Copper & biological health - Review Article
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Review Article
Indian J Med Res 128, October 2008, pp 448-461
Copper & biological health
S. Krupanidhi, Arun Sreekumar* & C.B. Sanjeevi **
Department of Biosciences, Sri Sathya Sai University, Prasanthi Nilayam, India; *Michigan Center for
Translational Pathology & Department of Pathology, University of Michigan, Ann Arbor, MI, USA &
**
Center for Molecular Medicine, Department of Molecular Medicine & Surgery, Karolinska Hospital,
Stockholm, Sweden
Received January 28, 2008
Cu being a transition metal is ubiquitously engaged in biological systems to derive electrons through
its participation in several enzymatic reactions. Upon bestowing the significance of Cu in biological
systems, an elaborate mechanism is set forth by nature for maintaining Cu homeostasis. As a
consequence, a wide variety of proteins viz., family of Cu bearing proteins, cuproenzymes, Cu
transporters and Cu chaperone proteins have been manifested for enabling Cu to show its relevance
in biological health. In addition, understanding the role of Cu in hepatic and neuronal functions
and also in angiogenesis keeps progressing with the advent of novel molecular tools. The studies on
genetic defects in Cu metabolism causing abnormalities are providing insights leading to the possible
prognostic cues to alleviate the sufferings.
Key words Ceruloplasmin - Cu carriers - Cu chaperones - Cu chelators - Cu metabolism - Cu transporters - Menkes disease -
oxygen binding Cu proteins - Wilson’s disease
Biological relevance of Cu respiration, iron metabolism, biosynthesis of
neurotransmitter, and free radical detoxification.
Cu is an essential micronutrient required by all life
Therefore, it is worth recalling that Cu is vital for normal
forms. Cu is a transition metal and hence involved in a
healthy functioning of organisms (Fig. 1).
variety of biological processes viz., embryonic
development, mitochondrial respiration, regulation of Source of copper: Rich amounts of copper along with
hemoglobin levels as well as hepatocyte and neuronal other essential elements found in the soil are taken up
functions. Being a transition metal, Cu gets biologically by plants using very elaborate transportation machinery.
converted between different redox states namely Plants, thus serve as a direct source of elemental copper
oxidized Cu (II) and reduced Cu (I). This unique for higher organisms. Additionally, human breast milk
attribute has made Cu metal to get manifested as an has the highest concentration of Cu (0.25 to 6.0 mg/l).
important catalytic co-factor for a variety of metabolic For its effective utilization, the elemental copper derived
reactions in biological systems. Several reviews 1-5 from these sources needs to be absorbed and transported
highlighted the participation of Cu in a myriad cellular to metabolically active sites. This process, termed
activities and physiological processes such as cellular bioavailability, is possibly regulated by four essential
448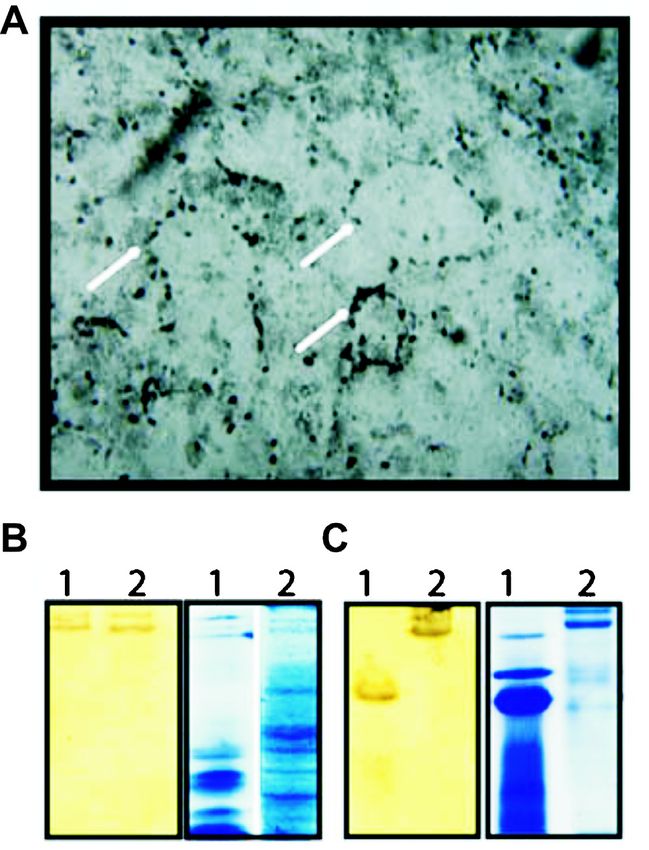
KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 449
molybdenum competitively inhibits intestinal Cu uptake.
On the contrary, various derivatives of copper such as
chlorides, acetates, sulfates and carbonates enhance its
bioavailability in higher organisms.
Plants as bioindicators and hyperaccumulators of Cu:
A few plants uniquely accumulate copper from their
habitats viz., soil or water. They are: Aeolanthus
biformifolius, Athyrium yokoscense, Azolla filiculoides,
Bacopa monnieri, Brassica juncea L., Callisneria
Americana, Eichhornia crassipes, Haumaniustrum
robertii, Helianthus annuus, Larrea tridentate, Lemna
minor, Pistia stratiotes and Thlaspi caerulescens. The
metal molybdenum is also accumulated by Thlaspi
caerulescens (Brassica). Sheep feeding on Thlaspi
caerulescens possibly face the deficiency of copper as
Fig. 1. Influence of Cu deficiency or excess on the response of this specific plant is also a hyperaccumulator of
organisms. (1) physiological efficiency, (2) tolerable levels with
inbuilt compensatory mechanisms, (3) therapeutic symptoms
molybdenum which inhibits intestinal copper uptake.
leading to fatal diseases. Copper transport and utilization
Dietary copper, absorbed in the stomach and upper
attributes as defined by Raul6. These include (i) quantum intestinal tract, reaches liver as a complex with serum
of intake; (ii) dependent variability; (iii) linearity proteins viz., albumin or transcuperin or the amino acid
between dose and response; and (iv) slope ratio analysis. histidine10. Importantly, liver is the major store house
Solubility of Cu in water or physiological fluids is a for intracellular copper11. Here, copper is reduced to
good indicator of bioavailability and digestibility. cupric state and transported across plasma membrane
Additionally, copper complexes with various by CTR1 transporters as described later. Importantly,
biomolecules, thus, facilitating its utilization. These as highlighted later in this review, intracellular copper
include complex of copper with lectins and needs to be maintained in a complex state so as to
glycoproteins as seen in grains or with amino acids as prevent the oxidative damage caused by free copper to
noticed in higher organisms including mammals. In fact, DNA, proteins and membrane components10. Hence,
amino acids exert a critical role in uptake of copper by copper transport and utilization involves a complex
the intestinal membranes. Further, among the essential interplay between transporters and binding proteins/
amino acids, methionine in the diet enhances Cu chaperones. Additionally, Cu plays a vital role as a
absorption by at least 2-fold. On the contrary, by the side catalytic co-factor for a variety of metalloenzymes.
of cysteine due to its ability to chelate by the side of Keeping the importance of cupric Cu in biological
copper coupled to its ability to potentially reduce copper function, an elaborate mechanism is set forth by Nature
to a monovalent state 7, leads to a reduction in its for maintaining Cu homeostasis, which includes a wide
bioavailability. On a similar note, tripeptide of glutathione array of proteins namely (i) family of Cu bearing
has significant post-absorptive importance in Cu proteins, (ii) cuproenzymes, (iii) Cu transporters and
transport. Glutathione forms an intermediary complex (iv) Cu chaperone proteins. It is not surprising for the
with Cu in the enterocytes before transferring the metal redundant machinery that Cu is enjoying out of several
to other target proteins- viz., superoxide dismutase or heavy metals for its transport and participation in
ceruloplamin (CP), etc., thus facilitating its assimilation. cellular metabolism, which guarantees the survival of
This ability of copper to complex with amino acids or living organisms as conditioned by the strategies and
organic acids is extensively exploited in animal nutrition mechanisms of the evolution of metallic proteins.
