Glucagon-mediated Ca2 signaling in BHK cells expressing cloned human glucagon receptors
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Glucagon-mediated Ca21 signaling in BHK cells
expressing cloned human glucagon receptors
LARS H. HANSEN,1 JESPER GROMADA,2 PIERRE BOUCHELOUCHE,3 TED WHITMORE,4
LAURA JELINEK,4 WAYNE KINDSVOGEL,4 AND ERICA NISHIMURA1
1Department of Molecular Signaling, Hagedorn Research Institute, DK-2820 Gentofte;
2Islet Cell Physiology, Novo Nordisk A/S, DK-2100 Copenhagen; 3Department of
Clinical Biochemistry, Roskilde County Hospital, DK-4600 Køge, Denmark; and
4ZymoGenetics Inc., Seattle, Washington 98102
Hansen, Lars H., Jesper Gromada, Pierre Bouche- The glucagon receptor belongs to the superfamily of
louche, Ted Whitmore, Laura Jelinek, Wayne Kindsvo- seven transmembrane-spanning receptors that couple
gel, and Erica Nishimura. Glucagon-mediated Ca21 signal- to heterotrimeric guanine nucleotide-binding proteins
ing in BHK cells expressing cloned human glucagon receptors. (G proteins). Furthermore, on the basis of structural
Am. J. Physiol. 274 (Cell Physiol. 43): C1552–C1562, 1998.— homology, glucagon receptors, together with those for
From video imaging of fura 2-loaded baby hamster kidney glucagon-like peptide 1 (GLP-1), gastric inhibitory
(BHK) cells stably expressing the cloned human glucagon polypeptide (GIP), secretin, vasoactive intestinal poly-
receptor, we found the Ca21 response to glucagon to be peptide (VIP), growth hormone-releasing factor, cortico-
specific, dose dependent, synchronous, sensitive to pertussis
tropin-releasing factor, pituitary adenylate cyclase-
toxin, and independent of Ca21 influx. Forskolin did not elicit
activating polypeptide, parathyroid hormone, and
a Ca21 response, but treatment with a protein kinase A
inhibitor, the Rp diastereomer of 8-bromoadenosine-38,58-
calcitonin (5, 18), form a subfamily of closely related
cyclic monophosphothioate, resulted in a reduced glucagon- receptors that is now emerging as a group of G protein-
mediated Ca21 response as well as Ca21 oscillations. The coupled receptors able to activate multiple signaling
specific phospholipase C inhibitor U-73122 abolished the pathways. All these receptors are able to stimulate
Ca21 response to glucagon, and a modest twofold increase in adenylate cyclase, but, in addition, many have been
inositol trisphosphate (IP3 ) production could be observed found to activate alternative intracellular second mes-
after stimulation with glucagon. In BHK cells coexpressing sengers. For example, stimulation of the parathyroid
glucagon and muscarinic (M1 ) acetylcholine receptors, carba- hormone receptor leads to intracellular accumulation
chol blocked the rise in intracellular free Ca21 concentrations of cAMP, inositol trisphosphates (IP3 ), and Ca21 (1),
in response to glucagon, whereas glucagon did not affect the and splice variants of the pituitary adenylate cyclase-
carbachol-induced increase in Ca21. Furthermore, carbachol, activating polypeptide receptor are able to differen-
but not glucagon, could block thapsigargin-activated in- tially couple to adenylate cyclase and phospholipase C
creases in intracellular free Ca21 concentration. These results (23). The cloned receptors for calcitonin (7), GLP-1 (9,
indicate that, in BHK cells, glucagon receptors can activate 28), GIP (29), and glucagon (16) have been shown to
not only adenylate cyclase but also a second independent G stimulate cAMP production and a rise in intracellular
protein-coupled pathway that leads to the stimulation of free Ca21 concentration ([Ca21]i ).
phospholipase C and the release of Ca21 from IP3-sensitive It had long been postulated that the cellular effects of
intracellular Ca21 stores. Finally, we provide evidence to
glucagon are mediated not only by cAMP but also by
suggest that cAMP potentiates the IP3-mediated effects on
[Ca21]i, since it has been reported that, in perfused rat
intracellular Ca21 handling.
liver or cultured hepatocytes, glucagon is able to in-
phospholipase C; adenosine 38,58-cyclic monophosphate; G crease the [Ca21]i by inducing a Ca21 influx as well as
proteins; inositol trisphosphate; intracellular calcium stores; by stimulating the release of intracellular Ca21 stores
baby hamster kidney cells (3, 6, 21). However, the mediator(s) of this rise in [Ca21]i
remains to be determined, particularly since there
appears to be some controversy as to whether this Ca21
response to glucagon is a cAMP-mediated effect or a
GLUCAGON, A PANCREATIC HORMONE that plays an impor-
result of a separate signaling pathway. Experiments
tant role in maintaining glucose homeostasis, is se- demonstrating that the effects of glucagon on Ca21
creted from the a-cells in response to low blood glucose mobilization in rat liver cells can be reproduced by
levels and stimulates hepatic glucose output by increas- cAMP analogs have led some investigators to conclude
ing glycogenolysis and gluconeogenesis while at the that the glucagon-mediated rise in [Ca21]i is mediated
same time inhibiting glycolysis (5). In addition to its by cAMP (8, 24, 25). In contrast, others have reported
metabolic actions in the liver, glucagon is also involved cAMP-independent effects of glucagon on intracellular
in the regulation of adipose, cardiac, renal, gastrointes- Ca21 mobilization (22, 27). On the basis of their studies
tinal, and pancreatic functions, including the potentia- demonstrating that TH-glucagon, a biologically active
tion of glucose-induced insulin secretion (5). These glucagon analog that is unable to activate adenylate
effects are mediated by specific glucagon receptors, cyclase, could stimulate inositol phosphate production
which have been shown to have a wide tissue distribu- in rat hepatocytes, Wakelam et al. (27) had proposed
