The solution structure of the complement deregulator FHR5 reveals a compact dimer and provides new insights into CFHR5 nephropathy
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JBC Papers in Press. Published on September 14, 2020 as Manuscript RA120.015132 The latest version is at https://www.jbc.org/cgi/doi/10.1074/jbc.RA120.015132 The solution structure of the complement deregulator FHR5 reveals a compact dimer and provides new insights into CFHR5 nephropathy Nilufar Kadkhodayi-Kholghi1, Jayesh S. Bhatt1, Jayesh Gor1, Lindsay C. McDermott2, Daniel P. Gale3 and Stephen J. Perkins1‡ 1 Department of Structural and Molecular Biology, Division of Biosciences, Darwin Building, University College London, Gower Street, London WC1E 6BT, United Kingdom 2 School of Life Sciences, University of Bedfordshire, Park Square, Luton LU1 3JU, United Kingdom 3 UCL Department of Renal Medicine, Royal Free Hospital, University College London, London NW3 2PF United Kingdom Running Title: Solution structure of FHR5 Downloaded from http://www.jbc.org/ by guest on September 24, 2020 ‡ Author to whom correspondence and requests for reprints should be addressed (SJP: Tel: 020-7679-7048; Fax: 020-7679-7193; email s.perkins@ucl.ac.uk). Keywords: analytical ultracentrifugation; atomistic modelling; FHR5; Complement; molecular dynamics; molecular modelling; Monte Carlo simulations; small angle X-ray scattering ABSTRACT arrangements of SCR domains for The human complement Factor H-related 5 scattering curve fits. Only compact domain protein (FHR5) antagonizes the main structures in this library fit well to the circulating complement regulator Factor H, scattering data, and these structures readily resulting in the deregulation of complement accommodated the extra SCR-1/2 domain activation. FHR5 normally contains nine pair present in CFHR5 nephropathy. This short complement regulator (SCR) model indicated that mutant FHR5 can domains, but a FHR5 mutant has been form oligomers that possess additional identified with a duplicated N-terminal binding sites for C3b in FHR5. We SCR-1/2 domain pair that causes CFHR5 conclude that the deregulation of nephropathy. To understand how this complement regulation by the FHR5 duplication causes disease, we mutant can be rationalized by the enhanced characterized the solution structure of binding of FHR5 oligomers to C3b native FHR5 by analytical deposited on host cell surfaces. Our FHR5 ultracentrifugation and small-angle X-ray structures thus explained key features of the scattering. Sedimentation velocity and X- mechanism and pathology of CFHR5 ray scattering indicated that FHR5 was nephropathy. dimeric, with a radius of gyration RG of 5.5 _________________________________ ± 0.2 nm and a maximum protein length of 20 nm for its 18 domains. This result Complement activation and indicated that FHR5 was even more regulation is of major importance in compact than the main regulator Factor H enabling clearance of pathogens, whilst which showed an overall length of 26-29 preventing complement-mediated host cell nm for its 20 SCR domains. Atomistic damage. Complement factor H related 5 modelling for FHR5 generated a library of protein (FHR5) was first identified co- 250,000 physically-realistic trial localised with C3 in glomerular immune 1
deposits from patients with data exist on whether FHR5 forms glomerulonephritis, and is a member of a heterodimers with other FHRs in vivo (6,7). family of structurally related proteins comprising the major serum complement CFHR5 nephropathy, a monogenic regulator Factor H and five complement cause of kidney failure endemic in Cypriots factor H related proteins. Factor H, (individuals residing in or with ancestry comprising 20 short complement regulator from the island of Cyprus), is characterised (SCR) domains, has been well in almost all affected individuals by characterised, both in terms of its structure persistent microscopic haematuria and, in a and function, binding to activated C3b and proportion of patients, episodes of kidney its fragment C3d, and regulating excess C3 damage and visible blood in the urine that activation (1). However, the principal occur at times of otherwise trivial mucosal physiological function of FHR5 is poorly infections, with repeated episodes typically understood. FHR5 circulates in plasma in resulting in progressive kidney damage and extremely low concentrations of 3-6 µg/ml eventually end stage kidney failure (2), which is approximately 100-fold lower occurring in >80% affected males and than Factor H. It is also the least abundant
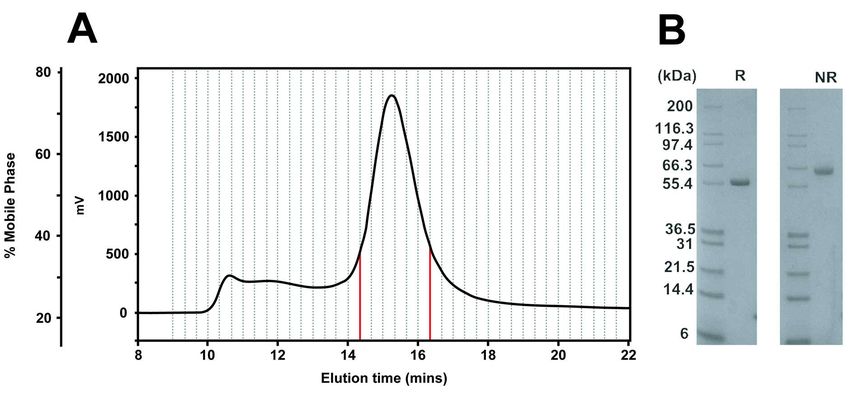
and eight residues (Figure 1), both of which Results make it difficult to crystallise in order to Purification of full-length FHR5 determine its three-dimensional Human FHR5 SCR-1/9 purchased appearance. To date, atomic-level from Creative Biolabs was subjected to gel structures have not been determined for any filtration chromatography to ensure small FHR5 fragments. However, monodispersity and removal of aggregates alternative methods can be used for prior to SAXS experiments. The protein structural studies. Previously for full-length eluted as a single symmetrical peak at factor H, electron microscopy, small-angle approximately 15 ml elution volume X-ray scattering (SAXS), analytical (Figure 2A). This was preceded by a ultracentrifugation (AUC), and molecular broader peak that was eluted between 10 modelling showed that full-length factor H and 14 ml, which was attributed to protein has a partially folded-back structure that is aggregates. Only the protein fractions relevant to its regulatory function (10-12). between 14.3-16.3 ml (red in Figure 2A) This combination of analytical were retained. By SDS-PAGE (Figure 2B), ultracentrifugation, X-ray solution a single band was seen at 60-66 kDa (non- scattering and atomistic modelling has been reduced) that corresponds well to the effective in determining many expected monomer molecular mass of 62.4 Downloaded from http://www.jbc.org/ by guest on September 24, 2020 macromolecular structures in solution (13- kDa. Reducing conditions resulted in 15). Many of the first structural another single band but at a slightly lower explanations for factor H-associated mass, this difference being attributed to the diseases such as atypical haemolytic presence of glycan chains on FHR5. uraemic syndrome were based on homology models for the SCR domains SEC-MALLS was used to (16-18). Here, these solution structural and determine the mass and self-association of modelling approaches were applied to FHR5 in our Tris-150 purification buffer, as determine the solution conformation of full- in previous work (4). FHR5 from a size- length FHR5 in order to explain its role in exclusion column was detected by UV healthy individuals and how CFHR5 (blue, Figure 3) and refractive index (green) nephropathy may arise through the SCR- measurements, in parallel with multi-angle 1/2 duplication. Following SAXS and AUC light scattering (red) to analyse size data collection, full-length FHR5 was distributions. Three peaks were observed in modelled using molecular dynamics, the elution profile. Peak 1 at 2.7-4.2 min followed by Monte Carlo simulations to was assigned as aggregated material, generate a large library of physically- because this had a lower UV and refractive realistic trial atomistic structures for the index, but high light scattering intensities FHR5 dimer (14,19). The theoretical that indicated very large sizes. Its molecular scattering profiles of this library were mass was calculated to be above 5,400 kDa. compared to the experimental SAXS curves Peak 2 at 4.9-5.2 min was the FHR5 dimer to determine best-fit FHR5 structures. We that eluted with higher UV and refractive thus defined a small subset of compact index values but with lower light scattering. folded-back solution structures. The extra Its molecular mass was estimated as 162 SCR-1/2 domain pair in mutant FHR5 was kDa, this being consistent with FHR5 dimer readily added to these structures, their formation, given that the mass of the presence leading to the formation of monomer was 62.4 kDa from its multivalent oligomers of FHR5. Our work composition (20). Despite a large inherent explains how FHR5 regulates complement error associated with light scattering, no activation in the kidney and how CFHR5 evidence of a FHR5 monomer peak was nephropathy arises. detectable. A small peak 3 at 7.6-7.9 min was assigned to fragments below 30 kDa. 3