experiments. As an example, Cu-lysine complex has been
shown to be effective as a supplement in feed for chicks The family of Cu bearing proteins plays a significant
than for lambs8. Similarly, formulations of proteins with role in metal detoxification and keeps the Cu in non ionic
minerals, also termed proteinates, have been found to be curpric state. They are metallothioneins, prion protein,
highly effective as feed for growing calves in areas with albumin, transcuperin, CP, phycocyanins of blue green
high molybdenum contents in foliage 9. Notably, algae and haemocyanins of blue blooded organisms.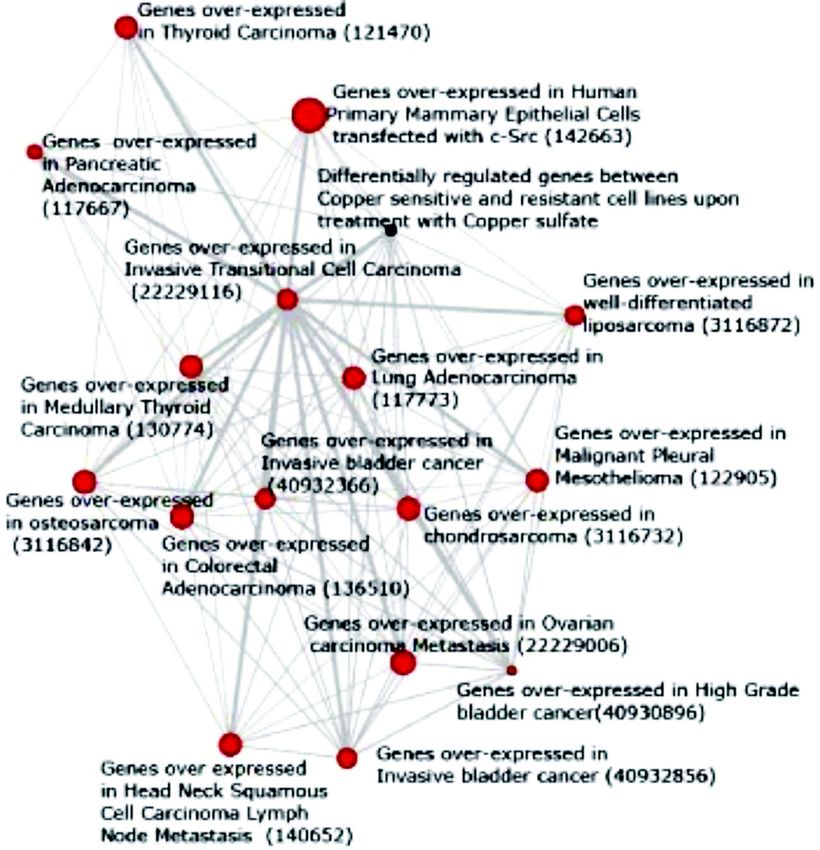
450 INDIAN J MED RES, OCTOBER 2008
Blue blooded organisms: An interesting copper binding
protein found in some of the lower eukaryotes is
hemocyanin (Hcy). Fig. 2 shows the UV spectrum of
Oxy-Hemocyanin (Oxy-Hcy) with a characteristic
absorbance at 340 nm revealing the presence of copper-
oxygen complex. Hcys are found in a majority of
arthropods and mollusks, and they are called “Blue
Blooded Organisms” by virtue of the fact that their blood
turns blue in color upon oxygenation. Importantly, in
these organisms, hemocyanin associated with blood (also
called hemolymph) serves as primary carrier of oxygen.
Hcy turns blue upon binding molecular oxygen, a
phenomenon that is readily reversible. Notably, such
binding occurs at high partial pressure of oxygen which
converts Hcy to Oxy-Hcy. The latter dissociates to release Fig. 2. The UV visible spectrum of blue blood of Pila globosa
molecular oxygen at the vicinity of tissues that have low containing hemocyanin revealing the presence of broad peak at
340 nm due to CU-O complex.
oxygen pressure, thus functioning as a mode for oxygen
transport12 (Fig. 3). % Oxygen saturation
Characteristically, Hcy is non-cellular and found
freely dissolved in haemolymph. By virtue of its large
molecular size with multiple epitopes, Hcy is a potent
immunogen as evidenced by the development of discrete
crescentic arcs in Ouchterlony double immunodiffusion
assay upon antibody challenge (Fig. 4).
As revealed in Figs. 5B and C the Hcy from
hemolymph of fresh water field crab and Indian apple
snail Pila, showed positive staining with rubeanic acid
stain (a stain to detect copper binding proteins),
confirming its ability to bind copper13. Additionally, the
detection of copper granules by the histochemical Partial pressure of oxygen
staining of hepato-pancreas in the pulmonate garden (mm Hg)
snail, Cryptozona ligulata, potentially reveals the
Fig. 3. Typical sigmoid oxygen equilibrium curve of hemocyanin.
existence of a copper store, probably complexed with The P50 values indicating the affinity of hcy towards oxygen are
metallothioneins, that could be possibly recruited for influenced by the factors shown in the diagram. DPG,
Hcy biosynthesis (Fig. 5A)14,15. Structurally, molluscan diphosphoglycerate.
haemocyanins are composed of multiple subunits (eight)
that result from duplications in the gene encoding for
the protein (Fig. 6). These subunits assemble into a
quaternary folded architecture with 160 oxygen binding
sites in the native protein16. This is in contrast to the
Hcy from arthropods, that have only 3 subunits that are
folded up to generate 48 oxygen binding sites 3 .
Importantly, in all these cases, each of the oxygen
binding sites contains 2-Cu atoms and each of the Cu
atoms anchors to 3 histidine residues. Further, the two
molecules of copper are bridged together by 2 molecules
of oxygen resulting in the formation of a dioxygen
bridge. Thus on the whole, hcy derived from molluscs
and arthropods contain 320 and 96 copper atoms Fig. 4. Immunocrescentric arcs developed by hemocyanin of Pila
respectively. globosa along with its antibodies harvested in mouse.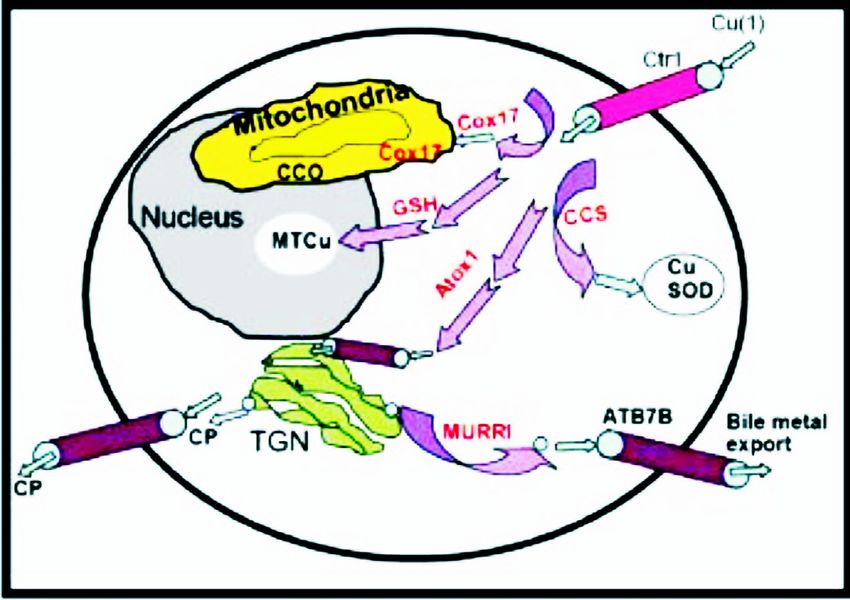
KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 451
Phenoloxidase is another such copper binding
protein that binds to dioxygen with a different
physiological function viz., browning of fruits and
vegetables in plants as well as wound healing, skin
pigmentation etc in higher organisms. Also, it has a role
during sclerotization of new exoskeleton in molting
insects. This contrasting physiological role for
phenoloxidase compared to Hcy could be attributed to
the ability of the former to trigger the catecholase
activity3.
Transporter proteins: In higher organisms and plants,
principal copper binding proteins belonging to the
family of P-type ATPases serve the function of
intracellular copper transport. Included among these are
the two proteins ATP 7A and ATP 7B. Interestingly, the
presence of such a network for Cu transport seems to
be evolutionarily conserved. Notably, prokaryotes
possess metal transporting enzymes also termed heavy
metal ATPases that protect them from stress caused by
heavy metals found in their natural environment. These
according to Nigel et al17 are encoded by the structural
genes (cutA, cut-B…cut-F) and the regulatory protein,
cutR. Additionally, some mutant forms of E.coli harbor
a plasmid borne version of copper resistance genes also
called pco that confers resistance to approximately five
Fig. 5. A. Cu granules (possibly prohcy) in the hepatopancreas of fold higher concentrations of cupric ions than wild type
a pulmonate snail, Cryptozona ligulata stained with rubeanic acid. strains18. Significantly, bacteria endowed with such
Rubeanic acid stains copper binding proteins B. Native PAGE of
heavy metal transporter proteins are now being
the proteins of hepatopancreas of fresh water field crab (1) and
Indian apple snail, Pila (2) stained with rubeanic acid and coomasie exploited commercially in a process termed
blue. C. Native PAGE of the blood proteins of crab (1) and Pila “Bioleaching”. The latter is an environmentally friendly
(2) stained similarly. process for metal recovery which is a cost-effective
process for treating ores that are remote and difficult to
access. One of the most exploited microorganisms in
bioleaching is Acidithiobacillus ferrooxidans.
As mentioned above, a similar paradigm consisting
of metal transporters have been described to be existent
in plants, specifically in edible portions such as seeds19.
