Genetic sensor regulators functional in Clostridia - Oxford ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
https://doi.org/10.1007/s10295-020-02303-6
GENETICS AND MOLECULAR BIOLOGY OF INDUSTRIAL ORGANISMS - ORIGINAL
PAPER
Genetic sensor‑regulators functional in Clostridia
SongI Han1 · Younghwan Kim2 · Mukund Karanjikar2 · Ka‑Yiu San1,3 · George N. Bennett3,4
Received: 9 June 2020 / Accepted: 18 August 2020 / Published online: 26 August 2020
© Society for Industrial Microbiology and Biotechnology 2020
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
Abstract
This study addressed the functionality of genetic circuits carrying natural regulatory elements of Clostridium acetobutyli-
cum ATCC 824 in the presence of the respective inducer molecules. Specifically, promoters and their regulators involved in
diverse carbon source utilization were characterized using mCherryOpt or beta-galactosidase as a reporter. Consequently,
most of the genetic circuits tested in this study were functional in Clostridium acetobutylicum ATCC 824 in the presence of
an inducer, leading to the expression of reporter proteins. These genetic sensor-regulators were found to be transferable to
another Clostridium species, such as Clostridium beijerinckii NCIMB 8052. The gradual expression of reporter protein was
observed as a function of the carbohydrates of interest. A xylose-inducible promoter allows a titratable and robust expression
of a reporter protein with stringency and efficacy. This xylose-inducible circuit was seen to enable induction of the expres-
sion of reporter proteins in the presence of actual sugar mixtures incorporated in woody hydrolysate wherein glucose and
xylose are present as predominant carbon sources.
Keywords Sensor · Regulator · Clostridium acetobutylicum · mCherryOpt · Beta-galactosidase
Introduction scale by anaerobic fermentation [28]. The metabolism of
this organism can be characterized as two distinct phases
Clostridium acetobutylicum ATCC 824 (C. acetobutylicum initiated from the acidogenic phase, followed by the solven-
ATCC 824) is a gram-positive and spore-forming obligate togenic phase [1]. Rapid cell growth is observed in the aci-
anaerobe [38]. It has been known to metabolize a wide vari- dogenic phase producing acetate and butyrate accompanied
ety of carbon sources to produce acetone, butanol, ethanol by a pH decrease, which triggers the solventogenic phase
(ABE), and other chemicals [36]. Depending on the type of where ABE is produced, leading to the uptake of acids and
carbon sources supplemented, it reveals a distinct solvent a pH increase [44].
production profile. Butanol has a high energy content as well An advantage of Clostridia as a host organism derives
as low vapor pressure, and it can be manufactured on a large from their ability to employ diverse carbon sources, includ-
ing mono-, oligo- and polysaccharides that would be found
in waste products [13], making the fermentation of indus-
Electronic supplementary material The online version of this trial, agricultural and waste products conceivable [32]. Pro-
article (https://doi.org/10.1007/s10295-020-02303-6) contains cessing of waste products to useful compounds by biological
supplementary material, which is available to authorized users.
fermentation has gained much attention since it would have
* George N. Bennett the possibility of reducing the cost of feedstocks [10]. In
gbennett@rice.edu general, biomass and waste-based feedstocks are presumed
1
to contain a mixture of sugars [37]. For example, hemicel-
Department of Bioengineering, Rice University, Houston,
TX 77005, USA
luloses consist of primary monomeric subunits such as glu-
2
cose, xylose, mannose, galactose, rhamnose, and arabinose
Technology Holding LLC, West Valley City, UT 84119, USA
[3] while dried distiller’s grains contain a mixture of glu-
3
Department of Chemical and Biomolecular Engineering, cose, xylose, arabinose, and mannose [7]. Alongside cel-
Rice University, Houston, TX 77005, USA
lulosic biomass, food waste holds nutrients such as amino
4
Department of BioSciences, Rice University, Houston, acids, fatty acids, and minerals to support culture growth as
TX 77005, USA
13
Vol.:(0123456789)610 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
well as containing significant amounts of sugars and starch genetic circuits responding to different carbon sources. A
[10]. Fermentation of carbohydrate mixtures presumably broad dynamic range of detection and low endogenous level
incorporated in waste products may rely on the regulatory in the strain of interest are critical features to be considered
response of the organism to carbohydrate and nutrient avail- as a reporter protein [11]. A lacZ from Thermoanaerobac-
ability [32]. To increase the efficiency of ABE fermentation terium thermosulfurigenes (encoding beta-galactosidase)
of carbohydrate blends, in-depth knowledge of the organ- [38] was exploited to demonstrate an assay as we have used
ism’s capacity to respond to, take up, and utilize carbon this reporter previously [31]. While many fluorescent pro-
sources is essential. In this regard, identification of key teins are restricted to use in aerobic environments due to
sensor-regulators is required for the improved utilization of their requirement of oxygen for chromophore maturation
waste and eventually allow butanol production to be eco- [6], other anaerobic fluorescent proteins have been reported
nomically feasible in Clostridia. If sensors have the capa- (evoglow-Pp1 which is fluorescent under both aerobic and
bility of responding to small amounts of sugar derived from anaerobic conditions [15], FMN-based fluorescent protein
polymers and then leading to the production of enzymes independent of oxygen [11, 33, 34]). Among them, mCher-
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
responsible for directing metabolic flux to butanol formation ryOpt [30] with suitable maturation was used in this study
by metabolic engineering, they would be very beneficial in as one of the reporter proteins. The assay can be conducted
the efficient utilization of waste products. It necessitates the with a fluorescence plate reader to show fluorescence inten-
investigation of genetic elements acting as promoters and sity levels.
their regulators in response to the substances incorporated Herein, a set of sensor-regulators responding to differen-
in waste products, particularly carbon sources. An increasing tial carbon sources was characterized. Their functionality
number of studies are addressing Clostridia related inducible was evaluated quantitatively by the reporter proteins. The
promoters and their application relevant to carbon sources, genetic circuit proven to be superior regarding sensitivity
which in part would allow circumventing the deleterious and selectivity compared to other circuits has been further
effects of inducer toxicity inherent in the Ptet system [23, explored for gradual induction of reporter proteins as a
30]. Inducible promoters, for example, xylose-inducible pro- function of inducer as well as enabling transfer of traits to
moters, have been developed in Clostridia, including one other organisms as genetic tools as regulatory mechanisms
assessed with a β-glucuronidase reporter exhibiting 17-fold controlling gene expression differ from one bacterium to
induction with xylose [8]. Tightly regulated expression of another [8]. Notably, the sensitivity of the xylose-inducible
the chloramphenicol acetyltransferase reporter was dem- circuit was assessed in the presence of actual mixed sugars
onstrated in Clostridium perfringens where regulation of incorporated in a woody biomass hydrolysate.
