS-
ODECC1 protein and therefore VRC01 binding ODEC and ODECConsensus proteins show similar
affinity is much lower for protein purified from exchange kinetics with near saturation observed
E. coli as compared to protein displayed on near around 20 minutes, though 4-6 protons
yeast. exchange between 20 minutes to 1 hour, while
ODEC attains complete saturation at 20 minutes
(Figure 7A-C). This indicates a small
HDX-MS studies. We next performed improvement in the local structural rigidity for
hydrogen-deuterium exchange (HDX) studies both the designs. Surprisingly ODECC2, which
coupled to mass spectrometry to probe the elicited sera with moderate pseudoviral
conformational flexibility of both the initial neutralization appears to have faster hydrogen
ODEC construct and of those OD immunogens exchange kinetics as compared to ODEC (Figure
which elicited measurable neutralization in 7D). In case of ODECCycV4, a fast exchange of
TZM-bl assays. In these HDX experiments, about 115 protons is seen as in ODEC. A near
protonated protein is placed in D2O (pH ~6.8). saturation value is reached at about 20 minutes.
Under these conditions, exchangeable protons, However, a slow exchange of about 4 protons is
such as amide protons and other protons seen up to 2 hours, after which saturation is
attached to polar/charged atoms, are replaced by reached. Thus, cyclic permutation results in a
deuterium. Protons present in side chains significantly slower exchange of some protons
exchange very fast but hydrogens linked to (Figure 7E). In all cases, around 30% of protons
9Design of HIV-1 outer domain immunogens
show rapid back exchange and hence could not In contrast to the ODEC refolded in PBS, the CD
be probed. HDX-MS profiles of corresponding spectrum of the ArgHCl refolded protein showed
regions in full length gp120 follow similar a characteristic spectrum of a beta-sheet
exchange profiles as outer domain derived consistent with its known structure in the context
immunogens for most of the peptides (38). For of gp120. The ArgHCl refolded ODEC bound
most of the peptides in the outer domain, about CD4 and b12 respectively with approximately
80% exchange is complete by 30 minutes with 15 and 80 times higher affinity than the ODEC
the exception of peptides 282-286 and 436-443 refolded in the absence of ArgHCl. ODEC, when
which show less than 20% exchange even after 4 displayed on the yeast surface, binds VRC01
hours (38). In case of the BG505 SOSIP.664 with a KD of 400nM, ODEC refolded in the
gp140 trimer, most of the outer domain region presence of ArgHCl also binds VRC01 with a
peptides show exchange of about 75% protons in similar affinity indicating that this in vitro
30 minutes with the exception of 284-287 and refolded molecule is probably as well folded as
446-453 peptides which show less than 20% the one folded in vivo in a eukaryotic system.
exchange even after 4 hours (39). The slightly However, these affinities were still 5-15 times
faster exchange seen for our constructs weaker than those of core gp120 for VRC01 and
compared to full length gp120 and trimeric therefore we attempted to stabilize ODEC further
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
gp140 may be due to truncation of these through mutations.
fragment immunogens from a large protein, Two such modified ODEC immunogens (∆BS-
leading to greater solvent accessibility of several ODEC and ODECConsensus), when purified using
regions. the improved refolding protocol, bound various
CD4bs ligands with KD’s of ~100nM. However,
Discussion their binding affinities for various CD4bs
Monomeric gp120, when used as an immunogen ligands were still significantly lower than the
has failed to elicit broadly neutralizing full-length gp120 molecule. Additionally, the
antibodies against HIV-1 (40-45). This in part, is binding of ∆BS-ODEC with CD4 and bNAb b12
due to the very high conformational flexibility of was reduced significantly, due to the removal of
the gp120 molecule. In an effort to reduce the the bridging sheet. Hydrogen exchange mass
size of the immunogen and focus the immune spectrometric studies showed these molecules
response to a smaller number of epitopes, an are still highly flexible. To further reduce the
outer domain immunogen (ODEC) lacking the conformational flexibility of ODEC, cavity filling
flexible loops and the inner domain was and disulfide mutations were introduced. As
designed previously by us (7). It was expressed binding to CD4 normally leads to a large change
in E. coli to prevent glycosylation and increase in entropy in gp120, previous studies on full-
yield. The protein was found to bind CD4 and length gp120 had shown that filling in the Phe-
the neutralizing antibodies b12 weakly (KD of 3- 43 cavity using S375W/T257S mutations
12µM) but did not bind the non-neutralizing reduces this entropy change and thereby leads to
antibodies b6 and F105. The far-UV CD a higher affinity for CD4 (22,23). In this work,
spectrum of ODEC indicated that it had we tested these mutations in the ODEC
considerable secondary structure, though the background. An ODEC mutant with these two
large dip near 210nm also indicated that there cavity-filling mutations (ODECCF), when
was a substantial amount of an unfolded displayed on the yeast surface showed ~6 fold
population. Therefore, we searched for improvement in the VRC01 binding affinity. An
alternative ways of refolding ODEC. In the ODEC disulfide mutant (ODECSS) with an
current work, we modified the purification additional disulfide between NHB pairs 293 and
protocol for ODEC by including ArgHCl in the 448 was also designed and expressed on the
refolding buffer. ArgHCl is a well-known yeast surface. It showed a marginal
stabilizing osmolyte, known to help in the improvement in binding with VRC01 bNAb.