Notably, Cu plays a vital role in the physiology of plants
viz., respiration and photosynthesis. Further,
photosynthetically active cells require more Cu than
other cells. Two families of Cu transporter proteins have
been recognized among plants19. Among these, P-type
ATPases (PAA) belong to the family called heavy metal
ATPases (HMA). In Arabidopsis, they function to
transport Cu to the stroma of the chloroplast, where
they play a critical role in maintaining copper
homeostasis. Importantly, mutation in these ATPases
Fig. 6. Alkaline dissociation of haemocyanin (H) of Pila globosa affects the photosynthetic electron transport, which can
showing 8 subunits at 8.5 pH shown on Native PAGE gel performed be reversed by addition of Cu. Additionally, these are
using PHAST system. M, molecular weight marker. also involved in the transport of Cu in roots and flowers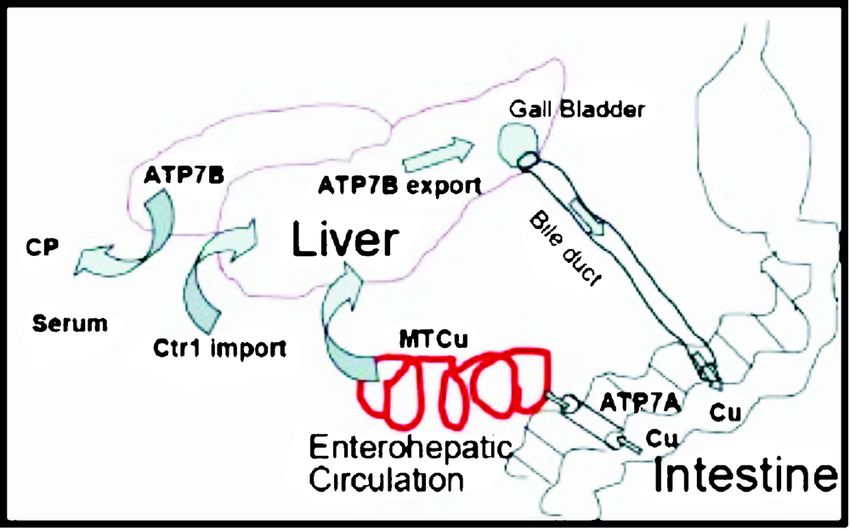
452 INDIAN J MED RES, OCTOBER 2008
of plants. The latter is supported by the detection of
transcripts for HMA in these sites19,20. The second family
of Cu transporters viz., COPT (Cu transporters) are also
identified in plants. The homologous transporter
proteins of the same have been reported in yeast and
mammals20. Notably, Arabidopsis exposed to decreased
levels of copper for a period of 18 h was shown to turn
on a compensatory mechanism that involved increased
synthesis of COPT mRNA21. In addition, the phenotypic
manifestation of reduced copper levels in these plants
involved an increase in root length which could be
reversed by the addition of Cu19. Further, the importance
of COPT1 knockdown using an anti-sense strategy
Fig. 7. Enterohepatic uptake, distribution and excretion of Cu.
resulted in an increased frequency of pollen
abnormalities even though the experimental plants were
grown under standard nutrient conditions. The latter transcripts are seen at high levels in muscle, kidney,
phenotype was rescued by exogenous addition of Cu lung and brain and low levels in placenta and pancreas,
highlighting the importance of this element for the while liver contains only trace amounts of this
developing pollen19. In addition to transporters, plants transporter22-24. ATP7A regulates Cu- efflux when the
also contain a class of molecules termed the metallo- levels of the latter become high in epithelial cells. In
chaperones that bind metals and facilitate their transport contrast, ATP7B expression is higher in the liver where
to target proteins/sites. The expression of such Cu it regulates the release of copper into bile. Importantly,
chaperone mRNA is ubiquitously seen in the tissues of the transcript levels of both these transporters are
root, stem, leaf and inflorescence indicating its role as positively regulated by intracellular levels of copper.
intercellular Cu delivery and recycling. One such copper Further, Cu ATPases also appear in the placenta and
chaperone seen in plants is cytochrome oxidase 17 lactating breast tissue for transporting Cu to fetus and
(COX17). Defects in COX17 lead to the respiratory through milk to neo-nates respectively. ATP7A have
deficiency due to the failure of protein to deliver Cu to been reported to be present within syncytiotrophoblasts,
mitochondrial cytochrome oxidase complex. cytotrophoblasts and fetal vascular endothelial cells.
Interestingly, the various metal transporter proteins like This is consistent with their role in the transport of Cu
PAA, HMA, COPT, CCH, COX17, etc., form potential from these tissues into the fetal circulation25. In contrast
targets that could be manipulated to enhance mineral ATP7B has been shown to facilitate the export of Cu
deposits in plants that could possibly alleviate mineral from the placenta to the maternal tissues, a mechanism
deficiency in humans and live stock. that protects excessive copper from reaching the
developing fetus. Additionally, ATP7A is also reported
In higher organisms, the absorbed dietary Cu enters to be expressed in luminal epithelial cells of alveoli
liver through entero-hepatic circulation and is further and ducts of breast tissue, with its expression levels
transported as a complex with CP or excreted into bile, being positively regulated by lactation26. Further, Cu
a process facilitated by a number of Cu chaperones and ATPase activity is also seen in the central nervous
transporting proteins (Fig. 7). ATP7A (MND) and system where both ATP7A and ATP7B regulate neuronal
ATP7B (WND) are also the principal Cu transporters Cu homeostasis. Also, both these are expressed within
in higher eukaryotes. These transporters contain 8- retinal pigment epithelium where they regulate the
transmembrane domains in addition to six Cu-binding release of CP that in turn maintains iron homeostasis.
motifs at the N-terminus (MXCXXC; M=methionine,
C=Cysteine, X=any amino acid). These Cu binding An alternate class of copper transporters found in
motifs of ATPases reveal that Cu ions are typically yeast are, Ctr1 that regulate the influx of copper into
bound to sulphur containing amino acids. They function the cytoplasm. These transporters contain three
like cation exchangers and use energy from ATP transmembrane domains with methionine rich
hydrolysis to translocate metal cations across lipid extracellular motifs at the N-terminal that bind copper
bilayers. Both ATP7A and ATP7B are predominantly and enable its import27. Importantly, dietary Cu (CuII)
localized in the transgolgi-network (TGN) and involved needs to be converted to its reduced form (CuI) prior to
in the delivery of Cu into nascent cuproproteins. ATP7A its transport by the Ctrls. The process of biochemical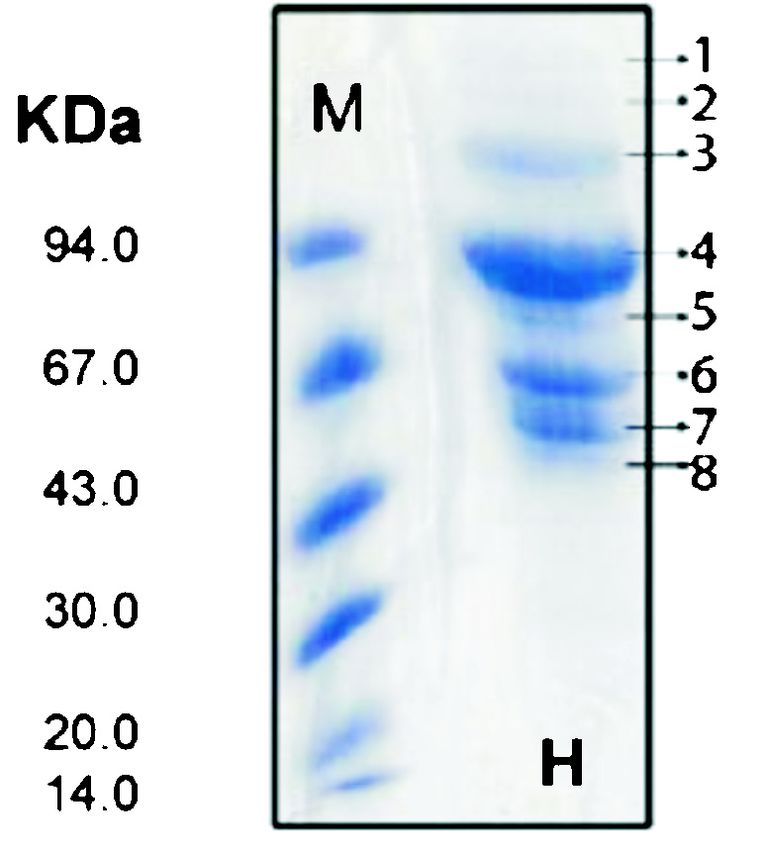
KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 453
reduction is carried out by various plasma-associated neurodegenerative disorders that includes Creutzfeld-
reductases. Jakob disease. In these disorders, the ability of the
mutant prion to transport copper is significantly
The mammalian homologue of yeast Ctr1 is called
impaired making neuronal cells susceptible to oxidative
MURR1; while it’s human orthologue is named CTR1/
stress24. Also, tripeptide glutathione (GSH) binds copper
2. MURR1 is a recently discovered protein chaperone
and enables its transport across the blood-brain barrier.
whose absence has been shown to cause Cu toxicosis,
potentially due to hepatic Cu overload (Fig. 8). The latter Importantly, in serum, most of the copper is
has been documented in Bedlington terriers with transported by CP. It is synthesized by both hepatocytes
MURR1 gene mutation that is characterized by deletion and activated macrophages. It is a 132 KDa monomer.
