LINC00665 functions as a competitive endogenous RNA to regulate AGTR1 expression by sponging miR 34a 5p in glioma
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
1202 ONCOLOGY REPORTS 45: 1202-1212, 2021
LINC00665 functions as a competitive endogenous RNA to
regulate AGTR1 expression by sponging miR‑34a‑5p in glioma
YONGYUE DAI1, YUCHENG ZHANG2, MAOLIN HAO1 and RENWU ZHU2
1
Department of Pathophysiology, Wenzhou Medical University, Wenzhou, Zhejiang 325035; 2Department of General Surgery,
Wenzhou Hospital Integrated Traditional Chinese and Western Medicine, Wenzhou, Zhejiang 325000, P.R. China
Received January 20, 2020; Accepted August 21, 2020
DOI: 10.3892/or.2021.7949
Abstract. Glioma is the most aggressive tumor of the central substantial mortality (4). Most glioma patients succumb to the
nervous system. Long non‑coding RNAs (lncRNAs) may be disease within 2 years after first diagnosis (5). The capacities
involved in modulating tumor generation. The present study to migrate, rapidly diffuse and invade paracancerous tissues,
analyzed an lncRNA microarray of glioma and selected long heterogeneity, and incessant proliferation of glioma cells
intergenic non‑protein coding RNA 665 (LINC00665) as the contribute to the overall survival of approximately 15 months
research object. The mode of expression and biological function for most patients with glioma at the late stage (6‑8). Hence,
of LINC00665 in glioma were assessed using lncRNA micro- improved understanding of novel mechanisms governing
array and RT‑qPCR analyses. Gain‑of‑function assays and/or glioma cell growth and metastasis is a key to the exploitation
loss‑of‑function assays were implemented to explore the role of early diagnostic regimens and personalized treatment.
of LINC00665 in the progression of glioma. Dual‑luciferase Long non‑coding RNAs (lncRNAs) are ncRNAs at least
reporter and RNA immunoprecipitation assays explored the 200 nucleotides in length (9,10). They have been implicated
downstream molecular mechanism of LINC00665. The func- in diverse epigenetic regulatory processes, including histone
tion of the molecular pathway in progression of glioma was modification, chromatin remodeling, RNA alternative splicing,
analyzed using rescue assays. High expression of LINC00665 and transcriptional regulation (11‑14). Due to their specificity
was marked in glioma tissues and cells, which correlated with and easy detection, lncRNAs can be used as biomarkers and
an unsatisfactory prognosis. Upregulation of LINC00665 treatment targets (15‑17). For example, Tamang et al confirmed
significantly promoted the proliferation and invasion of that SNHG12 is a potential therapeutic target and biomarker
glioma cells. LINC00665 acted as a competing endogenous for human cancer (18). Chen et al reported that lncRNAs
RNA by sponging miR‑34a‑5p to upregulate angiotensin II can be biomarkers and treatment targets in non‑small cell
receptor type 1 (AGTR1). LINC00665 promoted the progres- lung cancer (19). The long intergenic non‑protein coding
sion of glioma by acting as a competitive endogenous RNA RNA 665 (LINC00665) lncRNA promotes impacts in diverse
to competitively bind to miR‑34a‑5p and mediate AGTR1 tumors, including gastric cancer (20,21), non‑small cell lung
expression. cancer (22), lung adenocarcinoma (23) and hepatocellular
carcinoma (24). However, the involvement of LINC00665 in
Introduction the development of glioma is unclear.
In the present study, the high expression of LINC00665
Gliomas are the most widely encountered solid tumors of was reported in glioma tissues and cell lines. LINC00665
the central nervous system (CNS) (1,2). As reported in 2018, overexpression (OE) enhanced the proliferative, invasion, and
~100,000 people worldwide are diagnosed as having diffuse migratory potentials of glioma cells. The findings verified
gliomas every year (3). Although it comprises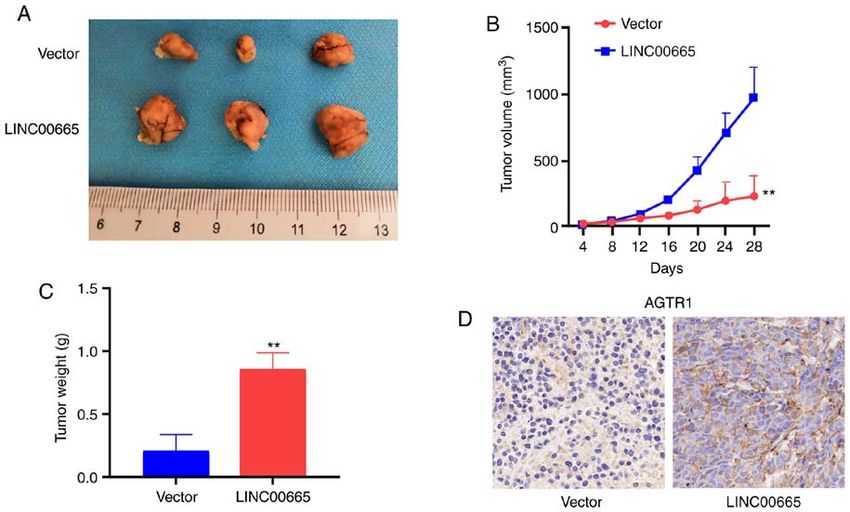
DAI et al: LINC00665 PROMOTES GLIOMA 1203
excision at Wenzhou Hospital Integrated Traditional Chinese Table I. Association of LINC00665 expression with clinico-
and Western Medicine from January 2017 to June 2019. The pathological features of glioma.
patients had not received chemotherapy or radiotherapy before
tissue excision. Prior to RNA extraction, all isolated specimens Expression of
were rapidly cryopreserved at ‑80˚C. Data concerning the LINC00665
‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑‑
association of LINC00665 expression with clinicopathological
Characteristics No. High Low P‑value
features of glioma are provided in Table I.