refolding of proteins by destabilizing expanded As all the above designs (∆BS-ODEC,
states like the unfolded state of a protein (46,47). ODECConsensus, ODECCF and ODECSS) showed
10Design of HIV-1 outer domain immunogens
some improvement in binding with VRC01 ODECCycV4 was the slowest to undergo
bNAb, we decided to combine these mutations exchange, indicating increased rigidity.
with the glycosylation-site mutations identified Broadly NAbs against HIV-1 show a very high
earlier by our group since these mutations also level of affinity maturation and it has been
improve binding for CD4bs ligands (18). The proposed that targeting the germline form of
first combined construct (ODECC1) had these bNAbs is a pre-requisite for generating
mutations at all the 14 outer-domain bNAbs (48-51). Full-length glycosylated gp120
glycosylation sites, cavity-filling mutations, an fails to engage the germline forms of bNAbs and
additional disulfide and was devoid of the is thus considered a poor immunogen for the
bridging sheet region. Another combination was activation of B-cells carrying germline variants
made (ODECC2) by the introduction of four of bNAbs. Apart from binding mature VRC01
glycosylation-site mutations, consensus with nM KD, ODECCycV4 also showed high-
mutations, cavity-filling mutations and a affinity binding (~10nM KD) with its germline
disulfide mutation in the ∆BS-ODEC background variant (GL-VRC01), indicating that it is a
(Table 1). Both these combined constructs, when promising immunogen. It is important to note
displayed on the yeast surface, bound bNAb that all our OD immunogens are produced from
VRC01 with very high-affinity (3-4nM KD). E.coli and are thus glycan free. All these OD
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
E.coli purified ODECC1 and ODECC2 showed a immunogens bind germline reverted VRC01
co-operative transition in urea mediated with very high-affinity. Even the original ODEC
denaturation with an apparent Cm of 4.2 M and immunogen without any additional stabilizing
3.26 M urea respectively. This is a significant mutation binds GL-VRC01 with ~4nM KD
improvement over ∆BS-ODEC and whereas full-length glycosylated gp120 does not
ODECConsensus immunogens, which show show measurable binding with GL-VRC01,
apparent Cm’s of 2.4 and 2.1 M urea indicating that glycosylation is one of the main
respectively. These results demonstrate that barriers in germline recognition of broadly
combining glycosylation site mutations with neutralizing antibodies. However, this
rational stabilizing mutations results in hypothesis needs to be tested for the germlines
significantly improved affinity and chemical of other CD4bs targeting bNAbs.
stability, even when the molecules are The OD designs with the best in vitro properties
bacterially expressed and lack glycans. were tested in a rabbit immunization study.