of exon 2, resulting in complete absence of the In addition to serving as a prime transporter of copper
functional protein product in liver of affected animals28. in serum, CP also plays a major role in intestinal
Importantly, these terriers exhibit elevated levels of absorption of iron. Significantly, in its role as a carrier
lysosomal Cu content and pronounced reduction in of copper in serum, each molecule of CP can bind to
bilary Cu excretion29. This is suggestive of cooperativity seven molecules of copper. Notably, elevated plasma
between MURR1 and ATP7B, to mediate excretion of CP has been shown to have adverse effects on
excess copper into bile. Further, imported intracellular cardiovascular system.
copper has been shown to bind Atox1, which then
Transcriptional regulation of proteins involved in
transfers the metal to its docking partners in the
Cu translocation
secretory pathway (Fig. 8). This reveals a potential role
of Atox1 in the ATP7B-mediated bilary excretion of Prokaryotic Cu homeostatic system has been well
excess Cu. Atox1 has also been implicated in mediating characterized in Enterococcus hirae 31. Four genes
copper transfer to CP and tyrosinase. The latter which (copY, copZ, copA and copB) are reported to be
is a critical step in melanin generation is supported by arranged in the cop operon of E.hirae. CopA and copB
the observation wherein Atox1-null mice have been encodes for Cu transporting P-type ATPases which are
shown to exhibit hypo-pigmentation30. highly conserved, stabilized and possibly extended into
eukaryotes. CopY encodes for Cu responsive repressor
An alternate mode of copper entry involving
and copZ encodes for a chaperone protein. The cop
endocytosis is facilitated by a class of proteins termed
operon allows growth of E. hirae in Cu-limiting
prion protein (PrPC). These are glycoproteins that are
conditions (up to 8 mM Cu). CopA ATPases take Cu
expressed on the plasma membrane. By virtue of its
while it is limiting and copB ATPases bale out excess
expression in the central nervous system as well as
Cu. CopY regulates the expression of cop operon and
peripheral tissues, mutation in PrPC lead to a number of
copZ translocates Cu intracellularly32,33. CopY is a Zn
containing homodimeric repressor that binds to the
promoter region of the cop operon, thereby regulating
the synthesis of ATPases and chaperones. It is reported
that copY is dimeric and belongs to winged-helix type
repressor34. Thus, initially, the package of molecular
machinery for the regulation of heavy metal ions gained
relevance in the survival of bacteria and hence it would
not be a surprise for the eukaryotes to adopt them. The
expression of the cop operon is low in standard growth
media whereas induced by 50 fold upon exposure of
bacteria to extracellular Cu35. CopY repressor binds to
the consensus binding site TACANNTGTA, called ‘cop
box’36. Experimentally induced mutation in cop-box
prevented its interaction with the repressor. The kinetics
Fig. 8. Pictorial illustration of copper transport in liver. Cu chaperones of the interaction between the repressor and promoter
(violet), storing site (MT), distribution (CCO, CuSOD, TGN), of cop operon in E. hirae are elaborated by David
secretion of CP and excretion of copper from a hepatocyte are shown. Magnani and Marc Solioz 35. The induction of cop
Ctr1 and ATP7B are Cu transporters. The abbreviations shown in
operon is facilitated by excess Cu which makes the
the figure are cited in the text.
repressor (CopY) to dissociate from the cop box. This
454 INDIAN J MED RES, OCTOBER 2008
E. hirae model has yielded an insight into possible Table. Classes of copper binding proteins defined by InterPro (http:/
existence of a similar molecular architecture in /www.ebi.ac.uk/interpro)
eukaryotes. Group 1: Copper type II, ascorbate-dependent monooxygenase
Copper-complexes…. A necessity for cellular 1. DBH, Dopamine beta-hydroxylase (dopamine beta-
monooxygenase)
function 2. MOXD1, Monooxygenase, dbh-like 1
In addition to being transported, intracellular copper 3. PAM, Peptidylglycine alpha-amidating monooxygenase
Group 2: Di-copper centre-containing
has to be sustained in a complexed configuration in 1. DCT, Dopachrome tautomerase
order to prevent its deleterious effects. The latter, 2. TYR, Tyrosinase
possibly are due to the generation of hydroxyl free 3. TYRP1, Tyrosinase-related protein 1
radicals by chemical reaction of monomeric copper with Group 3: Copper amine oxidase
hydrogen peroxide. Thus, elemental copper that is 1. ABP1, Amiloride binding protein 1 [amine oxidase
(copper containing)]
trafficked into cells is kept in bound state by a group of 2. AOC2, Amine oxidase, copper containing 2 (retina
copper binding proteins or chaperon proteins (Fig. 8). specific)
These include Atox1 (antioxidant protein), CCS (Cu 3. RHBDF1, Rhomboid 5 homolog 1 (drosophila)
chaperone for SOD), COX17, MT1, MT2 4. AOC3, Amine oxidase, copper containing 3 (vascular
(metallothionein) and APP (amyloid precursor protein). adhesion protein 1)
Group 4: Multicopper oxidase, type 1
In order to understand the biological processes 1. CP, Ceruloplasmin (ferroxidase)
regulated by copper binding proteins, we adopted an 2. F5, Coagulation factor V (proaccelerin, labile factor)
3. F8, Coagulation factor Viii, procoagulant component
enrichment strategy. Firstly, all proteins having either a (haemophilia a)
copper binding domain/functional site were culled from 4. HEPH, Hephaestin
the InterPro database (http://www.ebi.ac.uk/interpro). Group 5: Copper/Zinc superoxide dismutase
This resulted in a total of 36 proteins that were distributed 1. CR1, Complement component (3b/4b) receptor 1 (knops
across 7 groups based on function/domains/functional blood group)
2. PSORS1C1, Psoriasis susceptibility candidate 1
sites (Table). Each group included 3-12 proteins. Proteins 3. KIAA0467, Kiaa0467
from all groups were then used for enrichment analyses 4. SOD1, Superoxide dismutase 1, soluble (amyotrophic
using a bioinformatics tool called Oncomine Concept lateral sclerosis 1)
Maps (OCM) (www.oncomine.org), developed by Daniel 5. SOD3, Superoxide dismutase 3, extracellular
Rhodes and colleagues37,38. OCM, is an enrichment tool, 6. CCS, Copper chaperone for superoxide dismutase
Group 6: Blue (type 1) copper domain
that allows to systematically linking groups of protein/ 1. NR1H3, Nuclear receptor subfamily 1
genes that have a common biological nuance to various 2. LYST, Lysosomal trafficking regulator
molecular concepts thus generating novel hypothesis. 3. FLJ25006, hypothetical protein flj25006
Notably, we believe that such an enrichment analysis of 4. APR-2, Apoptosis related protein
copper binding proteins could potentially reveal various 5. SLC35B2, Solute carrier family 35, member b2
6. IGHG4, Immunoglobulin heavy constant gamma 4 (g4m
cellular processes that could be initiated by their action. marker)
The various molecular concepts that were used in this 7. SIRT7, Sirtuin (silent mating type information regulation
enrichment analyses were derived from both gene and 2 homolog) 7 (S. cerevisiae)
protein annotations from external databases, and 8. CCDC14, Coiled-coil domain containing 14
computationally-derived regulatory networks. The Group 7: Multicopper oxidase, copper-binding site
1. SLC15A4, Solute carrier family 15, member 4
external annotation included chromosomal locations, 2. CP, Ceruloplasmin (ferroxidase)