shift in the FHR5 dimer peak was visible in AUC analyses of FHR5 the c(s) distribution plots (vertical dashed AUC sedimentation velocity lines, Figure 4). This result indicated a experiments on FHR5 studied its conformational change in FHR5, where the oligomerisation and shape using size smaller s20,w values at high NaCl distribution c(s) analyses to determine its concentration indicated a more elongated molecular mass and sedimentation FHR5 domain structure that formed as the coefficient s20,w. Absorbance data for FHR5 ionic strength was increased (Figure 5). at 0.16 mg/ml in PBS were collected for five different salt concentrations between SAXS analyses of FHR5 20-250 mM NaCl. SEDFIT analyses SAXS was used to study the involved as many as 500 absorbance scans. solution structure of the FHR5 dimer in The experimental sedimentation concentration series in three different boundaries (left, Figure 4) gave good fits to buffers, two being physiological (PBS-137 the Lamm equation to give the size- and Tris-150) and one being low salt (PBS- distribution c(s) profiles (right, Figure 4), 50). The FHR5 samples were purified by despite the low concentrations in use. These gel filtration (Figure 2). In Tris-150, data fits were obtained by floating the meniscus, were collected using 0.04-0.5 mg/ml FHR5. Downloaded from http://www.jbc.org/ by guest on September 24, 2020 bottom of the cell, the baseline and the In PBS-137 and PBS-50, data were frictional ratio f/f0 of around 1.5. collected using 0.04-0.17 mg/ml FHR5. Guinier analyses of the solution structure Protein aggregation was visible in gave high quality linear plots in two distinct the earliest boundaries that sedimented regions of the I(Q) curves that rapidly at the start of the runs, to leave corresponded to the radius of gyration RG behind the FHR5 dimer that sedimented and the cross-sectional radius of gyration more slowly (Figure 4). This agreed with RXS from two distinct Q-ranges (Figure 6). SEC-MALLS. A major c(s) peak at 6.0 S These values are measures of the overall was observed for FHR5 in PBS-137 that and the shorter dimensions of corresponded to an average molecular mass macromolecular elongation respectively. of 134 kDa. This mass confirmed the Their values were deduced according to presence of dimer in solution. The Equations (1) and (2) respectively, within aggregates made little contribution to the satisfactory Q.RG and Q.RXS limits close to c(s) analyses between 3-12 S, even though 1.0: they contributed as much as half the protein (i) In the overall structural Guinier present. The molecular masses for the five RG analyses in a low Q-range of 0.1 - 0.27 buffers were between 133 kDa to 139 kDa nm-1 (Figure 7A), in Tris-150 and PBS-137 (Table 1), showing that the FHR5 dimer buffers with similar NaCl concentrations, was stable between 20 mM to 350 mM the mean RG values were 5.36 ± 0.14 nm NaCl. The c(s) analyses did not reveal any and 5.48 ± 0.17 nm respectively. However, FHR5 monomer at lower s values. The in the PBS-50 buffer with lower NaCl, the reproducibility of these data was tested at mean RG value increased slightly to 5.91 ± two different rotor speeds of 40,000 rpm 0.13 nm. This increase was attributed to and 50,000 rpm, to show no difference. trace aggregation in FHR5 that affected the lowest Q values (Figure 7A). No The solution structure of FHR5 concentration dependence was observed for between 20 mM to 350 mM NaCl was the RG values between 0.04 and 0.17 mg/ml, monitored using the mean s20,w values however a slightly increased RG value of up (Table 1). A significant decrease of 0.9 S to 0.2 nm was seen at 0.2-0.5 mg/ml FHR5. from 6.48 S to 5.35 S was seen on going (ii) In the cross-sectional Guinier from 20 mM NaCl to 350 mM NaCl. This RXS analyses, using a Q-range of 0.32 – 0.55 4

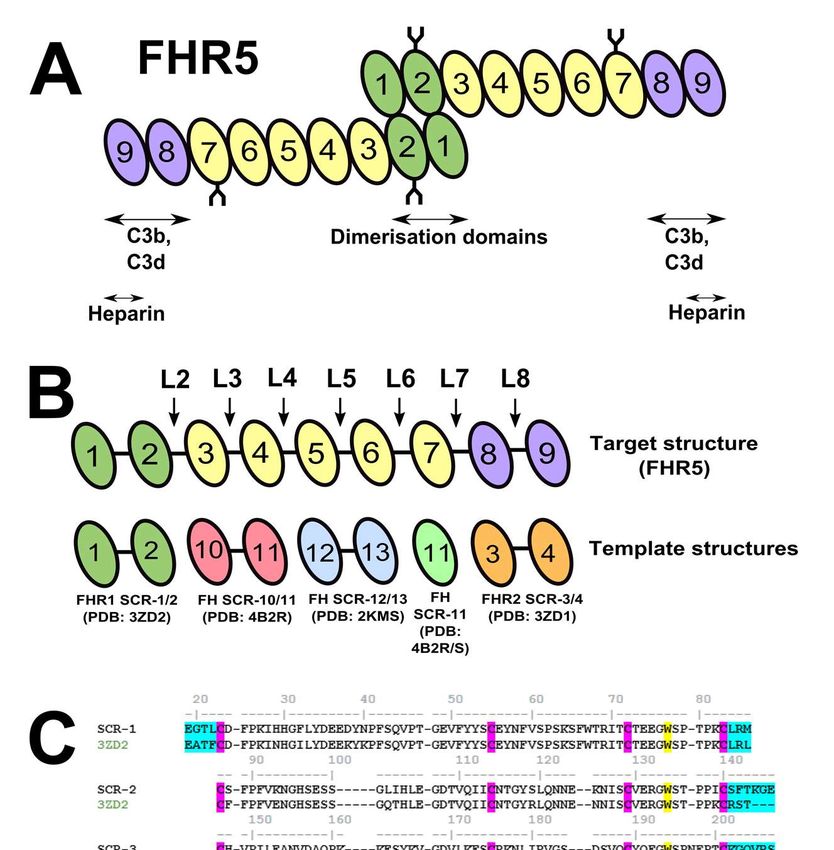
nm-1 (Figure 7B), the mean RXS values in based on four known SCR crystal structures each buffer were 2.41 ± 0.06 nm, 2.29 ± as structural templates (Figures 1B,C). Two 0.09 nm, and 2.46 ± 0.14 nm for Tris-150, used related crystal structures of the N- PBS-137 and PBS-50 respectively (Table terminal FHR1 SCR-1/2 domains and the 2). No significant changes in the RXS values C-terminal FHR2 SCR-3/4 domains with were seen between the data sets for these high sequence identities of 85.2% and NaCl and protein concentrations, indicating 61.7% respectively with SCR-1/2 and SCR- that the cross-sectional structure of FHR5 8/9 of FHR5. The SCR-3/7 domains of was unchanged in conformation. FHR5 shares significant sequence similarities with the SCR-10/14 domains of The distance distribution function Factor H. Although templates for individual P(r) in real space represents all the SCR3/7 domains in FHR5 were searched distances between pairs of atoms in FHR5. for in PDB-Blast, the best choices were This was calculated from Fourier these domain structures from Factor H due transformation of the full I(Q) scattering to their direct sequence similarities (Figure curve following the specification of the 1C). FHR5 SCR-3/4 was represented by maximum dimension Dmax (Equation (3); Factor H SCR-10/11 with a high sequence Figure 8). The P(r) curve provided an identity of 57.4%. FHR5 SCR-5/6 was Downloaded from http://www.jbc.org/ by guest on September 24, 2020 independent RG value for FHR5 for represented by Factor H SCR-12/13, also comparison with the Guinier value (Table with a high sequence identity of 53.9%. 2). The RG values from the P(r) analyses While FHR5 SCR-7 is similar to Factor H were in good agreement with those from the SCR-14, no structure existed for Factor H Guinier analyses (Table 2). The P(r) curve SCR-14. Searches showed that the best also gave the maximum length L of FHR5 template structure for FHR5 SCR-7 was from the value of r when P(r) = 0. The mean that of SCR-11 of Factor H with a sequence L values were 19.5 ± 0.4 nm in Tris-150 identity of 34.5%. The individual template- (Figure 8C), 19.6 ± 0.5 nm in PBS-137 target sequence alignments (Figure 1C) (Figure 8B) and 21.0 nm in PBS-50 (Figure showed no significant indels in the 8A). The L value for PBS-50 was slightly structure, because the number of residues in higher than those in Tris-150 and PBS-137, these were well aligned. Thus the FHR5 most likely due to trace aggregation that SCR-7 and SCR-8/9 sequences had only resulted from the lower ionic strength used one gap inserted in each. The individual (see above). A single maximum M was modelled domains satisfied validation observed in all the P(r) curves. This checks using PROCHECK, where the corresponded to the most frequent Ramachandran plots showed that 70% of interatomic distance within the FHR5 the residues were in the most favoured structure (Table 2). The mean M values steric regions. The FHR5 dimer was were 4.9 ± 0.3 nm, 4.9 ± 0.1 nm, and 5.4 ± generated from its monomer structure by 0.3 nm for Tris-150, PBS-137 and PBS-50 aligning its SCR-1/2 domains with the respectively. The M values were relatively crystal structure of the FHR1 SCR-1/2 stable, although slightly higher for PBS-50 dimer (Experimental Procedures), followed as the result of trace aggregates. by energy minimisation to relax this starting structure. Initial model for the FHR5 dimer Currently, there is no atomic level Modelling the solution structure of the structural information on FHR5. To FHR5 dimer determine an atomistic-level solution Atomistic modelling of the FHR5 structure for the FHR5 dimer, a starting scattering data established the best-fit model for the monomer was required. This FHR5 dimer structures, hence providing a was created by comparative modelling molecular explanation for its solution 5