Designing circular permutants of proteins is an Following monomeric gp120 boosts, groups
attractive approach for protein design as it primed with ODECC2 and ODECCycV4 showed
allows manipulation of protein topology without the highest anti-gp120 titers relative to the rest
changing its amino acid composition or the of the molecules tested (ODECBS,
three-dimensional structure. In an alternate ODECConsensus and ODECC1).
attempt to stabilize the ODEC design, we made a The week 22 sera elicited by ODECC2 and
cyclic permutant of ODEC. E.coli expressed, ODECCycV4 immunization could neutralize Tier
cyclic-permutant ODECCycV4 protein bound 1 viruses from clade B as well as clade C with a
CD4, bNAb b12 and bNAb VRC01 with higher potency as compared to the ∆BS-ODEC
affinities comparable to that of full-length WT and ODECConsensus immunized groups boosted
gp120. It bound VRC01 with 16±5nM KD, while with monomeric gp120. Clade B viruses are
the original ODEC construct with native more prevalent in North America and Europe,
connectivities binds VRC01 with 359±54nM whereas Clade C viruses are more common in
KD. As we have not introduced any additional Africa and Asia. This cross-clade neutralization
stabilizing mutations in the ODECCycV4 elicited by our immunogens is encouraging since
constructs, it indicates that cyclic-permutation the week 22 sera from ODEC immunized animals
indeed helped in better structural organization of could not show any measurable neutralization.
the CD4bs. HDX-MS studies also showed that Our results indicate that improvement in stability
amongst all the ODEC variants tested, and affinity help in faster elicitation of anti-
11Design of HIV-1 outer domain immunogens
gp120 neutralizing antibodies. However, we VRC01 binder) and was subsequently boosted
could not observe any Tier 2 neutralization. with a series of different immunogens with a
We expect that the combination of ODECC2/ decreased affinity toward germline VRC01
ODECCycV4 prime and V1cycP gp120 boost, compared to mature VRC01. However, the sera
possibly coupled with a longer immunization could only neutralize viruses lacking glycans
protocol should further increase the near the CD4bs. Whether these germline VRC01
neutralization potency and breadth of the sera. A binders induce VRC01 lineage antibodies when
longer immunization study is required because IGHV1-2*02HC and VRC01LC are present at
anti HIV-1 broadly neutralizing antibodies physiological frequencies remains to be tested
typically take a long time to develop, as has been (61). Recently, an OD construct based on
observed in primate experiments (52) and subtype C strain 1084i lacking V3 loop regions,
patients (2,53,54) and also in one of our recent having stabilizing disulfides and a cavity filling
studies (55). mutation (OD3) was able to elicit a Tier 1 and a
Various studies attempting to focus the immune weak Tier 2 neutralizing response in guinea pig
response to the outer domain of HIV-1 Env have immunization studies (62). Our OD
been carried out previously. An OD construct immunogens also lack the V3 loop and the
(OD1) based on YU2 gp120 strain contained neutralization of very different HIV-1 viruses
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
residues 252-482, retained V1V2 and V3 (MN.3 and MW965.26) observed in this study is
variable loops, was glycosylated was expressed therefore unlikely to be due to V3 directed
in Drosophila S2 cells, but the sera from rabbit antibodies.
immunizations failed to neutralize homologous Till date, we have tested our OD immunogens
YU2 virus (56). Mice immunization studies with only in rabbits. However, germlines for most
clade-C OD as a fusion of human IgG1 Fc anti-HIV-1 antibodies are absent in rabbits. As
domain did not elicit any neutralizing response. our OD immunogens bind both mature and
(57,58). In another study two OD immunogens germline bNAbs with very high-affinity and
(monomeric and trimeric) based on group M elicit sera in rabbits with cross-clade
consensus sequence, were used to elicit neutralization, the improved OD immunogens
heterologous Tier 1 neutralizing responses can be tested in other animal models including
though higher neutralization titers for all viruses non-human primates for estimating their true
were seen with sera immunized with monomeric potential for neutralizing antibody generation.
gp120 (59). A mammalian cell expressed, Although rational modifications in ODEC
cyclically permutated OD derivative (eOD-Base) resulted in significant improvements in the
lacking glycans at N276 and N463 was evolved binding affinity and chemical stability, the
using Rosetta design followed by random HDX-MS studies show that there is only a
mutagenesis and yeast display library screening marginal improvement in the rigidity of the new
to form eOD-GT6 mutant, which bound GL- OD immunogens. Hence it is desirable to further
VRC01 with 44 nM KD, and several other GL stabilize these molecules using yeast surface
antibodies with high affinity while retaining display or other methodologies (63,64). Despite
binding with the mature bNAbs. A modified these lacunae, it is encouraging that OD
variant of eOD-GT6 (eOD-GT8 60 mer) (60) derivatives lacking variable loops can be
was used to immunize knock-in mice modified expressed and purified in reasonably high yield,
to express the germline-reverted heavy chain of very low cost, and that such molecules elicit
VRC01 (VH1-2*02) under the control of a antibodies that bind well to fully glycosylated
mouse VH promoter. eOD-GT8 60 mer gp120. In future studies we will combine the
immunized mice could activate VRC01 class best priming and boosting immunogens
precursors whereas BG505 trimers failed to do identified in the present work in an attempt to
so, possibly because these trimeric molecules do elicit sera with improved neutralization breadth
not bind germline variants of VRC01. In another and potency.