All cases 48 24 24
Cell culture and transfection. Glioma cell lines U87 MG
(glioblastoma of unknown origin, ATCC® HTB‑14; ATCC), Age (years) 0.5639
LN229 (ATCC ® CRL‑2611), A172 (ATCC ® CRL‑1620), ≤48 25 14 11
U373 MG (ATCC ® HTB‑17), U251 (U251 MG; cat. >48 23 10 13
no. YS448C; YaJi Biological), human normal astrocytes Sex 0.5612
NHA (cat. no. YS2144C; YaJi Biological) and 293T cells (cat. Male 27 12 15
no. YS005C; YaJi Biological) were cultured and preserved in Female 21 12 9
DMEM (GIBCO‑BRL; Thermo Fisher Scientific, Inc.) supple-
Clinical stage 0.0189
mented with 100 U/ml penicillin, 10% fetal bovine serum, and
100 mg/ml streptomycin (Beyotime Institute of Biotechnology) I‑II 21 6 15
in a humidified atmosphere containing 5% CO2 at 37˚C. STR III‑Ⅳ 27 18 9
profiling analysis was performed for the authentication of cell
Total data from 48 tumor tissues of glioma patients were analyzed.
lines.
For the expression of LINC00665 which was assayed by RT‑qPCR,
As per the guidance of the manufacturer (Shanghai the median expression level was used as the cutoff. Data were
GenePharma Co., Ltd.), LINC0 0 665 overexpression analyzed by chi‑squared test and Fisher's exact test. The P‑value in
(OE) plasmid/small interfering (si)RNA and microRNA bold indicates a statistically significant difference.
(miR)‑34a‑5p mimics/inhibitor were used for transfec-
tion assays with Lipofectamine 2000 Reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). Cells grown to approximately
50‑60% confluence in culture dishes were used for transfection. 2 h, followed by three rinses with phosphate‑buffered saline
Transfection was performed in serum‑free medium for one day. (PBS; 0.5 g/ml). Subsequently, 4',6‑diamidino‑2‑phenylindole
(DAPI; Invitrogen; Thermo Fisher Scientific, Inc.) was used to
RNA extraction and reverse transcription‑quantitative PCR stain nuclei of the washed cells for 10 min at room tempera-
(RT‑qPCR). Total RNA was extracted from the tissues and ture in the dark. The DAPI‑stained cells were washed more
cultured cells using TRIzol® (Invitrogen; Thermo Fisher than twice with PBS. Stained cells were analyzed using the
Scientific, Inc.) following the manufacturer's guidelines. FACSCalibur DxP flow cytometer (BD Biosciences).
Approximately 1 µg of total RNA was reversely transcribed to
cDNA using a reverse transcriptase cDNA synthesis kit (Toyobo Cell migration and invasion assays. Cell migration was
Co., Ltd.). qPCR was performed using the SYBR Green PCR examined using a wound healing assay. Cells (5x105) were
kit (Roche Diagnostics) by initial denaturation at 94˚C for seeded in a six‑well plate and cultured to confluence. When
5 min, followed by 40 cycles including denaturation at 94˚C the cells grew to nearly 100% confluency, a 200‑µl pipette
for 30 sec, annealing at 55˚C for 30 sec and extension at 72˚C tip (QIAGEN,) was used to scratch the confluent monolayer
for 90 sec. Comparative quantification was assessed using the of cells. Suspended cells and cell debris were removed by
2‑ΔΔCq method with glyceraldehyde 3‑phosphae dehydrogenase washing three times with PBS. After adding fresh serum‑free
(GAPDH) or U6 used as the endogenous control (25). U6 was medium, the plate was incubated for 24 h with 5% CO2 at
used for normalization of the miRNA whereas GAPDH was 37˚C for 1 h. The wound was photographed regularly using
used for the normalization of other genes, such as AGTR1. The a computer‑assisted microscope (magnification, x100; Nikon
PCR primers used are summarized in Table II. Corporation).
Cell invasion was assessed in a Matrigel assay using
Cell proliferation assays. Approximately, 1.0x103 transfected a 24‑well invasion chamber system from BD Biosciences
U87 MG and U251 cells were cultured in 96‑well plates. Cell equipped with polycarbonic membranes (diameter 6.5 mm;
Counting Kit‑8 (CCK‑8; 10 µl) reagent (Beyotime Institute of pore size 8 µm). Subsequent to incubation at 37˚C for 24 h,
Biotechnology) was added and incubated at 37˚C for 1 h. The a fluorescence microscope (magnification, x200) was used
absorbance at 450 nm was recorded using an Infinite M200 to quantify cells co‑cultured with exosomes and invading
multimode microplate reader (Tecan Group, Ltd.). through the membranes in four fields that were randomly
After approximately 48 h of transfection, the selected. Each assay was repeated at least three times with
5‑ethynyl‑2´‑deoxyuridine (EdU) assay kit provided by triplicate samples each time.