tion corresponding to the observed multiple functions the possible existence of two distinct hepatic glucagon
of glucagon (12). receptors: one that couples to adenylate cyclase and the
C1552 0363-6143/98 $5.00 Copyright r 1998 the American Physiological SocietyGLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING C1553
other to the breakdown of inositol phospholipids. How- nated with alternating wavelengths (340 and 380 nm) by
ever, we have since demonstrated that the cloned rat using interference filters mounted on a filter wheel and
hepatic glucagon receptor is capable of mediating both observed through a 510-nm emission filter. The images were
a cAMP and a Ca21 response (16). captured in real time with a low-light-level intensified charge-
coupled devise videocamera system. The system was cali-
In the current study we have used baby hamster
brated for free Ca21 concentrations by use of the formula
kidney (BHK) cells expressing the cloned human gluca-
gon receptor (19) to further characterize our original [Ca21]i 5 Kd · b · (R 2 R340/380 min)/(R340/380 max 2 R)
observation that the glucagon receptor is able to stimu-
late an increase in [Ca21]i. By video imaging of fura where Kd is the dissociation constant of the dye (225 nM for
2-loaded cells, we have monitored the Ca21 response to fura 2), b is the proportionality coefficient (4), and R is the
glucagon and have addressed the questions as to the wavelength ratio. Calibrations of the fura 2 fluorescence
signal were performed by dialyzing single cells with 100 µM
source of the glucagon-mediated rise in [Ca21]i and the fura 2 pentapotassium salt and Ca21-EGTA buffers with free
possible molecular mechanisms involved. In view of Ca21 concentrations ranging from 0 to 39.8 µM. Thus
glucagon’s central role in regulating glucose metabo- R340/380 max was determined under saturating Ca21 concentra-
lism and our previous findings that a mutated glucagon tions, whereas R340/380 min was measured for the same field
receptor may in some manner contribute to the develop- under Ca21-free conditions. The ratio images (R340/R380 ) were
ment of non-insulin-dependent diabetes (10, 11), the obtained by dividing the 340-nm images by the 380-nm image
characterization of the molecular signaling pathways on a pixel-by-pixel basis, whereas the numeric R340/R380
activated by glucagon is not only important for under- values were calculated after the pixel gray values obtained for
standing the normal physiological mechanisms in- each wavelength were averaged. Ratio values were then
volved in glucagon action in target tissues such as the converted to Ca21 concentrations by use of the calibration
curve, and the background (field without cells) was sub-
liver, fat, or pancreas, but it may also lead to the tracted.
identification of defects in glucagon signal transduction For the experiments shown in Table 1 and Fig. 1, the
that may occur in diabetes. recordings were made using the Tardis imaging software
(Applied Imaging, Tyne & Wear, UK). For the toxin sensitiv-
MATERIALS AND METHODS ity studies, the cells were pretreated with 10 µg/ml pertussis
or cholera toxin for 24 h at 37°C in the culture medium. The
Cell culture and transfections. BHK cells were cultured in recordings in Figs. 3, 4, and 6–8 were made using a fluores-
DMEM supplemented with 10% heat-inactivated FCS, 100 cence imaging system (IonOptix, Milton, MA).
U/ml penicillin, 10 µg/ml streptomycin, and 2 mM L-glutamine IP3 determinations. BHK cells transfected with the human
at 37°C in 5% CO2-95% air. The transfection of BHK cells with glucagon receptor or the human M1 acetylcholine receptor
the human glucagon receptor cDNA pLJ6 (19) was carried out were seeded (5 3 104 cells) into 60-mm tissue culture dishes
using the Liptofectamine reagent, as previously outlined in and allowed to grow for 3–4 days to ,90% confluency. For an
detail (11). The clones analyzed in the present study have estimate of cell number, the cells from two dishes were
been characterized previously for the level of glucagon recep- counted for each cell line. The cells were then incubated for
tor expression (,3 3 106 receptors/cell) and for their ability to $1 h in DMEM culture medium containing 10 mM myo-
bind 125I-labeled glucagon (dissociation constant ,10 nM) as inositol and for an additional 30 min in the presence of 10 mM
well as stimulate cAMP production in response to glucagon myo-inositol and 10 mM LiCl2 in DMEM. Cells were subse-
(EC50 ,2 pM) (11). For the BHK cells coexpressing the quently stimulated for 30 s with various concentrations of
glucagon and muscarinic (M1 ) receptors, the glucagon receptor- glucagon or carbachol prepared in DMEM containing 1%
expressing BHK cells were transfected with the M1 choliner- BSA. The reactions were terminated by the addition of 2 ml of
gic receptor plasmid pM1-R by use of the calcium-phosphate ice-cold 15% TCA. The cells were then scraped off and
method (20). transferred onto ice for 20 min, and then they were centri-
[Ca21]i measurements. BHK cells (,3 3 104 ), seeded onto
thin, circular coverslips (22 mm) that had been precoated
with poly-D-lysine (10 µg/ml), were cultured for 3 days. Before Table 1. Toxin sensitivity and specificity
Ca21 measurements, the cells were loaded with 7 µM fura of glucagon-induced increase in [Ca21 ]i
2-AM in the presence of the nonionic detergent Pluronic in BHK cells stably transfected
F-127 (25%) at 37°C for 30 min, and then an equal volume of with human glucagon receptor cDNA
HEPES-buffered RPMI culture medium was added. The cells
were then incubated for a further 30 min to allow complete No. of Cells Increase in
deesterification of fura 2. Subsequently, the cells were washed Treatment Responding [Ca21 ]i , nM
three times in NaCl-HEPES buffer (in mM: 145 NaCl, 5 KCl,
50 nM glucagon 30/30 488 6 44
1 Na2HPO4, 0.5 Mg2SO4, 20 HEPES, 5 glucose, 1 CaCl2, pH