study, the immune responses were primed with
eOD-GT6 60mer (a high-affinity germline Materials and Methods
12Design of HIV-1 outer domain immunogens
5ml of Luria-Broth (LB). The primary culture
Construct descriptions. A summary of OD was grown overnight at 37°C. One litre of LB
construct details is provided in Table 1. The new was inoculated with 1% of the primary inoculum
OD immunogens are described in greater detail and grown at 37°C till an OD600 of ~0.6-0.8.
in the Results section. Cells were then induced with 1mM IPTG
(Isopropyl β-D-1-thiogalactopyranoside) and
Electrostatic calculations using Tanford- grown for another 8-12 hours at 37°C. Cells
Kirkwood model. An accessibility corrected were harvested at 3,500g and resuspended in
Tanford-Kirkwood model together with the 35ml phosphate buffered saline (PBS), pH 7.4
Bashford-Karplus approximation (TKBK) was containing 100µM PMSF and 0.2% Triton X-
used for calculating electrostatic interactions 100. The cell suspension was lysed by sonication
between charged residues in the immunogens on ice and centrifuged at 15,000g. The
(65). A program implementing the algorithm for supernatant was discarded, the pellet was
these calculations as described in Loladze et al. resuspended in 35ml 0.2% Triton X-100, PBS
(17) was kindly provided by Dr. JM Sanchez- (pH 7.4), sonicated again on ice and subjected to
Ruiz (Facultad de Ciencias, Departamento de centrifugation at 15,000g. The pellet was
Quimica Fisica, Spain). Classically, this theory solubilized in 40ml of 6M Guanidine
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
was developed for calculating charge-charge hydrochloride (GdnCl) in PBS (pH 7.4)
interactions assuming a spherical shape for any overnight at room temperature. The solution was
protein, and ionizable amino acids were centrifuged at 15,000g for 30 min. The
represented by point charges smeared over the supernatant was bound to 3ml Ni-NTA beads
spherical surface of the protein. All globular (GE Healthcare), washed twice with 25ml of
proteins were assumed to have an average 50mM imidazole containing 6M GdnCl in PBS
specific volume of 0.72ml/g. Hence, the radius (pH 7.4), and finally denatured protein was
of any protein sphere can be calculated based on eluted with 6M GdnCl in PBS (pH 7.4)
the molecular weight of the protein. The solvent containing 500mM imidazole at room
was assumed to be a continuum with a high temperature.
dielectric constant of 78 and protein had a low The first four eluted fractions (each 5ml) were
dielectric constant of 2 or 4. Subsequently, when pooled together and then rapidly diluted 10 fold
3D structures of proteins became available, the with PBS containing 1mM EDTA to reduce the
interaction energy between a pair of amino acids denaturant concentration from 6M to 0.6M. The
was corrected for by their surface accessibility in resulting solution was again concentrated back
the 3D structure, as the energy for a pair of to the original volume in an Amicon
residues on the surface of a protein would be concentrator. This was followed by desalting
lower relative to that at a buried position due to into PBS (pH 7.4) containing 1mM EDTA using
dielectric screening by the solvent (14). A a HiTrap desalting column (Amersham) to
positive value for the charge-charge interaction remove the remaining denaturant. The desalted
energy (E-int) indicates unfavourable interaction protein was concentrated to a final concentration
and vice versa. of 0.5mg/ml, flash-frozen in liquid nitrogen and
Cloning, expression and protein purification. stored in aliquots at -80°C. The yield was
The E.coli, codon-optimized gene sequences of determined by densitometry analysis from SDS-
our designed immunogens were synthesized PAGE using standard proteins of known
(GenScript, USA) and cloned into the concentrations.