Guangzhou Ribo Co., Ltd., was used to examine the
proliferation of U87 MG and U251 cells. Specifically, cells were Subcellular distribution. The Cytoplasmic and Nuclear RNA
grown in culture medium containing EdU (cat. no. A10044; Purification Kit (Norgen Biotek Corp.) was used to examine
Invitrogen; Thermo Fisher Scientific, Inc.) solution (1,000:1). RNA degradation in the cytoplasm or nucleus. U87 MG and
At the proliferative stage, the cells were labeled with EdU for U251 cells were lysed on ice for 5 min and then centrifuged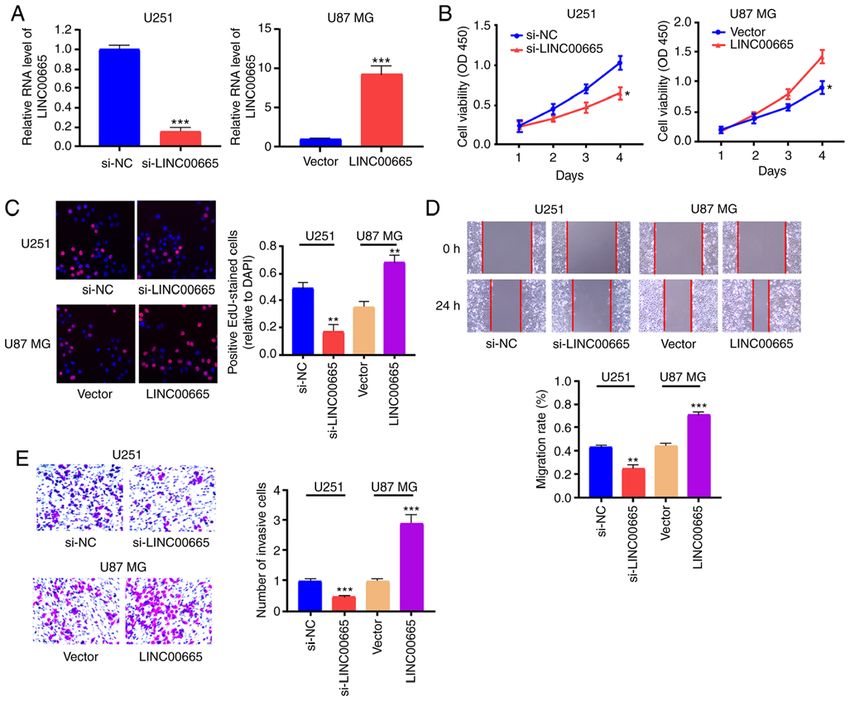
1204 ONCOLOGY REPORTS 45: 1202-1212, 2021
Table II. Sequences of primers for RT‑qPCR and miRNA‑related sequences.
Name Sequence
LINC00665 F: 5'‑GGTGCAAAGTGGGAAGTGTG‑3'
R: 5'‑CGGTGGACGGATGAGAAACG‑3'
miR‑34a‑5p F: 5'‑ACACTCCAGCTGGGTGTTGGTCGATTCTGT‑3'
R: 5'‑CTCAACTGGTGTCGTGGAGTCGGC AATTCAGTTGAGGTGACGGT‑3'
AGTR1 F: 5'‑ATTTAGCACTGGCTGACTTATGC‑3'
R: 5'‑CAGCGGTATTCCATAGCTGTG‑3'
U6 F: 5'‑GGTCGGGCAGGAAAGAGGGC‑3'
R: 5'‑TGGTATCGTGGAAGGACTC‑3'
GAPDH F: 5'‑AGTAGAGGCAGGGATGATG‑3'
R: 5'‑AGGGGCCATCCACAGTCTTC‑3'
si‑LINC00665 Sense, 5'‑AAUAGCCCAAGACUGAGGACUCACA‑3'
Antisense, 5'‑UGUGAGUCCUCAGUCUUGGGCUAUU‑3'
miR‑34a‑5p mimics Sense, 5'‑UGGCAGUGUCUUAGCUGGUUGU‑3'
Antisense, 5'‑ACAACCAGCUAAGACACUGCCA‑3'
miR‑34a‑5p inhibitor Sense, 5'‑ACAACCAGCUAAGACACUGCCA‑3'
F, forward; R, reverse; AGTR1, angiotensin II receptor type 1.
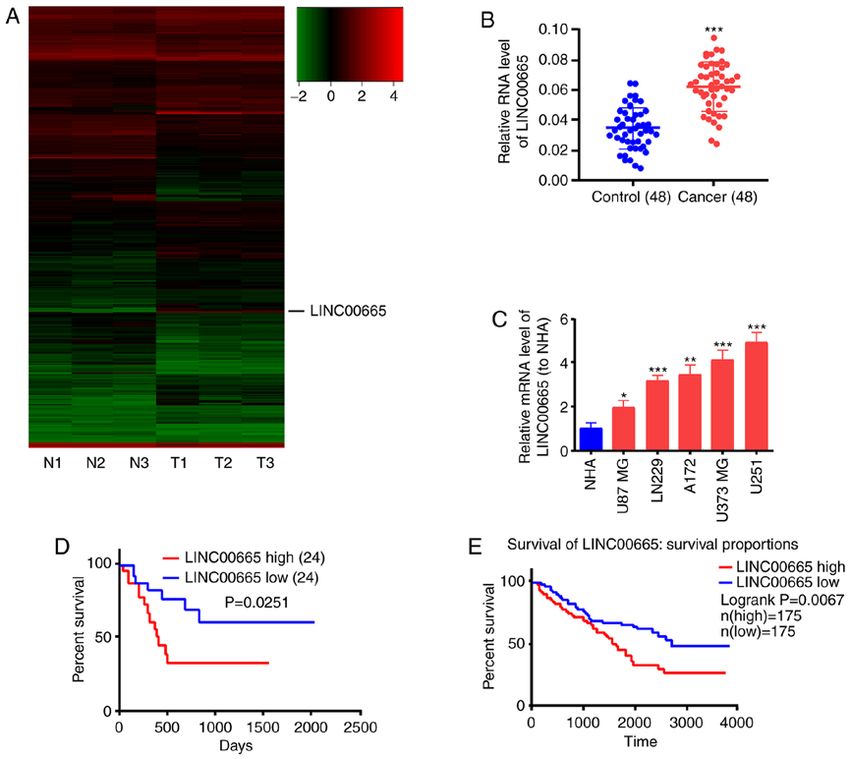
Figure 1. LINC00665 expression in glioma tissues. (A) Heatmap of differentially expressed lncRNAs between glioma tissues and paracarcinoma tissues.