protein domains and families, molecular functions, 3. F5, Coagulation factor V (proaccelerin, labile factor)
cellular localizations, biological processes, signaling and 4. HYAL4, Hyaluronoglucosamindase 4
metabolic pathways, protein-protein interaction 5. SNAI3, Snail homolog 3 (drosophila)
networks, protein complexes, and gene expression 6. HEPHL1, Hephaestin-like 1
7. ITIH3, Inter-alpha (globulin) inhibitor h3
signatures. The regulatory networks were derived by 8. CCDC73, Coiled-coil domain containing 73
scanning human promoters for known transcription factor 9. SLC14A1, Solute carrier family 14 (urea transporter),
motifs and by comparative genomics analyses that member 1
identified conserved promoter and 3’UTR elements. A 10. SLC14A2, Solute carrier family 14 (urea transporter),
P-value cutoff of 5X10-2 was used to cull significant member 2
11. CCIN, Calicin
concepts. In total, data from 12 databases and 335 high- 12. HEPH, Hephaestin
through put datasets were collected and analyzed.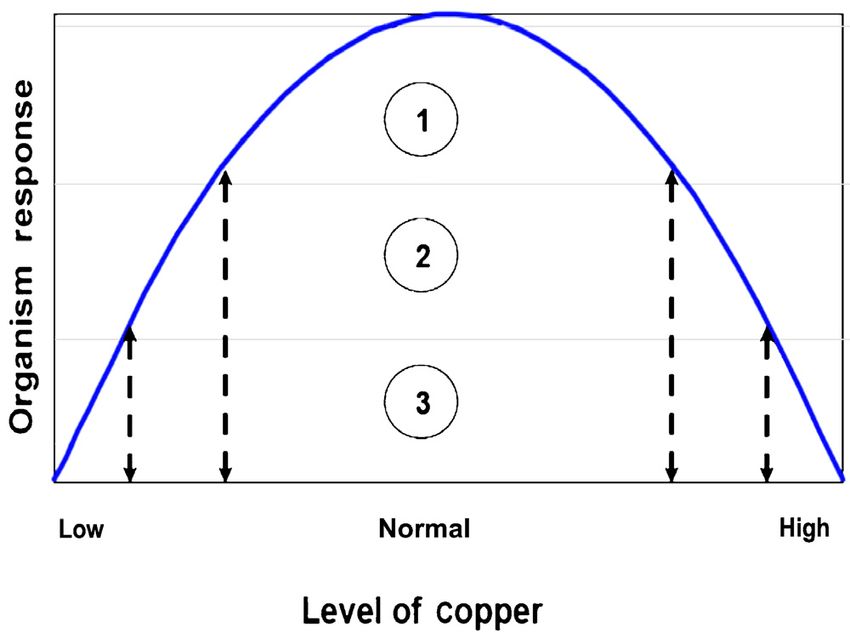
KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 455
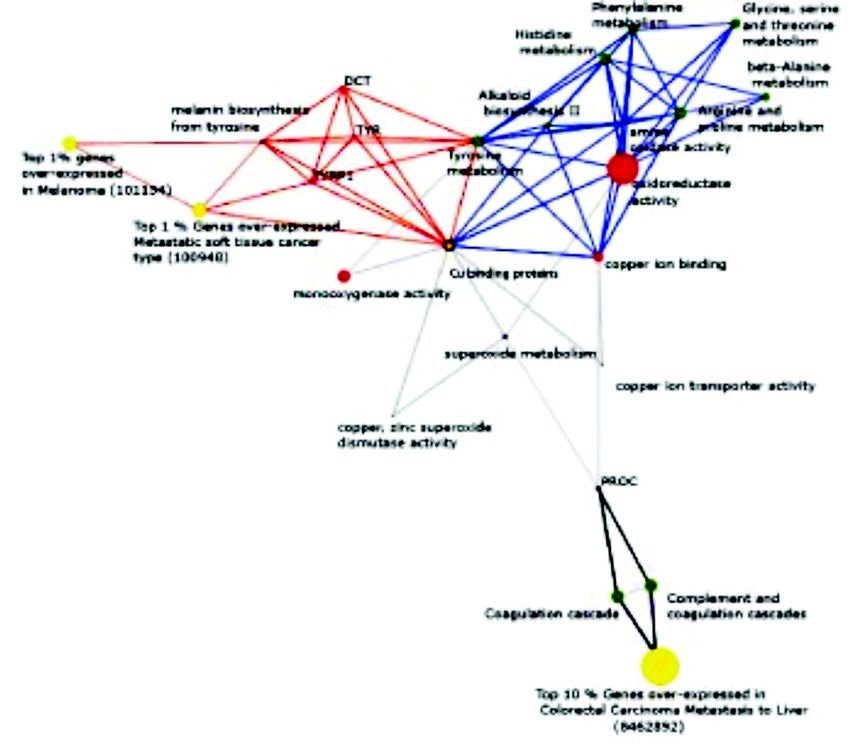
Interestingly, as shown in Fig. 9, the copper possesses three functional domains. Domain I contains
containing proteins play an active role in 3 major cellular Cu-binding site, domain II is homologous to SOD and
processes. These include tyrosine metabolism and domain III contains cysteines essential in the transfer of
melanin biosynthesis (red bridges), amino acid Cu to apo-SOD. CCS deletion has been documented to
metabolism (blue bridges) and coagulation cascade (black markedly reduce SOD activity in mice39,40. Third class
bridges). Further, included in the concept that portrayed of chaperone includes COX17, which delivers Cu to
“tyrosine metabolism and melanin biosynthesis” were cytochrome C oxidase (CCO). CCO is a large protein
multiple protein-protein complexes involving the proteins found in the cytoplasm and mitochondrial inner
Dopachrome tautomerase, Tyrosinase and Tyrosinase- membrane. It has two subunits I and II, each containing
related protein 1, all of which are copper binding proteins Cu binding sites. Fourth class of copper chaperone
and play a critical role in the above bioprocess. Similarly includes metallothioneins (MT). These are cysteine rich
copper binding proteins, potentate amino acid proteins (30%) composed of 61 amino acids. Due to their
metabolism, by having a functional role in two biological high redox potential, MT’s regulate intracellular levels
processes, namely amine oxidase and oxidoreductase of Zn and Cu in addition to serving as potent mediators
activity. Additionally, copper binding proteins regulate of toxic metal detoxification. As a part of the former,
the coagulation cascade by forming protein complex with MT levels tightly regulate copper homeostasis in liver.
the PROC protein (inactivator of coagulation factors Va Interestingly, the pool of MT-Cu complex progressively
and VIIIa). Also the proteins that bind copper were decreases with age in mammals41,42. A fifth class of copper
intimately involved in superoxide metabolism. chaperone comprises of the membrane protein β amyloid
precursor protein (APP) that regulates import of the metal
Among the proteins involved in superoxide into brain. This is supported by the observation wherein
metabolism, CCS plays a key role in the transmission of copper levels in the brain of APP null mice are higher
Cu to pro-form of superoxide dismutase (apo-SOD). CCS compared to their wild type counterparts43.
Fig. 9. Oncomine concept map analysis of copper binding proteins. Network view of the molecular concept analysis for the copper binding
proteins derived from InterPro (orange node). Each node represents a molecular concept or a set of biologically related genes. The node size is
proportional to the number of genes in the concept. Each edge represents a statistically significant enrichment (P456 INDIAN J MED RES, OCTOBER 2008
In addition to the transporters, there are a number cerebellar) degeneration and connective tissue
of enzymes that require copper as cofactors for their abnormalities during the first 2-4 yr of infancy46. Similar
catalytic activity. Some of the members of this class of condition has been reported to occur in sheep where
cuproenzymes include Cu/Zn SOD (antioxidant the disorder is termed as Kinky Hair Disease47. The
defense), cytochrome C oxidase (mitochondrial primary mode of diagnosis involves the use of genetic
respiration), CP (iron metabolism), tyrosinase screens. Early diagnosis coupled with supplementation
(pigmentation), lysyl oxidase (collagen maturation in of copper (as Cu-histidine complex) could avoid
connective tissue), Hephaestin (intestinal iron efflux), neurodegeneration and lead to reinstatement of normal
dopamine β-hydroxylase (catecholamine synthesis), development46. Interestingly, cells derived from patients
Peptidylglycine α -amidating monooxygenase (peptide- with Menkes disease exhibit copper accumulation when
hormone processing), amine oxidase (removal of cultured in vitro48.
hormones), ascorbate oxidase and catalase oxidase Wilson’s disease is a rare autosomal recessive trait
(oxidation of primary alcohols to aldehydes)44. manifested in the chromosome, 13q14.3. The frequency
Interestingly, as would be discussed later, the OCM of occurrence of Wilson’s disease is about 1/30,000 to
also portrayed an enrichment of copper binding proteins 1/50,000 with a carrier frequency of 1 per cent and
in invasive tumors especially those associated with soft heterozygote frequency of 0.86 per cent, (confined to
tissue (melanoma) and liver. western world). Notably, this defect is caused due to
the mutation in the gene coding for ATP7B, whose
Prognosis through copper metabolism: Cu is found in original function is to regulate the bilary excretion of
all living organisms in trace quantities with an uptake excess copper. ATP7B is encoded by the WND gene.
range in humans being 0.9 to 10 mg/day. The metal, Cu Among several mutations that have been reported for
is found as a prosthetic group in metalloenzymes this gene (>200), the most well studied one is a point
binding to sulphur residues. Several physiological mutation involving replacement of the amino acid
reactions such as electron transfer, detoxification of histidine by glutamine at position 1069. The mutated
reactive oxygen species, connective tissue development, protein thus loses the ability to orient ATP in its catalytic
oxygen transport, oxygenation reactions are being site, thus impairing its normal function49. The resultant
mediated by Cu containing metalloenzymes. When Cu is the accumulation of Cu in liver leading to cirrhosis
regulation fails, a variety of biochemical disturbances and hemolysis. Advanced stages of the disorder are
develop. The failure in Cu elimination and its efflux characterized by deposition of excess Cu in brain and
leads to Wilson’s and Menkes diseases respectively. eyes in the form of Kayaer-Fleischer ring, which serves
Another intriguing role of Cu is reported in the as diagnostic marker for Wilson’s disease 50 . The
promotion of angiogenesis for facilitating tumor to therapeutic measures for this disorder revolve around
progress. Therefore, by examining the distinguishing chelating the excess copper using chelating agents such
features of symptoms due to copper imbalance and its as tetrathiomolybdate, trientine and penicillamine.