pET28a(+) vector (Novagen) between the NdeI This purification protocol was further modified
and BamHI sites with an N-terminal His tag. to include arginine-hydrochloride (ArgHCl) in
The proteins were over-expressed in E.coli the refolding buffer. Briefly, the first three
BL21(DE3) cells and purified from the insoluble elution fractions (5ml each) were pooled
fraction of the cell culture lysate. Briefly, a together and rapidly diluted 10 fold with 0.5M
single colony of E.coli BL21(DE3) transformed ArgHCl (SRL, 99% purity) in PBS containing
with the plasmid of interest was inoculated into 1mM EDTA at 4°C to reduce the denaturant
13Design of HIV-1 outer domain immunogens
concentration from 6M to 0.6M. The resulting
solution was again concentrated back to the Fluorescence spectroscopy. All fluorescence
original volume in an Amicon concentrator. The spectra were recorded at 25°C on a JASCO FP-
concentrated solutions from the first three 6300 spectrofluorimeter. The concentration of
elution fractions were combined (total protein used was 1M either in PBS, pH 7.4 or
15ml),dialyzed once against one litre of 0.5M in the presence of 6M GdnCl in PBS, pH 7.4.
ArgHCl in PBS containing 1mM EDTA at 4°C The excitation wavelength was fixed at 280 nm
to remove the denaturant (GdnCl), and then and emission was recorded from 300 to 400 nm.
dialyzed extensively (3 times) against PBS, The excitation and emission slit widths were 3
1mM EDTA (using two litres of buffer each and 5 nm, respectively. Each spectrum was an
time) at 4°C to remove ArgHCl. The dialyzed average of three consecutive scans. Buffer
protein was concentrated to a final concentration spectra were also acquired under similar
of 0.5mg/ml, and flash-frozen in liquid nitrogen conditions and subtracted.
and stored in aliquots at -80°C. Protein yield was
determined by densitometry analysis from SDS- Isothermal urea denaturation. Isothermal urea
PAGE using standard proteins of known denaturation studies were carried out for all the
concentrations. All proteins in this study were proteins by monitoring their tryptophan
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
purified using a similar protocol and their fluorescence as a function of urea concentration
identity was confirmed by ESI-MS. as described previously (67). The final
concentration of protein used was ~1-3μM in
Far UV circular dichroism (CD). Far UV-CD PBS buffer (pH 7.4). Proteins were incubated
spectra were recorded on a Jasco J-715C with varying concentrations of urea (0-6M) until
spectropolarimeter flushed with nitrogen gas. equilibria were established. Urea concentrations
The concentration of all protein samples was ~5- were estimated by measurement of refractive
10M and buffer used was PBS, pH 7.4. indices of various urea dilutions. The
Measurements were recorded at 25°C in a 1mm denaturation was monitored at the wavelength
path length quartz cuvette with a scan rate of maximum of the difference spectra of native (in
50nm/min, a response time of 4 seconds and a 0 M urea) and denatured protein (in 6 M urea).
bandwidth of 2nm. Each spectrum was an This was typically in the range of 320-325 nm.
average of three scans. Mean residue ellipticities Fluorescence spectra were acquired on a JASCO
(MREs) were calculated as described previously FP-6300 spectrofluorimeter with an excitation
(66). Buffer spectra were also acquired under wavelength at 280 nm. The excitation and
similar conditions and subtracted from protein emission slit widths were 3 and 5 nm,
spectra, before analysis. The experimental data respectively. The data obtained was fit to a two-
plotting and fitting were done using state model as described previously (67,68).
SigmaPlot™ for Windows™ scientific graphing Experimental data plotting and fitting were done
software. using SigmaPlot™ for Windows™ scientific
graphing software.
Near UV-CD. Near UV-CD (tertiary CD)
spectra were recorded on a Jasco J-715C HPLC. 20g of the native protein in PBS, pH
spectropolarimeter flushed with nitrogen gas. 7.4 was injected into a Discovery C5 analytical
Measurements were done over a wavelength column (150 Х 4.6mm, 5m particle size)
range of 250 to 300 nm at 25C with 10M to (Supelco) while for reduced samples, the 20g
80M of protein in PBS buffer, pH 7.4 with a protein was incubated with 4M GdnCl in PBS,
1cm path length cuvette, a scan-rate of pH 7.4 and 5mM DTT at 37C, prior to
10nm/min, a response time of 8 seconds and a injection. Proteins were eluted with a gradient of
bandwidth of 2nm. Data reported were averaged 5 to 95% acetonitrile containing 0.1% formic
over 5 scans and were corrected for buffer acid at a flow rate of 2% per minute.
signals.