LINC00665 expression was increased in glioma tissues. (B) LINC00665 expression was increased in glioma tissue samples. (C) LINC00665 expression was
significantly higher in glioma cell lines relative to the NHA cell line. Kaplan‑Meier survival curve revealing the overall survival of glioma patients stratified
by LINC00665 expression based on (D) our dataset and the (E) TCGA dataset. *P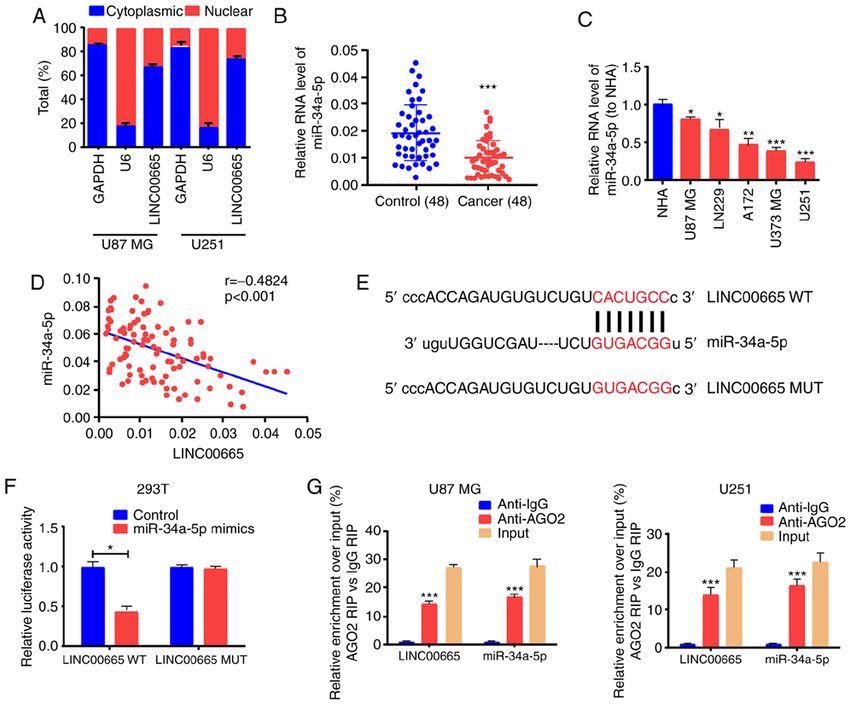
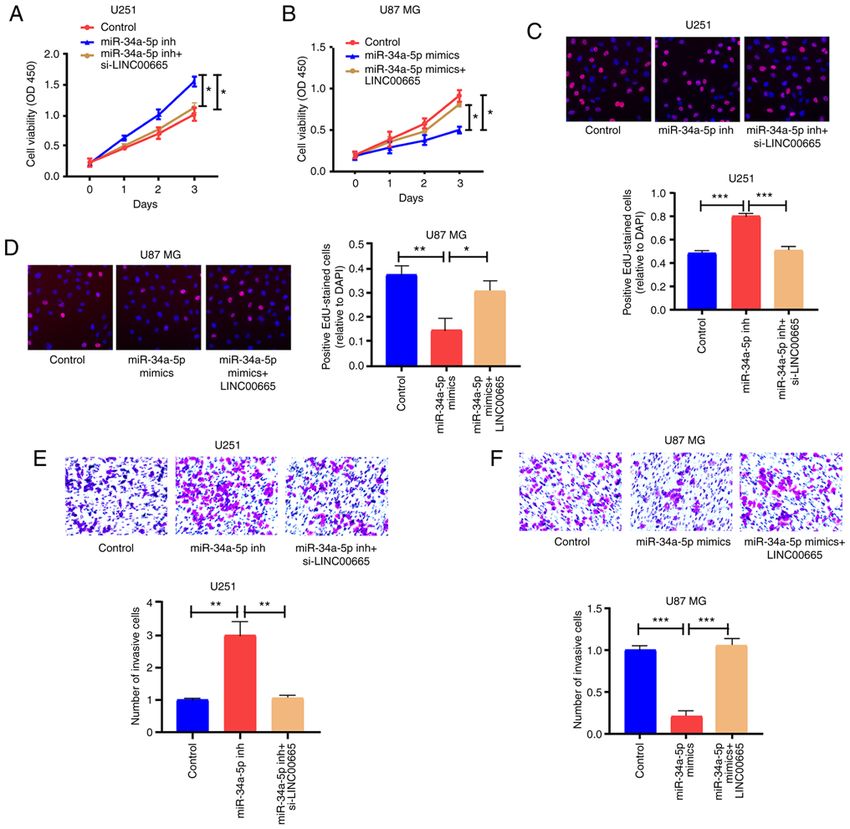
DAI et al: LINC00665 PROMOTES GLIOMA 1205 Figure 2. Regulatory effect of LINC00665 on proliferation, migration, and invasion of glioma cells. (A) LINC00665 expression levels in U251 and U87 MG cells were detected by RT‑qPCR after transfection with LINC00665 siRNA or LINC00665 OE vector. (B and C) CCK‑8 and EdU assays revealed proliferation of U251 cells treated with LINC00665 siRNA and U87 MG cells undergoing LINC00665 OE vector treatment. (D) Wound healing assay demonstrating the migration of glioma cells. (E) Transwell assay demonstrating the invasion of glioma cells. Images were captured by light microscopy (x200). *P
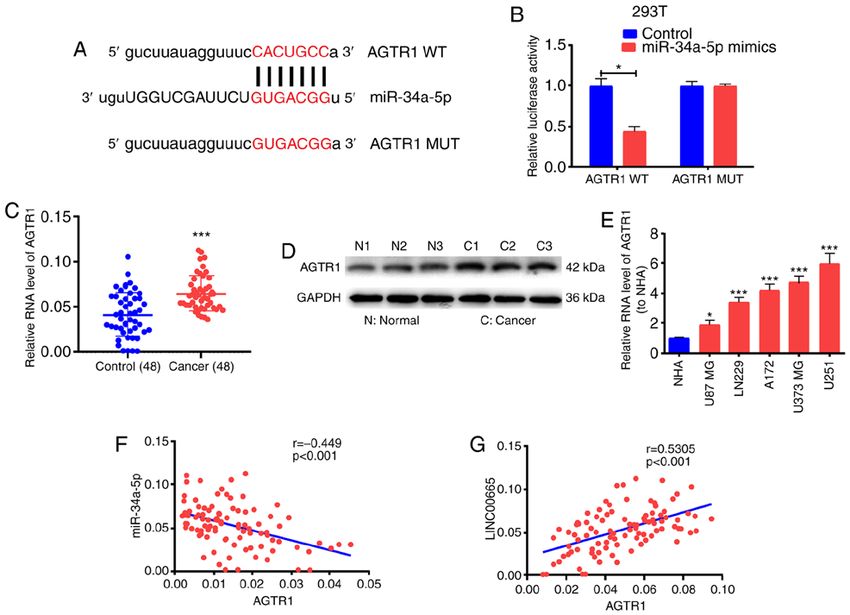
1206 ONCOLOGY REPORTS 45: 1202-1212, 2021 Figure 3. LINC00665 directly and mutually acts with miR‑34a‑5p. (A) Cytoplasmic and nuclear levels of LINC00665 in U251 and U87 MG cells assessed by RT‑qPCR. (B) Decreased miR‑34a‑5p expression in glioma and paracancerous tissue samples. (C) miR‑34a‑5p expression in glioma cell lines and NHA cells detected by RT‑qPCR. (D) Pearson correlation analysis of the correlation of LINC00665 with miR‑34a‑5p in glioma. (E) Identification of the miR‑34a‑5p binding site sequence with LINC00665. (F) Dual‑Luciferase reporter gene assay for 293T cells. (G) RIP assays for quantifying LINC00665 and miR‑34a‑5p in U251 and U87 MG cells. *P
DAI et al: LINC00665 PROMOTES GLIOMA 1207 Figure 4. AGTR1 is a direct target of miR‑34a‑5p. (A) The possible binding sites in the AGTR1 sequence. (B) Direct target sites verified by the Dual‑Luciferase reporter gene assay. (C and D) AGTR1 expression in glioma tissues detected by RT‑qPCR and western blotting. (E) AGTR1 expression in glioma cells and NHA cells. (F) Pearson correlation analysis of the correlation of AGTR1 with miR‑34a‑5p in glioma. (G) Pearson correlation analysis of the correlation of LINC00665 with AGTR1 in glioma. *P1 or log2FC