metabolism, the possible prophylactic and
chemotherapeutic agents could be designed. Contrary to copper accumulation, its deficiency
can lead to hypocupremic state. Zatta and Frank 44
Disease symptoms due to Cu deficiency and overload reported that there was an incidence of 11.3 million
The disturbance in the levels of Cu is primarily due clinically identifiable Cu deficiency cases in 1970,
to genetic defects. The most prominent among these which has since been on the rise. Copper deficiency
are Menkes and Wilson’s diseases. could be a result of either inadequate dietary intake
(also termed primary copper deficiency) or due to
Menkes disease is a rare X-linked (Xq13) fatal impairment in its uptake (secondary copper
disorder affecting one out of 200,000 newborn infants, deficiency). The latter could be caused by the presence
resulting from a mutation in the gene encoding ATP7A. of additional heavy metals in the diet that could
The mutant protein is no longer able to regulate the competitively diminish copper uptake in the lining of
flux of copper resulting in a systemic deficiency of gastrointestinal tract. Among these, molybdenum is
copper45. Specifically, most of the Cu accumulates in the most common competitor of copper absorption.
intestinal epithelium and kidney while suboptimal levels Importantly, the relative ratio of dietary Cu: MO have
of the metal are found in other tissues such as liver and been defined to be 4 and 8 51 respectively to achieve
brain. Menkes disease is a fatal disorder, wherein optimal control in nutritional balance and hence copper
lethality is preceded by neuronal (cerebral and homeostasis in ruminants.KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 457
Additional disorders are caused by mutations in induction of tumor angiogenesis53. This is supported by
various cuproenzymes as reported by Prohaska52. These experiments conducted by Parke et al58, wherein dose-
include, (i) Albinism, wherein an impairment of an dependent neovascularisation (angiogenesis) is noticed
enzyme tyrosinase which is a critical intermediate in upon implanting Cu pellet into rabbit cornea.
melanin biosynthesis, (ii) Over gene dose effect of Cu-
Zn SOD noticed in Down Syndrome (trisomy 21) due Notably, copper exerts its effect on angiogenesis by
to the presence of this gene on the chromosome 21, inducing endothelial cell proliferation and migration by
(iii) X-linked Cutis laxa (or an analogous disorder in the way of activation of various angiogenic factors. The
mouse termed blotchy mouse), which are characterized latter include vascular endothelial growth factor (VEGF),
by defects in cross-linking of collagen due to decreased basic fibroblast growth factor (bFGF), tumor necrosis
lysyl oxidase activity, (iv) Mottled mice, an X-linked factor α (TNFα) and Interleukin 1 (IL-1)53. These
disorder analogous to Menkes disease wherein Cu angiogenic factors in turn activate resting endothelial cells
metabolism is affected. These mice have a mottled (which are otherwise in G0 phase of the cell cycle) and
appearance due to decreased melanin pigmentation initiate their proliferation by transitioning them to the
resulting from a reduction in tyrosinase activity, and G1 phase of the cell cycle. This process of endothelial
(v) Toxic milk mutant mouse, a homozygous trait caused activation by copper can be reversed using chelating
by Cu accumulation in liver. This results in a decreased agents like penicillamine59, a property that is widely
copper content in milk of lactating mothers which is exploited in designing therapeutic regimens (see section
toxic to the suckling offspring. By virtue of its similarity below). Additionally copper has been thought to exert
to Wilson’s disease in accumulating copper in liver, the its effect by binding to proteins like heparin, CP, etc.,
toxic milk mutant mouse could serve as a paradigm for making them angiogenic60. The angiogenic property of
understanding the mechanism that underlies the the latter is evident in the observation wherein CP has
development of Wilson’s disease. been reported to induce the formation of capillaries in
the cornea of rabbits58.
Role of copper in tumor development and progression:
Copper metabolism is a critical component of tumor To understand the effect of copper in cancer, we used
progression. Concentration of copper in serum has been the data from an interesting study aimed at predicting
found to correlate well with tumor development, size, the chemosensitivity of human cancer cell lines61. In this
progression as well as recurrence53. Elevated levels of study chemosensitivity predictions were based on
circulating copper in serum have been documented in transcriptomic profiling done upon treatment with various
cancers of lung, breast, gastrointestinal tract, brain as compounds on a panel of 60 cancer cell lines (NCI-60
well as gynecological cancers54,55. Importantly, copper panel)61. A set of 50 genes were found to be differentially
levels are higher in metastatic disease compared to regulated between copper sensitive and resistant cell lines
localized tumors54. This increase in serum copper levels upon treatment with 0.0001M copper sulfate. These set
during neoplastic progression is reflected in of 50 genes were used for enrichment analyses to
concomitant increase in the levels of CP, the primary understand the role of copper in tumor progression. The
carrier of copper in serum53. Interestingly, CP has been enrichment analyses was done using OCM as described
nominated as potential marker for diagnosis of advanced above37. Interestingly, the differentially expressed genes
solid tumors56. Additional evidence for the role of between copper sensitive and resistant cell lines mapped
copper in tumor development is derived from to multiple gene expression signatures (red nodes)
experiments that show existence of Cu salts in tumor derived from tumors that included sarcoma, lung
extracts that could stimulate the migration of endothelial carcinoma, colorectal cancer, etc. (Fig. 10). Furthermore,
cells in vitro57. copper induced genes also mapped to a subset of genes
that are activated upon Src over expression (red node)
The role of copper in tumor progression is best
(Fig.10). This is important in the context of earlier studies
understood in the light of the knowledge that developing
that have described a critical role for Src in tumor
tumors require an ample supply of oxygen and nutrients
that necessitates the development of a well defined development and progression. These observations
provide evidence at the molecular level for the role of
vasculature. The process termed angiogenesis is critical
copper in tumor progression.
for tumor proliferation and metastatic spread. Among the
various factors that lead to initiation of the angiogenic Brain disorders due to Cu deficiency and/or excess: The
process, tumor associated hypoxia seems to play a major brain is an organ of bewildering complexity with
role. Importantly, copper also plays a major role in the multifaceted serendipitous effects. In such a resilient458 INDIAN J MED RES, OCTOBER 2008
etc.,67-69. In Alzheimer’s, the proteolytic product (Aß) is
mutated resulting in its accumulation which is assumed
to trigger free radical mediated injury viz., neuronal
injury70. Notably, under normal conditions, APP is known
to bind copper in its reduced state and facilitates its
transport along the length of the neuron from the cell
body to the axonal surface and to plasma membrane of
dendrites 63. However, in Alzheimer’s disease, APP
function is disrupted leading to oxidation of its bound
copper in presence of H2O2. This is accompanied by
fragmentation of APP resulting in Aß peptides. These
fragments are thought to aggregate and lead to oxygen
free radical injury in Alzheimer’s disease71. Additionally,
Cu also binds to extracellular plaques and causes
interference in Cu trafficking devices and in turn depletes
intracellular Cu repertoire. This would reduce the
activities of cytochrome oxidase and SOD. Thus,
increased oxidative stress coupled with the reduction in
key metabolic and defense mechanism could contribute
significantly to neuronal damage. Oral treatment of
Fig. 10. Oncomine concept map analysis of genes up regulated in transgenic mouse with clioquinol resulted in halving of
NCl-60 cancer cell lines upon treatment with 0.0001 M copper Aß levels and significantly increased the levels of Cu
sulfate (black node). Each node represents a molecular concept or and Zn in the brain72.
a set of biologically related genes. The node size is proportional to
the number of genes in the concept. Each edge represents a The other neuronal cellular membrane protein is prion.
statistically significant enrichment (PKRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 459
has been shown to be causal in inducing the de- progression. By its unique attribute of being a catalytic
myelination in affected animals77. cofactor, Cu occupies an important niche in biological
systems. Cu transporters, chaperone proteins and carrier
Cu chelation therapy: Dietary excess Cu intake is not
proteins make Cu available to the intricate network of
very common, although there are genetic disorders as
biochemical systems. Developments in the field of plant
discussed in the previous sections. The increased
genetic engineering have been pivotal in defining means
accumulation leads to hepatitis and neurological
to combat copper deficiency. In the clinical field,
disorders. Human Wilson’s disease and Toxic milk
management of disorders caused by impaired copper
mouse are associated with excess accumulation of
homeostasis are being combated either using metal
cellular Cu. In the former, the defect is manifested in
chelators or by supplementing the metal in a complex
Wilson protein (ATP7B) which in its normal form does
state with various carriers.
facilitate to eliminate excess Cu ions into bile.
Therapeutic approaches to Cu toxicity include the drugs Acknowledgment
and formulations such as D-penicillamine or trientine The authors thank to Dr M. Sivakumar, University of
to prevent neurodegenerative disorder78. Similarly, Wollongong, Australia, for designing a few of the figures shown in
tetrathiomolybdate, as a specific Cu chelator have been the text. One of the authors (SKN) acknowledges UGC and DST
used in Toxic milk mouse model in reducing abnormally (India) for providing financial support through SAP DRS and FIST
high Cu79. Since copper plays an important role in tumor programmes respectively.
development and progression (as discussed above), References
strategies employing Cu chelators are also being
pursued for cancer therapy80. In contrast, in conditions 1. Aaseth J, Flaten TP, Andersen O. Hereditary iron and copper
like Menkes disease that results from copper deficiency, deposition: diagnostics, pathogenesis and therapeutics. Scand
an approach to supplement copper complexed with J Gastroenterol 2007; 42 : 673-81.
histidine or albumin are being tested79. 2. Araya M, Pizarro F, Olivares M, Arredondo M, Gonzalez M,
Mendez M. Understanding copper homeostasis in humans and
Homeopathic formulations using Cu metal: copper effects on health. Biol Res 2006; 39 : 183-7.