expression exploited a xylose-regulatory system of Clostrid-
ium difficile [24]. A recent study established the genetic tools
of CRISPR interference by using xylose as an inducer in C. Materials and methods
difficile [23] along with demonstrating the uniform and tun-
able expression of xylose-inducible heterologous reporter Strains and culture conditions
proteins. A xylose-inducible transposon system was devel-
oped, allowing highly efficient random mutagenesis in C. The strains and plasmids used in this study are shown in
acetobutylicum ATCC 824 [51]. Beyond xylose-inducible Table 1. The plasmid pAN1 was utilized for the methyla-
promoters, carbohydrate-inducible promoters have been elu- tion of pSOS94 originated vectors, which is mediated by
cidated, including arabinose [49] and lactose [2, 9, 47] that the B. subtilis phage ϕ3TI methyltransferase as previously
are endogenous promoters in Clostridia. described [19]. E. coli strains were cultivated in Luria–Ber-
We are interested in testing promoters for the practical tani (LB) medium supplemented with appropriate antibiotics
development of controllable gene expression. In this study, at 37 °C for the amplification of plasmids. C. acetobutyli-
an attempt was made to find functional sensor-regulators in cum ATCC 824 strains were cultivated in clostridial growth
C. acetobutylicum ATCC 824 that respond to diverse sig- medium (CGM) consisting of 2 g (NH4)2SO4, 0.5 g KH2PO4,
nal molecules, in this case, carbon sources, and some of 1 g K2HPO4, 0.01 g MnSO4·2H2O, 0.1 g MgSO4·7H2O,
the candidates were chosen based on previously conducted 0.015 g FeSO4·7H2O, 0.01 g C aCl2, 0.02 g C
oCl2, 0.02 g
omics research [32, 40]. The transcriptional analysis of dif- ZnSO4, 2 g tryptone, 1 g yeast extract, and 20 g glucose
ferential carbohydrate utilization helped the design and con- (glucose as default carbon source unless otherwise stated)
struction of sensor-regulators in this study by tabulating the per liter in an anaerobic chamber (Coylab, MI, US) at 37 °C.
differential expression pattern of genes under the influence For the transformation, recombinant strains were grown
of different carbohydrates [32]. Each of the genetic circuits anaerobically on 2 × YTG (10 g yeast extract, 16 g tryptone,
has a reporter protein, either an enzymatic protein or a fluo- 5 g glucose, 4 g NaCl per liter, pH 5.8) agar plates sup-
rescent protein, to confirm the functionality of the designed plemented with 40 μg/ml of erythromycin at 37 °C [28]. A
13Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620 611
Table 1 Strains and plasmids used in this study
Strain or plasmid Relevant characteristics Source or references
Clostridium acetobutylicum ATCC 824 Wild type ATCC [17]
Clostridium beijerinckii NCIMB 8052 Wild type Supplied by Dr. Yi Wang [48]
Escherichia coli DH10β F− mcrA Δ(mrr-hsdRMS-mcrBC) φ80lacZΔM15 Invitrogen
ΔlacX74 recA1 endA1 araD139 Δ(ara-leu)7697 galU galK
λ− rpsL(StrR) nupG
−
Escherichia coli DH5α F φ80lacZΔM15 Δ(lacZYA-argF) Invitrogen
U169 recA1 endA1 hsdR17(rK−, mK+) phoA supE44
λ− thi-1 gyrA96 relA1
Escherichia coli MG1655 Wild type (F− λ−) ATCC 47,076
pAN1 p15A ori, Cmr ϕ3TI gene Mermelstein et al. 1993 [20]
pSOS94 ColE ori, pIM13 ori, Apr MLSr P_ptb ctfA/B adc Soucaille and Papoutsakis, unpublished
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
pSBX1 XylR_CAC3673-P-CAC2611-Thermoanaerobacterium This study
thermosulfurigenes lacZ
pSBX2 XylR_CAC3673-P_CAC3451-TtlacZ This study
pSBX3 XylR_CAC3673-P_Ccel1133-TtlacZ This study
pSBF1 FruR_CAC0231-P_CAC0231-TtlacZ This study
pSBF2 FruR_CAC0231-P_CAC0234-TtlacZ This study
pSBA1 AraR_CAC1340-P_CAC1341-TtlacZ This study
pSBA2 AraR_CAC1340-P_CAC1343-TtlacZ This study
pSBS1 LicT_CAC0422-P_CAC0423-TtlacZ This study
pSBM1 CcpA_V302N_P_CAC0532-3_TtlacZ This study
pSMX1 XylR_CAC3673-P_CAC2611-mCherryOpt This study
pSMX2 XylR_CAC3673-P_CAC3451-mCherryOpt This study
pSMX3 XylR_CAC3673-P_Ccel1133-mCherryOpt This study
pSMF1 FruR_CAC0231-P_CAC0231-mCherryOpt This study
pSMF2 FruR_CAC0231-P_CAC0234-mCherryOpt This study
pSMA1 AraR_CAC1340-P_CAC1341-mCherryOpt This study
pSMA2 AraR_CAC1340-P_CAC1343-mCherryOpt This study
pSMS1 LicT_CAC0422-P_CAC0423-mCherryOpt This study
pSMM1 CcpA_V302N_P_CAC0532-3_mCherryOpt This study
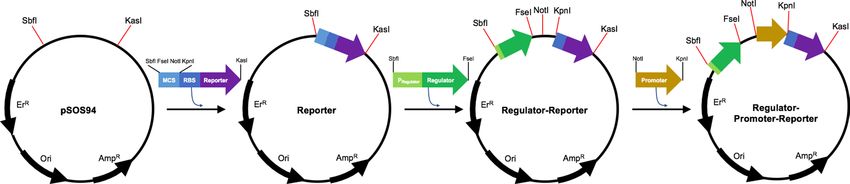
stock of spores of C. beijerinckii NCIMB 8052 was gener- with the traditional restriction enzyme cloning method,
ously granted by Dr. Yi Wang (Auburn University) and was which is placed in an order of SbfI-FseI-NotI-KpnI-reporter
cultivated by using CGM. gene-KasI. Then, the chemically synthesized regulator gene
was inserted into the plasmid carrying reporter genes by
Plasmid construction using SbfI and FseI. Lastly, chemically synthesized pro-
moters were individually cloned into the plasmid harboring
All oligonucleotide primers synthesis, gene synthesis, and regulator-reporter genes by using NotI and KpnI (Fig. 1).
DNA sequencing were conducted by GenScript Ltd (Piscata-
way, NJ, USA). For the construction of a list of genetic cir- Transformation
cuits, a modular genetic assembly was conducted where each
genetic part can be easily switched by specific restriction The recombinant plasmids were electro-transformed
enzymes. The pSOS94 plasmid, which can be replicated in into C. acetobutylicum ATCC 824, following the improved
both organisms such as E. coli and C. acetobutylicum ATCC protocol reported previously [21, 28]. The C. acetobu-
824, was used as a backbone plasmid. The gene sequence tylicum ATCC 824 cells were incubated at 37 °C under
of optimized beta-galactosidase or mCherryOpt, which is anaerobic conditions until the OD600 reached 0.6–0.8
a reporter protein, was chemically synthesized and cloned (logarithmic phase). Then the cultures were centrifuged
into the pSOS94 vector by using SbfI and KasI. Additional and washed twice using cold ETM buffer (270 mM sucrose,
restriction enzyme sites were inserted upstream of the 0.6 mM Na2HPO4·12H2O, 4.4 mM N aH2PO4·2H2O, 10 mM
reporter gene to make the next cloning process compatible MgCl2·6H2O), and the cells were resuspended in 1–2 ml ET
13612 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
Fig. 1 Plasmid construction consisting of a regulator, promoter, and reporter protein by using restriction enzymes
buffer (270 mM sucrose, 0.6 mM Na2HPO4·12H2O, 4.4 mM 50 ml tubes. Inoculum was typically 10% (v/v) of a CGM
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
aH2PO4·2H2O). The plasmids were mixed with the com-
N grown pre-culture. An artificial sugar mixture containing dif-
petent cells and electro-transformed under conditions of ferent glucose (G) to xylose (X) ratios (50:0, 40:10, 30:20,
2000 V, 25 μF of capacitance, and infinite Ω of resistance 25:25, 20:30, 10:40, and 0:50 G:X g/l) was supplemented
(4 mm cuvette). After incubating in a pre-warmed 2 × YTG to CGM for investigating the gradual expression of reporter
overnight, cells were spread onto a 2 × YTG agar plate sup- proteins by xylose-inducible circuits. The initial total sugar
plemented with 40 µg/ml erythromycin and incubated at concentration was kept at 50 g/l.