1208 ONCOLOGY REPORTS 45: 1202-1212, 2021 Figure 5. LINC00665/miR‑34a‑5p axis is critical for AGTR1 expression. (A and B) miR‑34a‑5p expression levels in U251 and U87 MG cells were detected by RT‑qPCR after transfecting with miR‑34a‑5p inhibitor/mimics. (C and D) miR‑34a‑5p inhibitor with or without LINC00665 siRNA were transfected into U251 cells and miR‑34a‑5p mimics with or without LINC00665 OE vector were transfected into U87 MG cells. AGTR1 expression levels in U251 and U87 MG cells were detected by RT‑qPCR and western blot analysis. (E and F) Relative mRNA and protein levels of AGTR1 following transfection with LINC00665‑WT/MUT OE plasmid. **P
DAI et al: LINC00665 PROMOTES GLIOMA 1209 Figure 6. LINC00665 regulates cell function through miR‑34a‑5p. (A‑D) CCK‑8 and EdU assays were performed to examine U251 and U87 MG cell prolifera- tion. (E and F) The invasion ability of U251 and U87 MG cells after different treatments. *P
1210 ONCOLOGY REPORTS 45: 1202-1212, 2021
Figure 7. LINC00665 in U87 MG cells promotes tumor growth in vivo. (A) Typical images of xenografts in nude mice. (B) Tumor volume and (C) weight.
(D) Typical images of AGTR1 in immunohistochemical staining analysis. AGTR1, angiotensin II receptor type 1
competitive endogenous RNA (ceRNA) in glioma. Analysis (Fig. 4E). Correlation analysis revealed an inverse relationship
using the starBase bioinformatics prediction database between miR‑34a‑5p and AGTR1 expression (Fig. 4F) as well
demonstrated that sequences in miR‑34a‑5p were mark- as a positive correlation between AGTR1 and LINC00665
edly similar to the LINC00665 3'untranslated region (UTR) expression (Fig. 4G).
(Fig. 3E). RT‑qPCR also demonstrated that the expression To determine the modulation of LINC00665 on AGTR1
of miR‑34a‑5p was associated with a decreasing trend in expression by targeting miR‑34a‑5p, the expression level
glioma tissues and cells (Fig. 3B and C). Correlation analysis of AGTR1 in glioma cells was examined after altering
revealed that miR‑34a‑5p and LINC00665 expression were LINC00665 or miR‑34a‑5p expression. The transfection
inversely associated (Fig. 3D). Next, pGL3‑LINC00665‑WT effectiveness of miR‑34a‑5p mimics/inhibitors was assessed
and pGL3‑LINC00665‑MUT were constructed on the basis (Fig. 5A). Then, AGTR1 expression was increased by treating
of binding sequences (Fig. 3E). A significant decrease in the U251 cells with miR‑34a‑5p inhibitors. The increased expres-
luciferase activity of 293T cells was evident during treatment sion was abrogated by treatment with LINC00665 siRNA
with LINC00665‑WT and miR‑34a‑5p mimics, however, no (Fig. 5B and C). Furthermore, AGTR1 expression in U87
change was apparent after treatment with LINC00665‑MUT MG cells treated with miR‑34a‑5p mimics was impeded, and
and miR‑34a‑5p mimics (Fig. 3F). The RIP assay revealed that was reversed by LINC00665 OE treatment (Fig. 5B and D).