24-h Pretreatment with pertussis toxin
7.5). Finally, the coverslips were mounted in a recording followed by 50 nM glucagon 8/40 72.5 6 13
chamber and placed on an inverted microscope (model 135 24-h Pretreatment with cholera toxin fol-
TU, Zeis axioscope, Oberkochen, Germany). The recording lowed by 50 nM glucagon 20/20 279 6 7
chamber was continuously perfused with NaCl-HEPES buffer 1 µM GLP-1 0/20 ND
supplemented with 1% BSA at 1.5–2 ml/min. All test com- 1 µM VIP 0/20 ND
pounds were applied through the perfusate to give the
Increases in intracellular free Ca21 concentration ([Ca21 ]i ) were
concentrations indicated. For experiments that required Ca21- determined as difference between peak and baseline responses and
free medium, the CaCl2 in the NaCl-HEPES buffer was are presented as means 6 SD from data obtained from at least 2
replaced with 1 mM EGTA. experiments with use of different passages of cell lines. BHK, baby
The [Ca21]i measurements were performed essentially as hamster kidney; GLP-1, glucagon-like peptide 1; VIP, vasoactive
outlined previously (4, 9, 26). Briefly, the cells were illumi- intestinal polypeptide; ND, not detectable.C1554 GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING
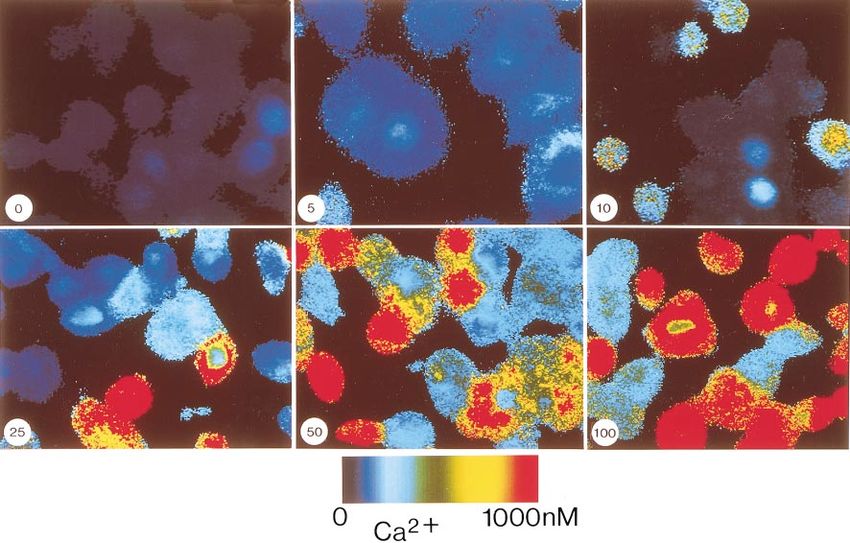
Fig. 1. Glucagon-induced mobilization of Ca21 in baby hamster kidney (BHK) cells expressing cloned human
glucagon receptors. Each panel represents pseudocolor ratio images of fura 2-labeled cells obtained at time of peak
response to concentration of glucagon (in nM) indicated at bottom left. Colors represent range of free Ca21
concentrations according to scale. Original magnification, 3300.
fuged for 15 min at 250 g at 4°C. The supernatants were then diameter) glass coverslips were purchased from Struers Kebo
transferred to polypropylene tubes containing 10 µl of 5 mM Lab (Albertslund, Denmark). Cell culture medium and compo-
EDTA. To the remaining pellets 0.5 ml of 15% TCA was added, nents, FCS, and Lipofectamine reagent were from GIBCO
and the pellets were incubated for an additional 20 min on ice. BRL (Paisley, Scotland). Poly-D-lysine, forskolin, pertussis
The resulting supernatant was combined with the first and toxin, cholera toxin, thimerosal, EGTA, LaCl3, carbachol, and
extracted four times with one volume of diethyl ether (H2O myo-inositol were purchased from Sigma Chemical (St. Louis,
saturated). The aqueous phase was freeze-dried, resuspended MO). Glucagon was obtained from Novo Nordisk (Bagsvaerd,
in 2 ml of H2O, and adjusted to pH 7.5 with 1 M NaHCO3. Denmark); GLP-1-(7—36) amide and VIP were from Penin-
Amersham’s D-myo-inositol 1,4,5-trisphosphate (IP3 ) 3H sula Laboratories (Belmont, CA). The cAMP antagonist, an
assay system was used to measure IP3 in the BHK cell Rp diastereomer of 8-bromoadenosine-38,58-cyclic monophos-
extracts according to the manufacturer’s instructions. For phothioate (Rp-8-BrcAMPS), and the cAMP agonistic Sp
each cell extract, duplicate determinations of a 1:10 dilution diastereomer of adenosine 38,58-cyclic monophosphothioate
were assayed. The samples were counted for 4 min in a beta (Sp-8-BrcAMPS) were obtained from BIOLOG Life Science
scintillation counter, and the amount of IP3 in each sample Institute (Bremen, Germany). Ryanodine was from Alomone
was determined by interpolation from the standard curve. Labs (Jerusalem, Israel). The phospholipase C inhibitor
cAMP measurements. Cells were seeded out in six-well U-73122 and its inactive analog U-73343 were obtained from
plates (3 3 105 cells/well) and cultured overnight. Before Calbiochem (Bad Soden, Germany). Fura 2-AM and fura 2
stimulation the cells were washed once in Hanks’ balanced pentapotassium salt were from Molecular Probes (Eugene,
salt solution and again in RPMI 1640 medium, and finally in OR). The IP3 3H assay system and cAMP 125I scintillation
1 ml of RPMI medium supplemented with 0.5% fetal bovine proximity assay system (dual range) from Amersham (Little
serum and 0.45 mM IBMX was added to each well. The cells Chalfont, England) were used for IP3 and cAMP measure-
were stimulated with glucagon and carbachol, and after 20 ments.
min of incubation at 37°C, 1 ml of 65% ethanol was added to
each well. The cells were scraped off and transferred to RESULTS
Eppendorf tubes, which were then centrifuged at 300 g for 15
min. The resulting supernatants were dried down overnight Specific, dose-dependent rise in [Ca21]i mediated by
in a Speed-Vac and stored at 220°C until they were assayed. glucagon. In BHK cells stably expressing the cloned
The cAMP concentrations were determined using the cAMP human glucagon receptor, we found that stimulation by
125I scintillation proximity assay (Amersham). The dried cell
glucagon results in a transient, synchronized, and
extracts were resuspended in 1 ml of assay buffer and diluted
1:150. For the assay the acetylation protocol described by the dose-dependent rise in [Ca21]i (Figs. 1 and 2). The
manufacturer was followed, and the cAMP determinations response is rapid and recovers to prestimulus levels
were normalized to cell number. within 2–3 min. As can be noted in Fig. 1, it is not only
Reagents. All tissue culture flasks and dishes were from the amplitude of the response to glucagon that in-
Nunc (Roskilde, Denmark). Thin (0.2-mm), circular (22-mm- creases in a dose-dependent manner but also theGLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING C1555
further 1 mM EGTA. Under these conditions, glucagon
was still able to elicit a robust rise in [Ca21]i of the same
magnitude as obtained in the presence of extracellular
Ca21 (Fig. 3, A and B), suggesting that this transient
rise in [Ca21]i is a result of the release of intracellular
Ca21 stores. This is also confirmed by our finding that
LaCl3, a general Ca21 channel blocker, did not affect the
Ca21 response to glucagon (Fig. 3C).