structure. The scattering curves for 0.17 Searches 1-3, 86,732 structures with no mg/ml and 0.5 mg/ml FHR5 in Tris-150 steric clashes were accepted for Search 1, were used in order to assess good quality and likewise 72,755 structures for Search 2, curves with no traces of aggregation and and 123,776 structures for Search 3 (yellow better signal-noise ratios at 0.5 mg/ml in Figure 10). To verify the Monte Carlo- (Figure 9). Data for 0.5 mg/ml were not generated conformations, a grid density available in PBS-137 or PBS-50, and traces plot was generated for the Search 2 library of aggregates were present in PBS-50 of models (Figure 9). The volumetric data buffer, thus these data sets were not used. showed that a full conformational range of structures had been sampled, in comparison The starting structure for the FHR5 with the starting FHR5 dimer model at the dimer represented an extended centre of the grid. Significantly, the conformation of the 18 SCR domains experimental RG value of 5.36 nm occurred (Figure 9). Each SCR domain was held at the left of the distribution plots in Figure fixed in conformation. Because as many as 10, clearly indicating that FHR5 has a 14 linkers between the 18 domains were compact domain structure. In distinction, potentially variable, three different Monte linear FHR5 models showed higher RG Carlo conformational searches were set up. values of over 8 nm. Downloaded from http://www.jbc.org/ by guest on September 24, 2020 As detailed in Table 3, these varied all 14 linkers (Search 1), or eight linkers in which The three sets of 72,755-123,776 the crystal structure-observed linkers were models were each filtered to identify the kept fixed (Search 2), or four linkers after best-fit structures (Table 3). The every third SCR domain (Search 3) (Figure appearance of the RG vs. R-factor graphs 1B) (Experimental Procedures). Initial was similar in all six fits (Figure 10). This Monte Carlo conformational simulations in showed that the outcome of the modelling Searches 1-3 gave many models that were was independent of the assumption used to too elongated with too large RG values and generate the linkers. As required, the dimer few models with low RG values close to the models with the lowest R-factors of 4-5% experimental RG value of ~5.5 nm. Thus, in agreed well with the experimental RG value further simulations, models were selected of 5.36 nm. The most extended FHR5 with RG values closer to the experimental structures with the largest RG values of 8 nm RG value to generate further conformers, and above showed the highest R-factors of but now using an RG cut-off of 6.0 nm as ~30%. No models had an RG of 4.5 nm or constraint to generate more compact FHR5 less because such a dimer would be too dimers. This resulted in more structures compact to be sterically allowed. Filters with lower RG values; however, many of were now used to reject poor-fit structures. these models were rejected by the workflow First, a ± 5% experimental RG filter was because the more compact shapes gave rise used to reject models that had RG values to physically-disallowed steric clashes outside this range, followed by a ± 5% RXS between the SCR domains. filter. Models with an R-factor below 6% were then selected. For the two fits of All six analyses from the three Search 1 (Table 3), totals of 28 and 131 Searches at two FHR5 concentrations gave models were identified (green in Figure a clear single minimum in the distribution 10A). For Search 2, totals of 55 and 52 of R-factor goodness of fit values (Figure models were identified (Figure 10B). For 10). A lower R-factor indicated a better fit Search 3, totals of 694 and 749 models were to experiment. Thus all three Searches identified (Figure 10C). These best-fit successfully generated good-fit solution models formed a single cluster of fits at the structures for the FHR5 dimer. Starting R-factor minima. The best-fit models with from 200,000-250,000 trial structures in the lowest R-factors (red in Figure 10) had 6

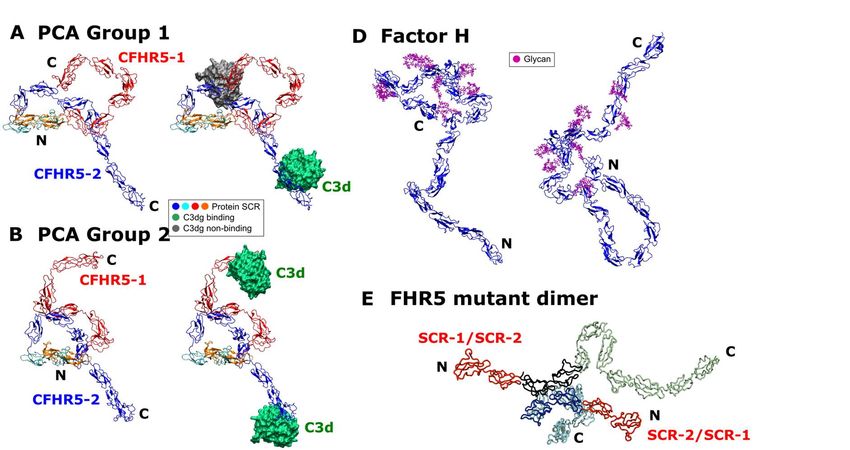
R-factors of 4.5% and 4.2% for Search 1, of this matrix each have an associated 4.7% and 3.9% for Search 2, and 4.3% and eigenvalue that characterises the clustering 3.8% for Search 3. For comparison, the of the models based on structural parameters for the best-fit 100 models were coodinates (or variance). By this, the first also shown in Table 3. three eigenvalue rankings (PC1 to PC3) accounted for a variance of 68.9% in the 55 Visual inspection of the fits between best-fit FHR5 models. The median FHR5 the theoretical and experimental SAXS I(Q) structure from each principal component and P(r) curves showed good agreement analysis group consistently revealed (Figure 11). The M and L values of the P(r) folded-back N-terminal domains and curves were well reproduced. Kratky plots extended C-terminal domains (Figure 14). of the SAXS curves monitor whether the protein was compact and globular or was Sedimentation coefficient modelling of the extended and disordered in its structure. FHR5 dimer The normalised Kratky analyses of As an independent test of the SAXS (Q.RG)2.I(Q)/I(0) vs Q.RG for the three best- modelling, the theoretical s20,w values were fit models from Searches 1-3 and the calculated using HYDROPRO for the best- experimental curve at 0.5 mg/ml showed fit FHR5 dimer models obtained from the Downloaded from http://www.jbc.org/ by guest on September 24, 2020 that a clear peak was seen at a Q.RG of 2.26 three Searches 1-3 (Table 3). The six best- (Figure 12). Good fits to the experimental fit models gave a mean s20,w value of 5.3 ± curve were also obtained at larger Q.RG 0.2 S. This compared well with the values for all three best-fit models. The experimental s20,w value in PBS-137 of 5.97 Kratky plot thus showed that FHR5 ± 0.2 S (Table 1). The typical accuracy of possessed a globular structure with little the s20,w calculation is ± 0.21 S (23). The inter-domain flexibility. In comparison, our difference of 0.67 S may result from recent Factor H models showed poorer fits potential trace aggregates remaining in the at larger Q.RG values, indicating that the 20 X-ray sample which would increase the SCR domains in Factor H had more experimental and modelled RG values of flexibility (12). This comparison indicated FHR5 and in turn decrease the modelled that the structure of FHR5 was well- s20,w value. formed, and this was less flexible in structure than full-length Factor H. Discussion Up to now, the domain organization Because all three searches gave of FHR5 was unknown. Here we present similar good fits, Search 2 was selected for the first protein structures for the FHR5 the final output because this most closely dimer by a combination of SAXS and AUC resembled the crystal structures for the SCR in conjunction with molecular simulations. domain pairs used to construct it. To Previously, it was often thought that FHR5 understand better the 55 best-fit structures possessed nine SCR domains in a flexible from Search 2 (available in Supplementary linear conformation (4,8,24-26). Instead, Materials), they were clustered into our analyses now show that FHR5 is conformational families using principal dimeric and adopts a compact domain component analysis (Figure 13) (21,22). conformation. Such a structure readily Principal component analysis determines leads to FHR5 oligomer formation in the the correlated motions of protein residues presence of mutant FHR5 protein (see as linearly uncorrelated variables termed below). This structure revises our principal components. These “essential understanding of how FHR5 interacts with motions” are extracted from a covariance its target ligand C3b and its C3d fragment, matrix of the atomic coordinates of the as well as others such as heparin-like frames in the trajectory. The eigenvectors 7