37 °C under anaerobic conditions. The identification of the Woody biomass hydrolysate containing both glucose
transformants containing the plasmid was performed by the and xylose as major carbon sources was supplemented to
method of colony PCR [27] by using corresponding primers the medium for testing the response of the xylose-inducible
for the beta-galactosidase gene or mCherryOpt gene partial genetic circuit in the presence of actual waste products.
fragment. Alternatively, the plasmids were extracted and Woody biomass hydrolysate was obtained from NREL,
purified using a modified standard mini-prep protocol [18]. wherein 480 mM glucose and 330 mM xylose (36.5%
The isolated plasmids were then re-transformed into E. coli. xylose) were characterized to be present as predominant
Extracted plasmids from E. coli were confirmed by restric- carbon sources. The fermentation media examined include
tion enzymes digestions. The identical procedure was dem- CGM supplemented with (1) 13 g/l glucose + 7 g/l xylose;
onstrated for the electroporation of C. beijerinckii NCIMB (2) woody biomass hydrolysate containing 20 g/l total sugars
8052. (36.5% xylose); (3) 32 g/l glucose + 18 g/l xylose; (4) woody
biomass hydrolysate containing 50 g/l total sugars (36.5%
Batch fermentation xylose). Other carbon sources were analyzed to be negligent
in woody biomass hydrolysate.
For all scales of fermentations, CGM was used (glucose as
default carbon source unless otherwise stated). Erythromy- Beta‑galactosidase assay
cin (80 µg/ml) was added to maintain a selection pressure
for plasmid harboring strains. A single colony selected from The beta-galactosidase activity was measured as described
freshly transformed recombinants was grown overnight at by Thakker et al. [35] with the following modifications. Ten
37 °C in the medium supplemented with erythromycin for ml of culture broth was centrifuged at 2000g for 10 min at
securing seed culture. For the experiments with different 4 °C and washed once by suspending the pellet with chilled
kinds of carbohydrate addition, glucose was replaced with Z-buffer. One ml of Z-buffer (60 mM Na2HPO4·7H2O,
the respective carbon source. The modified CGM supple- 40 mM NaH2PO4·H2O, 10 mM KCl, 1 mM MgSO4·7H2O,
mented with 20 g/l of the respective carbon source depend- 50 mM β-mercaptoethanol, pH 7.0) was used for re-sus-
ing on the type of genetic circuits or CGM with glucose as a pension of cells, and OD600 was measured. Dilution was
control in the presence of erythromycin was used for anaero- made 1:10 in a total of 1 ml Z-buffer and mixed with 100 μl
bic cultivations. A 4% (v/v) of a CGM grown pre-culture chloroform and 50 μl 0.1% SDS followed by brief vortex-
at OD600 = 1.0–2.5 was inoculated into the modified CGM, ing. After 10 min of incubation at 60 °C, 0.2 ml of ONPG
and fermentation was carried out at 37 °C for 24 h. Sam- (4 mg/ml) was added, and the suspension was briefly vor-
ples were taken for further assay (beta-galactosidase assay texed and incubated further at 60 °C for 1 h. The reaction
or fluorescence detection). pH was not adjusted throughout was stopped by the addition of 0.5 ml of 1 M N
a2CO3. After
the fermentation. centrifugation, OD420 and OD550 values were measured
The fermentation with glucose and xylose mixtures as using the supernatant. Beta-galactosidase activity in Miller
substrates was carried out in a 25 ml working volume in units was calculated as 1000 × [(OD420-1.75 × OD550)]/
13Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620 613
(t × v × OD600) where t is the time in minutes, v is the vol- The second part is composed of the regulated promoter by
ume of culture used in the assay in milliliters [22]. its associated small molecule (carbon source) and its cor-
responding regulator, and the third part is the ribosome
Measurement of mCherryOpt fluorescence binding site with the reporter gene. For endowing the inter-
changeable features between each genetic cassette conveni-
mCherryOpt fluorescence was measured as described by ently for future use, restriction enzyme cut sites were thor-
Ransom et al. [30] with the following modifications. The oughly explored and designed to be compatible between
culture broth was taken outside from the anaerobic chamber genetic parts. Carbon sources to be exploited in this study
after 24 h of incubation at 37 °C and 500 µl of culture broth were determined based on the composition of sugar blends
was centrifuged at 13,000g for 5 min at RT, and cells were in waste products. Xylose, fructose, arabinose, sucrose, and
washed three times by suspending the pellet in 1 ml 1 × PBS maltose were examined in this work. For the regulator part,
(137 mM NaCl, 2.7 mM KCl, 10 mM Na2HPO4, and 1.8 mM XylR (CAC3673), FruR (CAC0231), AraR (CAC1340),
KH2PO4, pH 7.4). The final cell pellet was suspended in LicT (CAC0422), or CcpA V302N (CAC0531 mutant)
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
30 µl of 1 × PBS. For chromophore maturation, the cell sus- derived from the genome sequence of C. acetobutylicum
pension was stored in the dark at RT or 4 °C. mCherryOpt ATCC 824 was chosen for xylose, fructose, arabinose,
fluorescence can generally be detected within 30 min and sucrose, and maltose regulator, respectively. The promoters
reaches a maximum in about 3 h [29]. After chromophore regulated by the corresponding carbon source are listed in
maturation, 170 µl of 1 × PBS was added, and the entire Table 2. The codon-optimized sequence of mCherryOpt or
200 µl of cell suspension was transferred to a single well of beta-galactosidase was inserted downstream of the promoter
a 96-well plate. (Fig. 1).
A microplate reader (Tecan Infinite M200 Pro plate In C. acetobutylicum ATCC 824, three operons were
reader) was used for monitoring optical density and red fluo- discovered to be induced by xylose not arabinose in a tran-
rescence. For mCherryOpt, the following software settings scriptomics study while showing negligible expression when
were used as described by Ransom et al. [30]: OD600 with fed with glucose [32]. An operon, including CAC2611 and
the number of flashes = 25; fluorescence excitation = 554, CAC2610, encoding an unidentified hypothetical protein and
emission = 610, mode = top, Z-position = 20,000 µm, num- a fucose isomerase, respectively, was found to be induced
ber of flashes = 25, gain = 100 manual, and integration by xylose. The second operon includes xynB (CAC3451)
time = 20 µs. Microplates compatible with the reader were and xynC (CAC3452), encoding a putative xylose symporter
used (Advangene, CC Plate-PS-96S-F–C-S). and a xylosidase, respectively. These two xylose-inducible
promoters were employed to construct xylose cassettes 1
Analytical methods and 2, respectively. Xylose promoter 3 was derived from
the Clostridium cellulolyticum chromosome sequence used
A previously established analytical method was used [28]. in the study of promoter screening with fluorescent protein
Residual carbon sources and the primary fermentation [34]. This promoter was characterized to be a xylan-induci-
products including acetate, butyrate, ethanol, acetone, and ble promoter and was found to have a relatively higher fluo-
butanol were determined using a Shimadzu 20A LC Sys- rescence intensity of the reporter protein in the presence of
tem high-performance liquid chromatography (Shimadzu xylose compared to other carbon sources tested following
Corporation, KS, USA) with a refractive index detector
(RID-10A), and equipped with an Aminex HPX-87H HPLC
column (Bio-Rad Laboratories Inc., CA, USA). A 5 mM Table 2 Controlled promoters, regulatory protein genes, and effectors
H2SO4 solution was used as the mobile phase at a flow rate of expression
of 0.5 ml/min, and the column was kept at 30 °C. In addi- Promoters Regulatory protein Small
tion, cell density was analyzed by determining the OD600 molecule
using a spectrophotometer (Shimadzu UV-2600, TX, USA). effector
CAC2610-12, XylR (CAC3673) Xylose
CAC3451-2
Results and discussion Ccel1133
CAC0231, 0234 FruR (CAC0231) Fructose
Design and construction of the genetic circuits CAC1341-2 AraR (CAC1340) Arabinose
CAC1343
Genetic cassettes consist of three main parts (regulator- CAC0423-5 LicT (CAC0422) Sucrose
promoter-reporter gene). The regulator element consists of CAC0532-3 CcpA (CAC0531) V302N Maltose
mutant
the regulatory protein coding gene and its native promoter.