Fig. 2. Concentration-dependent rise in intracellular free Ca21 con-
centration ([Ca21]i ) in response to glucagon in BHK cells expressing
human glucagon receptors. Increase in [Ca21]i in response to gluca-
gon was determined as difference between peak and baseline Ca21
values and is represented as means 6 SD from data obtained from 24
cells from at least 3 different experiments.
number of responding cells rises with increasing concen-
trations of glucagon, such that at high levels ($25 nM)
all cells respond. Although the amplitude of the Ca21
response to glucagon varies from cell to cell, it appears
that all responding cells do so in a synchronous man-
ner. There is a delay in the response that becomes
progressively less when higher concentrations of gluca-
gon are used; there is a 75 6 3 s lapse from the time 5
nM glucagon is introduced to the time the rise in [Ca21]i
peaks, whereas the delay is only 53 6 1, 38 6 1, and
31 6 2 s for 10, 25, and 50 nM glucagon, respectively
(n 5 6). The observed effects of glucagon on the increase
in [Ca21]i appear to be specific, mediated by the binding
and activation of specific glucagon receptors, since the
related hormones, GLP-1-(7—36) amide and VIP, were
not able to elicit a rise in [Ca21]i in these cells (Table 1).
Finally, in nontransfected BHK cells, no Ca21 response
to glucagon could be observed.
Toxin sensitivity of the glucagon-mediated rise in
[Ca21]i. The glucagon-induced rise in [Ca21]i was found
to be mediated by the activation of a pertussis toxin-
sensitive G protein, since pretreatment of the human
glucagon receptor expressing BHK cells with pertussis
toxin resulted in the near ablation of the Ca21 response
to glucagon (Table 1). Under identical conditions, per-
tussis toxin had no effect on the carbachol-stimulated
Ca21 response in BHK cells expressing the M1 choliner-
gic receptors. In contrast, a glucagon-mediated rise in
[Ca21]i was still observed in cells pretreated with
cholera toxin, although the amplitude of the response
was decreased in comparison to that observed in non- Fig. 3. Characterization of glucagon-induced rise in [Ca21]i in BHK
treated cells (Table 1). cells expressing human glucagon receptors. A: stimulation with 200
Source of the glucagon-mediated rise in [Ca21]i. To nM glucagon over time period covered by horizontal line (control).
determine the source of the observed increase in [Ca21]i, B: stimulation with 200 nM glucagon in Ca21-free medium supple-
mented with 1 mM EGTA. C: stimulation with 200 nM glucagon in
we examined the Ca21 response to glucagon in the presence of 0.6 mM LaCl3. Each trace represents response in a single
absence of extracellular free Ca21, which was achieved cell and is representative of at least 30 single-cell recordings from 3
by using Ca21-free medium to which was added a different experiments.C1556 GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING
Mechanisms involved in the glucagon-mediated rise
in [Ca21]i. Binding of glucagon to its receptors results in
activation of heterotrimeric G proteins (Gs subtype),
stimulating the production of cAMP, which ultimately
functions as the second messenger mediating many of
glucagon’s cellular effects. However, because it has also
been reported that stimulation of hepatocytes with
glucagon leads to activation of phospholipase C and
production of IP3, we have examined the putative role
of this second messenger in glucagon receptor signal-
ing. As evident from Fig. 4A, incubation of the cells with
the phospholipase C inhibitor U-73122 completely abol-
ished the ability of glucagon to mobilize intracellular
Ca21. The inactive isoform of the inhibitor U-73343 was
without effect (Fig. 4B), indicating that activation of
phospholipase C is indeed involved in the glucagon-
induced Ca21 response. Furthermore, incubation of the
cells with 50 µM ryanodine had no effect on the Ca21
response to subsequent stimulation with glucagon (Fig.
4C). These findings are consistent with our observation
that glucagon can stimulate a twofold increase in IP3
production (Fig. 5A). This response, however, appears
to be relatively modest compared with the 100-fold
increase obtained when carbachol is used to stimulate
BHK cells expressing M1 cholinergic receptors, which
are known to activate phospholipase C, resulting in the
formation of IP3 and release of IP3-sensitive intracellu-
lar Ca21 stores. Figure 5B also shows that stimulation
with carbachol does not lead to any concomitant in-
crease in cAMP production, indicating that signaling by
the M1 receptors, when transfected into BHK cells, is
essentially normal. The effectiveness of the phospholi-
pase C inhibitor U-73122 is illustrated in Fig. 5C,
where treatment of the cells with this agent (15 µM)
decreases the carbachol-mediated IP3 response by 67.5 6
7.8% (n 5 4) compared with untreated cells. In addi-
tion, the specificity of U-73122 can be seen in Fig. 5D,
where this inhibitor was found to have no effect on
adenylate cyclase activity as determined by the accumu-
lation of cAMP in response to glucagon.