analogues. It also explains the molecular that the RG and RXS values for FHR5 were defect underlying CFHR5 nephropathy. relatively constant in 50-150 mM NaCl and between 0.1-0.5 mg/ml, although residual New understandings of the FHR5 trace aggregates were detectable in 50 mM solution structure were determined: NaCl buffer. The maximum length L of (i) Our SEC-MALLS and AUC data FHR5 was 20-21 nm in all buffers (Table showed that full-length FHR5 SCR-1/9 is a 2). A single SCR domain is about 4 nm in dimer (Figures 3 and 4), in agreement with length. A hypothetical fully-extended previous results for the FHR proteins FHR5 domain arrangement (Figure 1A) (4,6,7). In addition, AUC monitors would be predicted to be of length 64 nm, macromolecular shapes through the s20,w or over three-fold longer than seen values which measures macromolecular experimentally (Figure 8). Likewise Factor elongation. Of interest here was that, not H is predicted to be 80 nm in length if fully- only did the s20,w values correspond to a extended, but was observed to be only 26- much more compact protein than expected 29 nm in length, so again such an extended from the 18 domains in the dimer (Table 3), structure is also predicted to be three-fold but also these s20,w values decreased with an longer than seen experimentally (12). Both increase in the NaCl concentration of the FHR5 and Factor H thus have similar Downloaded from http://www.jbc.org/ by guest on September 24, 2020 buffer. This decrease implied that the folded-back domain structures. compact structure became more elongated (iii) Because no high resolution through the weakening of charge-charge FHR5 domain structures were available, the interactions between the SCR domains. The starting model for FHR5 was generated by predicted pI values of the N-terminal five standard homology modelling methods domains SCR-1/5 were mostly acidic at 4.6, based on sequence similarities. The FHR5 5.4, 8.5, 4.7 and 4.3 in that order, while the SCR-1/2 and SCR-8/9 structures were predicted pI values of the four C-terminal readily modelled on other FHR proteins. domains SCR-6/9 were mostly basic at 9.6, These modelled domain pairs were notable 6.3, 8.9 and 8.4 in that order for their short linker lengths of three (http://web.expasy.org/protparam/). residues each, suggesting that these linkers Differences in these individual pI values were relatively inflexible (Figure 1C). The may facilitate the formation of a more longest inter-SCR linkers occurred between compact FHR5 domain structure through SCR-3/7, which were six, six, eight and charge attractions in physiological 137 mM seven residues in length respectively. NaCl salt. Interestingly, the same linker lengths (ii) The SAXS data provided more occurred in SCR-10/14 of Factor H. In fact, detailed views of the FHR5 structure. sequence similarities showed that these five Interestingly, given that the SAXS SCR domains resembled SCR-3/7 of technique is sensitive to aggregate FHR5. These Factor H domains contributed formation, both FHR5 and Factor H turned significantly to its folded-back solution out to be aggregation-prone. The RG value structure (11,12,27,28). These long linkers of Factor H was originally reported to be in Factor H and FHR5 contained a high 12.4 nm in the first SAXS studies in 1991 proportion of charged residues, particularly for reason of being aggregated; with lysine and glutamate, and are conserved in improved Factor H purifications, this value mouse and bovine factor H (29). Indeed, has now diminished to 7.22-7.77 nm (12). SCR-10/14 of factor H, not only has longer Factor H aggregates in storage conditions. inter-domain linkers, but also shorter SCR FHR5 as supplied for our study showed sequences and higher glycosylation levels aggregation by SEC-MALLS and AUC, (30). These similarities imply that these and these aggregates were removed by size- middle domains act as conformational exclusion chromatography. SAXS showed spacers that result in more compact domain 8

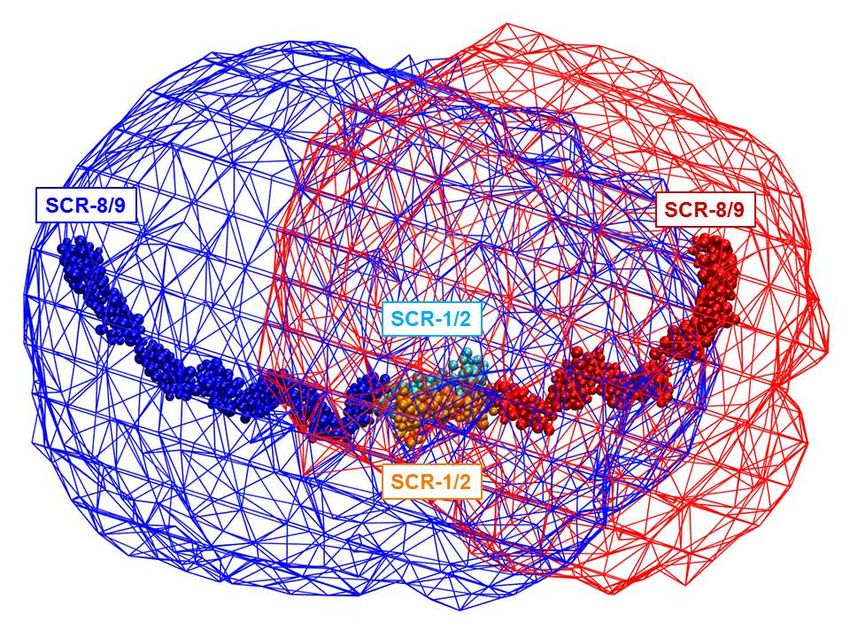
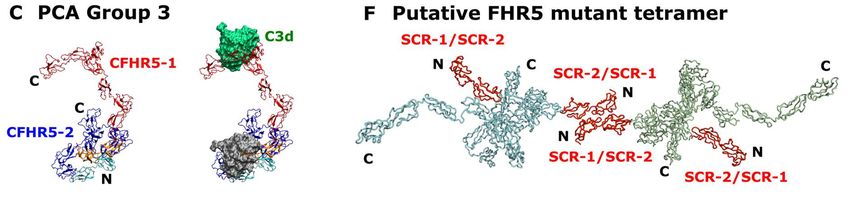
structures that enable the multiple factor H C3b/C3d on surfaces is low, the much more and FHR5 binding sites to act abundant Factor H will preferentially bind synergistically. to inhibit and degrade C3b there via Factor (iv) The Monte Carlo simulations I-mediated cleavage. When the density of generated a large conformational library of C3b/C3d is great enough to allow dimeric possible SCR arrangements in FHR5, from FHR5 binding to be functionally bivalent which best-fit structures were identified. (Figure 14B) the FHR5-C3d interaction These best-fit structures accounted for the becomes stronger than the monovalent experimental SAXS and AUC data for Factor H-C3d interaction. This reasoning FHR5. Interestingly the Kratky plots indicates a mechanism for FHR5 to (Figure 12) did not show evidence of modulate Factor H activity. disorder or flexibility in the FHR5 solution structure, meaning that its structure was In CFHR5 nephropathy, the well-defined. The molecular structures for heterozygous duplication of SCR-1/2 FHR5 and Factor H show similar folded results in a more elongated FHR5 molecule back and compact SCR structures (Figure that is detectable in the blood of patients 14A-D). From the principal component (Figure 14E,F) (8). Other heterozygous analyses, the three best-fit FHR5 structures genomic rearrangements that result in the Downloaded from http://www.jbc.org/ by guest on September 24, 2020 (Supplementary Materials) showed that, production of more elongated FHR proteins while the SCR-1/2 dimer pair was with additional N-terminal SCR-1/2 consistently buried in the dimer core in all domains have been described in association three structures, the two SCR-3/4 domain with autosomal dominant C3 pairs looped back across the SCR-1/2 core glomerulopathy (31,32). Conversely, clear in a compact arrangement with SCR-1/2. loss-of-function variants in CFHR5 occur at The two C-terminal ends with SCR-5/9 high frequency in the population and are not were solvent-exposed and either extended known to be pathogenic. Among the away from the SCR-1/4 core or looped back ~245,000 alleles tested in GnomAD towards this core. The functional SCR-8/9 (//gnomad.broadinstitute.org/gene/ENSG0 domains thus showed a range of folded- 0000134389), ~3000 variants predicted to back or extended conformations relative to stop FHR5 protein translation before the a more compact SCR-1/4 core. The three final exon are documented. In addition, 3% best-fit conformations were able to interact of the UK population is homozygous for a with one or two C3d ligands (Figure 14A- CFHR3/CFHR1 deletion polymorphism C). that results