13614 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
xylan addition to the media [34]. Teng et al. (2015) showed and its application as a reporter in anaerobic bacteria has
this promoter to have approximately five times higher fluo- been described increasingly [23, 30]. The use of mCherry-
rescence intensity when grown on xylose than that on cel- Opt benefits from its features such as brighter fluorescence
lobiose, which is the routinely used carbon source for C. and faster maturation, as well as less interference [30]. For
cellulolyticum cultivation [34]. Additionally, the reporter testing the functionality of these genetic circuits, the car-
protein abundance and fluorescence intensity were closely bon source of interest was supplemented to the medium
correlated (R2 = 0.87) [34]. In the absence of xylose, the for each individual genetic circuit accordingly. At the same
transcription of the xylose-inducible promoters is engineered time, glucose was used as a control carbon source in par-
to be repressed by XylR (CAC3673) for all three constructs. allel cultures. The initial concentration of carbon sources
The promoter sequences of the operon carrying CAC0231 was controlled at 20 g/l for this experiment. The cultiva-
(DeoR-type transcriptional regulator) and CAC0234 (PTS tion of cells harboring distinct genetic cassettes was per-
IIBC) were examined for a fructose study, respectively. formed concurrently to be comparable for analysis. After
These promoters were established to be under the control 24 h fermentation, samples were taken outside of the anaero-
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
of fructose regulator FruR (CAC0231) [39]. For arabinose bic chamber to allow the chromophore of mCherryOpt to
promoters, the operon encompassing the gene responsible mature in the presence of oxygen for more than 3 h, which
for an arabinose utilization pathway, CAC1341 (araD), was has been proven to be a suitable amount of time required for
selected. It has been reported that the transcription of the the maximum chromophore maturation [30]. Fixation was
pentose phosphate pathway genes is also controlled by AraR omitted because maintaining cellular architecture seems to
(CAC1340). Among them, the xfp gene (CAC1343, phos- be not essential in this study. After chromophore maturation,
phoketolase) that was the most prominently induced gene optical density and fluorescence intensity of mCherryOpt
in an AraR inactivated mutant, was chosen for this study were measured by using a microplate reader. As a result,
to generate another type of arabinose genetic circuit [50]. all the genetic circuits tested in this work led to the expres-
For sucrose, the promoter sequence of CAC0423 (scrA) was sion of the reporter protein in response to the carbon source
manipulated to be under the control of the LicT (CAC0422) of interest. They showed higher fluorescence intensity in
regulator. The promoter regulating the CAC0532-3 operon comparison to glucose as control (Fig. 2). The fold-change
was fused to the mutated catabolite repression protein CcpA of fluorescence intensity of xylose-inducible cassettes is
V302N to construct the maltose-inducible circuit. A com- three-fold (X1), 300-fold (X2), and five-fold (X3) higher
plex set of sugar metabolic genes is affected by CcpA, and a when induced, respectively (Fig. 2). The dynamic range of
genetically modified CcpA enables all the sugars, including expression varied significantly depending on the sequence
xylan, starch, arabinosides, arabinogalactan, and sucrose to of the promoters, although they are all under the control of
be readily utilized by alleviating catabolite repression [42]. the same carbon source as an inducer, where the 50-fold dif-
Therefore, this regulator with the mutation was chosen for ference was observed between pSMX2 and pSMX3 in the
constructing a genetic circuit responding to maltose. The presence of xylose (Fig. 2). Among xylose cassettes, pSMX2
sequence information of genetic parts we used is described exhibited a significant difference in fluorescence intensity
in supplementary material 1 in detail. Most of the sequences in the presence of xylose and glucose (300-fold change).
were originated from the C. acetobutylicum ATCC 824 This stringency and efficacy suggest its potential use in other
genome sequence database (Biocyc) (Table 2). Clostridia strains since it is highly activated upon induction
and efficiently repressed in the absence of an inducer.
It is noted that most of the promoters seem to be tightly
Genetic circuit function test by mCherryOpt regulated in the absence of an inducer. On the other hand,
fructose circuits exhibited high leakage without fructose as
Each regulated cassette under the control of the indicated an inducer. A considerable basal expression from the un-
carbon source is annotated as X1, X2, X3, F1, F2, A1, A2, induced promoter was observed for both fructose promoters.
S1, and M1 according to the type of promoter (xylose (X), This might be due to the level of repressor protein expres-
fructose (F), arabinose (A), sucrose (S), and maltose (M)). sion being inadequate for the regulation. Basal expression
One of the reporter proteins, mCherryOpt, which has been without an inducer could be mitigated by supplying a higher
beneficial to use in anaerobic bacteria, was inserted down- level of repressor [25]. Alternatively, there may be cross-
stream of each promoter [30]. Fluorescent reporter proteins talk between operons toward diverse carbon sources in C.
are useful to examine the gene expression because adding an acetobutylicum ATCC 824, as seen in the transcriptional
exogenous substrate or permeabilizing cells are not neces- expression study [32], which might account for the induction
sary for determining the level of fluorescent proteins [14]. of fructose promoter in the absence of fructose. Arabinose
Particularly, mCherryOpt has been codon-optimized to be constructs also showed good stringency of the promoter
expressed in low G + C content bacteria such as Clostridium, in the absence of inducer, yielding a 60-fold and a 21-fold
13Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620 615
Fig. 2 Fluorescence intensity of
mCherryOpt reporter after over-
night maturation. Anaerobic
culture of individual recom-
binant cells was taken outside
the anaerobic chamber for
chromophore maturation which
requires oxygen. Glucose was
used as a control carbon source.
The respective carbon source
was supplemented instead of
glucose for each genetic circuit,
accordingly. Fructose respond-
ing genetic circuits appear not
stringent due to leaky expres-
sion of mCherryOpt in the
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
presence of glucose. X: xylose,
F: fructose, A: arabinose, S:
sucrose, and M: maltose. Error
bars represent SD of tripli-
cate experiments. *p < 0.05,
**p < 0.01, ***p < 0.001 vs. the
glucose supplemented control
as a sole carbon source
elevated fluorescence intensity for pSMA1 and pSMA2 analyzed with a list of promoters required for sugar uptake
when induced, respectively (Fig. 2). A 21-fold or an 11-fold and utilization in parallel with those using the mCherryOpt
induction was achieved in the presence of sucrose or malt- reporter protein. This reporter protein is useful because C.
ose for pSMS1 or pSMM1, respectively (Fig. 2). However, acetobutyliucm ATCC 824 lacks endogenous beta-galac-
sucrose and maltose did not exhibit fluorescence intensity tosidase activity [46] so that the expression of heterologous
levels comparable with that found with the xylose, fructose, beta-galactosidase in the presence of an inducer would
and arabinose promoters in the presence of a carbon source allow verification of the functionality of genetic circuits
of interest as an inducer. Increasing the concentration of more sensitively with low background activity [38]. Before
carbon source seems to be one strategy to expand its opera- liquid broth fermentation, a simple colorimetric method by
tional range. using X-gal in the presence of the decisive inducer was car-
Herein, the fluorescence intensity profile, which would ried out on agar plates. However, it was not successful in
presumably correlate to the reporter protein expression, is detecting the color change of the substrate on an agar plate
in agreement with the transcriptional analysis study of dif- in the presence of an inducer very qualitatively. Therefore,
ferential carbohydrate utilization conducted by Servinsky a large-scale liquid culture was used to generate an appro-
et al. [32]. This is because promoters examined in this study priate amount of enzyme sufficient to metabolize the sub-
were selected in the context where significant up-regulation strate. The analogous method exploited in the cultivation
of RNA expression units was found when growing on a car- of strains carrying the mCherryOpt circuit was used in the
bon source of interest compared to glucose. It is noted that cultivation of strains harboring the beta-galactosidase cir-
there was very little difference in RNA expression when cuit. After 24 h of fermentation, samples were taken out,
comparing cells grown on glucose and fructose [32], which and the cells were subjected to beta-gal assay to determine
led to a certain level of gene expression when growing on the response of each promoter. The beta-gal assay was per-
glucose, consistent with a relatively high background level formed at 60 °C without the sonication of harvested cells.