LINC00665 was enriched in anti‑AGO2 antibody. Similar Subsequently, U251 cells were transfected with LINC00665
results were revealed for miR‑34a‑5p (Fig. 3G). The findings OE plasmid/MUT OE plasmid, and AGTR1 expression was
indicated that miR‑34a‑5p probably binds to LINC00665 examined. RT‑qPCR and western blotting revealed that
in vitro. LINC00665 WT OE increased the expression of AGTR1 in
glioma cells, while LINC00665 MUT had no influence on
LINC00665 regulates the target gene AGTR1 of miR‑34a‑5p. AGTR1 expression (Fig. 5E and F). The findings indicated
To ascertain the possible function of miR‑34a‑5p in glioma that LINC00665 directly binds to miR‑34a‑5p to positively
growth, the starBase bioinformatics prediction system was modulate AGTR1 expression.
used to screen miR‑34a‑5p target genes. AGTR1 was identified
for subsequent assessment. Subsequent to the establishment LINC00665/miR‑34a‑5p axis regulates the behaviors of
of pGL3‑AGTR1‑WT and pGL3‑AGTR1‑MUT (Fig. 4A), glioma cells. CCK‑8 and EdU assay results revealed that
293T cells were co‑treated with miR‑34a‑5p mimics/control. miR‑34a‑5p inhibition significantly contributed to the ability
Luciferase activity was blocked in the WT reporter group, of U251 cells to proliferate, in contrast to controls. LINC00665
but not in the MUT reporter group (Fig. 4B). These findings siRNA partially abrogated this ability (Fig. 6A and C).
implied that AGTR1 probably is the target gene for miR‑34a‑5p. Additionally, overexpressed miR‑34a‑5p restricted the prolifera-
The levels of AGTR1 mRNA and protein were significantly tion of U87 MG cells, but LINC00665 OE partially reversed this
increased in glioma tissues (Fig. 4C and D). AGTR1 expres- potential (Fig. 6B and D). Moreover, miR‑34a‑5p‑mediated down-
sion was higher in glioma cell lines than in the NHA cell line regulation induced invasion of U251 cells, which was partiallyDAI et al: LINC00665 PROMOTES GLIOMA 1211
reversed by LINC00665 siRNA (Fig. 6E). Overexpressed to LINC00665 in strengthening the malignant phenotype of
miR‑34a‑5p blocked the invasion capability of U87 MG cells, glioma cells.
which was partially reversed by LINC00665 OE (Fig. 6F). Nevertheless, the present study has a number of limitations.
Firstly, a larger tissue sample size of glioma is required to further
LINC00665 in U87 MG cells stimulates tumor growth. Nude explore the clinical value of LINC00665. Secondly, in situ
mice were subcutaneously injected with stably expressed U87 hybridization fluorescence would be valuable to verify the rela-
MG cells transfected with vector or LINC00665 OE to assess tionship between LINC00665 and miR‑34a‑5p in future studies.
the function of LINC00665 in glioma in vivo. Upregulation of In addition, whether there are other target genes or miRNAs
LINC00665 increased the tumor volume (Fig. 7A and B) and which can interact with LINC00665 requires further exploration.
weight (Fig. 7C). Immunohistochemical results demonstrated In conclusion, LINC00665 was increased in human
that mice treated using LINC00665 OE treatment had a higher glioma cell lines and tissues, and its decrement in glioma cells
AGTR1 level (Fig. 7D). impeded proliferation, invasion, and migration of glioma cells.
LINC00665 is a ceRNA that modulated AGTR1 expression
Discussion by sponging miR‑34a‑5p, thus modulating glioma growth. The
present findings could aid in the discovery of new targets for
An increasing number of lncRNAs have been implicated the diagnosis and treatment of glioma.
as biomarkers for glioma growth. For example, lncRNA
PAXIP1‑AS1 enhanced cell invasion and blood vessel formation Acknowledgements
of glioma utilizing transcription factor ETS1 to increase KIF14
expression (30). lncRNA GAS5 inversely regulated miR‑18a‑5p Not applicable.
to modulate glioma cells to proliferate, migrate, and invade (31).
Thus, lncRNAs are likely markedly influential in the onset Funding
and growth of glioma. Continued examinations of the possible
molecular mechanisms and biological functions of lncRNAs in No funding was received.
glioma will identify novel molecular targets for disease treatment.
Presently, increased LINC00665 expression was demon- Availability of data and materials
strated in glioma tissues and cells. In addition, decreased
LINC00665 expression significantly decreased glioma cell The datasets used and/or analyzed during the present study
proliferation, migration, and invasion in vitro, indicating that are available from the corresponding author on reasonable request.
LINC00665 acts as an oncogene to modulate the growth of
glioma cells. A tumor xenograft model was used to confirm the Authors' contributions
role of LINC00665 in glioma. In vivo assays revealed that over-
expressing of LINC00665 in U87 MG cells promoted tumor RZ designed the experiments. YD and YZ performed the
growth. The findings highlight the importance of determining experiments. YD and MH wrote the manuscript. All authors
the role of LINC00665 in enhancing the growth of glioma cells analyzed the results and revised the manuscript. All authors
to better understand the onset, growth, and migration of glioma. have read and approved the final version of the manuscript.
The cross‑regulation between lncRNAs and miRNAs has
been demonstrated. lncRNAs may serve as ceRNAs to modu- Ethics approval and consent to participate
late the expression and functions of miRNAs, and thus have
been termed are ‘miRNA sponges’ (32,33). To understand the The study was approved by the Ethics Committee of Wenzhou
potential oncogenic mechanisms of LINC00665 in glioma Hospital Integrated Traditional Chinese and Western
cells, the starBase bioinformatics database was utilized to iden- Medicine. All participants provided written informed consent.
tify miR‑34a‑5p as a target of LINC00665. Gao et al revealed The mouse experiments were approved by the Animal Care
that miR‑34a‑5p suppressed colorectal cancer metastasis and and Use Committee of Wenzhou Medical University.
predicted recurrence in patients with stage II/III colorectal
cancer (34). Previous studies revealed that miR‑34a‑5p can Patient consent for publication
suppress tumorigenesis and progression of glioma (35‑37). The
present results demonstrated that miR‑34a‑5p was decreased Not applicable.
in glioma tissues and cells. Transfection of miR‑34a‑5p
mimics inhibited glioma cell proliferation and invasion, which Competing interests
could be reversed by LINC00665 OE. It can be concluded
that both LINC00665 and miR‑34a‑5p may be involved in the The authors declare that they have no competing interests.
development and progression of glioma.