in the complete deficiency of FHR1 and FHR3 (33). Together, these In terms of new functional insight observations suggest that a gain-of-function obtained from this study, FHR5 is a mechanism underlies CFHR5 nephropathy complement deregulator that competitively and that tandem duplication of the two N- inhibits factor H, an important regulator of terminal SCR-1/2 domains is necessary and C3b activation at host cell surfaces (4,5). sufficient to cause this. Structural From the nine best fit structures (Table 3), simulations using our FHR5 models show the two C3d or C3b binding sites found in that the extra SCR-1/2 domains of the dimeric FHR5 will have a C-terminal mutant FHR5 protein are readily added, and separation of around 10-20 nm. FHR5 these will be accessible to other FHR5 would increase its avidity for C3d- or C3b- molecules (Figure 14E,F). At least two coated host cell surfaces only if bound C3d distinct mechanisms can be proposed by or C3b were present at a great enough which the mutation in CFHR5 nephropathy spatial density on this surface, thus causes augmented function (i.e. increased displacing the binding of the much more avidity for C3-coated surfaces). In one, as abundant Factor H. If the spatial density of proposed previously, the presence of two 9
accessible SCR-1/2 dimerization motifs on Mammalian-expressed (HEK293 the single mutant protein would allow cells) human FHR5 SCR-1/9 was trimers or higher order oligomers to form purchased from Creative Biolabs (Shirley, that would be tri- or multi-valent with NY, USA). This was prepared with a His respect to C3d (Figure 13E,F). In addition, tag which was cleaved off by the the greater length of macromolecules manufacturer. This protein was prone to containing mutant FHR5 would reduce the aggregation. Aggregate-free FHR5 for density of C3d on a host cell surface SAXS was successfully purified from required for multivalent binding to occur, approximately 1 mg of protein that was since the longer protein would have a pooled and concentrated using a Vivaspin greater steric range. 20 spin concentrator (Sartorius) with a 10 kDa molecular weight cut-off, then purified Overall, it is expected that different using a Superdex 200 10/300 GL gel- tissues will function differentially in respect filtration column (Cytiva) equilibrated in 50 of FHR5 or Factor H binding activity. It is mM Tris, 150 mM NaCl, 1 mM EDTA, pH possible that a high blood flow rate, such as 7.4, using a Gilson HPLC system kindly that in the renal glomeruli, enables the made available by Dr A.J. Beavil (Kings density of C3b or C3d deposition to become College London). The FHR5 concentration Downloaded from http://www.jbc.org/ by guest on September 24, 2020 high enough to allow FHR5 dimers to bind was checked by the absorbance reading at bivalently. This explains why FHR5 is 280 nm. Its purity and integrity was enriched in C3-coated glomeruli and why checked by SDS-PAGE before and after CFHR5 gain-of-function mutations result each SAXS and AUC experiment under in the purely renal disease of CFHR5 reducing and non-reducing conditions nephropathy, which manifests clinically at using a Novex® 8-12% Bis-Tris Gel 1.0 times of infection when the complement mm (Invitrogen, Paisley, UK). system is systemically activated. The striking clinical and histological similarity The amino acid composition of of IgA nephropathy to CFHR5 human FHR5 SCR-1/9 was determined nephropathy, combined with the co- from its sequence (SWISSPROT accession localization of FHR5 and C3 in the code: Q9BXR6). Two potential N-linked glomerulus in IgA nephropathy (1), raises glycan sites were present at Asn126 and the possibility that the FHR proteins, Asn400 (Figure 1A), and may be occupied including FHR5, play an important role in by biantennary glycans as reported for both diseases. This possibility is supported Factor H (30). However, there was no by the observation that, in IgA evidence that these sites are occupied, in nephropathy, a common polymorphic particular at Asn126, where glycan was not deletion of CFHR1 (which encodes the present in the crystal structure of HEK293- smaller dimeric complement deregulator expressed FHR1 SCR-1/2 (PDB code: FHR1) is protective (34). Our 3ZD2) (4). Since FHR1 SCR-1/2 has the demonstration of a compact FHR5 dimer same glycosylation sequence as that in structure at a molecular level therefore FHR5 (Figure 1C), glycosylation was reveals new aspects of how FHR5 disregarded here. The mass of glycan-free antagonises Factor H function, amplifying wild-type FHR5 was predicted to be 62,377 complement activation at host cell surfaces Da from its sequence. Using the program when C3 deposition reaches a critical SLUV (20), it has an unhydrated volume of density, and leading to renal damage. 79.76 nm3, a hydrated volume of 105.23 nm3, a partial specific volume of 0.7278 Experimental Procedures nm3, and an absorption coefficient of 15.59 Purification and composition of full- (1%, 280 nm, 1 cm path length). length FHR5 10
FHR5 samples were run through viscosity of 0.01002 cp was used SEC-MALLS. This determines protein throughout in the AUC analyses. molecular masses using a standard HPLC system equipped with a Superdex 200 Sedimentation velocity data collection and Increase 5/150 GL gel filtration column analyses for FHR5 (Cytiva). The instrument was equipped AUC data were obtained on a with three detectors, namely a miniDawn Beckman XL-I instrument, equipped with detector (Wyatt Technology) which is a an eight-hole AnTi50 rotor (Beckman- triple-angle light scattering detector, an Coulter Inc., Palo Alto, CA). Optilab DSP Interferometric Refractometer Ultracentrifugation caused any aggregates (Wyatt Technology) which measures present to sediment rapidly, leaving the refractive index changes, and an SPD-20A soluble FHR5 protein visible for analysis. UV absorbance detector (Shimadzu Approximately 400 µl of FHR5 sample was Scientific). In multiple runs, 60 µl aliquots loaded into standard AUC double-sector of FHR5 were loaded on the column via an cells for sedimentation velocity injection loop. Following separation by experiments at 20°C, equipped with size-exclusion, the three different detectors sapphire windows and with 12 mm column were combined in parallel to provide a heights. Sample concentrations were 0.16 Downloaded from http://www.jbc.org/ by guest on September 24, 2020 molecular mass for the eluted sample. The mg/ml, therefore absorbance optics was chromatograms were analysed using used to collect data. Up to 500 consecutive ASTRA software (Wyatt Technology). scans were recorded until the protein had fully sedimented. The AUC runs were For AUC and SAXS experiments, performed using two rotor speeds of 40,000 FHR5 was dialysed into the appropriate rpm and 50,000 rpm to check for buffer at 4°C prior to data collection. For reproducibility. AUC, these buffers were; PBS-20 (20 mM NaCl, plus 8.2 mM Na2HPO4, 2.6 mM KCl, Data analysis was performed using 1.5 mM KH2PO4, pH 7.4), PBS-50 with 50 SEDFIT software (version 14.6) (36, 37), mM NaCl, PBS-90 with 90 mM NaCl, using direct boundary Lamm fits of up to 50 PBS-137 with 137 mM NaCl (standard selected scans at appropriately spaced time physiological salt), PBS-250 with 250 mM intervals. A c(s) size-distribution analysis NaCl, and PBS-350 with 350 mM NaCl. was carried out, which assumes that all For SAXS, three buffers were used, namely species have the same frictional ratio f/f0. Tris-150 (50 mM Tris, 150 mM NaCl, 1 The c(s) distribution was optimized by mM EDTA, pH 7.4), and PBS-50 and PBS floating the value of the meniscus and 137 as above. The experimental buffer bottom of the cell positions, the baseline densities were measured at 20°C using an and the frictional ratio f/f0 (set at 1.2 to Anton Paar DMA 5000 density meter, and begin with). Fits were carried out until their theoretical values were calculated satisfactory visual fits and overall root from SEDNTERP (35). The resulting mean square deviations were obtained. The densities were 1.000538 g/ml for PBS-20 final SEDFIT analysis used a resolution of (theoretical, 1.00052 g/ml), 1.001714 g/ml 200, and the sedimentation coefficient s20,w for PBS-50 (theoretical, 1.00176 g/ml), for FHR5 was determined from the peak 1.003382 g/ml for PBS-90 (theoretical, maximum in the c(s) size-distribution plot. 1.00342 g/ml), 1.005054 g/ml for PBS-137 The c(s) integration function was also used (theoretical, 1.00524 g/ml), 1.009960 g/ml to derive the percentage of oligomers in the for PBS-250 (theoretical, 1.00999 g/ml), total loading concentration if required. 1.013920 g/ml for PBS-350 (theoretical, 1.01406 g/ml), and 1.00650 g/ml for Tris- SAXS data collection and data analyses 150 (theoretical, 1.00603 g/ml). A solvent for FHR5 11