of a reporter protein in this study. This result indicates that A couple of constructs exhibited a notable fold-change
fructose is among the preferred carbohydrates for this organ- in Miller units in the presence of an inducer compared to
ism [32]. the control carbon source, glucose. These constructs were
pSBX1, pSBX2, and pSBF2 (two xylose constructs and one
Genetic circuit function test by beta‑galactosidase fructose construct). These constructs were verified to func-
tion efficiently in Clostridium acetobutylicum ATCC 824
The expression of lacZ originated from Thermoanaero- in comparison to the rest of the constructs. The beta-gal
bacterium thermosulfurigenes EM1 as a reporter gene was assay revealed analogous behavior of constructs with genetic
13616 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
circuits carrying mCherryOpt, confirming the induction of tested. Both species are recognized as model organisms for
a reporter protein in the presence of an inducer. Both xylose the industrial production of ABE.
cassettes led to the 60-fold increased beta-galactosidase The identical procedure was applied for the characteri-
activity in a reporter assay, with the fructose circuit being zation of genetic circuits with mCherryOpt as a reporter
less than a two-fold increase (Fig. 3). As shown in Fig. 3, protein in C. beijerinckii NCIMB 8052. Simultaneous
xylose sensing circuits were observed to be more stringent fermentation of strains harboring each genetic circuit was
in the absence of inducer since Miller units in the presence carried out. Consequently, it was possible to use the same
of glucose appeared to be negligible compared to the value toolkit in another Clostridium species, although the level of
in xylose media. However, the fructose sensing circuit was fold change in Clostridium beijerinckii NCIMB 8052 was
not tightly regulated by repressor in the presence of glucose, slightly less than in C. acetobutylicum ATCC 824. It was
which correlates with the result of the fluorescence signal of observed that the fluorescence intensity increases in the
the corresponding mCherryOpt genetic circuits, exhibiting presence of carbon sources, indicating the proper function-
relatively small fold-change difference. Surprisingly, other ing of genetic circuits in Clostridium beijerinckii NCIMB
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
genetic circuits carrying beta-galactosidase as a reporter 8052 except for pSMX1. As shown in Fig. 4, the utiliza-
protein did not lead to the development of yellow color in tion of xylose as the sole carbon source led to a 20-fold
the process of the beta-gal assay. It might be relevant to the and a two-fold higher mCherryOpt expression and its cor-
limited suitability of these circuits with a specific type of responding fluorescence signal for pSMX2 and pSMX3,
reporter protein, or mCherryOpt might be more sensitive respectively. The plasmids pSMA1 and pSMA2 resulted
than beta-galactosidase. in a 32-fold and a 27-fold higher fluorescence signal when
induced by arabinose, respectively (Fig. 4). As observed in
Transferrable traits of genetic circuits to other C. acetobutylicum ATCC 824, fructose responding circuit
organisms pSMF2 exhibited leaky background expression, which led
to only a two-fold higher fluorescence signal when induced
Genetic tools and methodologies are frequently strain- by fructose (Fig. 4). Overall, the relative fold change pat-
dependent within Clostridia [26]. To explore the compat- tern was shown to be similar to that of C. acetobutylicum
ibility and portable traits of the genetic toolkits to another ATCC 824. Also, this genetic toolkit was confirmed to be
Clostridia species besides the base strain C. acetobutylicum compatible with E. coli and showed enhancement of expres-
ATCC 824, Clostridium beijerinckii NCIMB 8052 was sion upon induction with the corresponding carbon sources,
Fig. 3 Miller units of genetic circuits after beta-galactosidase assay.
Anaerobic cultures of individual recombinant cell lines were taken
outside the anaerobic chamber for beta-gal assay. Glucose was used Fig. 4 Fluorescence intensity of mCherryOpt reporter after overnight
as a control. The respective carbon source was supplemented instead maturation from C. beijerinckii NCIMB 8052. FI/OD measured from
of glucose for each genetic circuit. Miller units measured from two two xylose responding circuits (pSMX2, X3), one fructose respond-
xylose responding circuits and one fructose responding circuits are ing circuit (pSMF2), and two arabinose responding circuits (pSMA1,
illustrated (pSBX1, X2, and pSBF2). Error bars represent SD of trip- A2) is illustrated. Error bars represent SD of triplicate experiments.
licate experiments. *p < 0.05, **p < 0.01, ***p < 0.001 vs. the glucose *p < 0.05, **p < 0.01, ***p < 0.001 vs. the glucose supplemented con-
supplemented control as a sole carbon source trol as a sole carbon source
13Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620 617
which is comparable with the signal shown in C. acetobu- for the repressor encoded by xylR gene in the mixture of glu-
tylicum (data not shown). cose and xylose [12]. This phenomenon was also observed
It would be expected that these genetic circuits would in the beta-galactosidase assay with pSBX2 bearing cells. In
perform better in the organism in which the correspond- such situations, the remaining non-glucose sugar may not be
ing sensor-regulators are absent as shown in the study dem- completely used until glucose is completely consumed even
onstrated by Nariya et al. [24], thereby indicating that the with these inducible constructs unless other modifications
xylose-responding circuits could be applied to C. perfrin- are made to the host. This catabolic repression effect would
gens lacking xylose metabolism genes, for instance. be anticipated to be attenuated if the maltose-inducible cir-
cuit was tested in the mixture of glucose and maltose as the
Mixed sugar experiment mutant CcpA was applied as a repressor protein, thereby the
concomitant carbon utilization would likely be seen.
The next step was to determine the performance of genetic
circuits in C. acetobutylicum as host in the presence of Fermentation of woody hydrolysate as a substrate
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
diverse carbon sources together. Mixed sugar fermentations
are frequently used to mimic the composition of hydrolysates The further approach for verifying the compatibility of these
with synthetic sugars [4]. In this regard, carbon source mix- circuits with actual mixed sugars from the waste stream/
tures were supplemented to the medium for the characteriza- biomass hydrolysate as a substrate was examined in C. ace-
tion of particular genetic circuits. For this study, pSMX2 and tobutylicum as host. Glucose and xylose are abundant sugar
pSBX2 genetic circuits were characterized in the presence of monomers in lignocellulosic hydrolysates. It has been ana-
altered ratios of glucose and xylose to analyze the function- lyzed that the medians of total sugars, glucose, and xylose
ality of xylose circuits. Total sugar concentration was estab- are 41.8, 23.6, and 10.8 g/l, respectively, for lignocellulosic
lished as 50 g/l. A gradual increase of xylose proportion hydrolysates [4]. The deviation from the general trend, high
was demonstrated by setting up the ratio of sugar mixtures in xylose, was reported in soybean hull hydrolysis yield-
as 50G, 40G10X, 30G20X, 25G25X, 20G30X, 10G40X, ing 50% xylose (49 g/l glucose and 48 g/l xylose) [45],
and 50X g/l (G: glucose, X: xylose). Then, the expression and horticultural waste with 90% xylose (6 g/l glucose and
of the reporter protein was measured for cells exposed to the 58 g/l xylose) [43]. Herein, actual mixed sugars from woody
range of xylose concentrations from 0 to 50 g/l. The signal hydrolysate in a sample obtained from NREL was tested to
of the reporter protein increased in a stepwise manner as a evaluate the capacity of strain to be able to express reporter
function of xylose for both genetic circuits with different protein in the presence of the corresponding carbon source
reporter proteins, as shown in Fig. 5. A drastic increase of in the waste stream. Woody hydrolysate obtained from
fluorescence intensity was observed when 50 g/l of xylose NREL was characterized as containing 480 mM glucose and
was supplemented as a sole carbon source compared to the 330 mM xylose (36.5% xylose) as major sugar components.