Homeopathy is based on the argument that the body is 3. Decker H, Terwilliger N. Cops and robbers: putative evolution
a self-healing entity, and that symptoms are the of copper oxygen-binding proteins. J Exp Biol 2000; 203 :
1777-82.
expression of the body attempting to restore its balance.
Homeopathic physicians are trained to match the 4. Goodman VL, Brewer GJ, Merajver SD. Copper deficiency
as an anti-cancer strategy. Endocr Relat Cancer 2004; 11 :
patient’s symptoms with the accurate remedy. They 255-63.
believe that the remedies themselves never destroy
5. Srivastava S, Singh BR, Tripathi VN Application of bacterial
disease, but stimulate the body’s own healing action to biomass as a potential metal indicator. Curr Sci 2005; 89 :
get rid itself of the problem. Minerals in the body can 1248-51.
be used as healing agents for specific health problems. 6. Wapnir RA. Copper absorption and bioavailability. Am J Clin
Minerals are used in homeopathic remedies to stimulate Nutr 1998; 67 : 1054S-60S.
corresponding body cells towards metabolic activity and 7. Baker DH, Czarnecki-Maulden GL. Pharmacologic role of
health restoration. A few tinctures with the combination cysteine in ameliorating or exacerbating mineral toxicities.
of copper are: (i) Cuprum aceticum, (ii) Cuprum J Nutr 1987; 117 : 1003-10.
Arsenicosum, (iii) Cuprum Metallicum and (iv) Cuprum 8. Pott EB, Henry PR, Ammerman CB, Merritt AM, Madison
JB, Miles RD. Relative bioavailability of Cu in a Cu-lysine
Sulphuricum81 . complex for chicks and lambs. Anim Feed Sci Technol 1994;
All life forms exploit naturally available Cu for 45 : 193-203.
myriad physiological functions. Bacteria, plants, blue 9. Kincaid RL, Blauwiekel RM, Cronath JD. Supplementation
blooded organisms and vertebrates have developed the of Cu sulphate or Cu proteinate for growing calves fed forages
containing molybdenum. J Diary Sci 1986; 69 : 160-3.
molecular mechanisms to upkeep the Cu homeostasis.
10. Lowndes SA, Harris AL. The role of copper in tumour
The bioavailability of Cu, by complexing with proteins angiogenesis. J Mammary Gland Biol Neoplasia 2005; 10 :
or amino acids or organic acids constituting 299-310.
organometallic complex, facilitates its ease in uptake and 11. Gu M, Cooper JM, Butler P, Walker AP, Mistry PK, Dooley
distribution in ecosystem. Literature review reveals that JS, et al. Oxidative-phosphorylation defects in liver of patients
the Cu imbalance could be causal in Menkes disease, with Wilson’s disease. Lancet 2000; 356 : 469-74.
Wilson’s disease, Kuru, Creutzfeld- Jakob disease, mad 12. Krupanidhi S. Respiratory pigments. Biol Educ (India)1988;
cow disease as well as induce tumor development and 4 : 104-14.460 INDIAN J MED RES, OCTOBER 2008
13. Krupanidhi S, Laksmikanth T. Detection of haemocyanin in pharamacology and toxicology. Tofowa, NJ: Humana Press;
native PAGE gels. Natl Acad Sci Lett 2005; 28 : 353-5. 2002. p. 527-43.
14. Krupanidhi S, Venkata Reddy V, Padmanabha Naidu B. Some 32. Odermatt A, Krapf R, Solioz M. Induction of the putative
studies on copper metabolism in the garden snail,Cryptozona copper ATPases, CopA and CopB, of Enterococcus hirae by
ligulata. Indian J Exp Biol 1978; 16 : 249-50. Ag+ and Cu2+, and Ag+ extrusion by CopB. Biochem Biophys
Res Commun 1994; 202 : 44-8.
15. Krupanidhi S. Copper granules in the hepatopancreas of the
snail, Crytpzona ligulata. Curr Sci 1985; 53 : 431-2. 33. Wunderli-Ye H, Solioz M. Effects of promoter mutations on
the in vivo regulation of the cop operon of Enterococcus hirae
16. van Holde KE, Miller KI. Hemocyanins. Adv Protein Chem
by copper(I) and copper(II). Biochem Biophys Res Commun
1995; 47 : 1-81.
1999; 259 : 443-9.
17. Brown NL, Camakaris J, Lee BT, Williams T, Morby AP,
34. Gajiwala KS, Burley SK. Winged helix proteins. Curr Opin
Parkhill J, et al. Bacterial resistances to mercury and copper.
Struct Biol 2000; 10 : 110-6.
Cell Biochem 1991; 46 : 106-14.
35. Magnani D, Solioz M. Copper chaperone cycling and
18. Rouch DLB, Camakaris J. Metal on homeostasis: Molecular
degradation in the regulation of the cop operon of
biology and chemistry. Supplement: UCLA Symposia on
Enterococcus hirae. Biometals 2005; 18 : 407-12.
Molecular & Cellular Biology 1989; 38 : 439-46.
36. Portmann R, Magnani D, Stoyanov JV, Schmechel A,
19. Grotz N, Guerinot ML. Molecular aspects of Cu, Fe and Zn Multhaup G, Solioz M. Interaction kinetics of the copper-
homeostasis in plants. Biochim Biophys Acta 2006; 1763 : responsive CopY repressor with the cop promoter of
595-608. Enterococcus hirae. J Biol Inorg Chem 2004; 9 : 396-402.
20. Sancenon V, Puig S, Mira H, Thiele DJ, Penarrubia L. 37. Rhodes DR, Kalyana-Sundaram S, Tomlins SA, Mahavisno
Identification of a copper transporter family in Arabidopsis V, Kasper N, Varambally R, et al. Molecular concepts analysis
thaliana. Plant Mol Biol 2003; 51 : 577-87. links tumors, pathways, mechanisms, and drugs. Neoplasia
21. Petris MJ. The SLC31 (Ctr) copper transporter family. Pflugers 2007; 9 : 443-54.
Arch 2004; 447 : 752-5. 38. Tomlins SA, Mehra R, Rhodes DR, Cao X, Wang L,
22. Chelly J, Tumer Z, Tonnesen T, Petterson A, Ishikawa-Brush Dhanasekaran SM, et al. Integrative molecular concept
Y, Tommerup N, et al. Isolation of a candidate gene for Menkes modeling of prostate cancer progression. Nature Genet 2007;
disease that encodes a potential heavy metal binding protein. 39 : 41-51.
Nat Genet 1993; 3 : 14-9. 39. Prohaska JR, Gybina AA. Intracellular copper transport in
23. Mercer JF, Livingston J, Hall B, Paynter JA, Begy C, mammals. J Nutr 2004; 134 : 1003-6.
Chandrasekharappa S, et al. Isolation of a partial candidate 40. Wong PC, Waggoner D, Subramaniam JR, Tessarollo L,
gene for Menkes disease by positional cloning. Nat Genet Bartnikas TB, Culotta VC, et al. Copper chaperone for
1993; 3 : 20-5. superoxide dismutase is essential to activate mammalian Cu/
24. Mufti AR, Burstein E, Duckett CS. XIAP: cell death regulation Zn superoxide dismutase. Proc Natl Acad Sci USA 2000; 97 :
meets copper homeostasis. Arch Biochem Biophys 2007; 463 2886-91.
: 168-74. 41. Coyle P, Philcox JC, Carey LC, Rofe AM. Metallothionein: the
25. La Fontaine S, Mercer JF. Trafficking of the copper-ATPases, multipurpose protein. Cell Mol Life Sci 2002; 59 : 627-47.
ATP7A and ATP7B: role in copper homeostasis. Arch Biochem 42. Hamza I, Faisst A, Prohaska J, Chen J, Gruss P, Gitlin JD.
Biophys 2007; 463 : 149-67. The metallochaperone Atox1 plays a critical role in perinatal
26. Ackland ML, Anikijenko P, Michalczyk A, Mercer JF. copper homeostasis. Proc Natl Acad Sci USA 2001; 98 : 6848-
Expression of menkes copper-transporting ATPase, MNK, in 52.
the lactating human breast: possible role in copper transport 43. White AR, Reyes R, Mercer JF, Camakaris J, Zheng H, Bush
into milk. J Histochem Cytochem 1999; 47 : 1553-62. AI, et al. Copper levels are increased in the cerebral cortex
27. Guo Y, Smith K, Lee J, Thiele DJ, Petris MJ. Identification of and liver of APP and APLP2 knockout mice. Brain Res 1999;
methionine-rich clusters that regulate copper-stimulated 842 : 439-44.
endocytosis of the human Ctr1 copper transporter. J Biol Chem 44. Zatta P, Frank A. Copper deficiency and neurological disorders
2004; 279 : 17428-33. in man and animals. Brain Res Rev 2007; 54 : 19-33.