SAXS experiments were carried out has no effect, and the forward scattering at in one beam session on the BM29 zero angle I(0). BioSAXS beamline at the European ln ( ) = ln (0) − Synchrotron Radiation Facility, Grenoble, France, operating with a ring energy of 6.0 GeV. Data was acquired using a Pilatus 1M (1) two-dimensional detector with a pixel size of 172 µm. The sample-to-detector distance ln[ ( ). ] = was 3.0 m. The beamline was equipped [ ( ). ] ⟶ − with an automatic sample changer, and the (2) samples were loaded using the thermo- regulated PCR tube configuration in the The Guinier plots are usually valid in a Q BsxCuBE control interface. The FHR5 range up to Q.RG values of 1.5 (39). If the samples were measured in three buffers macromolecular structure is elongated, the (above) at concentrations of 0.04 mg/ml, mean cross-sectional radius of gyration RXS 0.09 mg/ml, 0.13 mg/ml, and 0.17 mg/ml. is obtained from plots of I(Q).Q against Q2 Additional data sets were collected at 0.2 in a larger Q range than those used for the mg/ml, 0.3 mg/ml, 0.4 mg/ml, and 0.5 Downloaded from http://www.jbc.org/ by guest on September 24, 2020 RG values. Using the SCT software package mg/ml concentrations in Tris-150 buffer. (40), the Q ranges for the RG and RXS values Data was collected in triplicate from a total were 0.1 - 0.27 nm-1 and 0.32 – 0.55 nm-1 sample volume of 50 µl per run. An respectively. exposure time of 1 sec was used, and the absence of radiation damage was monitored Indirect Fourier transformation of from continuous automatic online checks. the scattering curve I(Q) in reciprocal space A total of 10 frames were collected as the (units in nm-1) into real space (units in nm) sample was passed continuously through a gives the distance distribution function quartz capillary tube (1.8 mm in diameter) P(r). This transformation was carried out to minimise radiation damage due to using the program GNOM (41). exposure. The final time-frames were ( ) = merged, excluding any damaged data, to improve the signal-to-noise ratio. Between ∫ ( ) sin( ) each sample measurement, the sample (3) capillary was cleaned using Hellmanex® and water to ensure the removal of any P(r) corresponds to the distribution of residual protein or aggregates on the interatomic distances r in the capillary walls (38). macromolecule. In order to obtain the distance distribution P(r) curve, the full The raw scattering data files were measured scattering curve was utilised. By corrected by subtraction of the buffer data specifying an assumed maximum from the sample data. The resulting one- dimension Dmax, the P(r) curve provides the dimensional scattering curve I(Q) in a Q- macromolecular length L and the most range between 0.05 nm-1 to 2 nm-1 (where common distance M. The P(r) curves also Q = 4π sin θ/λ; 2θ is the scattering angle and provide an alternative calculation of RG for λ is the wavelength) represented the comparison with the Guinier analysis. macromolecular structure. Guinier analysis of I(Q) against Q2 at low Q values gave the Generating of the starting model for radius of gyration RG, which is a measure of FHR5 structural elongation if the internal Protein structural analyses of FHR5 inhomogeneity of the scattering densities were initiated from homology models for the nine SCR domains. Firstly, suitable 12
templates were selected based on high The PDB file for the dimer of FHR5 sequence and structural similarities. This was generated by superimposing SCR-1/2 was achieved by a combination of PDB- of each FHR5 monomer model onto the BLAST searches and sequence alignments FHR1 SCR-1/2 dimer crystal structure between the five FHR-related proteins and (PDB code: 3ZD2), using PyMOL (DeLano Factor H. The final template was selected Scientific). This structure was inputted from the quality of the sequence alignment directly into the atomistic modelling and its structural relevance. The template workflow of the SASSIE scattering curve structures were taken from closely related fit package (19). First, the PDB file was structures in the Protein Data Bank (PDB). manually corrected for gaps or errors in the In the process, the amino acid sequence of amino acid sequence. A protein structure FHR5 SCR-1/9 was used to replace the file (PSF), which contained molecule- sequence of the template structure. These specific information for the application of a were constructed using closely-related force field, was generated via PSFGEN structural templates using MODELLER using Visual Molecular Dynamics (VMD) (version 9.15) (42). (version 1.9.2) (47). To create a physically- realistic atomistic model, the structure was The closest template for each of the subjected to 10 ps of energy minimisation Downloaded from http://www.jbc.org/ by guest on September 24, 2020 nine SCR domains, defined in terms of using the molecular dynamics simulation sequence identity and minimum insertions package NAMD (version 2.9) (47, 48). The and deletions, was identified using force field for this was CHARMM-36 (49, CLUSTALO alignments (43). Four 50) and energy minimisation was template structures for eight domains were performed using the conjugate gradient used as follows (Figure 1B,C); FHR1 SCR- method. 1/2 (PDB code: 3ZD2), Factor H SCR- 10/11 (PDB code: 4B2R), Factor H SCR- Molecular simulations and SAXS fitting 12/13 (PDB code: 2KMS) and FHR2 SCR- of FHR5 3/4 (PDB code: 3ZD1). The ninth domain By excluding the dimerization was SCR-7, for which a multiple sequence interface at SCR-1/2 and linker L1 which alignment (44) was performed using the do not vary in conformation (Figure 1A), NMR structures of Factor H SCR-10/11 FHR5 contains seven poentially flexible (PDB code: 4B2R) (28) and Factor H SCR- inter-SCR linkers L2 – L8 (Figure 1B). The 11/12 (PDB code: 4B2S) (28), which linkers were subjected to peptide dihedral provided an experimental structure for angle variations in the Monte Carlo Factor H SCR-11. The full-length FHR5 simulations through the Markov sampling model was evaluated using the SAVES of backbone torsion angles (19). This server allowed the rapid generation of a large (https://services.mbi.ucla.edu/SAVES/), conformational library of physically which incorporated validation criteria realistic atomistic models of the FHR5 including PROCHECK and Ramachandran SCR-1/9 dimer through the Complex plots. The secondary structure and surface Monte Carlo module of SASSIE. The same accessibilities of the FHR5 model were linkers on either monomer of the dimer analysed using the Definition of Secondary were varied independently of each other, Structure of Protein (DSSP) program (45). thus the resulting dimer structures were Structures were also modelled using asymmetric in shape. In Search 1, all seven SWISS-MODEL (46) to cross-check the linkers (L2 – L8) were varied. These were models from MODELLER using another defined as follows: L2 141SFTKGE146, L3 202 tool. KGQVRS207, L4 263VEQVKT268, L5 323 VATHQLKR330, L6 382TEKREQF388, L7 443 VESTAY448, and L8 504LDP506. In 13