other conditions where glucose and xylose were fed concur- Sugars in the woody hydrolysate were diluted with CGM
rently for pSMX2. This could be explained since glucose to contain a total of 20 g/l or 50 g/l. The identical ratio of
exerts catabolite repression and exhibits a direct influence glucose:xylose (total of 20 g/l, 12.8 g/l of glucose and 7.2 g/l
on reporter protein expression by acting as an anti-inducer of xylose or total of 50 g/l, 32 g/l glucose and 18 g/l of
Fig. 5 Mixed sugar fermentation with genetic circuits responding to 30G20X, 25G25X, 20G30X, 10G40X, and 50X g/l (G: glucose, X:
xylose in C. acetobutylicum as host. a mCherryOpt. b Beta-galac- xylose). Error bars represent SD of triplicate experiments. *p < 0.05,
tosidase as a function of xylose. Total sugar concentration was estab- **p < 0.01, ***p < 0.001 vs. the glucose supplemented control as a
lished as 50 g/l and a gradual increase of xylose concentration was sole carbon source
demonstrated by setting up the ratio of sugar mixtures: 50G, 40G10X,
13618 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
xylose) was supplemented for artificial mixed sugar fer- up to approximately 5 OD600 at 24 h. The two conditions
mentation as a comparison. The hydrolysate components where 20 g/l of total sugar was supplemented were found to
exhibit variability [41, 52] due to the difference in the initial consume all glucose. The profile of metabolites, such as ace-
composition of the biomass and pretreatment process used. tate, ethanol, acetone, butyrate, and butanol, was shown to
The woody hydrolysate used in this work is rich in xylose be comparable to each other at 24 h. At 24 h, the strains fed
(36.5%). with 20 g/l of sugars in woody hydrolysate produced 0.82,
For implementing this experiment, the pSMX2 genetic 2.07, and 0.47 g/l of ABE, respectively, while the strains
circuit was explored. Samples were taken after 24 h of fer- fed with pure sugar mixtures produced 0.61, 2.04, 0.45 g/l
mentation for the measurement of the fluorescence intensity of ABE, respectively. Additional incubation did not increase
of the reporter protein. The strain harboring pSMX2 fed with the xylose utilization and butanol production much for the
20 g/l of artificial sugar mixtures exhibited a 1.5-fold higher 20 g/l of total sugar fermentation (data not shown). The sup-
fluorescence intensity compared to the one fed with 20 g/l plementation of 50 g/l of total sugar led to higher production
of sugar in woody hydrolysate, indicating analogous induc- of ABE. Although the initial total substrate concentrations
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
tion response between two substrates. The toxicity arising of woody hydrolysate and mixed sugar fermentations were
from various inhibitors potentially present in hydrolysate almost the same (50 g/l), the latter had 16% higher ABE
likely accounts for the adverse effect on the reporter protein solvents at 48 h. This difference has been found due with
expression and its corresponding fluorescence intensity [5, higher butanol production in pure mixed sugar fermentation
16]. This phenomenon was also observed with 50 g/l of total wherein 5.53 g/l of butanol was produced while 4.15 g/l of
sugar supplementation, showing a 1.2-fold higher fluores- butanol was produced in the woody hydrolysate fermentation
cence intensity of pure mixed sugar compared to sugar in (Table 3). The toxicity of woody hydrolysate likely accounts
woody hydrolysate (data not shown). The fluorescence inten- for inferior performance as has generally been observed.
sity is placed in descending order from pure mixed sugar Given genetic circuits were functional in actual sugar
50 g/l, woody hydrolysate 50 g/l, pure mixed sugar 20 g/l, mixtures with varying concentrations, making a cassette
and woody hydrolysate 20 g/l. The low sugar concentration consisting of genetic parts having different inducer response
may cause a very weak induction effect on these promoters. and with a metabolic gene that can improve butanol produc-
This drawback could be alleviated by the addition of xylose tion in a Clostridium strain such as adhE1 or adhE2 gene
to increase the ratio of xylose: glucose, which is a common modifications would be the next stage of work. Alternatively
practice to increase the total sugar concentration with glu- or concurrently, the regulator of the maltose-inducible cir-
cose addition in many fermentations with hydrolysates [4]. cuit, which is the mutant of CcpA protein, could likely be
In alignment with reporter protein fluorescence intensity linked to other carbon source-inducible circuit as the effector
measurement, the extracellular metabolites were analyzed protein as a modified CcpA was exemplified to allow a hand-
(Table 3). No significant differences were found in the pro- ful of sugars to be readily utilized by alleviating catabolic
file of metabolites between four conditions until 24 h. With- repression [42]. The use of the CcpA mutant may be use-
out further modification, the diauxic growth was observed ful for an organism subject to exposure of sugar mixture as
wherein cells preferentially utilize glucose during fermenta- opposed to the preferential glucose metabolism displayed
tion when both glucose and xylose are available. Under all by wild type strains. Once suggested genetic cassettes are
conditions, strains exhibited comparable growth, reaching proven to be efficient in butanol production in the presence
Table 3 Sugars consumption and solvents production by C. acetobutylicum ATCC 824 harboring pSMX2 fermented in CGM supplemented with
the total sugar of woody hydrolysate 20 g/l, 50 g/l or mixed sugar 20 g/l, 50 g/l (glucose and xylose mixture)
Time (h) Total sugar (g/l) Glucose uti- Xylose uti- Acetone (g/l) Butanol (g/l) Ethanol (g/l) ABE (g/l)
Xylose 36.5% lized (g/l) lized (g/l)
24 h Hydrolysate 20 g/l 12.6 0.23 0.82 ± 0.00 2.07 ± 0.02 0.47 ± 0.00 3.36
Mixed Sugar 20 g/l 12.5 0.24 0.61 ± 0.06 2.04 ± 0.03 0.45 ± 0.00 3.10
Hydrolysate 50 g/l 11.68 0 1.14 ± 0.09 2.22 ± 0.14 0.45 ± 0.00 3.96
Mixed Sugar 50 g/l 10.39 0 0.70 ± 0.08 2.01 ± 0.02 0.42 ± 0.00 3.13
48 h Hydrolysate 20 g/l 12.8 0.51 0.84 ± 0.19 2.22 ± 0.10 0.51 ± 0.03 3.57
Mixed Sugar 20 g/l 12.8 1.72 0.51 ± 0.30 2.14 ± 0.45 0.48 ± 0.06 3.13
Hydrolysate 50 g/l 18.8 0 2.37 ± 0.61 4.15 ± 0.20 0.90 ± 0.12 7.41
Mixed Sugar 50 g/l 24.4 0 2.33 ± 0.82 5.53 ± 0.33 0.77 ± 0.09 8.63
Data are means ± standard deviations
13Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620 619
of a mixture of carbon sources such as glucose and xylose, reporter system and an inducible promoter-repressor system for
various waste product streams can be more carefully exam- Clostridium acetobutylicum. Appl Environ Microbiol 69:4985–
4988. https://doi.org/10.1128/aem.69.8.4985-4988.2003
ined as a feedstock. 9. Hartman AH, Liu H, Melville SB (2011) Construction and char-
acterization of a lactose-inducible promoter system for con-
trolled gene expression in Clostridium perfringens. Appl Envi-
Conclusions ron Microbiol 77:471–478. https://doi.org/10.1128/AEM.01536
-10
10. Huang H, Singh V, Qureshi N (2015) Butanol production from
We have characterized a set of inducible promoters natively food waste: a novel process for producing sustainable energy and
found in Clostridia species. Genetic cassettes were con- reducing environmental pollution. Biotechnol Biofuels 8:147.