Further characterization of the Ca21 response to
glucagon was carried out in BHK cells coexpressing
glucagon and M1 receptors. As expected, repeated stimu-
lation of these cells with the same agonist, glucagon
(Fig. 6A) or carbachol (Fig. 6B), results in desensitiza-
tion of the Ca21 response. Interestingly, we found that Fig. 4. Mechanisms involved in glucagon-stimulated increase in
although prior stimulation with carbachol could pre- [Ca21]i. Fura 2-loaded BHK cells expressing human glucagon recep-
tors were stimulated with 200 nM glucagon after incubation for 10
vent the Ca21 response to a subsequent glucagon min with 15 µM U-73122 (A), 15 µM U-73343 (B), or 50 µM ryanodine
treatment (Fig. 6D), glucagon pretreatment was not (C) for time periods indicated by horizontal lines. Each trace repre-
able to prevent the rise in [Ca21]i elicited by a subse- sents change in [Ca21]i within a single cell and is representative of at
quent carbachol stimulation (Fig. 6C). These results least 20 single-cell recordings from 3 experiments.
indicate that glucagon releases Ca21 from the IP3-
sensitive intracellular stores but that, unlike carba- the thapsigargin-releasable intracellular Ca21 stores,
chol, glucagon is not capable of emptying these stores since treatment with thapsigargin immediately after
completely. This is further substantiated by our finding glucagon stimulation can still elicit a rise in [Ca21]i
that, after depletion of thapsigargin-sensitive intracel- (Fig. 6E).
lular Ca21 stores, glucagon is unable to induce a rise in To investigate whether cAMP may be involved in the
[Ca21]i (data not shown). In addition, unlike carbachol, glucagon-induced rise in [Ca21]i, we examined the
which can deplete thapsigargin-sensitive intracellular effects of forskolin, which stimulates the accumulation
Ca21 stores (Fig. 6F), glucagon only partially empties of intracellular cAMP by directly activating adenylateGLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING C1557
Fig. 5. Accumulation of D-myo-inositol
1,4,5-trisphosphate (IP3 ) and cAMP in
BHK cells expressing human glucagon
receptors (BHK; Glu) or muscarinic
(M1 ) receptors (BHK; M1). A: glucagon-
and carbachol-stimulated IP3 produc-
tion. B: glucagon- and carbachol-stimu-
lated cAMP production. Effect of 10
min of pretreatment with 15 µM U-
73122 on carbachol-stimulated accumu-
lation of IP3 (C) or glucagon-stimulated
cAMP production (D) is shown. [IP3]i
and [cAMP]i, intracellular free IP3 and
cAMP concentration, respectively. Val-
ues are means 6 SD of 4 determina-
tions.
cyclase. As can be seen in Fig. 7A, forskolin was not in results, the activator of protein kinase A (PKA), Sp-8-
itself able to elicit a Ca21 response in these cells, BrcAMPS, was also unable to stimulate an increase in
whereas the subsequent addition of glucagon was still [Ca21]i (Fig. 7C). In addition, we examined the effects of
able to stimulate a rise in [Ca21]i. Consistent with these Rp-8-BrcAMPS, a cAMP analog that specifically inhib-C1558 GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING Fig. 6. Desensitization of glucagon- and carbachol-mediated rise in [Ca21]i in BHK cells coexpressing human gluca- gon receptors and M1 acetylcholine re- ceptors. Cells were stimulated for time intervals indicated by horizontal lines. Each trace represents response in a single cell and is representative of at least 30 single-cell recordings from 3 different experiments. its PKA (8), the effectiveness of which has been demon- whereas on its own, thimerosal does not affect the strated in BHK cells by its ability to inhibit the [Ca21]i (data not shown). A Ca21 response of normal translocation of the catalytic subunit of PKA, as deter- magnitude is observed after stimulation with glucagon, mined by video-imaging techniques (O. Thastrup, per- which is then followed by a number of Ca21 oscillations sonal communication). Here we found that Rp-8- (Fig. 8A). To further test this hypothesis that cAMP BrcAMPS inhibits the Ca21 response to glucagon, as potentiates the Ca21 response to IP3, we examined the can be seen by the diminished [Ca21]i transients (Fig. effect of cAMP on the carbachol-stimulated rise in 7B). Furthermore, in some of the Rp-8-BrcAMPS- [Ca21]i in BHK cells expressing M1 receptors. We found treated cells, glucagon-evoked Ca21 oscillations could that subthreshold concentrations of carbachol could be observed; the amplitude of the repetitive spikes induce a Ca21 response in the presence of forskolin (Fig. appeared to be somewhat lower than that of the initial 8, B and C). Finally, in BHK cells transfected with a response (Fig. 7B). These data suggest that the rise in mutant (Gly40Ser) glucagon receptor that had previ- [Ca21]i after glucagon stimulation is most likely not ously been identified in some diabetic patients (10), we mediated by cAMP itself but that it may in some way observed a reduction in the Ca21 response to glucagon modulate or potentiate the Ca21 response. Further (Fig. 7D), and, interestingly in some of these cells, support of this suggestion is the finding that, together glucagon-induced Ca21 oscillations occurred that were with forskolin, the IP3 receptor-sensitizing agent thi- similar to those seen in cells treated with Rp-8- merosal is able to elicit a Ca21 response (Fig. 8A), BrcAMPS or thimerosal (Figs. 7B and 8A, respectively).
GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING C1559
Fig. 7. Role of protein kinase A in
glucagon-mediated Ca21 response. BHK
cells expressing wild-type human gluca-
gon receptor were stimulated with 200
nM glucagon after a previous stimula-
tion with 10 µM forskolin (A), 100 µM
Rp diastereomer of 8-bromoadenosine
38,58-cyclic monophosphothioate (Rp-8-
BrcAMPS, B), or 100 µM Sp diaste-
reomer of 8-bromoadenosine 38,58-cyclic
monophosphothioate (Sp-8-BrcAMPS, C)
for time period indicated by horizontal
bar. D: BHK cells expressing mutant
(Gly40Ser) glucagon receptors were
stimulated with 200 nM glucagon. Each
trace represents response in a single
cell and is representative of at least 20
single-cell recordings from 3 different
experiments.