The RAS component AGTR1 has the potential to References
stimulate cell growth, migration, or invasion and to promote
angiogenesis, inflammation and immunity (38). The present 1. Deng MY, Sill M, Sturm D, Stichel D, Witt H, Ecker J,
findings affirmed that LINC00665 elevation could increase Wittmann A, Schittenhelm J, Ebinger M, Schuhmann MU, et al:
Diffuse glioneuronal tumour with oligodendroglioma‑like
AGTR1 expression, giving rise to significant proliferation, features and nuclear clusters (DGONC)‑a molecularly‑defined
invasion, and migration of glioma cells. We intend in future glioneuronal CNS tumour class displaying recurrent mono-
studies to investigate other mechanisms that may be related somy 14. Neuropathol Appl Neurobiol 46: 422‑430, 2019.1212 ONCOLOGY REPORTS 45: 1202-1212, 2021
2. Xi J, Sun Q, Ma L and Kang J: Long non‑coding RNAs in glioma 23. Cong Z, Diao Y, Xu Y, Li X, Jiang Z, Shao C, Ji S, Shen Y, De W
progression. Cancer Lett 419: 203‑209, 2018. and Qiang Y: Long non‑coding RNA linc00665 promotes lung
3. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA and adenocarcinoma progression and functions as ceRNA to regulate
Jemal A: Global cancer statistics 2018: GLOBOCAN estimates AKR1B10‑ERK signaling by sponging miR‑98. Cell Death
of incidence and mortality worldwide for 36 cancers in Dis 10: 84, 2019.
185 countries. CA Cancer J Clin 68: 394‑424, 2018. 24. Shan Y and Li P: Long intergenic non‑protein coding RNA
4. Ferlay J, Colombet M, Soerjomataram I, Mathers C, Parkin DM, 665 regulates viability, apoptosis, and autophagy via the
Piñeros M, Znaor A and Bray F: Estimating the global cancer MiR‑186‑5p/MAP4K3 axis in hepatocellular carcinoma. Yonsei
incidence and mortality in 2018: GLOBOCAN sources and Med J 60: 842‑853, 2019.
methods. Int J Cancer 144: 1941‑1953, 2019. 25. Livak KJ and Schmittgen TD: Analysis of relative gene expres-
5. Saxena S and Jha S: Role of NOD‑like receptors in glioma sion data using real‑time quantitative PCR and the 2(‑Delta Delta
angiogenesis: Insights into future therapeutic interventions. C(T)) method. Methods 25: 402‑408, 2001.
Cytokine Growth Factor Rev 34: 15‑26, 2017. 26. Cui CL, Li YN, Cui XY and Wu X: lncRNA XIST promotes the
6. Rynkeviciene R, Simiene J, Strainiene E, Stankevicius V, progression of laryngeal squamous cell carcinoma by sponging
Usinskiene J, Miseikyte Kaubriene E, Meskinyte I, Cicenas J and miR‑144 to regulate IRS1 expression. Oncol Rep 43: 525‑535,
Suziedelis K: Non‑coding RNAs in glioma. Cancers (Basel) 11: 2020.
17, 2018. 27. Bao W, Cao F, Ni S, Yang J, Li H, Su Z and Zhao B: lncRNA
7. Wang Q, Li Q, Zhou P, Deng D, Xue L, Shao N, Peng Y and Zhi F: FLVCR1‑AS1 regulates cell proliferation, migration and invasion
Upregulation of the long non‑coding RNA SNHG1 predicts poor by sponging miR‑485‑5p in human cholangiocarcinoma. Oncol
prognosis, promotes cell proliferation and invasion, and reduces Lett 18: 2240‑2247, 2019.
apoptosis in glioma. Biomed Pharmacother 91: 906‑911, 2017. 28. Wang Y, Zeng X, Wang N, Zhao W, Zhang X, Teng S, Zhang Y
8. Gao Y, Yu H, Liu Y, Liu X, Zheng J, Ma J, Gong W, Chen J, and Lu Z: Long noncoding RNA DANCR, working as a competi-
Zhao L, Tian Y and Xue Y: Long non‑coding RNA HOXA‑AS2 tive endogenous RNA, promotes ROCK1‑mediated proliferation
regulates malignant glioma behaviors and vasculogenic mimicry and metastasis via decoying of miR‑335‑5p and miR‑1972 in
formation via the MiR‑373/EGFR axis. Cell Physiol Biochem 45: osteosarcoma. Mol Cancer 17: 89, 2018.
131‑147, 2018. 29. Miao H, Wang L, Zhan H, Dai J, Chang Y, Wu F, Liu T, Liu Z,
9. Lorenzen JM and Thum T: Long noncoding RNAs in kidney and Gao C, Li L and Song X: A long noncoding RNA distributed in
cardiovascular diseases. Nat Rev Nephrol 12: 360‑373, 2016. both nucleus and cytoplasm operates in the PYCARD‑regulated
10. Sun W, Yang Y, Xu C and Guo J: Regulatory mechanisms of apoptosis by coordinating the epigenetic and translational regu-
long noncoding RNAs on gene expression in cancers. Cancer lation. PLoS Genet 15: e1008144, 2019.