14Design of HIV-1 outer domain immunogens
80°C respectively. The data were collected in
Binding affinity measurements using Surface positive ion mode.
Plasmon Resonance (SPR). All SPR One spectrum was collected per second. Spectra
experiments were performed with a Biacore were combined in the MassLynx software and
2000 (Biacore, Uppsala, Sweden) optical processed using the following parameters: i)
biosensor at 25°C. 800-900 resonance units Background subtraction: polynomial order: 20;
(RU) 4-domain CD4, b12, VRC01, b6 or F105 below curve: 40%. ii) Smoothing: smooth
was attached by standard amine coupling to the windows (channels): 40; the number of smooths:
surface of a research-grade CM5 chip. A sensor 2; smooth mode: Savitzky Golay. iii) Centroid:
surface (without CD4 or an antibody) that had minimum peak width at half height: 2; centroid
been activated and deactivated served as a mode: top 80%. Following this, the spectrum
negative control for each binding interaction. was deconvoluted either automatically or
Different concentrations of analytes were passed manually by choosing two consecutive peaks at
across each sensor surface in a running buffer of a time. For deuterated samples, a high standard
PBS, pH 7.4 containing 0.005% P20 surfactant. deviation of 2-3 Da was usually obtained in the
Both binding and dissociation were measured at deconvoluted mass due to heterogeneity in the
a flow rate of 30 l/min. In all cases, the sensor amount of deuterium incorporated in different
Downloaded from http://www.jbc.org/ by guest on October 12, 2018
surface was regenerated between binding molecules in the mixture, hence the masses of
reactions by one to two washes with 4M MgCl2 individual charge states were monitored. The
for 30 seconds at 30 l/min. Each binding curve difference in mass of the protein obtained in the
was corrected for nonspecific binding by protonated solvent, as compared to that obtained
subtraction of the signal obtained from the after incubation in D2O was calculated for
negative-control flow cell. The kinetic different time points.
parameters were obtained by fitting the data to
the simple 1:1 Langmuir interaction model using Cloning of OD immunogens into a yeast
BIA EVALUATION 3.1 software. surface display vector. Genes for various OD
immunogens (ODEC, ∆BS-ODEC,
Hydrogen Deuterium Exchange Mass ODECConsensus, ODECCF, ODECSS, ODECC1
spectrometry (HDX-MS). For HDX-MS and ODECC2) were synthesized by GenScript
experiments, 50 picomoles of ODEC or an ODEC (USA). These gene products were PCR
variant protein (1.2µg in 3µl) at 20°C was amplified with pPNLS specific primers and
diluted 20 times in D2O (Sigma) to a final individually recombined into SfiI digested
volume of 100µl and incubated for varying linearized pPNLS vector by homologous
amounts of time. The pH of the resulting recombination in the EBY100 strain of S.
solution was ~6.8. The reaction was quenched cerevisiae (24,26,69).
by addition of 2µl of 10% formic acid to a final
concentration of 0.2% (pH 2.3) and immediately Yeast surface display of OD variants. pPNLS
used for LC ESI-MS. A Nano Acquity UPLC vector contains an AGA2p fusion at the N-
system from WATERS was used. The auxiliary terminus of the surface displayed protein along
solvent (which pushes the sample from the loop with two epitope tags, HA (YPYDVPDYA) and
into the trap column) used was 0.1% formic acid c-Myc (EQKLISEEDL) for detection (69). All
in water, while the binary solvents were water constructs were displayed on the cell surface of
and acetonitrile, each containing 0.1% formic S. cerevisiae strain EBY100 using a standard
acid. The sample was typically desalted using a protocol (26). Briefly, EBY100 cells were
3-minute acetonitrile gradient from 3-97%. For transformed with pPNLS plasmids containing
acquiring mass spectra, a WATERS Q-TOF genes for OD immunogens, and colonies were
mass spectrometer was used. The capillary grown in glucose-containing liquid SDCAA
voltage was set to 3kV, while the desolvation media (pH 4.0) till mid-log phase at 30°C,
and source temperatures were set to 200°C and followed by induction in galactose-containing
15You can also read

















