structed by exploiting genetic parts that have been shown https://doi.org/10.1186/s13068-015-0332-x
11. Joseph RC, Kim NM, Sandoval NR (2018) Recent developments
to be functional in response to diverse carbon sources in C. of the synthetic biology toolkit for Clostridium. Front Microbiol
acetobutylicum ATCC 824. The genetic circuits constructed 9:154. https://doi.org/10.3389/fmicb.2018.00154
exhibited differential fluorescence intensity of reporter pro- 12. Kim L, Mogk A, Schumann W (1996) A xylose-inducible Bacillus
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
teins in the presence of the respective inducer. Furthermore, subtilis integration vector and its application. Gene 181:71–76
13. Kolesinska B, Fraczyk J, Binczarski M, Modelska M, Berlowska
this work is meaningful for augmenting genetic tools encom- J, Dziugan P, Antolak H, Kaminski ZJ, Witonska IA, Kregiel D
passing other organisms beyond C. acetobutylicum ATCC (2019) Butanol synthesis routes for biofuel production: trends and
824. perspectives. Materials (Basel). https://doi.org/10.3390/ma120
30350
Acknowledgements This work was supported by US Army Contract 14. Kremers GJ, Gilbert SG, Cranfill PJ, Davidson MW, Piston DW
W911NF18C0078. (2011) Fluorescent proteins at a glance. J Cell Sci 124:157–160.
https://doi.org/10.1242/jcs.072744
15. Landete JM, Peiroten A, Rodriguez E, Margolles A, Medina M,
Compliance with ethical standard Arques JL (2014) Anaerobic green fluorescent protein as a marker
of Bifidobacterium strains. Int J Food Microbiol 175:6–13. https
Conflict of interest All the authors declare that he/she has no conflict ://doi.org/10.1016/j.ijfoodmicro.2014.01.008
of interest. 16. Lee KM, Min K, Choi O, Kim KY, Woo HM, Kim Y, Han SO,
Um Y (2015) Electrochemical detoxification of phenolic com-
pounds in lignocellulosic hydrolysate for Clostridium fermenta-
tion. Bioresour Technol 187:228–234. https://doi.org/10.1016/j.
References biortech.2015.03.129
17. Lin YL, Blaschek HP (1983) Butanol production by a butanol-
1. Amador-Noguez D, Brasg IA, Feng XJ, Roquet N, Rabinowitz tolerant strain of Clostridium acetobutylicum in extruded corn
JD (2011) Metabolome remodeling during the acidogenic-solven- broth. Appl Environ Microbiol 45:966–973
togenic transition in Clostridium acetobutylicum. Appl Environ 18. Maniatis T, Fritsch EF, Sambrook J (1982) Molecular cloning:
Microbiol 77:7984–7997. https: //doi.org/10.1128/AEM.05374- 11 a laboratory manual, vol Cold. Cold Spring Harbor Laboratory
2. Banerjee A, Leang C, Ueki T, Nevin KP, Lovley DR (2014) Lac- Press, Spring Harbor
tose-inducible system for metabolic engineering of Clostridium 19. Mermelstein LD, Papoutsakis ET (1993) In vivo methylation in
ljungdahlii. Appl Environ Microbiol 80:2410–2416. https://doi. Escherichia coli by the Bacillus subtilis phage phi 3T I meth-
org/10.1128/AEM.03666-13 yltransferase to protect plasmids from restriction upon transfor-
3. Berezina OV, Brandt A, Yarotsky S, Schwarz WH, Zverlov VV mation of Clostridium acetobutylicum ATCC 824. Appl Environ
(2009) Isolation of a new butanol-producing Clostridium strain: Microbiol 59:1077–1081
high level of hemicellulosic activity and structure of solventogen- 20. Mermelstein LD, Papoutsakis ET, Petersen DJ, Bennett GN (1993)
esis genes of a new Clostridium saccharobutylicum isolate. Syst Metabolic engineering of Clostridium acetobutylicum ATCC 824
Appl Microbiol 32:449–459. https://doi.org/10.1016/j.syapm for increased solvent production by enhancement of acetone for-
.2009.07.005 mation enzyme activities using a synthetic acetone operon. Bio-
4. Birgen C, Durre P, Preisig HA, Wentzel A (2019) Butanol produc- technol Bioeng 42:1053–1060. https://doi.org/10.1002/bit.26042
tion from lignocellulosic biomass: revisiting fermentation per- 0906
formance indicators with exploratory data analysis. Biotechnol 21. Mermelstein LD, Welker NE, Bennett GN, Papoutsakis ET (1992)
Biofuels 12:167. https://doi.org/10.1186/s13068-019-1508-6 Expression of cloned homologous fermentative genes in Clostrid-
5. Chandel ASS, Singh O (2013) Detoxification of lignocellulose ium acetobutylicum ATCC 824. Biotechnology (N Y) 10:190–195
hydrolysates: biochemical and metabolic engineering toward 22. Miller JH (1972) Experiments in Molecular Genetics, vol Cold.
white biotechnology. Bioenergy Res 6:388–401 Cold Spring Harbor Laboratory Press, Spring Harbor
6. Drepper T, Eggert T, Circolone F, Heck A, Krauss U, Guterl 23. Muh U, Pannullo AG, Weiss DS, Ellermeier CD (2019) A xylose-
JK, Wendorff M, Losi A, Gartner W, Jaeger KE (2007) Reporter inducible expression system and a CRISPRi-plasmid for targeted
proteins for in vivo fluorescence without oxygen. Nat Biotechnol knock-down of gene expression in Clostridioides difficile. J Bac-
25:443–445. https://doi.org/10.1038/nbt1293 teriol. https://doi.org/10.1128/JB.00711-18
7. Ezeji T, Blaschek HP (2008) Fermentation of dried distillers’ 24. Nariya H, Miyata S, Kuwahara T, Okabe A (2011) Development
grains and solubles (DDGS) hydrolysates to solvents and value- and characterization of a xylose-inducible gene expression system
added products by solventogenic clostridia. Bioresour Technol for Clostridium perfringens. Appl Environ Microbiol 77:8439–
99:5232–5242. https://doi.org/10.1016/j.biortech.2007.09.032 8441. https://doi.org/10.1128/AEM.05668-11
8. Girbal L, Mortier-Barriere I, Raynaud F, Rouanet C, Croux C, 25. Pogrebnyakov I, Jendresen CB, Nielsen AT (2017) Genetic tool-
Soucaille P (2003) Development of a sensitive gene expression box for controlled expression of functional proteins in Geobacillus
13620 Journal of Industrial Microbiology & Biotechnology (2020) 47:609–620
spp. PLoS ONE 12:e0171313. https://doi.org/10.1371/journ complex, multilayered response of Clostridium acetobutylicum
al.pone.0171313 to butanol and butyrate stress. BMC Syst Biol 7:120. https://doi.