DISCUSSION where the first transient rise in [Ca21]i has been
attributed to the mobilization of intracellular thapsigar-
We previously demonstrated that glucagon stimu-
gin-sensitive Ca21 stores and the second sustained rise
lates a concentration-dependent rise in intracellular
cAMP production in BHK cells expressing human in Ca21 has been found to be a result of Ca21 influx
glucagon receptors (11, 19). Here we provide evidence through a cation channel (29). Despite these differences
to indicate that activation of the cloned human gluca- among the Ca21 signaling mediated by the various
gon receptor expressed in BHK cells leads to a rise in related receptor types, the characteristics of the Ca21
[Ca21]i that does not appear to be due to cAMP- response that we observe after activation of the cloned
mediated activation of Ca21 influx. This suggests that human glucagon receptor nonetheless are supported by
the glucagon receptor, like many others of the secretin previous studies in which endogenous hepatic glucagon
receptor family, can stimulate two independent signal- receptors were found to have Ca21-mobilizing effects
ing pathways: one mediated by cAMP and the other by that could not be reproduced by cAMP (22, 27). We do
Ca21. recognize, however, that by overexpressing the gluca-
From studies aimed at characterizing the rise in gon receptor in BHK cells we run the risk of ‘‘forcing’’
[Ca21]i stimulated by glucagon, we found it to be the coupling to effectors that may not necessarily occur
independent of extracellular Ca21, since it was not in cells expressing the endogenous receptors. Neverthe-
affected by blocking of membrane Ca21 channels or by less, previous studies using cultured primary hepato-
removal of extracellular Ca21. These findings indicate cytes have demonstrated that glucagon is able to
that the source of the glucagon-stimulated rise in activate cAMP- and Ca21-mediated signaling path-
[Ca21]i is intracellular Ca21 stores. Thus the general ways, which supports our findings with use of the BHK
features of the glucagon-mediated rise in [Ca21]i appear cells transfected with the cloned glucagon receptor. In
to differ from those described in response to activation addition, signaling by the cloned M1 receptors in BHK
of GLP-1 and GIP receptors, the two most closely cells overexpressed to the same extent as the glucagon
related to the glucagon receptor. In contrast to the Ca21 receptors seems to have the same characteristics as
response to glucagon, the Ca21-mobilizing effects of signaling by endogenous M1 receptors, that is, activa-
GLP-1 in pancreatic b-cells have been demonstrated to tion of phospholipase C and release of intracellular
be dependent on extracellular Ca21 and mimicked by Ca21 without a concomitant increase in cAMP produc-
stimulation with cAMP. This response has been pro- tion (Fig. 5, A and B).
posed to involve influx of extracellular Ca21 through To address the question as to the mechanism of
voltage-dependent Ca21 channels as well as a release of glucagon-induced Ca21 signaling, we have shown that
Ca21 from intracellular stores (15). From further stud- phospholipase C and IP3 are indeed involved, resulting
ies on b-cells, it has been proposed that the mobiliza- in the release of intracellular Ca21 stores. This is
tion of Ca21 from intracellular stores in response to demonstrated by our observation that the phospholi-
GLP-1 reflects Ca21-induced Ca21 release, since it was pase C inhibitor U-73122 completely abolishes the rise
blocked by ryanodine (9). On the other hand, the rise in in [Ca21]i stimulated by glucagon (Fig. 4A). In addition,
[Ca21]i elicited by GIP through the cloned receptor we observed a twofold increase in IP3 in response to
expressed in COS-7 cells has been shown to be biphasic, glucagon stimulation, although this was small com-C1560 GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING
tially empties the Ca21 stores mobilized by IP3 after
activation of M1 cholinergic receptors. The fact that
stimulation with carbachol could desensitize the Ca21
response to a subsequent stimulation with glucagon
whereas the opposite is not the case suggests that
glucagon and carbachol mobilize Ca21 from the same
IP3-sensitive intracellular stores but to varying de-
grees. This is further exemplified by the finding that
thapsigargin is still able to release Ca21 stores after
stimulation by glucagon but not by carbachol (Fig. 6, E
and F). Finally, Hofer et al. (14) found that the intraor-
ganellar sequestration of Ca21 in BHK cells is not
affected by ryanodine, which supports our finding that
this agent has no effect on glucagon’s Ca21 response
(Fig. 4C). Although it is thus apparent that phospholi-
pase C-catalyzed IP3 production is involved in the Ca21
response to M1 and glucagon receptor activation, the
upstream signaling mechanisms leading to this phos-
pholipase C activation are clearly different, since we
find that pertussis toxin blocks the glucagon-mediated
rise in [Ca21]i (Table 1). Because M1 cholinergic recep-
tors couple to phospholipase C by a pertussis toxin-
resistant G protein (Gq ) (2), the glucagon-induced Ca21
signal must involve a G protein distinct from that used
by the M1 receptors. It has been shown that the
bg-subunit of the pertussis toxin-sensitive Gi is able to
activate the b2-isoform of phospholipase C (17), and
since it has been recently shown that the luteinizing
hormone receptor is able to couple to Gas and Gai to
activate adenylate cyclase and phospholipase C, with
the bg-subunits released from either G protein contrib-
uting to the stimulation of phospholipase C b-isoforms
(13), it is possible that this mechanism may also be
involved in the glucagon-mediated Ca21 response.
Our results also indicate that Gs- and Gi-activated
pathways are involved in the Ca21 response to gluca-
gon, since Rp-8-BrcAMPS, which acts as an inhibitor of
PKA, was found to reduce the glucagon-stimulated
increase in [Ca21]i (Fig. 7B), whereas neither forskolin
nor the cAMP agonist Sp-8-BrcAMPS was able to elicit
a Ca21 response on its own. Furthermore, the IP3-
sensitizing agent thimerosal, although unable to elicit
a Ca21 response itself, leads to an increase in IP3 if the
cells have previously been exposed to forskolin (Fig.
8A). Further substantiation of the possible intereaction
between cAMP and IP3 is given by our finding that a
subthreshold concentration of carbachol (1 nM) is also
Fig. 8. Modulation of glucagon- and carbachol-evoked Ca21 re-
able to evoke a Ca21 response after prior treatment
sponses. A: BHK cells expressing human glucagon receptors were with forskolin (Fig. 8, B and C). This suggests that
stimulated with 200 nM glucagon after pretreatment with 10 µM although cAMP is not responsible for the rise in [Ca21]i
forskolin and 100 µM thimerosal. BHK cells expressing human M1 stimulated by glucagon, cAMP can potentiate the Ca21
receptors were stimulated with 1 nM and 100 nM carbachol (B) or 1 response. It is interesting to note that the induction of
nM carbachol after previous stimulation with 10 µM forskolin (C).