Search 2, only L2, L4, L6 and L7 were based on their RG and RXS values as well as varied, because these were not part of the their goodness-of-fit R-factor values crystal structures that were used (Figure defined as: 1B). In Search 3, only Linkers L3 and L6 were varied as a control of Searches 1 and = 2. This strategy of independent simulations ∑ ( ) ‖ ( )‖ (Table 3) checked whether extra or fewer ∑ ( ) × 100 constraints in the linkers affected the (4) resulting best-fit structures. During the Monte Carlo simulations, models with where I Expt (Q ) and I Theor (Q) were the steric overlaps that were generated by SASSIE were excluded by specifying an experimental and theoretically calculated atomic overlap distance cut-off of 0.3 nm. scattered intensities, and was a scaling Simulations were continued to produce factor used to match the theoretical and models with RG values close to that of 6.0 experimental I(0) values. Typical best-fit R- nm obtained experimentally by filtering for factors for SAXS modelling are between a fixed range of RG values in the FHR5 2% and 8% (14). To visualise the initial and dimer models. The outputted structures best fit models for the FHR5 dimer, density Downloaded from http://www.jbc.org/ by guest on September 24, 2020 were generated as binary format DCD files plots were generated using the Density Plot and visualised on VMD. In the three module in SASSIE. The envelope was searches, a total of up to 250,000 models generated for the sterically-accepted trial were generated in order to sample a models, sampled to produce the volumetric sufficient number of conformations for the data, using the Gaussian cube file format. two monomers in the dimer. This was superimposed onto the initial FHR5 dimer model. The output files were Using the SCT module (40) in rendered, analysed and annotated in VMD. SASSIE, a theoretical scattering curve was Once the best-fit dimer models were calculated for each of the FHR5 dimer chosen, their sedimentation coefficients models. The atomic coordinates were were calculated for comparison with the converted into small spheres to generate a AUC data, based on the atomic coordinates coarse-grained sphere model. A cube side using the HYDROPRO shell modelling length of 0.53 nm in conjunction with a cut- program (53). off of four atoms was used to generate unhydrated sphere models. Because the Data Availability Statement hydration shell was visible by X-rays, a All data are contained within this hydration shell containing 0.3 g of H2O/g of manuscript. The 55 best-fit models from protein was added to each of the models by Search 2 and the 6 best-fit structures from HYPRO (51). The theoretical scattering Figure 11 are available in Supplementary curve I(Q) for each model was calculated Materials. using the Debye equation adapted to _______________________________ spheres (40, 52). Acknowledgements We thank Dr P. Pernot (ESRF, Grenoble) The theoretical scattering curves for for excellent X-ray instrumental support, the dimer models were compared to the Dr David W. Wright for expert experimental SAXS curves. In the SCT computational support, and Dr Andrew J. Analyse module of SASSIE, the RG and RXS Beavil (King’s College London) for expert values were calculated from the modelled assistance with SEC-MALLS. Initial work curves using the same Q ranges that were in this project won an award for Best Poster used for the experimental Guinier analyses. Presentation at the XXVIth International The curve fits were compared and filtered 14
Complement Workshop in Kanazawa, Conflict of interest: The authors declare Japan 4-8 September 2016. that they have no conflicts of interest with the contents of this article. References 1. Murphy, B., Georgiou, T., Machet, D., Hill, P., and McRae, J. (2002) Factor H-related protein-5: a novel component of human glomerular immune deposits. Am. J. Kidney Dis. 39, 24-27. 2. McRae, J.L., Duthy, T.G., Griggs, K.M., Ormsby, R.J., Cowan, P.J., Cromer, B.A., McKinstry, W.J., Parker, M.W., Murphy, B.F., and Gordon, D.L. (2005) Human factor H-related protein 5 has cofactor activity, inhibits C3 convertase activity, binds heparin and C-reactive protein, and associates with lipoprotein. J. Immunol. 174, 6250-6256. 3. Soares, D., and Barlow, P. N. (2005) Complement control protein modules in the regulators of complement activators. In Structural Biology of the complement system (Morikis D & Lambris J.D., eds), pp. 19-62, Taylor & Francis, Boca Raton, USA. 4. Goicoechea de Jorge, E., Caesar, J.J., Malik, T.H., Patel, M., Colledge, M., Johnson, S., Hakobyan, S., Morgan, B.P., Harris, C.L., Pickering, M.C., and Lea, S.M., (2013) Downloaded from http://www.jbc.org/ by guest on September 24, 2020 Dimerization of complement factor H-related proteins modulates complement activation in vivo. Proc. Natl. Acad. Sci. USA. 110, 4685–4690. 5. Jokiranta, T.S., Jaakola, V.P., Lehtinen, M.J., Pärepalo, M., Meri, S. and Goldman, A. (2006) Structure of complement factor H carboxyl-terminus reveals molecular basis of atypical haemolytic uremic syndrome. EMBO J., 25, 1784-1794. 6. Van Beek, A. E., Pouw, R. B., Brouwer, M. C., Rispens, V. T. Kuijper, T. W., and Wouters, D. (2016) Dimerization of Complement Factor H-related (FHR) proteins: FHR-5 forms homodimers whereas FHR-1 and FHR-2 both homodimerize and heterodimerize with each other. Immunobiology 221,1182-1183. 7. Ruseva, M. M., Malik, T. H., and Pickering, M. C. (2016) Insights into the role of FHR5 in C3 glomerulopathy. Immunobiology 221,1167. 8. Gale, D.P., Goicoechea de Jorge, E., Cook, H.T., Martinez-Barricarte, R., Hadjisavvas, A., McLean, A.G., Pusey, C.D., Pierides, A., Kyriacou, K., Athanasiou, Y., Voskarides, K., Deltas, C., Palmer, A., Fremeaux-Bacchi, V., de Cordoba, S.R., Maxwell, P.H., and Pickering, M.C. (2010) Identification of a mutation in complement factor H-related protein 5 in patients of Cypriot origin with glomerulonephritis. Lancet 376, 794–801. 9. Athanasiou, Y., Voskarides, K., Gale, D.P., Damianou, L., Patsias, C., Zavros, M., Maxwell, P.H., Cook, H.T., Demosthenous, P., Hadjisavvas, A., Kyriacou, K., Zouvani, I., Pierides, A., and Deltas, C. (2011) Familial C3 glomerulopathy associated with CFHR5 mutations: clinical characteristics of 91 patients in 16 pedigrees. Clin J Am Soc Nephrol. 6,1436-1446. 10. DiScipio, R. G. (1992) Ultrastructures and interactions of complement factors H and I. J. Immunol. 149, 2592-2599. 11. Okemefuna, A. I., Nan, R., Gor, J., and Perkins, S. J. (2009) Electrostatic interactions contribute to the folded-back conformation of wild type human factor H. J. Mol. Biol. 391, 98-118. 12. Osborne, A. J., Nan, R., Miller, A., Bhatt, J. S., Gor, J., and Perkins, S. J. (2018) Two distinct conformations of factor H regulate discrete complement-binding functions in the fluid phase and at cell surfaces. J. Biol. Chem. 293, 17166-17187. 13. Perkins, S. J., Nan, R., Li, K., Khan, S., and Abe, Y. (2011) Analytical Ultracentrifugation combined with X-ray and neutron scattering: Experiment and modelling. Methods. 54, 181-199. 15