Genet 216‑217: 105‑110, 2017. 30. Xu H, Zhao G, Zhang Y, Jiang H, Wang W, Zhao D, Yu H
11. Dastmalchi N, Safaralizadeh R and Nargesi MM: LncRNAs: and Qi L: Long non‑coding RNA PAXIP1‑AS1 facilitates cell
Potential novel prognostic and diagnostic biomarkers in invasion and angiogenesis of glioma by recruiting transcription
colorectal cancer. Curr Med Chem 27: 5067‑5077, 2020. factor ETS1 to upregulate KIF14 expression. J Exp Clin Cancer
12. Lu Q, Gong W, Wang J, Ji K, Sun X, Xu C, Du L, Wang Y and Res 38: 486, 2019.
Liu Q: Analysis of changes to lncRNAs and their target mRNAs 31. Liu Q, Yu W, Zhu S, Cheng K, Xu H, Lv Y, Long X, Ma L,
in murine jejunum after radiation treatment. J Cell Mol Med 22: Huang J, Sun S and Wang K: Long noncoding RNA GAS5 regu-
6357‑6367, 2018. lates the proliferation, migration, and invasion of glioma cells by
13. Sallam T, Jones M, Thomas BJ, Wu X, Gilliland T, Qian K, negatively regulating miR‑18a‑5p. J Cell Physiol 234: 757‑768,
Eskin A, Casero D, Zhang Z, Sandhu J, et al: Transcriptional 2018.
regulation of macrophage cholesterol efflux and atherogenesis by 32. Li J, Guo W, Xue W, Xu P, Deng Z, Zhang D, Zheng S and Qiu X:
a long noncoding RNA. Nat Med 24: 304‑312, 2018. Long noncoding RNA AURKAPS1 potentiates malignant
14. Li R and Fox AH: SPArking interest in the long noncoding RNA hepatocellular carcinoma progression by regulating miR‑142,
world: A new class of 5'SnoRNA‑stabilized LncRNA that influ- miR‑155 and miR‑182. Sci Rep 9: 19645, 2019.
ences alternative splicing. Mol Cell 64: 435‑437, 2016. 33. Wang Y, Jiang F, Xiong Y, Cheng X, Qiu Z and Song R: LncRNA
15. Xu ZM, Huang F and Huang WQ: Angiogenic lncRNAs: A TTN‑AS1 sponges miR‑376a‑3p to promote colorectal cancer
potential therapeutic target for ischaemic heart disease. Life progression via upregulating KLF15. Life Sci 244: 116936, 2020.
Sci 211: 157‑171, 2018. 34. Gao J, Li N, Dong Y, Li S, Xu L, Li X, Li Y, Li Z, Ng SS,
16. Tripathi MK, Doxtater K, Keramatnia F, Zacheaus C, Sung JJ, et al: miR‑34a‑5p suppresses colorectal cancer
Yallapu MM, Jaggi M and Chauhan SC: Role of lncRNAs metastasis and predicts recurrence in patients with stage II/III
in ovarian cancer: Defining new biomarkers for therapeutic colorectal cancer. Oncogene 34: 4142‑4152, 2015.
purposes. Drug Discov Today 23: 1635‑1643, 2018. 35. Ma S, Fu T, Zhao S and Gao M: MicroRNA‑34a‑5p suppresses
17. Chandra Gupta S and Nandan Tripathi Y: Potential of long tumorigenesis and progression of glioma and potentiates
non‑coding RNAs in cancer patients: From biomarkers to thera- Temozolomide‑induced cytotoxicity for glioma cells by targeting
peutic targets. Int J Cancer 140: 1955‑1967, 2017. HMGA2. Eur J Pharmacol 852: 42‑50, 2019.
18. Tamang S, Acharya V, Roy D, Sharma R, Aryaa A, Sharma U, 36. Xu H, Zhang Y, Qi L, Ding L, Jiang H and Yu H: NFIX circular
Khandelwal A, Prakash H, Vasquez KM and Jain A: SNHG12: RNA promotes glioma progression by regulating miR‑34a‑5p via
An LncRNA as a potential therapeutic target and biomarker for notch signaling pathway. Front Mol Neurosci 11: 225, 2018.
human cancer. Front Oncol 9: 901, 2019. 37. Di Bari M, Bevilacqua V, De Jaco A, Laneve P, Piovesana R,
19. Chen J, Wang R, Zhang K and Chen LB: Long non‑coding RNAs Trobiani L, Talora C, Caffarelli E and Tata AM: Mir‑34a‑5p
in non‑small cell lung cancer as biomarkers and therapeutic mediates cross‑talk between M2 muscarinic receptors and
targets. J Cell Mol Med 18: 2425‑2436, 2014. notch‑1/EGFR pathways in U87MG glioblastoma cells:
20. Yang B, Bai Q, Chen H, Su K and Gao C: LINC00665 induces Implication in cell proliferation. Int J Mol Sci 19: 1631, 2018.
38. Ma Y, Xia Z, Ye C, Lu C, Zhou S, Pan J, Liu C, Zhang J, Liu T,
gastric cancer progression through activating Wnt signaling Hu T, et al: AGTR1 promotes lymph node metastasis in breast
pathway. J Cell Biochem 121: 2268‑2276, 2020. cancer by upregulating CXCR4/SDF‑1α and inducing cell
21. Qi H, Xiao Z and Wang Y: Long non‑coding RNA LINC00665 migration and invasion. Aging (Albany NY) 11: 3969‑3992, 2019.
gastric cancer tumorigenesis by regulation miR‑149‑3p/RNF2
axis. Onco Targets Ther 12: 6981‑6990, 2019.
22. Liu X, Lu X, Zhen F, Jin S, Yu T, Zhu Q, Wang W, Xu K, Yao J This work is licensed under a Creative Commons
and Guo R: LINC00665 induces acquired resistance to gefitinib Attribution-NonCommercial-NoDerivatives 4.0
through recruiting EZH2 and activating PI3K/AKT pathway in International (CC BY-NC-ND 4.0) License.
NSCLC. Mol Ther Nucleic Acids 16: 155‑161, 2019.You can also read

















