Each trace represents change in [Ca21]i within a single cell and is Ca21 oscillations that occurred after thimerosal (Fig.
representative of at least 20 single-cell recordings from 3 experi- 8A) or Rp-8-BrcAMPS (Fig. 7B) treatment was similar
ments. to our observation in BHK cells expressing the Gly40Ser
mutant receptor (Fig. 7D), which we have reported to
pared with the response observed with carbachol (Fig. be present in a subset of diabetic patients (10). We
5A). This difference in the magnitude of IP3 formation previously demonstrated that this mutation results in
provides an explanation for our heterologous desensiti- a decreased sensitivity of the receptor to glucagon in
zation experiments, from which it is apparent that, terms of binding and cAMP production (11). However,
even at a maximal concentration, glucagon only par- the mechanisms underlying the induction of this oscil-GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING C1561
latory response to glucagon under certain conditions couple different receptors to PI hydrolysis in the same cell. Cell
and the significance of these oscillations remain to be 56: 487–493, 1989.
3. Benedetti, A., P. Graf, R. Fulceri, A. Romani, and H. Sies.
determined. Ca21 mobilization by vasopressin and glucagon in perfused
Thus, taking into consideration all our findings, we livers. Biochem. Pharmacol. 38: 1799–1805, 1989.
suggest the following model for glucagon-mediated 4. Bouchelouche, P. N. Dynamic, real time imaging of ion activi-
Ca21 signaling in which two distinct signaling path- ties in single living cells using fluorescence video microscopy and
image analysis. Scand. J. Clin. Lab. Invest. 53: 27–39, 1993.
ways are involved. On the basis of sensitivity to pertus- 5. Burcelin, R., E. B. Katz, and M. J. Charron. Molecular and
sis and cholera toxins, it appears that the glucagon cellular aspects of the glucagon receptor: role in diabetes and
receptor can couple to multiple G proteins: Gs and metabolism. Diabetes Metab. 22: 373–396, 1996.
possibly Gi. In addition, the glucagon-stimulated cAMP 6. Bygrave, F. L., A. Gamberucci, R. Fulceri, and A. Bene-
detti. Evidence that stimulation of plasma-membrane Ca21
production appears to play a permissive role in the rise inflow is an early action of glucagon and dibutyryl cyclic AMP in
in [Ca21]i, such that cAMP, although itself unable to rat hepatocytes. Biochem. J. 292: 19–22, 1993.
elicit a response, in concert with the small incremental 7. Chabre, O., B. Conklin, H. Lin, H. Lodish, E. Wilson, H.
release of IP3 is able to mediate the observed rise in Ives, L. Catanzariti, B. Hemmings, and H. Bourne. A
[Ca21]i in response to glucagon. Previously, we proposed recombinant calcitonin receptor independently stimulates aden-
osine 38,58-cyclic monophosphate and Ca21/inositol phosphate
that a similar mechanism underlies the GLP-1-stimu- signaling pathways. Mol. Endocrinol. 6: 551–556, 1992.
lated release of Ca21 from intracellular stores in pancre- 8. Connelly, P. A., L. H. Botelho, R. B. Sisk, and J. C. Garrison.
atic b-cells, such that a cAMP-induced phosphorylation A study of the mechanism of glucagon-induced protein phosphor-
of the IP3 receptor (mediated by PKA) may enhance the ylation in isolated rat hepatocytes using (Sp)-cAMPS and
(Rp)-cAMPS, the stimulatory and inhibitory diastereomers of
Ca21 mobilization in response to the low levels of IP3 adenosine cyclic 38,58-phosphorothioate. J. Biol. Chem. 262:
arising from activation of phospholipase C by localized 4324–4332, 1987.
Ca21 influx (9). Therefore, although we did not see an 9. Gromada, J., S. Dissing, K. Bokvist, E. Renström, J. Frøk-
increase in Ca21 in response to cAMP stimulation alone jaer-Jensen, B. S. Wulf, and P. Rorsman. Glucagon-like
peptide I increases cytoplasmic calcium in insulin-secreting
(via forskolin), the cAMP produced in response to bTC3 cells by enhancement of intracellular calcium mobilization.
activation of glucagon receptors in these cells may Diabetes 44: 767–774, 1995.
potentiate the effects of the rather small increase in IP3 10. Hager, J., L. Hansen, C. Vaisse, N. Vionnet, A. Philippi, W.
we observed after glucagon stimulation. This model is Poller, G. Velho, C. Carcassi, L. Contu, C. Julier, F. Cam-
bien, P. Passa, M. Lathrop, W. Kindsvogel, F. Demenais, E.
supported by our observation that thimerosal or a Nishimura, and P. Froguel. A missense mutation in the
subthreshold concentration of carbachol is able to in- glucagon receptor gene is associated with non-insulin-dependent
duce a Ca21 response in the presence of forskolin. In diabetes mellitus. Nat. Genet. 9: 299–304, 1995.
summary, we have analyzed the general characteristics 11. Hansen, L. H., N. Abrahamsen, J. Hager, L. Jelinek, W.
of the glucagon-mediated rise in [Ca21]i and have Kindsvogel, P. Froguel, and E. Nishimura. The Gly40Ser
mutation in the human glucagon receptor gene associated with
provided evidence to suggest that, contrary to earlier NIDDM results in a receptor with reduced sensitivity to gluca-
proposals of two glucagon receptor subtypes in the liver gon. Diabetes 45: 725–730, 1996.
(27), the cloned human glucagon receptor is able to 12. Hansen, L. H., N. Abrahamsen, and E. Nishimura. Glucagon
couple to multiple G proteins, thereby activating two receptor mRNA distribution in rat tissues. Peptides 16: 1163–
1166, 1995.
distinct signaling pathways. It remains a challenge, 13. Herrlich, A., B. Kÿhn, R. Grosse, A. Schmid, G. Schultz, and
however, to elucidate the physiological role of these T. Gudermann. Involvement of Gs and Gi proteins in dual
cAMP-Ca21 signaling pathways stimulated by gluca- coupling of the luteinizing hormone receptor to adenylyl cyclase
gon in its target tissues. and phospholipase C. J. Biol. Chem. 271: 16764–16772, 1996.