14. Perkins, S. J., Wright, D. W., Zhang, H., Brookes, E. H., Chen, J., Irving, T. C., Krueger, S., Barlow, D. J., Edler, K. J., Scott, D. J., Terrill, N. J., King, S. M., Butler, P. D. and Curtis, J. E. (2016) Atomistic modelling of scattering data in the Collaborative Computational Project for Small Angle Scattering (CCP-SAS). J. Appl. Crystallogr. 49, 1861-1875. 15. Perkins, S. J., Nan, R. D., Li, K. Y., Khan, S., and Miller, A. (2012) Complement Factor H-ligand interactions: Self-association, multivalency and dissociation constants. Immunobiology 217, 281-297. 16. Perkins, S.J., and Goodship, T.H. (2002) Molecular modelling of the C-terminal domains of factor H of human complement: a correlation between haemolytic uraemic syndrome and a predicted heparin binding site. J. Mol. Biol. 316, 217-224. 17. Saunders, R. E., Abarrategui-Garrido, C., Frémeaux-Bacchi, V., Goicoechea de Jorge, E., Goodship, T. H. J., López Trascasa, M., Noris, M., Ponce Castro, I. M., Remuzzi, G., Rodríguez de Córdoba, S., Sánchez-Corral, P., Skerka, C., Zipfel, P. F. & Perkins, S. J. (2007). The interactive Factor H - atypical Haemolytic Uraemic Syndrome mutation database and website: Update and integration of Membrane Cofactor Protein and Factor I mutations with structural models. Human Mutation, 28, 222-234. 18. Rodriguez, E., Rallapalli, P. M., Osborne, A. J., and Perkins, S. J. (2014) New Downloaded from http://www.jbc.org/ by guest on September 24, 2020 functional and structural insights from updated mutational databases for complement factor H, Factor I, membrane cofactor protein and C3. Bioscience. Rep. 34, 635-649. 19. Curtis, J. E., Raghunandan, S., Nanda, H., and Krueger, S. (2012) SASSIE: A program to study intrinsically disordered biological molecules and macromolecular ensembles using experimental scattering restraints. Comput. Phys. Commun. 183, 382-389. 20. Perkins, S. J. (1986) Protein volumes and hydration effects. The calculation of partial specific volumes, neutron scattering matchpoints and 280-nm coefficients for proteins and glycoproteins from amino acid sequences. Eur. J. Biochem. 157, 169-180. 21. Hui, G. K., Wright, D. W., Vennard, O. L., Rayner, L. E., Pang, M., Yeo, S. C., Gor, J., Molyneux, K., Barratt, J., and Perkins, S. J. (2015) The solution structures of native and patient monomeric human IgA1 reveal asymmetric extended structures: implications for function and IgAN disease. Biochem. J. 471, 167-185 22. David, C. C. and Jacobs, D. J. (2014) Principal component analysis: a method for determining the essential dynamics of proteins. Methods Mol. Biol. 1084, 193-226 23. Perkins, S. J., Okemefuna, A. I., Nan, R., Li, K., and Bonner, A. (2009) Constrained solution scattering modelling of human antibodies and complement proteins reveals novel biological insights. J. Royal Soc. Interface 6, S679-S696. 24. Holers, V. M. (2013) Human C3 glomerulopathy provides unique insights into complement factor H–related protein function. J. Clin. Invest. 123, 2357-2360. 25. Tortajada, A., Yébenes, H., Abarrategui-Garrido, C., Anter, J., García-Fernández, J.M., Martínez-Barricarte, R., Alba-Domínguez, M., Malik, T.H., Bedoya, R., Cabrera Pérez, R., López Trascasa, M., Pickering, M.C., Harris, C.L., Sánchez-Corral, P., Llorca, O. and Rodríguez de Córdoba, S. (2013) J. Clin. Invest. 6, 2434-2446. 26. Józsi, M., and Meri, S. (2014) Factor H-related proteins. The Complement System: Methods and Protocols, 225-236. 27. Schmidt, C. Q., Herbert, A. P., Mertens, H. D., Guariento, M., Soares, D. C., Uhrin, D., Rowe, A. J., Svergun, D. I., and Barlow, P. N. (2010) The central portion of factor H (modules 10-15) is compact and contains a structurally deviant CCP molecule. J. Mol. Biol. 395, 105-122. 28. Makou, E., Mertens, H.D., Maciejewski, M., Soares, D.C., Matis, I., Schmidt, C.Q., Herbert, A.P., Svergun, D.I., and Barlow, P.N. (2012) Solution structure of CCP 16
modules 10–12 illuminates functional architecture of the complement regulator, factor H. J. Mol. Biol. 424, 295-312. 29. Aslam, M., and Perkins, S. J. (2001) Folded-back solution structure of monomeric Factor H of human complement by synchrotron X-ray and neutron scattering, analytical ultracentrifugation and constrained molecular modelling. J. Mol. Biol. 309, 1117-1138. 30. Fenaille, F., Le Mignon, M., Groseil, C., Ramon, C., Riande, S., Siret, L., and Bihoreau, N. (2007) Site-specific N-glycan characterization of human complement factor H. Glycobiology 17, 932-944. 31. Tortajada, A., Yébenes, H., Abarrategui-Garrido, C., Anter, J., García-Fernández, J.M., Martínez-Barricarte, R., Alba-Domínguez, M., Malik, T.H., Bedoya, R., Cabrera Pérez, R., López Trascasa, M., Pickering, M.C., Harris, C.L., Sánchez-Corral, P., Llorca, O., and Rodríguez de Córdoba S. (2013) C3 glomerulopathy-associated CFHR1 mutation alters FHR oligomerization and complement regulation. J. Clin. Invest. 123, 2434- 2446. 32. Chen, Q., Wiesener, M., Eberhardt HU, Hartmann A, Uzonyi B, Kirschfink M, Amann K, Buettner M, Goodship T, Hugo C, Skerka C, Zipfel PF. (2014) Complement factor H-related hybrid protein deregulates complement in dense deposit disease. J. Clin. Invest. 124, 145-155. Downloaded from http://www.jbc.org/ by guest on September 24, 2020 33. Holmes, L. V., Strain, L., Staniforth, S. J., Moore, I., Marchbank, K., Kavanagh, D., Goodship, J. A., Cordell, H. J. & Goodship, T. H. J. (2013) Determining the population frequency of the CFHR3/CFHR1 deletion at 1q32. PLoS ONE 8, e60352. 34. Kiryluk, K., Li, Y., Scolari, F., Sanna-Cherchi, S., Choi, M., Verbitsky, M., Fasel, D., Lata, S., Prakash, S., Shapiro, S., Fischman, C., Snyder, H.J., Appel, G., Izzi, C., Viola, B.F., Dallera, N., Del Vecchio, L., Barlassina, C., Salvi, E., Bertinetto, F.E., Amoroso, A., Savoldi, S., Rocchietti, M., Amore, A., Peruzzi, L., Coppo, R., Salvadori, M., Ravani, P., Magistroni, R., Ghiggeri, G.M., Caridi, G., Bodria, M., Lugani, F., Allegri, L., Delsante, M., Maiorana, M., Magnano, A., Frasca, G., Boer, E., Boscutti, G., Ponticelli, C., Mignani, R., Marcantoni, C., Di Landro, D., Santoro, D., Pani, A., Polci, R., Feriozzi, S., Chicca, S., Galliani, M., Gigante, M., Gesualdo, L., Zamboli, P., Battaglia, G.G., Garozzo, M., Maixnerová, D., Tesar, V., Eitner, F., Rauen, T., Floege, J., Kovacs, T., Nagy, J., Mucha, K., Pączek, L., Zaniew, M., Mizerska-Wasiak, M., Roszkowska-Blaim, M., Pawlaczyk, K., Gale, D., Barratt, J., Thibaudin, L., Berthoux, F., Canaud, G., Boland, A., Metzger, M., Panzer, U., Suzuki, H., Goto, S., Narita, I., Caliskan, Y., Xie, J., Hou, P., Chen, N., Zhang, H., Wyatt, R.J., Novak, J., Julian, B.A., Feehally, J., Stengel, B., Cusi, D., Lifton, R.P., and Gharavi, A.G. (2014) Discovery of new risk loci for IgA nephropathy implicates genes involved in immunity against intestinal pathogens. Nat Genet. 46, 1187-1196. 35. Laue, T. M., Shah, B. D., Ridgeway, T. M., and Pelletier, S. L. (1992) Computer-aided interpretation of analytical sedimentation data for proteins. In Analytical Ultracentrifugation in Biochemistry and Polymer Science, (Harding, S. E., Rowe, A. J., and Horton, J. C., eds), pp. 90-125, The Royal Society of Chemisty, Cambridge, U.K. 36. Schuck, P. (1998) Sedimentation analysis of non-interacting and self-associating solutes using numerical solutions to the Lamm equation. Biophys. J. 75, 1503-1512. 37. Schuck, P. (2000) Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and Lamm equation modeling. Biophys. J. 78, 1606-1619. 38. Pernot, P., Round, A., Barrett, R., Antolinos, A. D., Gobbo, A., Gordon, E., Huet, J., Kieffer, J., Lentini, M., Mattenet, M., Morawe, C., Mueller-Dieckmann, C., Ohlsson, S., Schmid, W., Surr, J., Theveneau, P., Zerrad, L., and McSweeney, S. (2013) Upgraded ESRF BM29 beamline for SAXS on macromolecules in solution. J. Synchrotron Radiat. 20, 660-664. 17
You can also read

















