26. Pyne ME, Bruder M, Moo-Young M, Chung DA, Chou CP (2014) org/10.1186/1752-0509-7-120
Technical guide for genetic advancement of underdeveloped and 41. Wu H, Lee J, Karanjikar M, San KY (2014) Efficient free fatty
intractable Clostridium. Biotechnol Adv 32:623–641. https://doi. acid production from woody biomass hydrolysate using metaboli-
org/10.1016/j.biotechadv.2014.04.003 cally engineered Escherichia coli. Bioresour Technol 169:119–
27. Pyne ME, Moo-Young M, Chung DA, Chou CP (2013) Develop- 125. https://doi.org/10.1016/j.biortech.2014.06.092
ment of an electrotransformation protocol for genetic manipula- 42. Wu Y, Yang Y, Ren C, Yang C, Yang S, Gu Y, Jiang W (2015)
tion of Clostridium pasteurianum. Biotechnol Biofuels 6:50. https Molecular modulation of pleiotropic regulator CcpA for glucose
://doi.org/10.1186/1754-6834-6-50 and xylose coutilization by solvent-producing Clostridium ace-
28. Qi F, Thakker C, Zhu F, Pena M, San KY, Bennett GN (2018) tobutylicum. Metab Eng 28:169–179. https://doi.org/10.1016/j.
Improvement of butanol production in Clostridium acetobutyli- ymben.2015.01.006
cum through enhancement of NAD(P)H availability. J Ind Micro- 43. Xin F, Wu YR, He J (2014) Simultaneous fermentation of glucose
biol Biotechnol 45:993–1002. https://doi.org/10.1007/s1029 and xylose to butanol by Clostridium sp. strain BOH3. Appl Envi-
5-018-2068-7 ron Microbiol 80:4771–4778. https: //doi.org/10.1128/AEM.00337
29. Ransom EM, Ellermeier CD, Weiss DS (2015) Use of mCherry -14
Downloaded from https://academic.oup.com/jimb/article/47/8/609/6046100 by guest on 29 December 2020
Red fluorescent protein for studies of protein localization and 44. Yang X, Tu M, Xie R, Adhikari S, Tong Z (2013) A comparison
gene expression in Clostridium difficile. Appl Environ Microbiol of three pH control methods for revealing effects of undissociated
81:1652–1660. https://doi.org/10.1128/AEM.03446-14 butyric acid on specific butanol production rate in batch fermenta-
30. Ransom EM, Weiss DS, Ellermeier CD (2016) Use of mCher- tion of Clostridium acetobutylicum. AMB Expr 3:3. https://doi.
ryOpt fluorescent protein in Clostridium difficile. Methods Mol org/10.1186/2191-0855-3-3
Biol 1476:53–67. https://doi.org/10.1007/978-1-4939-6361-4_5 45. Yu L, Xu M, Tang IC, Yang ST (2015) Metabolic engineering of
31. Scotcher MC, Bennett GN (2005) SpoIIE regulates sporulation Clostridiumtyrobutyricum for n-butanol production through co-
but does not directly affect solventogenesis in Clostridium ace- utilization of glucose and xylose. Biotechnol Bioeng 112:2134–
tobutylicum ATCC 824. J Bacteriol 187:1930–1936. https://doi. 2141. https://doi.org/10.1002/bit.25613
org/10.1128/JB.187.6.1930-1936.2005 46. Yu PL, Smart JB, Ennis BM (1987) Differential Induction of Beta-
32. Servinsky MD, Kiel JT, Dupuy NF, Sund CJ (2010) Transcrip- galactosidase and phospho-beta-galactosidase activities in the
tional analysis of differential carbohydrate utilization by Clostrid- fermentation of whey permeate by Clostridium–Acetobutylicum.
ium acetobutylicum. Microbiology 156:3478–3491. https://doi. Appl Microbiol Biotechnol 26:254–257. https://doi.org/10.1007/
org/10.1099/mic.0.037085-0 Bf00286319
33. Shin H, Cho Y, Choe DH, Jeong Y, Cho S, Kim SC, Cho BK 47. Yu Y, Tangney M, Aass HC, Mitchell WJ (2007) Analysis of the
(2014) Exploring the functional residues in a flavin-binding mechanism and regulation of lactose transport and metabolism in
fluorescent protein using deep mutational scanning. PLoS ONE Clostridium acetobutylicum ATCC 824. Appl Environ Microbiol
9:e97817. https://doi.org/10.1371/journal.pone.0097817 73:1842–1850. https://doi.org/10.1128/AEM.02082-06
34. Teng L, Wang K, Xu J, Xu C (2015) Flavin mononucleotide 48. Zhang J, Hong W, Zong W, Wang P, Wang Y (2018) Markerless
(FMN)-based fluorescent protein (FbFP) as reporter for promoter genome editing in Clostridium beijerinckii using the CRISPR-
screening in Clostridium cellulolyticum. J Microbiol Methods Cpf1 system. J Biotechnol 284:27–30. https://doi.org/10.1016/j.
119:37–43. https://doi.org/10.1016/j.mimet.2015.09.018 jbiotec.2018.07.040
35. Thakker C, Lin K, Martini-Stoica H, Bennett GN (2016) Use 49. Zhang J, Liu YJ, Cui GZ, Cui Q (2015) A novel arabinose-induc-
of transposase and ends of IS608 enables precise and scarless ible genetic operation system developed for Clostridium cellulo-
genome modification for modulating gene expression and meta- lyticum. Biotechnol Biofuels 8:36. https://doi.org/10.1186/s1306
bolic engineering applications in Escherichia coli. Biotechnol J 8-015-0214-2
11:80–90. https://doi.org/10.1002/biot.201500205 50. Zhang L, Leyn SA, Gu Y, Jiang W, Rodionov DA, Yang C (2012)
36. Thakker C, Martinez I, Li W, San KY, Bennett GN (2015) Meta- Ribulokinase and transcriptional regulation of arabinose metabo-
bolic engineering of carbon and redox flow in the production of lism in Clostridium acetobutylicum. J Bacteriol 194:1055–1064.
small organic acids. J Ind Microbiol Biotechnol 42:403–422. https https://doi.org/10.1128/JB.06241-11
://doi.org/10.1007/s10295-014-1560-y 51. Zhang Y, Xu S, Chai C, Yang S, Jiang W, Minton NP, Gu Y
37. Tracy BP, Jones SW, Fast AG, Indurthi DC, Papoutsakis ET (2016) Development of an inducible transposon system for effi-
(2012) Clostridia: the importance of their exceptional substrate cient random mutagenesis in Clostridium acetobutylicum. FEMS
and metabolite diversity for biofuel and biorefinery applications. Microbiol Lett. https://doi.org/10.1093/femsle/fnw065
Curr Opin Biotech 23:364–381. https://doi.org/10.1016/j.copbi 52. Zhu F, Wang C, San KY, Bennett GN (2020) Metabolic engineer-
o.2011.10.008 ing of Escherichia coli to produce succinate from woody hydro-
38. Tummala SB, Welker NE, Papoutsakis ET (1999) Development lysate under anaerobic conditions. J Ind Microbiol Biotechnol.
and characterization of a gene expression reporter system for https://doi.org/10.1007/s10295-020-02259-7
Clostridium acetobutylicum ATCC 824. Appl Environ Microbiol
65:3793–3799 Publisher’s Note Springer Nature remains neutral with regard to
39. Voigt C, Bahl H, Fischer RJ (2014) Identification of PTS(Fru) jurisdictional claims in published maps and institutional affiliations.
as the major fructose uptake system of Clostridium acetobu-
tylicum. Appl Microbiol Biotechnol 98:7161–7172. https://doi.
org/10.1007/s00253-014-5809-1
40. Wang Q, Venkataramanan KP, Huang H, Papoutsakis ET, Wu CH
(2013) Transcription factors and genetic circuits orchestrating the
13You can also read

















