14. Hofer, A. M., W.-R. Schlue, S. Curci, and T. E. Machen.
The technical assistance of Ane Bøjet Baunsgaard is gratefully Spatial distribution and quantitation of free luminal [Ca] within
acknowledged. We thank Dr. Nils Billestrup for insightful comments the InsP3-sensitive internal store of individual BHK-21 cells: ion
regarding the manuscript. dependence of InsP3-induced Ca release and reloading. FASEB J.
This work was supported in part by the Danish Research Academy. 9: 788–798, 1995.
The Hagedorn Research Institute is an independent research unit of 15. Holz, G. G., IV, C. A. Leech, and J. F. Habener. Activation of a
Novo Nordisk. cAMP-regulated Ca21-signaling pathway in pancreatic b-cells by
Address for reprint requests: E. Nishimura, Dept. of Molecular the insulinotropic hormone glucagon-like peptide-1. J. Biol.
Signaling, Hagedorn Research Institute, Niels Steensens Vej 6, Chem. 270: 17749–17757, 1995.
DK-2820 Gentofte, Denmark. 16. Jelinek, L. J., S. Lok, G. B. Rosenberg, R. A. Smith, F. J.
Grant, S. Biggs, P. A. Bensch, J. L. Kuijper, P. O. Sheppard,
Received 19 March 1997; accepted in final form 9 February 1998. C. A. Sprecher, P. J. O’Hara, D. Foster, K. M. Walker, L. H. J.
Chen, P. A. McKernan, and W. Kindsvogel. Expression
cloning and signaling properties of the rat glucagon receptor.
REFERENCES
Science 259: 1614–1616, 1993.
1. Abou-Samra, A.-B., H. Jÿppner, T. Force, M. W. Freeman, 17. Katz, A., D. Wu, and M. I. Simon. Subunits bg of heterotrimeric
X.-F. Kong, E. Schipani, P. Urena, J. Richards, J. V. Bonven- G protein activate b2 isoform of phospholipase C. Nature 360:
tre, J. T. J. Potts, H. M. Kronenberg, and G. V. Segre. 686–689, 1992.
Expression cloning of a common receptor for parathyroid hor- 18. Laburthe, M., A. Couvineau, P. Gaudin, J.-J. Maoret, C.
mone and parathyroid hormone-related peptide from rat osteo- Rouyer-Fessard, and P. Nicole. Receptors for VIP, PACAP,
blast-like cells: a single receptor stimulates intracellular accumu- secretin, GRF, glucagon, GLP-1, and other members of their new
lation of both cAMP and inositol triphosphates and increases family of G protein-linked receptors: structure-function relation-
intracellular free calcium. Proc. Natl. Acad. Sci. USA 89: 2732– ship with special reference to the human VIP-1 receptor. In: VIP,
2736, 1992. PACAP, and Related Peptides: Second International Symposium,
2. Ashkenazi, A., E. Peralta, J. Winslow, J. Ramachandran, edited by A. Arimura and S. I. Said. New York: NY Acad. Sci.,
and D. Capon. Functionally distinct G proteins selectively 1996, p. 94–109. (Ann. NY Acad. Sci., vol. 805)C1562 GLUCAGON RECEPTOR-MEDIATED CA21 SIGNALING
19. Lok, S., J. L. Kuijper, L. J. Jelinek, J. M. Kramer, T. E. mic free Ca21 concentration in isolated rat hepatocytes. Biochem.
Whitmore, C. A. Sprecher, S. Mathewes, F. J. Grant, S. H. J. 238: 737–743, 1986.
Biggs, G. B. Rosenberg, P. O. Sheppard, P. J. O’Hara, D. C. 25. Staddon, J. M., and R. G. Hansford. Evidence indicating that
Foster, and W. Kindsvogel. The human glucagon receptor the glucagon-induced increase in cytoplasmic free Ca21 concentra-
encoding gene: structure, cDNA sequence and chromosomal tion in hepatocytes is mediated by an increase in cyclic AMP
localization. Gene 140: 203–209, 1994. concentration. Eur. J. Biochem. 179: 47–52, 1989.
20. Maniatis, T., E. Fritsch, and J. Sambrook. Molecular Clon- 26. Stroop, S., D. Thompson, R. Kuestner, and E. Moore. A
ing: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring recombinant human calcitonin receptor functions as an extracel-
Harbor Laboratory, 1989. lular calcium sensor. J. Biol. Chem. 268: 19927–19930, 1993.
21. Mine, T., I. Kojima, and E. Ogata. Sources of calcium mobi- 27. Wakelam, M. J. O., G. J. Murphy, V. J. Hruby, and M. D.
lized by glucagon in isolated rat hepatocytes. Acta Endocrinol. Houslay. Activation of two signal-transduction systems in hepa-
119: 301–306, 1988. tocytes by glucagon. Nature 323: 68–71, 1986.
22. Mine, T., I. Kojima, and E. Ogata. Evidence of cyclic AMP- 28. Wheeler, M., M. Lu, J. Dillon, X.-H. Leng, C. Chen, and A.
independent action of glucagon on calcium mobilization in rat Boyd III. Functional expression of the rat glucagon-like pep-
hepatocytes. Biochim. Biophys. Acta 970: 166–171, 1988. tide-1 receptor, evidence for coupling to both adenylyl cyclase and
23. Spengler, D., C. Waeber, C. Pantaloni, F. Holsboer, J. phospholipase-C. Endocrinology 133: 57–61, 1993.
Bockaert, P. H. Seeburg, and L. Journot. Differential signal 29. Wheeler, M. B., R. W. Gelling, C. H. S. McIntosh, J. Geor-
transduction of five splice variants of the PACAP receptor. giou, J. C. Brown, and R. A. Pederson. Functional expression
Nature 365: 170–175, 1993. of the rat pancreatic islet glucose-dependent insulinotropic poly-
24. Staddon, J. M., and R. G. Hansford. 4b-Phorbol 12-myristate peptide receptor: ligand binding and intracellular signaling
13-acetate attenuates the glucagon-induced increase in cytoplas- properties. Endocrinology 136: 4629–4639, 1995.You can also read

















