28. van De Sluis B, Rothuizen J, Pearson PL, van Oost BA, 45. Daniel KG, Harbach RH, Guida WC, Dou QP. Copper storage
Wijmenga C. Identification of a new copper metabolism gene diseases: Menkes, Wilsons, and cancer. Front Biosci 2004; 9
by positional cloning in a purebred dog population. Hum Mol : 2652-62.
Genet 2002; 11 : 165-73.
46. Gu YH, Kodama H, Sato E, Mochizuki D, Yanagawa Y,
29. Klomp AE, van de Sluis B, Klomp LW, Wijmenga C. The Takayanagi M, et al. Prenatal diagnosis of Menkes disease by
ubiquitously expressed MURR1 protein is absent in canine genetic analysis and copper measurement. Brain Dev 2002;
copper toxicosis. J Hepatol 2003; 39 : 703-9. 24 : 715-8.
30. de Bie P, van de Sluis B, Klomp L, Wijmenga C. The many 47. Menkes JH. Kinky hair disease: twenty five years later. Brain
faces of the copper metabolism protein MURR1/COMMD1. Dev 1988; 10 : 77-9.
J Hered 2005; 96 : 803-11. 48. Horn N. Menkes’ X-linked disease: prenatal diagnosis and
31. Wimmer R, Dameron CT, Solioz M. Molecular hardware of carrier detection. J Inherit Metab Dis 1983; 6 (Suppl 1) : 59-
Cu homeostasis in Enterococcus hirae. Handbook of Cu 62.KRUPANIDHI et al: COPPER & BIOLOGICAL HEALTH 461
49. Shim H, Harris ZL. Genetic defects in copper metabolism. fibrillogenesis in a rat brain model of amyloidosis: implications
J Nutr 2003; 133 : 1527S-31S. for Alzheimer’s therapy. Nature Med 1998; 4 : 822-6.
50. Sarkar B. Treatment of Wilson and menkes diseases. Chem 67. Huang X, Atwood CS, Hartshorn MA, Multhaup G, Goldstein
Rev 1999; 99 : 2535-44. LE, Scarpa RC, et al. The A beta peptide of Alzheimer’s disease
directly produces hydrogen peroxide through metal ion
51. Underwood EJ, Suttle NF. Cu. The mineral nutrition of livestock.
reduction. Biochemistry 1999; 38 : 7609-16.
3 rd ed. New York: CABI Publsihing Oxon; 2001.
p. 283-342. 68. Butterfield DA, Hensley K, Cole P, Subramaniam R, Aksenov
M, Aksenova M, et al. Oxidatively induced structural alteration
52. Prohaska JR. Genetic diseases of copper metabolism. Clin of glutamine synthetase assessed by analysis of spin label
Physiol Biochem 1986; 4 : 87-93. incorporation kinetics: relevance to Alzheimer’s disease.
53. Nasulewicz A, Mazur A, Opolski A. Role of copper in tumour J Neurochem 1997; 68 : 2451-7.
angiogenesis-clinical implications. J Trace Elem Med Biol 69. Stadtman ER. Metal ion-catalyzed oxidation of proteins:
2004; 18 : 1-8. biochemical mechanism and biological consequences. Free
54. Zowczak M, Iskra M, Torlinski L, Cofta S. Analysis of serum Radic Boil Med 1990; 9 : 315-25.
copper and zinc concentrations in cancer patients. Biol Trace 70. Wong PC, Rothstein JD, Price DL. The genetic and molecular
Elem Res 2001; 82 : 1-8. mechanisms of motor neuron disease. Curr Opin Neurobiol
55. Yoshida D, Ikeda Y, Nakazawa S. Quantitative analysis of 1998; 8 : 791-9.
copper, zinc and copper/zinc ratio in selected human brain 71. Multhaup G, Ruppert T, Schlicksupp A, Hesse L, Bill E,
tumors. J Neurooncol 1993; 16 : 109-15. Pipkorn R, et al. Copper-binding amyloid precursor protein
56. Senra Varela A, Lopez Saez JJ, Quintela Senra D. Serum undergoes a site-specific fragmentation in the reduction of
ceruloplasmin as a diagnostic marker of cancer. Cancer Lett hydrogen peroxide. Biochemistry 1998; 37 : 7224-30.
1997; 121 : 139-45. 72. Cherny RA, Atwood CS, Xilinas ME, Gray DN, Jones WD,
McLean CA, et al. Treatment with a copper-zinc chelator
57. Hu GF. Copper stimulates proliferation of human endothelial
markedly and rapidly inhibits beta-amyloid accumulation in
cells under culture. J Cell Biochem 1998; 69 : 326-35.
Alzheimer’s disease transgenic mice. Neuron 2001; 30 : 665-76.
58. Parke A, Bhattacherjee P, Palmer RM, Lazarus NR. 73. Cerpa W, Varela-Nallar L, Reyes AE, Minniti AN, Inestrosa
Characterization and quantification of copper sulfate-induced NC. Is there a role for copper in neurodegenerative diseases?
vascularization of the rabbit cornea. Am J Pathol 1988; Mol Aspects Med 2005; 26 : 405-20.
130 : 173-8.
74. Jones CE, Abdelraheim SR, Brown DR, Viles JH. Preferential
59. Pan Q, Kleer CG, van Golen KL, Irani J, Bottema KM, Bias Cu2+ coordination by His96 and His111 induces beta-sheet
C, et al. Copper deficiency induced by tetrathiomolybdate formation in the unstructured amyloidogenic region of the
suppresses tumor growth and angiogenesis. Cancer Res 2002; prion protein. J Biol Chem 2004; 279 : 32018-27.
62 : 4854-9.
75. Sigurdsson EM, Brown DR, Alim MA, Scholtzova H, Carp
60. Ziche M, Jones J, Gullino PM. Role of prostaglandin E1 and R, Meeker HC, et al. Copper chelation delays the onset of
copper in angiogenesis. J Natl Cancer Inst 1982; 69 : 475-82. prion disease. J Biol Chem 2003; 278 : 46199-202.
61. Staunton JE, Slonim DK, Coller HA, Tamayo P, Angelo MJ, 76. Picco SJ, De Luca JC, Mattioli G, Dulout FN. DNA damage
Park J, et al. Chemosensitivity prediction by transcriptional induced by copper deficiency in cattle assessed by the Comet
profiling. Proc Natl Acad Sci USA 2001; 98 : 10787-92. assay. Mutat Res 2001; 498 : 1-6.
62. Miranda S, Opazo C, Larrondo LF, Munoz FJ, Ruiz F, 77. Kumar N, Gross JB, Jr, Ahlskog JE. Copper deficiency
Leighton F, et al. The role of oxidative stress in the toxicity myelopathy produces a clinical picture like subacute combined
induced by amyloid beta-peptide in Alzheimer’s disease. Prog degeneration. Neurology 2004; 63 : 33-9.
Neurobiol 2000; 62 : 633-48. 78. Offen D, Gilgun-Sherki Y, Barhum Y, Benhar M, Grinberg L,
Reich R, et al. A low molecular weight copper chelator crosses
63. Selkoe DJ. The cell biology of beta-amyloid precursor protein
the blood-brain barrier and attenuates experimental autoimmune
and presenilin in Alzheimer’s disease. Trends Cell Biol 1998; encephalomyelitis. J Neurochem 2004; 89 : 1241-51.
8 : 447-53.
79. Czachor JD, Cherian MG, Koropatnick J. Reduction of copper
64. Soto C, Branes MC, Alvarez J, Inestrosa NC. Structural and metallothionein in toxic milk mice by tetrathiomolybdate,
determinants of the Alzheimer’s amyloid beta-peptide. but not deferiprone. J Inorg Biochem 2002; 88 : 213-22.
J Neurochem 1994; 63 : 1191-8.
80. Cai L, Li XK, Song Y, Cherian MG. Essentiality, toxicology
65. Yankner BA. Mechanisms of neuronal degeneration in and chelation therapy of zinc and copper. Curr Med Chem
Alzheimer’s disease. Neuron 1996; 16 : 921-32. 2005; 12 : 2753-63.
66. Soto C, Sigurdsson EM, Morelli L, Kumar RA, Castano EM, 81. Clarke J. A dictionary of practical materia medica. B. Jain
Frangione B. Beta-sheet breaker peptides inhibit Publishers Pvt. Ltd.; 1990; 1: 633-44.
Reprint requests: Dr S. Krupanidhi, Department of Biosciences, Sri Sathya Sai University
Prasanthi Nilayam 515 134, India
e-mail: krupanidhi_srirama@yahoo.comYou can also read

















































